1. Introduction
The juxtaposition of terrestrial and marine environments along the ocean's shores and the abundance and diversity of modern life observed there reflect the ecological and evolutionary significance of this environmental setting. Representatives of all major animal phyla spend some portion of their lives in intertidal settings and, despite the relatively small global surface area represented by these environments, littoral sequences are well represented in the geologic record. However, soft-bodied macrofossils recording life in ancient littoral settings are almost entirely absent from the fossil record. This dichotomy between living abundance and fossil rarity emphasizes the preservational biases associated with the erosional and biodegradational processes characteristic of the vast majority of coastlines. Background wave and tide energy, punctuated by periodic storm events, results in repeated high-energy deposition and reworking of poorly consolidated substrates. Soft tissues transported and/or exhumed by these events are, with rare exception, quickly broken apart by wave energy and/or are consumed by marine and terrestrial scavengers, and thus an unusually small proportion of organic matter is buried in littoral sediments. Additionally, the elevated porosity and permeability associated with most coastal substrates results in heightened flow of oxygenated pore fluids, enhancing the rapid decomposition of any remaining organic detritus. Due to the rarity with which fossilization of nonmineralized taxa occurs in these settings, those few nonmineralized beach inhabitants which do eventually make it to the fossil record provide exceptional windows into the palaeobiology and taphonomy of intertidal biotas. Conspicuously, fossils of Cambrian medusozoans, or ‘jellyfish’, are a substantial portion of this otherwise sparse record, suggesting that unusual preservational pathways, resulting from the interaction of medusozoan tissues with a unique set of environmental and ecological variables, were operating along early Palaeozoic shorelines.
Medusozoans are considered among the oldest and most widespread macrofaunal zooplankton groups in modern oceans (Collins, Reference Collins2002; Peterson & Butterfield, Reference Peterson and Butterfield2005; Collins et al. Reference Collins, Schuchert, Marques, Jankowski, Medina and Schierwater2006) and yet, despite the abundance, ubiquity and deep evolutionary history of medusozoans, the fossil record of this subphylum is exceedingly sparse. To supplement and foster further evaluation of the nature of the fossil record of Cambrian jellyfish, we describe a new medusa-bearing locality from the lower Cambrian Zabriskie Quartzite of the southern Nopah Range in the Death Valley region (southeastern California, USA) (Fig. 1). Discoidal structures occurring on a single bedding plane at this locality were examined; palaeobiologically, sedimentologically and taphonomically characterized; and compared to both modern and other fossilized examples of medusozoan remains. This analysis suggests that the Zabriskie fossils provide the oldest macrofossil evidence of cnidarian medusae from the Phanerozoic. Further, the sedimentological context and en masse occurrence of these fossils indicate that this assemblage records the oldest reported example of a metazoan mass stranding event. Further, the taphonomic character of the Zabriskie specimens suggests that previous preservational models for Cambrian medusozoans may be refined by further integrating conceptual models for Cambrian Earth system dynamics with uniformitarian sedimentological principles. Comparison to broadly similar preservational modes characteristic of Ediacaran Lagerstätten also highlights important distinctions between Ediacaran and Cambrian taphonomic pathways, with important implications for the preservational, as well as evolutionary, dynamics of the Precambrian–Cambrian transition (see Butterfield, Reference Butterfield2003; Buatois et al. Reference Buatois, Narbonne, Mángano, Carmona and Myrow2014).

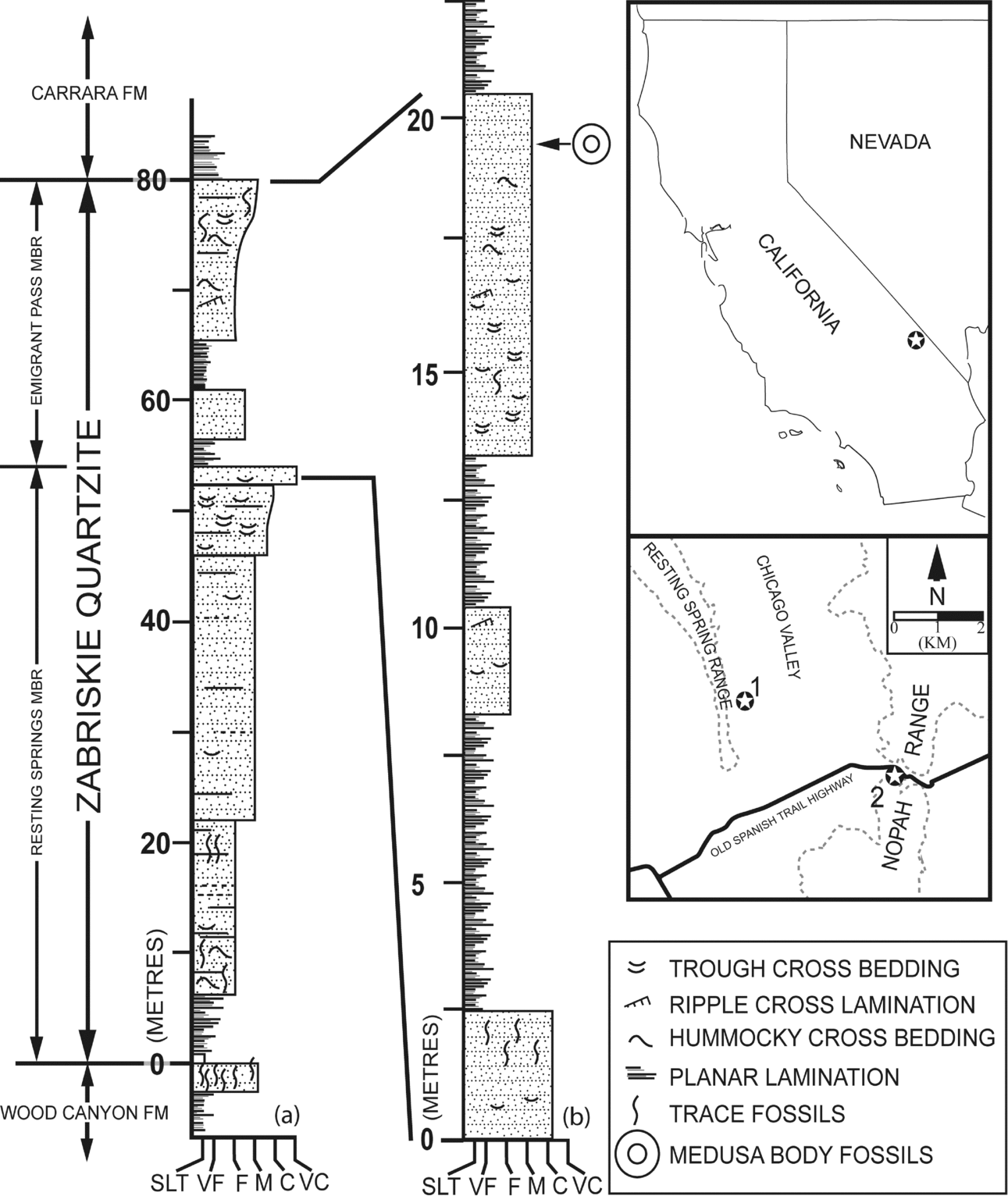
Figure 1. Location map and stratigraphic sections of the Zabriskie Quartzite. (a) Type section for Zabriskie Quartzite in Resting Springs Range (adapted from Prave, Reference Prave1992). (b) Nopah Range section indicating approximate stratigraphic position of medusa body fossils.
2. Geologic setting
Precambrian–Cambrian sediments are well exposed in western North America, largely as a result of protracted subsidence of the continental margin following separation of Laurentia from the supercontinent Rodinia (Stewart, Reference Stewart1970, Reference Stewart1972). Rifting was initiated and the ensuing tectonic subsidence was hinged along a sinuous craton margin that essentially traced what is now the southern and eastern perimeter of the Great Basin, continuing north-northwestward into Canada (Stewart, Reference Stewart1972; Fedo & Cooper, Reference Fedo and Cooper2001). Nearly coincident with this tectonic activity, sea level rise associated with the onset of the Sauk Marine Transgression resulted in further expansion of available sediment accommodation space (Sloss, Reference Sloss1963; Stewart, Reference Stewart1970, Reference Stewart1972). A sedimentary succession nearly 9 km thick accumulated in portions of the basin, resulting in an extensive and largely continuous record of the Precambrian–Cambrian transition, today well exposed throughout the western Cordillera (Stewart, Reference Stewart1970, Reference Stewart1972).
The establishment of passive margin sedimentation in the early Phanerozoic coupled with Sauk-associated rising sea levels likely resulted in, beginning in the early Cambrian, an eastward march of the palaeoshoreline. One interval of the shoreline's migration is recorded in the widespread and laterally continuous sandstones and siltstones assigned to the upper portions of the Zabriskie Quartzite (Stewart, Reference Stewart1970; Prave, Reference Prave1992). The Zabriskie Quartzite is a texturally mature siliciclastic unit exposed over 36000 km2 in SE California and portions of S Nevada (Barnes & Klein, Reference Barnes, Klein and Ginsburg1975). The formation ranges from 3 to c. 300 m in thickness, thickening from SE to NW, and is generally composed of massive to cross-bedded, fine- to medium-grained quartz arenite interbedded with mudstone and siltstone (Stewart, Reference Stewart1970; Prave, Reference Prave1992). The underlying Wood Canyon Formation and the overlying Carrara Formation have each yielded Bonnia-Olenellus faunas (Stewart, Reference Stewart1970; Diehl, Reference Diehl1979), thus constraining the age of the Zabriskie Quartzite to the early Cambrian. Exquisite examples of Skolithos piperock, the distinctively Cambrian trace fossil for which the Zabriskie Quartzite is best known, are readily apparent in the majority of exposures of the formation. Localized assemblages of Arenicolites and Planolites burrows are also present. Prior to this investigation, body fossils had not been reported from the Zabriskie Quartzite.
3. Palaeoenvironmental setting
Detailed facies analyses and sequence stratigraphic evaluation of the Zabriskie Quartzite suggest that deposition occurred in a range of coastal marine and terrestrial environments (Prave, 1984, 1992). Prave (1984) described 18 lithofacies preserved within multiple outcrops of the formation. These lithofacies are composed entirely of siliciclastic packages of varying textural and mineralogical maturity. Observation of the lateral and vertical distribution of lithofacies cropping out in California and Nevada suggests that the thickest exposures of the Zabriskie Quartzite consist of two distinct sequence tracts separated by a regional unconformity in the upper portion of the unit (Prave, Reference Prave1992). Facies associations in the lower portion of the Zabriskie Quartzite are indicative of a transition from marine to terrestrial deposition during a sea level lowstand, during which densely burrowed near-shore marine and coastal lithofacies gradually gave way to prograding braidplain deposits (Prave, Reference Prave1992). This succession is capped by a regionally persistent transgressive lag which marks the onset of sea level rise and associated coastal onlapping of marine facies (Prave, Reference Prave1992). Consistent with the division of these systems tracts, the Zabriskie Quartzite is divided into the lower Resting Spring Member and the upper Emigrant Pass Member (Fig. 1). The fossils reported herein were discovered in situ within the Emigrant Pass Member, c. 20 m above the base of the transgressive lag marking the base of this subunit (Fig. 1).
Beaches are generally high-energy environments characterized by sequential episodes of rapid deposition and erosion. This alternation, paired with the abundance and activity of infaunal and epifaunal communities, results, in modern littoral systems, in a highly dynamic and typically unstable substrate. Lacking a terrestrial biotic component, the early Cambrian coastline was likely to have been fundamentally distinct from the majority of modern beaches. Microbial and mat-forming communities were likely far more widespread in near-shore marine environments at this time than they are today (Hagadorn & Bottjer, Reference Hagadorn and Bottjer1997; Gehling, Reference Gehling1999; Noffke, Knoll & Grotzinger, Reference Noffke, Knoll and Grotzinger2002; Gehling & Droser, Reference Gehling and Droser2009). The proliferation of microbially bound substrates, in conjunction with the deposition of sheet quartzites, is recorded by the globally widespread occurrence of microbially induced sedimentary structures (MISS) or textured organic surfaces (TOS) in Precambrian through lowermost Palaeozoic strata (Hagadorn & Bottjer, Reference Hagadorn and Bottjer1997; Gehling, Reference Gehling1999; Noffke, Knoll & Grotzinger, Reference Noffke, Knoll and Grotzinger2002; Gehling & Droser, Reference Gehling and Droser2009). Animals expanded into the infaunal life mode during the early Phanerozoic and, even by the late early Cambrian, infaunalization was well advanced (Tarhan & Droser, Reference Tarhan and Droser2014), as evidenced by the abundance and size of Skolithos and Arenicolites burrows in marine facies of the Zabriskie Quartzite. However, concurrently, littoral sediments subject to exposure and desiccation were less well colonized by infauna, permitting denser communities of microbes to thrive relatively undisturbed. MISS or TOS are thus not uncommon in many early Cambrian littoral quartzite successions in the Great Basin. In the Zabriskie Quartzite these structures include wrinkle marks, pustules and ‘elephant skin’ textures.
4. Materials and methods
The Zabriskie medusozoan specimens reported herein occur along a single, east-dipping sandstone bedding plane, within a c. 9 m thick planar-laminated to ripple cross-laminated sandstone interval exposed in the upper portion (Emigrant Pass Member) of the Zabriskie Quartzite in the southern Nopah Range (Fig. 2). The specimen-bearing bed was examined and excavated in situ, exposing c. 6 m2 of fossiliferous bedding plane (Fig. 2). The fully excavated bed was divided into 25 cm × 25 cm grids, and the location, size, orientation and morphological features of all discoidal fossils occurring on the bed were recorded (Fig. 2). Similar structures observed in adjacent (within a 5 m radius) float were also catalogued; these specimens bear a marked lithologic similarity to the in situ fossiliferous bedding plane, strongly suggesting that these loose pieces were weathered from the immediately adjacent outcrop. Measurements taken in the field were supplemented using ImageJ image-processing and analysis software (Rasband, 1997–2008) on digital photographs. Latex moulds were also made in order to facilitate laboratory analysis, including the production of replicate casts.

Figure 2. Excavated fossiliferous bedding plane in the Zabriskie Quartzite. (a) Panoramic photo of excavated bedding plane following 25 cm × 25 cm grid application. (b) Line drawing of fossils mapped on bedding plane. Specimen labelling in (b) coincides with specimen labelling in Figure 3. Scale bars are 25 cm.
A total of 13 specimens were identified from the excavated surface and associated talus. Of these, nine were recorded along the in situ bedding plane and four were identified in immediately adjacent float. Specimens range from 3 cm to greater than 21 cm in diameter (Fig. 3), with fossil margins of ≤ 2–3 mm thickness preserved in both convex and concave epirelief. Specimens were systematically measured along the shortest axis; measurements thus represent a minimum size. All of the specimens are slightly ovoid, with the elongated axis roughly parallel to the predominant orientation of ripple crests preserved along the same bedding plane. Similarly, specimens are also characterized by variable topographic relief; commonly, the greatest variation in relief occurs in the largest specimens, along an axis parallel to that of the surrounding ripple crests and troughs (Fig. 2).

Figure 3. Lower Cambrian medusa fossils from the Zabriskie Quartzite. (a) Specimen preserving evidence of sediment infill (white arrow) (UCR11131-1). (b) Close-up of fossil shown in (a) showing evidence of folding adjacent to the central mound (‘A’) and along the fossil's marginal furrow (‘B’). ‘C’ indicates bulbous structure projecting from the fossil's central mound (UCR11131-1). (c) Large fossil containing ripples (white arrows indicate ripple crests) in the interior of the specimen, suggesting collapse of a portion of the carcass interior prior to burial (UCR11131-2). (d) Discoidal specimen with two primary concentric ridges (‘A’) and evidence of folding in the central mound of the fossil (‘B’) (UCR11131-3). (e, f) Discoidal specimens each characterized by two primary concentric ridges (A) and evidence of folding in the central mound of the fossil (B) (UCR11131-4 and UCR11131-5). (g) Specimen characterized by evidence of preburial deformation (‘rilling’). Corresponding image A–A′–A″’ shows cross-sectional view through this specimen.Undisturbed laminae immediately underlying the specimen are indicated by the white arrows (UCR11131-6). (h) Discoidal specimen with a single marginal ridge (white arrow). Corresponding image B–B′ shows cross-sectional view through the specimen. Undisturbed laminae immediately underlying the specimen are indicated by the white arrows (UCR11131-7). (i) White arrows indicate, for contrast, abiotic sedimentary structures (‘sand volcanoes’) from an outcrop of the Zabriskie Quartzite in the Montgomery Mountains (western Nevada). Corresponding image C–C′ shows cross-sectional view through one of these structures. Note evidence of fluidization and upward migration of underlying sediments (UCR11132-1). Scale bars equal 5 cm in (a–f) and 1 cm in (g–i).
Select fossils were removed from the exposure and transported back from the field for sectioning. This material was cut using an automated oil saw and subsequently polished on a vibrating lap in multiple stages, using a series of successively finer silicon carbide grits. Polished surfaces were scanned while submerged in a water reservoir to enable high-resolution photographic analyses of sedimentological features preserved in the fossils and surrounding horizons. Thin-sections were generated from these materials for further petrologic evaluation. Cross-sections cut and polished orthogonal to bedding indicate that the laminae underlying the specimens are entirely undisrupted (Fig. 3g, h).
Specimens typically occur as high-relief and topographically variable sandstone casts and moulds. Positive-relief (convex) portions of each of these fossils are distinctly lighter-coloured, allowing these fossils to be readily distinguished from the dark desert varnish that coats the remainder of the bed. This disparity in colour does not result from differences in either composition or grain-size, but instead likely reflects differential weathering of cements due to the elevated topography of these fossils.
In addition to the discoidal to elliptical shape, associated with laminated sediments and high relief described above, the Zabriskie specimens are characterized by a number of additional distinctive features. Three general preservational motifs characterize the Zabriskie specimens: (1) convex discoidal ridges, (2) concave discoidal ‘moats’ encircling a flat or low positive-relief interior and (3) convex mounds characterized by second-order elements of highly variable relief, indicative of plastic deformation. Specimens are commonly characterized by one or more of these motifs, and may occupy a continuum between them.
Discs preserved as convex discoidal ridges include both the largest and smallest of the Zabriskie specimens. These specimens are characterized by a general discoidal shape, without additional embellishment. Ripples preserved in the interior of the largest specimen (Fig. 3c) are consistent in orientation, size and general morphology with those present on the surrounding bedding plane.
Several specimens are characterized by concentric convex rings separated by a concave depression or ‘moat’ (Fig. 3d, e, f). The central regions of these fossils are generally characterized by low positive relief, with some negative-relief grooves or folds in their central regions (Fig. 3d, e, f). This preservational style was characteristic of all specimens ranging from c. 10 to 15 cm in diameter. Faint concentric rings were observed within the concave moats of at least two specimens (Fig. 3d, e).
Several specimens are distinctively preserved in convex relief. One particularly well-preserved example (Fig. 3a, b, d), collected from the in situ bedding plane, contains abundant striations of considerable relief. The striations associated with the specimen margins are characterized by similar relief to that of the striations associated with the specimen interior (Figs 3b, 4b).

Figure 4. Photograph (a) and corresponding line drawing (b) of major features observed in Zabriskie (UCR11131-1). See also Figure 3a, b. Scale bars equal 5 cm.
5. Interpretation
Discoidal structures generally comparable to the Zabriskie specimens are common elements of the rock record. This commonality reflects the frequency with which ring-like impressions are produced by both biotic and abiotic processes. Hypotheses surrounding discoidal rock impressions have repeatedly been reconsidered and revised as new materials are uncovered or as new perspectives take hold (Pickerill, Reference Pickerill1982, Reference Pickerill1990; Jensen et al. Reference Jensen, Gehling, Droser and Grant2002; MacGabhann, Reference MacGabhann, Vickers-Rich and Komarower2007; Young & Hagadorn, Reference Young and Hagadorn2010; Hagadorn & Miller, Reference Hagadorn and Miller2011). Interpretation of the origin of the Zabriskie specimens thus requires consideration of the wide spectrum of alternative hypotheses, including both abiogenic and alternative biogenic structures. Interpretations of affinity for the Zabriskie materials must be consistent with at least five distinctive features characteristic of individual specimens and shared among the assemblage. These are as follows: (1) sediments immediately underlying these fossils remain undisturbed; (2) specimens are preserved in both concave and convex epirelief; (3) ‘wrinkling’ along the margins of the fossils; (4) individual specimens are deformed parallel to the palaeocurrent; and (5) the specimens are, as an assemblage, non-uniform in size, distribution and preservational style.
Circular features observed on bedding planes are most commonly interpreted as scratch circles (Jensen et al. Reference Jensen, Gehling, Droser and Grant2002), trace fossils (e.g. Mata et al. Reference Mata, Corsetti, Corsetti, Awramik and Bottjer2012), gas/fluid escape features (Hagadorn & Miller, Reference Hagadorn and Miller2011) and concretions (Seilacher, Reference Seilacher2001), as well as being attributed to a variety of radially symmetrical body fossils (e.g. Gehling, Narbonne & Anderson, Reference Gehling, Narbonne and Anderson2000). The characteristics of each of these potential affinities are described and the Zabriskie specimens evaluated according to these criteria below.
Scratch circles (Kullingia) are discoidal features produced by currents which induce a passive interaction between ‘tethered’ (e.g. stalked and attached to the substrate by means of a holdfast) epibenthic organisms and the substrate (Jensen et al. Reference Jensen, Gehling, Droser and Grant2002). Most commonly, these structures are produced when currents spin tubular organisms attached to the seafloor at the sediment–water interface, producing a series of low-, negative-relief concentric rings in the substrate reflecting the presence of repeated annulations or ornamentation along the stalk of the scratch circle-producing organism (Jensen et al. Reference Jensen, Gehling, Droser and Grant2002). A conical depression, representing the organism's attachment site, is sometimes present at the centre of scratch circles, and very rarely some portion of the tubular organism may also be preserved (Jensen et al. Reference Jensen, Gehling, Droser and Grant2002). The temporal distribution of scratch circles in normal marine strata is non-uniform and there is an unusual abundance of scratch circles in Neoproterozoic and lower Palaeozoic shallow marine strata, potentially reflecting the presence of widespread firm and cohesive substrate conditions resulting from the lack of intensive bioturbation through this interval (Droser, Jensen & Gehling, Reference Droser, Jensen and Gehling2002a , Reference Droser, Jensen, Myrow and Narbonne b ; Jensen et al. Reference Jensen, Gehling, Droser and Grant2002; Tarhan et al. Reference Tarhan, Droser, Planavsky and Johnston2015b ).
In spite of a similar general morphology, the detailed features described above for the Zabriskie specimens are in stark contrast to those characteristic of scratch circles. The Zabriskie specimens are largely elliptical, rather than perfectly circular. This asymmetry is not uniformly distributed along the bed in a single compass direction (Fig. 2) nor is evidence of tectonic deformation observed in immediately adjacent beds. Moreover, trace fossils (e.g. Skolithos and Arenicolites), body fossils (e.g. trilobites, Harlaniella confusa, hyolithids, etc.) and abiotic sedimentary structures observed in strata above and below the fossil-bearing bed in the same section are not characterized by evidence of tectonic deformation on the scale required to produce the alignment observed in the Zabriskie fossils. Additionally, the variable internal morphology (e.g. folds and rilling) characteristic of the Zabriskie fossils is entirely dissimilar from the smooth surface and regular concentric rings of Kullingia. The positive relief of the ridges and raised central mounds observed in the epirelief Zabriskie fossils (Fig. 3) also suggests that a scratch circle origin is improbable, as the rotation of a tethered stalk would be expected to produce, as preserved in epirelief, a very low-relief structure with concentric furrows, rather than ridges.
Animal burrows (e.g. Berguaeria), gas/fluid escape structures (e.g. Astropolithon), and concretions are also commonly preserved as discoidal structures along bedding planes. Such structures can be quite large, can contain considerable ornamentation and commonly occur en masse and have thus been repeatedly mistaken for fossilized medusae (Pickerill & Harris, Reference Pickerill and Harris1979; Seilacher, Reference Seilacher2001; Hagadorn & Miller, Reference Hagadorn and Miller2011; Mata et al. Reference Mata, Corsetti, Corsetti, Awramik and Bottjer2012). Distinguishing true medusozoan body fossils from either discoidal trace fossils or discoidal abiotic sedimentary structures is discussed collectively below, as both of these processes involve perturbation of sediment at depth. Thus, distinguishing structures produced by these processes from body fossils is largely contingent upon three-dimensional data obtained from viewing materials in both bedding-plane and cross-section.
Slabbed specimens reveal that laminae immediately below the Zabriskie fossils are undisrupted (Fig. 3g, h), indicating that these structures did not form through either passive (abiotic) or active (infaunal) sediment disturbance. Sand volcanoes collected from an outcrop of the Zabriskie Quartzite in the Montgomery Mountains (Fig. 3i) contain ample evidence of sediment disruption, including fluidization and upward migration of sediments. Viewed in cross-section (Fig. 3i), laminae are continuously interrupted for centimetres below the bedding plane surface expression of each sand volcano. Sectioned Zabriskie medusa samples, however, contain undisrupted laminae within millimetres of each fossil specimen (Fig. 3g, h). In bedding plane view, the folds and asymmetry of the Zabriskie fossils provide further evidence that a trace fossil or non-biogenic sedimentary origin is improbable.
Discoidal holdfasts, the attachment structures of frondose and tubular organisms, are a common component of the terminal Ediacaran-aged Ediacara Biota; structures such as Aspidella are broadly morphologically comparable to the Zabriskie fossils, and likewise typically occur en masse (Gehling, Narbonne & Anderson, Reference Gehling, Narbonne and Anderson2000). Precambrian holdfasts may be preserved as positive-relief casts on either the tops or the bases of beds, are characterized by regular relief and pronounced external margins and may also contain concentric rings, which have been observed to incrementally gain relief toward the centre of the fossil (Gehling, Narbonne & Anderson, Reference Gehling, Narbonne and Anderson2000). However, the styles of preservation and general morphology of Ediacaran holdfasts make a comparable origin for the Zabriskie fossils unlikely. Evidence of an attached stalk, as might be expected from an epirelief composite holdfast (Tarhan et al. Reference Tarhan, Droser, Gehling and Dzaugis2015a ), is absent from the Zabriskie specimens. Further, the margins of the Zabriskie fossils are irregular and show signs of significant biostratinomic deformation, inconsistent with the preservation of Ediacaran holdfasts which, as previously noted, are typically characterized by regular margins with pronounced relief (Gehling, Narbonne & Anderson, Reference Gehling, Narbonne and Anderson2000). The absence of disrupted laminae immediately beneath these fossils, as well as the absence of unidirectional strain marks – in spite of the evidence of alignment characterizing the Zabriskie fossil assemblage (Fig. 2) – is inconsistent with a pullout origin for these fossils (cf. Tarhan, Droser & Gehling, Reference Tarhan, Droser and Gehling2010), as the forcible removal of a stalked organism from the substrate would have disturbed the immediately underlying sediment.
The features observed within individual Zabriskie specimens and their occurrence as moderately dense, spatially associated assemblages of potentially monospecific and current-deformed discoidal fossils (without chitinous or biomineralized components) along a single bedding plane are most consistent with a cnidarian mass stranding interpretation (though some modern ctenophores have also been observed to form dense aggregates, albeit in deeper marine (i.e. non-intertidal) settings (Reisser et al. Reference Reisser, Proietti, Sazima, Kinas, Horta and Secchi2013)). Medusozoans are, among modern groups, considered to be among the oldest and most widespread macrofaunal zooplankton groups (Collins, Reference Collins2002; Peterson & Butterfield, Reference Peterson and Butterfield2005; Erwin et al. Reference Erwin, Laflamme, Tweedt, Sperling, Pisani and Peterson2011). Phylogenetic and molecular clock data are consistent with a Precambrian origin for the Medusozoa (Collins, Reference Collins2002; Peterson & Butterfield, Reference Peterson and Butterfield2005; Peterson et al. Reference Peterson, Cotton, Gehling and Pisani2008), and previous reports of definitive medusa body fossils, likely representing extant crown-group lineages, extend as far back as the middle to upper Cambrian (Furongian) (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Cartwright et al. Reference Cartwright, Halgedahl, Hendricks, Jarrard, Marques, Collins and Lieberman2007; Hagadorn & Belt, Reference Hagadorn and Belt2008; Young & Hagadorn, Reference Young and Hagadorn2010; Erwin et al. Reference Erwin, Laflamme, Tweedt, Sperling, Pisani and Peterson2011). Incontrovertible fossils of mass strandings of Cambrian jellyfish have also been previously reported from the Potsdam Group of New York (Hagadorn & Belt, Reference Hagadorn and Belt2008) and the Elk Mound Group of Wisconsin (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Tarhan, unpub. MA thesis, Amherst College, Amherst, MA, 2008).
Dense aggregations or ‘blooms’ of medusae are common in modern coastal environments and can be associated with either reproductive or feeding behaviours (Hamner, Hamner & Strand, Reference Hamner, Hamner and Strand1994; Purcell, Reference Purcell2003, Reference Purcell2005; Magome et al. Reference Magome, Yamashita, Kohama, Kaneda, Hayami, Takahashi and Takeoka2007; Albert, Reference Albert2011). Inhabitation of wave-influenced, near-shore marine environments by modern medusae commonly results in mass strandings in the intertidal zone (Fig. 4). Behavioural observations of the scyphozoan medusa Aurelia suggest that migration to tidally influenced portions of the shoreline may even be active as well as passive (Albert, Reference Albert2011). Medusae will position themselves in the upper portion of the water column during incoming tides in order to be washed into the intertidal zone to forage, typically riding the ebb tide back out to sea (Albert, Reference Albert2011). Medusae damaged by wave activity during the shoreward venture or trapped in coastal depressions following the ebbing of high tide or storm waters become stranded in the intertidal zone, commonly resulting in portions of these aggregations washing ashore en masse (Fig. 4). Unlike mineralized fossils such as trilobites which bear features that, even following biostratinomic degradation, can commonly be readily identified, even to the species level, the simple, radially symmetrical body plan characteristic of medusozoans is convergent with that of a number of disparate taxa. This, coupled with a lack of mineralized sclerites, can make even class- or phylum-level identification of discoidal fossils, let alone categorization to a more specific level, a challenging endeavour. Here, the presence of multiple fossils along a single bedding plane becomes a vital tool in interpreting these structures. The range and variability of detail provided by concurrent preservation of immediately spatially associated individuals under largely similar environmental conditions provides a useful metric for distinguishing anatomical from taphonomic structures, as well as shedding light on the behavioural implications of medusozoan mass accumulations. The strong probability that this fossiliferous bedding plane is part of an intertidal facies lends further credence to interpretation of the Zabriskie fossil assemblage as a medusozoan mass stranding.
The anatomy of modern cnidarian medusae generally consists of a radially symmetrical, umbrella-shaped body or ‘bell’, reproductive organs, a ventrally located mouth attached to the tip of a stalk-like manubrium which is, in turn, attached to, in certain groups, oral arms, as well as, among certain groups, tentacles trailing from the bell margin. Medusae are most hydrodynamically stable with their subumbrella (ventral side) down and thus most commonly come to rest with the subumbrella in direct contact with the substrate (Schäfer & Craig, Reference Schäfer and Craig1972; Bruton, Reference Bruton, Simonetta and Morris1991). Other areas of a medusa carcass, conversely, will commonly be disrupted by wave activity or heterogeneous rates of desiccation.
Flat-topped or convex central mounds, consistent with those observed in modern medusae and with previous reports of fossilized medusae (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Hagadorn & Belt, Reference Hagadorn and Belt2008) are present in many of the Zabriskie specimens (Figs 2, 3a, c, d, e, h, 4). The best-preserved example among the Zabriskie fossils (Figs 3a, b, 4) contains a pronounced central convex mound, encircled by ridges of variable relief strongly indicative of tissue deterioration and deformation (Figs 3b, 4). The lack of disturbance of sediments directly underlying these structures suggests that these structures were produced at the sediment–water interface. Further, as previously mentioned, several of the Zabriskie specimens are characterized by a lack of perfect radial symmetry; for instance, the raised sediment mound in the best preserved of the Zabriskie fossils (Figs 3a, b, 4) is not centralized, which is atypical for non-medusozoan discoidal structures.
The variety of preservational motifs characteristic of the Zabriskie fossils, even along a single bedding plane, is a common theme among medusa body fossils (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Hagadorn & Belt, Reference Hagadorn and Belt2008; Young & Hagadorn, Reference Young and Hagadorn2010). The prevalence of modern medusozoan strandings suggests that, in spite of their gelatinous structure, medusa carcasses possess sufficient ductility to pass through the shore-break with their bells intact. However, once stranded and subaerially exposed, their high water content and immobility make them particularly susceptible to desiccation and scavenging.
Several of the features common among the Zabriskie specimens have also been invoked as key characters for the identification of other fossil medusa assemblages across the Phanerozoic fossil record (Young & Hagadorn, Reference Young and Hagadorn2010). Young & Hagadorn (Reference Young and Hagadorn2010) grouped these characters into the following categories: (1) fossils hav a general morphology consistent with that of modern cnidarian medusae; (2) fossils display the influence of biostratinomic factors, including pre-burial transport and desiccation; (3) fossils lack the perfect radial symmetry associated with abiotic sedimentary structures; (4) fossiliferous strata are interpreted to have been deposited in a near-shore environment similar to settings in which modern medusa strandings are commonly observed; (5) specimens occur en masse; and (6) the absence, in either bedding-plane or cross-sectional views, of evidence suggesting an abiotic, trace fossil or other body fossil origin. These criteria were formulated on the basis of uniformitarian principles supported by a comprehensive review of reports in the literature of putative fossilized medusae and of taphonomic studies of modern medusae (Schäfer & Craig, Reference Schäfer and Craig1972; Norris, Reference Norris1989). The independent observation of features of the Zabriskie specimens consistent with each of these criteria provides additional support for a medusozoan affinity for the Zabriskie specimens.
6. Taphonomy
Given the rarity with which nonmineralized tissues are preserved in the fossil record, taphonomic models detailing the biostratinomic and diagenetic histories of those relatively few examples of fossilized soft tissues are commonly the centrepiece around which new discoveries are described. These efforts have culminated in a well-defined taphonomic pathway for stranded jellyfish body fossils which emphasizes the variety of preservational features observed among these fossils in the gross context of a unique Cambrian shoreline (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Hagadorn & Belt, Reference Hagadorn and Belt2008; Young & Hagadorn, Reference Young and Hagadorn2010). Common taphonomic features shared by the Zabriskie fossils and other medusozoan fossil deposits include the absence of a well-developed sedimentary mixed layer, as well as evidence of subaerial exposure, including shrinkage and desiccation structures (cf. Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Hagadorn & Belt, Reference Hagadorn and Belt2008; Young & Hagadorn, Reference Young and Hagadorn2010). The discovery and analysis of the Zabriskie specimens thus not only enhances and pushes deeper in time the known fossil record of this important taxonomic group, but also tests several hypotheses concerning the biological, ecological and environmental requirements for medusozoan preservation in an entirely new sedimentary succession of significantly greater age than those hosting previously described assemblages, and from a new palaeogeographic setting.
The high abundance of specimens observed in the Zabriskie Quartzite, Potsdam Sandstone and Elk Mound Group suggests that the aggregation of medusozoans in near-shore modern environments was a strategy employed as early as the Cambrian period (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002). The apparently actualistic shoreward transport of medusozoans indicated by the preservation and facies associations of the Zabriskie fossils implies that the ductile and neutrally buoyant body plan of these organisms must have played an important role in their intact transport and preservation. Modern medusozoans are, in spite of their unmineralized anatomy, able to passively maintain their structural integrity in high-energy conditions and under turbulent flow while, conversely, skeletonized carcasses are commonly, under similar conditions, broken apart. Given the high morphological disparity characteristic of the Zabriskie fossil assemblage, it is possible that these fossils represent multiple generations of transport (i.e. strandings) and preservation. However, the absence of cross-cutting relationships or overlap suggests that the morphological disparity characteristic of these fossils reflects variable rates of decay, disparity between individuals stranded live and those stranded post-mortem, and potentially post-stranding secondary transport, rather than time-averaging (Fig. 5).

Figure 5. Transport model for the Zabriskie cnidarian medusa strandings. (a) Near-shore medusa bloom. Relatively undisturbed microbial mat (darker shading) in the intertidal zone. (b) Medusae migrate to the upper portions of the water column, potentially using incoming tide or wave energy to assist in migration further toward shore. Some individuals are damaged by wave activity. (c) Receding tide results in stranding and subaerial exposure of some of the medusae. Labels in (c) correspond to labelling of taphonomic models presented in Figure 6.
The morphology of the Zabriskie specimens, as well as that of other Cambrian medusozoan fossils, appears to have been strongly shaped by biostratinomic processes. Taphonomically mediated (rather than anatomically primary) features include striations – which we interpret to record fluid-mediated rilling (Fig. 3g) and/or folding (Fig 3d, e) – ripples within the central mound of the fossil (Fig. 3c), concentric rings interpreted to represent sequential stages in carcass desiccation (Fig. 3d, e) and striations at the fossils’ margin potentially linked to folding of the organism's bell (Fig. 3a, d). The preservation of Cambrian medusae in both concave and convex epirelief suggests that disparate individuals were characterized by disparity in collapse, or disparity in accumulation (within or immediately adjacent to a medusozoan) or winnowing of sediments associated with medusozoan carcasses. Disparity in topography both within and between specimens likely reflects some combination of: (1) sediment build-up around the perimeter of the animal's carcass, (2) sediment sealing by the body of the organism, (3) sediment deposition during collapse of the sand-filled endodermal elements, (4) active substrate disturbance by the attempts of the medusozoan to free itself from stranding, and (5) desiccation of the body of the organism during subaerial exposure (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Young & Hagadorn, Reference Young and Hagadorn2010). These scenarios are not mutually exclusive, so multiple conditions may and commonly do exist within a single fossil (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002).
The fossil margins of most medusa body fossils do not form distinct boundaries, but rather are gradational, making it difficult to distinguish the external margin of the carcass from the surrounding sedimentary matrix. This is particularly true among specimens characterized by external margins preserved as a rounded and gently sloping convex ridge (Fig. 3c, h). Previous models have interpreted these ridges to result from sediment accumulation around the perimeter of the organism (Young & Hagadorn, Reference Young and Hagadorn2010). However, in the absence of a well-defined fossil margin, it is difficult to discern whether sediment accumulated around and on top of the margins of the jellyfish or was stabilized below the organisms as a result of local sealing and/or capping of the substrate (thus preventing subsequent erosion) by the carcass (Fig. 6b). Constraining the true outermost perimeter of these fossils is essential, not only to accurately gauge the size of these organisms, but also to reconstruct the sedimentological processes which contributed to biostratinomic conditions. It should be noted that observational data suggest that the burial of modern jellyfish does not commonly result in additional sediment accumulation around the margins of the organism. Rather, most commonly, as waves wash over the organism, deflection of flow vectors toward those margins of the carcass oriented parallel to flow results in, as water flows oceanward, an increase in fluid velocity and thus sediment erosion around those peripheral margins of the carcass located parallel to flow. In contrast, certain specimens preserved as positive-relief ridges extending several centimetres above the surrounding substrate suggest that, occasionally, sediment baffling may occur below or surrounding some carcass margins (e.g. those margins located perpendicular to the direction of flow). Convex peripheral ridges observed in the largest Zabriskie specimen are oriented parallel to ripples preserved within low-relief regions interior of the fossil margins (Fig. 3c), suggesting that oscillatory currents across a partially decayed carcass or in the area vacated by reworking and removal of a carcass may have occurred prior to final burial of the bedding plane (Fig. 6).

Figure 6. Taphonomic model schematics for the various preservational modes observed in Zabriskie cnidarian medusae. Note that these preservational models are not mutually exclusive and that cnidarian medusa fossils are capable of preserving steps along any or multiple of these preservational pathways. Also note, the bells of modern medusae will typically experience significant flattening and widening upon stranding. For clarification of taphonomic features observed in the Zabriskie fossils, vertical relief in these medusa schematics has been exaggerated: (a) Initial medusa stranding. (b1a) Tide-mobilized sediment is removed from the margins of the medusa oriented parallel to current flow, or, (b1b) tide-mobilized sediment begins accumulating at the margins of the medusa. (b2) Medusa carcass either collapses or is completely removed by wave activity. (b3) Sedimentation continues, resulting in ripple formation in the depression left by the collapsed or removed medusa carcass. (b4) Example of this preservational style fromthe Zabriskie Quartzite (UCR11131-1). (c1a) Stranded medusa begins to pulsate, excavating a moat between two prominent ridges representing the proximal (inner ridge) and distal (outer ridge) reaches of the bell, or (c1b) stranded medusa begins to desiccate, sequentially excavating a moat between two prominent ridges representing the distal (outer ridge) reach of the bell and intermittent phases of desiccation. (c2) Medusa is buried and collapses. (c3) Collapse or flattening of the medusa results in deformation or flattening of the interior of the fossil. (c4) Example of this preservational style from the Zabriskie Quartzite (UCR11131-2). (d1) Deformation of the substrate below a stranded jellyfish, resulting in sediment mounding beneath the subumbrella and production of a well-defined marginal furrow. (d2) Medusa burial results in collapse of the organism. (d3) Lithification of the underlying sediments results in casting of the carcass. (d4) Example of this preservational style from the Zabriskie Quartzite (UCR11131-5). Scale bars are 5 cm.
Concave discoidal rings, or depressions between two ridges, are also common among medusa body fossils (Fig. 3d, e, f). Previous models have attributed external concave rings to the contracting bell of a live-stranded medusa (Norris, Reference Norris1989; Bruton, Reference Bruton, Simonetta and Morris1991; Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Young & Hagadorn, Reference Young and Hagadorn2010). The inner and outer edges of the concave ring are interpreted to represent the proximal and distal reaches, respectively, of the bell (Fig. 6c1a). In a similar but unidirectional action, shrinkage of a medusa carcass may also produce circumscribing moats during contraction of the organism's body (Fig. 6c1 b). Shrinkage likely reflects desiccation; gradual desiccation would result in sequential shrinkage and, at each stage, sediment ‘strand lines’ would form at the new external margins of the medusa. Multiple distinct, concentric, low-relief convex ridges surrounding several Zabriskie specimens suggest that certain specimens likely experienced this type of sequential desiccation of the medusa carcass (Fig. 3d).
The finest detail preserved in stranded medusa body fossils occurs in specimens preserved wholly in convex relief, commonly as a generally smooth surface bearing some impression of not only the dorsal but also the ventral portion of the organism (Figs 3a,b, d). These impressions are characterized by a mound of sediment near the centre of the fossil, with some examples preserving quadriradiate spurs extending from the centre of this mound (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002). Previous models for comparable specimens collected from the Elk Mound Group have invoked incorporation of suspended sediment prior to stranding of the medusa (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Young & Hagadorn, Reference Young and Hagadorn2010). This model is explained by the following sequence:
(1) Prior to being washed onto shore, medusae transported to shallow, turbid water in the intertidal zone may pulse their bells and, in so doing, incorporate sediment into their gastrovascular cavity.
(2) Following stranding and deterioration of the medusa tissues, the sediment load carried by the animal may result in formation of a distinct mound beneath the subumbrella. Deposition of the medusa may result in folding of some portion of the bell and endodermal elements, as well as intercalation with sediment, resulting in striations and uneven topography in the centre of the resulting medusa fossil.
(3) This sediment mound may be augmented by excavation of the surrounding sediment by the bell of the medusa, if stranded alive as an escape mechanism.
While sediment ingestion may explain mounding in some Zabriskie specimens (Fig. 6d), this model is not required for the preservation of all positive-relief specimens. Mounds comparable to those observed in the fossil record are commonly produced below modern jellyfish, entirely as a result of sedimentological processes with little to no evidence of sediment ingestion.
During decomposition of the organism, the body of the jellyfish and the mucus lining the umbrella and tentacles facilitate sealing of the underlying substrate, making the area temporarily more resistant to erosion. Given the relatively limited extent of grazing along Cambrian beaches and resulting relative abundance of microbial mats in near-shore setting, it is most likely that the substrate was further sealed by growth of microbial mats along the sediment–water interface and between sand grains, thus inhibiting organic matter degradation through macrofaunal and meiofaunal scavenging or aerobic microbial respiration (Seilacher, Reference Seilacher1999; Noffke, Knoll & Grotzinger, Reference Noffke, Knoll and Grotzinger2002; Gehling & Droser, Reference Gehling and Droser2009). This hypothesis is supported by the presence of sedimentary structures of likely microbial origin within the Zabriskie section, bracketing the fossiliferous horizon, and abundant organically mediated structures preserved in immediate spatial association with several assemblages of medusozoan body fossils from Cambrian strata of Wisconsin and New York (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Hagadorn & Belt, Reference Hagadorn and Belt2008).
The collective taphonomic history of these fossils is fundamentally distinct from body fossils of other nonmineralized forms in that the general morphology of the Zabriskie specimens largely reflects ambient sedimentological processes rather than post-burial diagenetic processes associated with tissue degradation. The taphonomic framework established for the Zabriskie fossils suggests that, in contrast to fossils preserved as carbonaceous compressions or through permineralization, sandstone-hosted medusozoan fossils largely reflect sedimentary dynamics; in particular, the morphology of medusozoan fossils is a function of (1) interactions between the living organism, the surrounding fluids and the substrate; and (2) post-mortem fluid–carcass interactions that result in variations in sediment-carrying capacity and thus locally variable sediment removal and accumulation. Although these fluid–organism interactions have actualistic analogues, the factors responsible for their preservation in the fossil record reflect anactualistic environmental and ecological conditions unique to the Cambrian shoreline.
7. An analogue for Ediacaran-style preservation?
Preservation of soft-bodied macrofauna in shallow marine sandstones is largely limited to the Neoproterozoic and early Palaeozoic. As the vast majority of Ediacaran organisms disappeared with the onset of the Phanerozoic Eon, consideration of the requirements for soft-bodied preservation across the Precambrian–Cambrian transition, rather than taxonomic comparison alone, is essential to distinguish between a potential mass extinction event and a preservational bias in the fossil record (cf. Seilacher, Buatois & Mangano, Reference Seilacher, Buatois and Mangano2005; Laflamme et al. Reference Laflamme, Darroch, Tweedt, Peterson and Erwin2013; Buatois et al. Reference Buatois, Narbonne, Mángano, Carmona and Myrow2014). The discovery of the Zabriskie specimens extends the record of Phanerozoic examples of preservation of medusae in sandstones back in time by several tens of millions of years, considerably closer to the Precambrian–Cambrian boundary, increasing the need for additional comparison of Neoproterozoic and early Palaeozoic taphonomic models.
While the taphonomy of Cambrian medusa body fossils is generally comparable to that of Ediacaran-aged fossil assemblages, the preservational histories and palaeoenvironmental distribution of these two groups are markedly distinct. Conditions which may have heightened the preservation potential of soft-bodied organisms in near-shore settings across the Precambrian–Cambrian transition include limited scavenging and the absence of a well-developed mixed layer (Tarhan & Droser, Reference Tarhan and Droser2014; Tarhan et al. Reference Tarhan, Droser, Planavsky and Johnston2015b ), which, if well developed, would have increased the oxygenation of seafloor sediments and thus rates of bacterial decomposition, as well as the erodibility of the substrate (Bottjer, Hagadorn and Dornbos, Reference Bottjer, Hagadorn and Dornbos2000; Droser, Jensen & Gehling, Reference Droser, Jensen and Gehling2002a ; Dornbos, Bottjer & Chen,et al Reference Dornbos, Bottjer and Chen2005; Jensen, Droser & Gehling, Reference Jensen, Droser and Gehling2005; Seilacher, Buatois & Mangano, Reference Seilacher, Buatois and Mangano2005; Dornbos, Reference Dornbos2006; Tarhan et al. Reference Tarhan, Droser, Planavsky and Johnston2015b ). Ediacara-style preservation of soft-bodied organisms has long been attributed to unique diagenetic conditions resulting from the presence of extensive microbial matgrounds, which acted as sediment seals and loci for authigenic mineralization, resulting in the formation of detailed sandstone casts and moulds of a diverse array of organisms (Seilacher & Pflüger, Reference Seilacher, Pflüger and Paterson1994; Gehling, Reference Gehling1999; Mapstone & McIlroy, Reference Mapstone and McIlroy2006; Callow & Brasier, Reference Callow and Brasier2009; Gehling & Droser, Reference Gehling and Droser2009, Reference Gehling and Droser2013; Tarhan et al. Reference Tarhan, Droser, Gehling and Dzaugis2015a ). In contrast, Cambrian examples of mouldic preservation of soft-bodied organisms in near-shore sandstones likely reflect environmental controls upon sediment reworking, scavenging and bioturbation. While microbial fabrics likely helped to bind the substrate and also limit infaunal disturbance (Seilacher & Pflüger, Reference Seilacher, Pflüger and Paterson1994; Jensen, Droser & Gehling, Reference Jensen, Droser and Gehling2005), the presence of microbial mats does not appear to have exercised a direct control upon the diagenetic processes that led to the preservation of Cambrian medusae. In addition to these diagenetic differences, the palaeoenvironments represented by sandstone-hosted soft-bodied macrofossils are also distinct across the Precambrian–Cambrian boundary. Fossils of the Ediacara Biota are exceedingly rare in shoreface sandstone facies (Gehling & Droser, Reference Gehling and Droser2013), whereas taphonomically similar Cambrian faunas are more common in near-shore (intertidal) palaeoenvironments. These facies associations suggest a significant disparity between Ediacaran and Cambrian palaeoenvironments conducive to fossil preservation. As depositional environment is a first-order control on preservation, recognition of the differences among the palaeoenvironments represented by Ediacaran and lower Cambrian fossiliferous sandstones is key to discussions regarding the fidelity and continuity of the fossil record through this critical time in the history of life.
8. Significance
The rarity of fossilized medusae, coupled with the importance of these animals in modern ecosystems, highlights the significance of expanding both the temporal and spatial distribution of the medusozoan fossil dataset. The discovery of these fossils not only extends the fossil record of cnidarian medusa strandings back several tens of millions of years to the early Cambrian, but also further substantiates claims that jellyfish were abundant and widespread components of Cambrian marine ecosystems (Hagadorn, Dott & Damrow, Reference Hagadorn, Dott and Damrow2002; Hagadorn & Belt, Reference Hagadorn and Belt2008; Young & Hagadorn, Reference Young and Hagadorn2010; Erwin et al. Reference Erwin, Laflamme, Tweedt, Sperling, Pisani and Peterson2011).
Additionally, these fossils contain a wealth of taphonomic information that may shed considerable light upon the anactualistic biostratinomic and diagenetic conditions present within near-shore settings at the onset of the Phanerozoic. In spite of the high water content and near-neutral buoyancy of these organisms, the unique physiological properties of medusozoans are likely what, to some extent, allowed these animals to remain intact during transport to the shoreline to produce the sedimentary structures now preserved in the rock record.
While rare, the highest concentrations of putative medusozoan body fossils occur in early Phanerozoic and particularly Cambrian strata, consistent with the temporal distribution of nonmineralized fossil-bearing deposits as a whole (Briggs, Reference Briggs2003; Gaines, Kennedy & Droser, Reference Gaines, Kennedy and Droser2005; Gaines, Briggs & Yuanlong, Reference Gaines, Briggs and Yuanlong2008; Garson et al. Reference Garson, Gaines, Droser, Liddell and Sappenfield2012). Along the early Cambrian shoreline, the confluence between a depauperate terrestrial biosphere and an ocean teeming with complex metazoan life would have yielded a fundamentally distinct environment that, in rare instances, served as a preservational sanctuary for medusozoan fossilization. The preservation of the Zabriskie fossils in early Cambrian sediments thus supports the model first introduced by Hagadorn, Dott & Damrow, (Reference Hagadorn, Dott and Damrow2002) that the preferred environment for soft-bodied preservation in sandstone shifted, over the Precambrian–Cambrian transition, from subtidal to intertidal environments. This shift in preferred environment from below fair-weather wavebase to the shoreline could reasonably account for the disparity in taxa and differences in preservation represented on either side of the Precambrian–Cambrian boundary. Thus, in evaluating the role of taphonomy through the Precambrian–Cambrian transition as a potential driver for the disappearance of the Ediacara Biota, it is important to recognize that the persistence of matgrounds into the Phanerozoic cannot be used as the sole litmus test for a taphonomic continuum across the Precambrian–Cambrian boundary, as has recently been proposed (Laflamme et al. Reference Laflamme, Darroch, Tweedt, Peterson and Erwin2013; Buatois et al. Reference Buatois, Narbonne, Mángano, Carmona and Myrow2014).
9. Conclusions
Reconstruction of not only the palaeobiology and palaeoecology, but also taphonomic controls upon the lower Cambrian fossil record, is critical to understanding of environmental and ecological evolution during one of the most significant intervals in the history of life on Earth. The fossil record of cnidarian medusae provides a unique resource for reconstructing both environmental and palaeobiological dynamics concomitant with the diversification of early metazoan life. The preservational pathway described herein refines previous models for the preservation of cnidarian medusae by emphasizing the significance of sedimentological processes surrounding a gelatinous organism stranded along a microbially bound substrate, rather than attributing this unique style of preservation to carcass diagenesis alone. The morphological similarities of modern stranded medusae and the medusozoan fossil record, as well as the actualistic record of shoreline sedimentological processes, suggest that a uniformitarian approach can be applied to evaluating the origin of the features characterizing the Zabriskie fossils. However, as indicated by the paucity of fossilized medusozoan strandings, anactualistic environmental requirements for medusozoan fossilization in marginal marine settings – such as the absence of bioturbation and scavenging, and sediment stabilization by microbial binding – were largely lacking during the later Phanerozoic. Thus, a significant portion of the fossil record of cnidarian medusae is intricately linked to early Palaeozoic substrate evolution and, in particular, the protracted development of infaunal sediment mixing (Tarhan et al. Reference Tarhan, Droser, Planavsky and Johnston2015b ).
Acknowledgements
We acknowledge funding for field work from National Science Foundation Grant EAR-9219731 (to M.L.D.), the Paleontological Society, SEPM, Southern California Academy of Sciences, American Federation of Mineralogical Societies, Geosyntec Consultants, Inc. and the Geological Society of America (to A.S.). L.G.T.’s research was supported by a National Science Foundation Graduate Research Fellowship, Yale University Postdoctoral Fellowship and National Science Foundation Earth Sciences Postdoctoral Fellowship. J. Esteve, S. Henry, J. Wine and C. Sappenfield assisted with field work.