1. Introduction
Late Triassic and Jurassic fissure fill deposits occur on both sides of the Bristol Channel in southwest England and South Wales (Fig. 1). These deposits, which occur within the Carboniferous Limestone, have produced internationally important faunas from which many new genera of reptiles (principally archosaurs and sphenodontian lepidosaurs) and mammals have been described (Table 1). Indeed, it was from the fissure deposits of Durdham Down in Bristol that the small prosauropod dinosaur Thecodontosaurus was described in 1836 and 1840 by Riley & Stutchbury, thus making this one of the the first dinosaurs to be named (Benton et al. Reference Benton, Juul, Storrs and Galton2000). The history of research has been reviewed by Savage (Reference Savage1993) and the fissure faunas themselves are reviewed by Fraser (Reference Fraser, Fraser and Sues1994), Evans & Kermack (Reference Evans, Kermack, Fraser and Sues1994) and Benton & Spencer (Reference Benton, Spencer, Wimbledon and Palmer1995). Significant publications that post-date these reviews include Rauhut & Hungerbühler (Reference Rauhut and Hungerbühler2000), Fraser et al. (Reference Fraser, Padian, Walkden and Davis2002), Renesto & Fraser (Reference Renesto and Fraser2003), Yates (Reference Yates2003), Cuny (Reference Cuny2004) and Säila (Reference Säila2005).
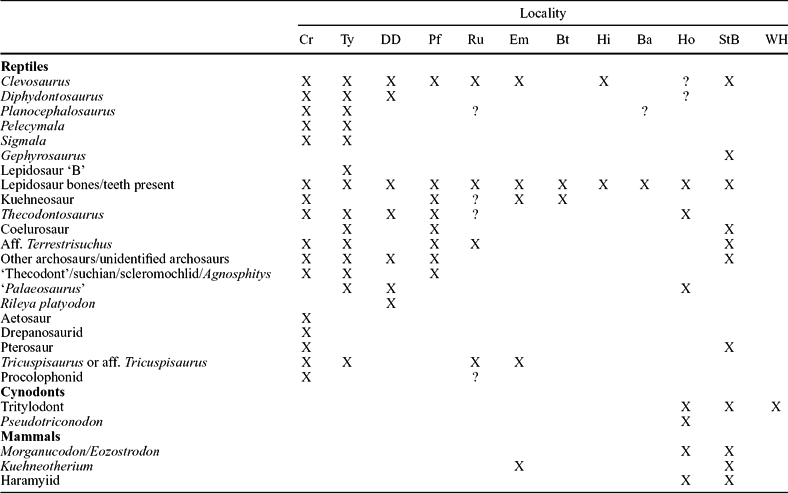
Table 1. Tetrapod faunas from Late Triassic and Early Jurassic fissure localities in SW Britain
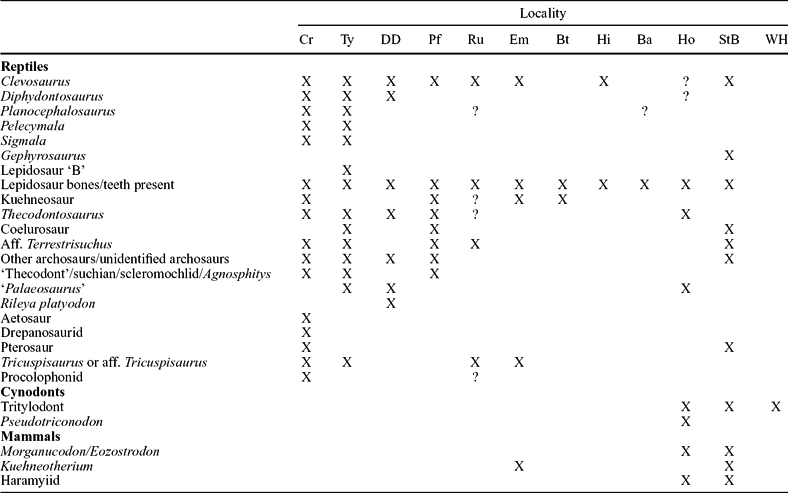
Data from D. I. Whiteside (unpub. Ph.D. thesis, Univ. Bristol, 1984), Walkden & Fraser (Reference Fraser, Lucas and Morales1993), Fraser (Reference Fraser, Fraser and Sues1994), Evans & Kermack (Reference Evans, Kermack, Fraser and Sues1994), Benton & Spencer (Reference Benton, Spencer, Wimbledon and Palmer1995), Fraser et al. (Reference Fraser, Padian, Walkden and Davis2002), Renesto & Fraser (Reference Renesto and Fraser2003), Cuny (Reference Cuny2004), Säila (Reference Säila2005), S. E. Evans (pers. comm.) and personal observations. X – confirmed presence and ? – unconfirmed presence. Cr – Cromhall; Ty – Tytherington; DD – Durdham Down; Pf – Pant-y-ffynon; Ru – Ruthin; Em –Emborough; Bt – Batscombe; Hi – Highcroft; Ba – Barnhill; Ho – Holwell; StB – quarries on St Brides palaeo-island (Pant, Duchy, Ewenny and Pontalun) and WH – Windsor Hill. Note the similarities of the fauna in the first nine localities from Cromhall to Barnhill which produce the ‘sauropsid-type’ fauna of Robinson (Reference Robinson1957a) where mammals are generally lacking (except Emborough), and the last three quarries that have distinctive cynodont and mammalian components. Note also that there are no records of amphibians (confirmed for the Welsh fissures by S. E. Evans, pers. comm.) in any fissure deposit.
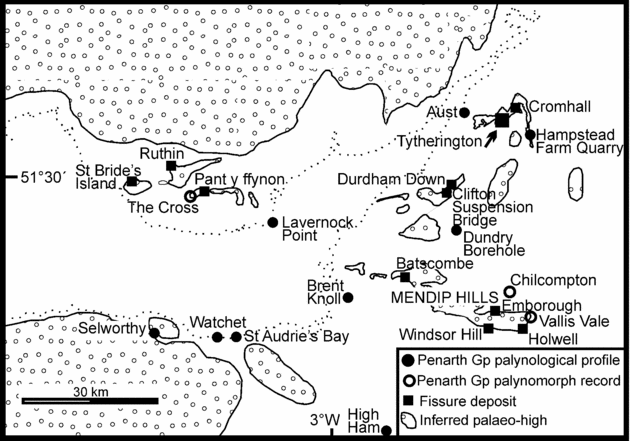
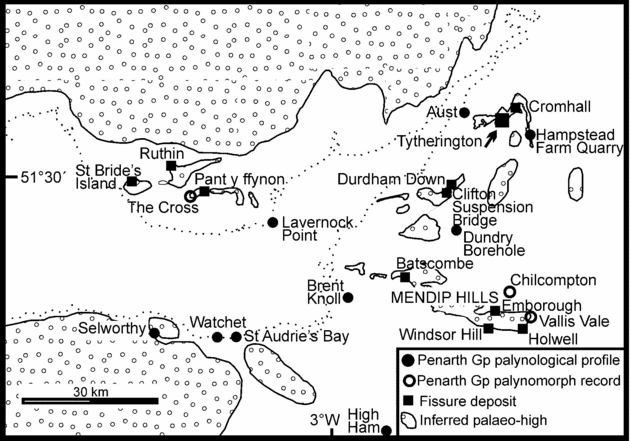
Figure 1. Map showing the principal Upper Triassic/Lower Jurassic tetrapod-bearing fissure deposits of the English West Country around Bristol and in South Wales. The three important groups of fissure localities are Durdham Down (in Bristol), Tytherington and Cromhall, north of Bristol; Holwell, Emborough, Windsor Hill and Batscombe in the Mendip Hills; and a west of Cardiff group including Pant-y-ffynon and Ruthin, together with those on St Bride's palaeo-island (e.g. Pant, Duchy, Ewenny and Pontalun). The map shows the inferred latest Triassic palaeo-highs that would have formed an archipelago of small limestone islands lying in a Rhaetian sea. A UK location map is shown on Figure 2. The map also shows the locations of complete Penarth Group palynological profiles and the more limited spot sample palynomorph records. Compiled from Ordnance Survey Digimap © and the BGS 1:250000 solid geology Bristol Channel sheet with input from Audley-Charles (Reference Audley-Charles1970), Tucker (Reference Tucker1977), Kellaway & Welch (Reference Kellaway and Welch1993) and Robinson (Reference Robinson1957a). Sources for the palynological data points are The Cross & Lavernock Point (Orbell, Reference Orbell1973); Brent Knoll Borehole (Warrington, Reference Warrington, Neale and Brasier1981); Watchet and St Audries Bay (Warrington & Whittaker, Reference Warrington and Whittaker1984); Selworthy (Warrington in Edwards, Reference Edwards1999; Warrington et al. Reference Warrington, Ivimey-Cook, Edwards and Whittaker1995); High Ham (Warrington, Whittaker & Scrivener, 1986); Chilcompton and Vallis Vale (Warrington, Reference Warrington1984); Dundry Borehole (Warrington in Kellaway & Welch, Reference Kellaway and Welch1993); Aust (Manor Farm Quarry, pers. obs.) and Hampstead Farm Quarry, Chipping Sodbury (this paper).
The many fissure-derived assemblages are comprised of extremely well-preserved individual bones and some complete or partially complete skeletons. Huge amounts of material have been processed by many researchers, but there is no evidence (e.g. teeth or phalanges) of any animals bigger than about 3 m. The variety of small animals from these fissure deposits has made an unparalleled contribution to a more complete understanding of the Late Triassic–Early Jurassic tetrapod biota. The Tytherington fissure system itself is noteworthy as the type locality for Diphydontosaurus avonis Whiteside, Reference Whiteside1986, and a spectacular new find of Thecodontosaurus antiquus Morris, Reference Morris1843, the ‘Bristol dinosaur’.
However, despite this long history of study, the dating of the mainly reptile-bearing deposits of the Bristol region and South Wales and the mammal-bearing fissures of South Wales has been problematic. Although similar reptiles are found with ‘Rhaetic’ invertebrates and fish at Holwell, most authors have usually regarded the reptile-bearing fissures (the ‘sauropsid fissures’ of Robinson, Reference Robinson1957a, also known as the ‘complex-A’ assemblage of Shubin & Sues, Reference Shubin and Sues1991) as having formed in underground water-courses during ‘Keuper’ or Norian times and therefore pre-dating the Penarth Group. This age assignment was based on two arguments outlined by Robinson (Reference Robinson1957a). The first concerns apparent lithological similarities to the bedded red and green mudstone in the local Blue Anchor Formation. The second concerns field relationships at Emborough Quarry, where Robinson proposed that the ‘Rhaetic’ would have capped the reptile-bearing fissure, which therefore must have been ‘pre-Rhaetic’. Except for two Kuehneotherium teeth from Emborough (Fraser, Walkden & Stewart, 1985), the sauropsid fissures have not yielded any confirmed mammalian fossils and the main mammal-bearing fissures have, therefore, generally been regarded as younger. The fissure faunas have also been regarded as representing a unique ‘upland’ fauna, on the basis that the fossils are from Mesozoic caves in limestone hills and comprise small reptiles, including small dinosaurs (Robinson, Reference Robinson1957a; Tarlo, Reference Tarlo1962; Halstead & Nicoll, Reference Halstead and Nicoll1971; Halstead & Halstead, Reference Halstead and Halstead1981).
Both the pre-Penarth Group age and the postulated ‘upland’ environment for the sauropsid fissures were challenged by Marshall & Whiteside (Reference Marshall and Whiteside1980), who discovered terrestrial and marine palynomorphs in a reptile-bearing fissure system in Tytherington Quarry. The palynomorphs included dinoflagellate cysts and indicated a Rhaetian age, and a stratigraphical level equivalent to the top of the Westbury Formation, and showed that the environment was not upland but marginal marine. Whiteside & Robinson (Reference Whiteside and Robinson1983) provided further evidence, from a glauconitic clay mineral out of the same fissure, for a marginal marine location with the system being located at the margin of a freshwater lens abutting saline waters. Whiteside (Reference Whiteside1986) reported the osteichthyan fish, Pholidophorus, from Tytherington and Cromhall, thus indicating that both quarries contained Rhaetian-age fissure deposits.
Despite this direct and independent age for the age of the Tytherington fissures, the arguments for an earlier origin for ‘sauropsid fissures’ were restated by Simms, Ruffell & Johnston (1994), who regarded them as having formed during an early Late Triassic pluvial episode (Carnian) with infilling in later Triassic times. This was despite an admission that no definitive Carnian sediments had been found and with no direct evidence for a Norian age. Walkden & Fraser (Reference Fraser, Lucas and Morales1993) had developed a similar argument for the age of the Cromhall fissure infills. By matching a sequence of sedimentological and diagenetic associations against proposed Late Triassic climatic and eustatic events, they dated their three faunal associations as ?Carnian, mid-late Norian and Rhaetian. There was, however, no incontrovertible palaeontological evidence for this interpretation.
In this paper we present the first detailed description of the morphology of the Tytherington fissures, the nature of their infill and their fossil content. This, the only detailed modern account of the geology of such a fissure system, provides key evidence for the age and environment of the reptiles including Thecodontosaurus. The evidence from Tytherington demonstrates that the reptile-bearing deposits are of Rhaetian age and occupy fissures formed on a small limestone island. There is no prima facie evidence that any of the ‘sauropsid fissure’ infills in southwest Britain and their tetrapods date from pre-Penarth Group times. In this fundamental respect we disagree with Fraser (Reference Fraser1988a), Fraser & Walkden (Reference Fraser and Walkden1983), Benton (Reference Benton, Fraser and Sues1994), Benton & Spencer (Reference Benton, Spencer, Wimbledon and Palmer1995) and Lucas (Reference Lucas1999), who generally regard these reptile-bearing fissures, particularly those at Cromhall that have yielded the most diverse herpetofauna, as older than the Rhaetian transgression and therefore as Norian in age. Although Walkden & Fraser (Reference Fraser, Lucas and Morales1993) and Fraser (Reference Fraser, Fraser and Sues1994) now consider that some deposits containing Clevosaurus hudsoni Swinton, Reference Swinton1939, and Diphydontosaurus at Cromhall as Rhaetian in age, they still maintain that most reptile-bearing fissures at that locality pre-date the basal Penarth Group.
The dating of the Tytherington reptiles also has implications for the debate about a single, abrupt, end-Triassic mass extinction event versus a prolonged multi-phase event, as the ranges of the reptile families represented at Tytherington are shown to extend into the Rhaetian rather than terminating in the Norian.
2. The Tytherington fissures
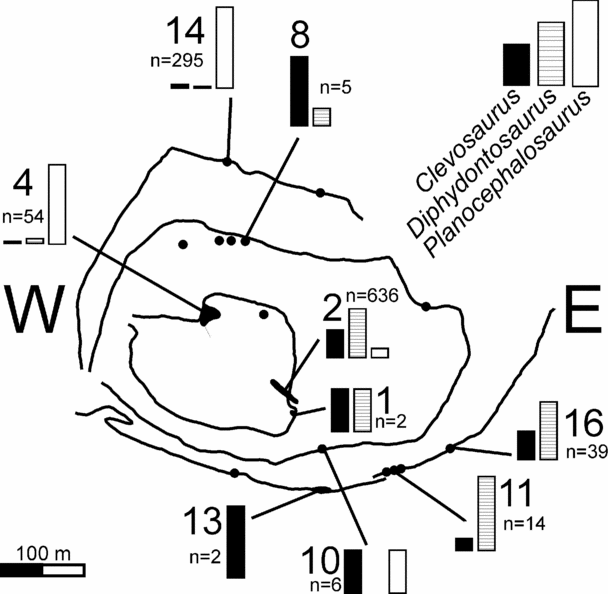
In 1975 an important new vertebrate fossil-bearing fissure system was discovered by the amateur geologists Mike Curtis and Tom Ralph in Tytherington Quarry, South Gloucestershire, UK (ST 660890). Active quarrying rendered much of the rock infilling the fissure available for study, although this necessitated the immediate removal, by R. J. G. Savage, of several tonnes of rock to the then Department of Geology at the University of Bristol. Collection of this new material initiated a systematic study of this and other fissures subsequently discovered in the quarry. The initial findings were described by D. I. Whiteside (unpub. Ph.D. thesis, Univ. Bristol, 1984), and these contribute to some of the data and conclusions reported in this paper. Sixteen fissures were discovered (Figs 2, 3) and documented during regular and frequent visits from 1976 to 1983, and infrequent visits up to 1992, paralleled by research on the deposits up to the present day. Quarrying operations destroyed many of the fissures, and observations and sampling could only be carried out when safe access was available. Hence some fissures are known in less detail. Some of the vertebrate fossils have been described by Whiteside (Reference Whiteside1986), and preparation and study of the collection continues.
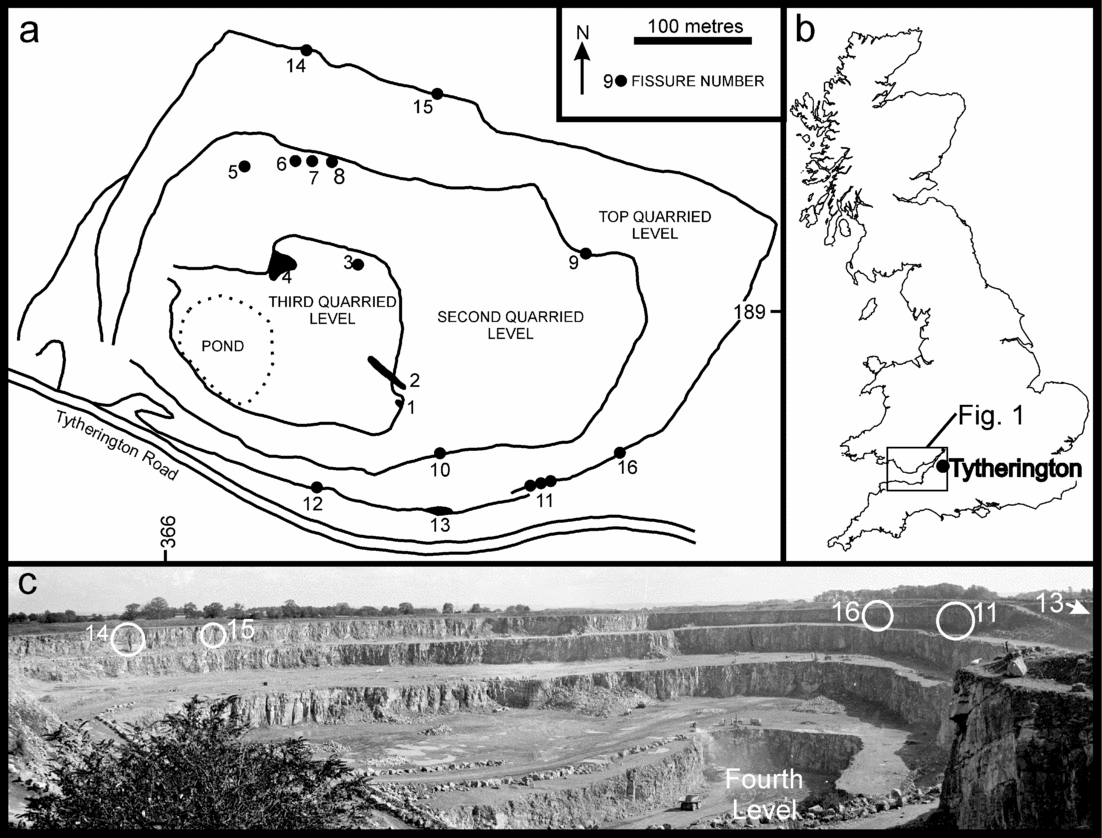
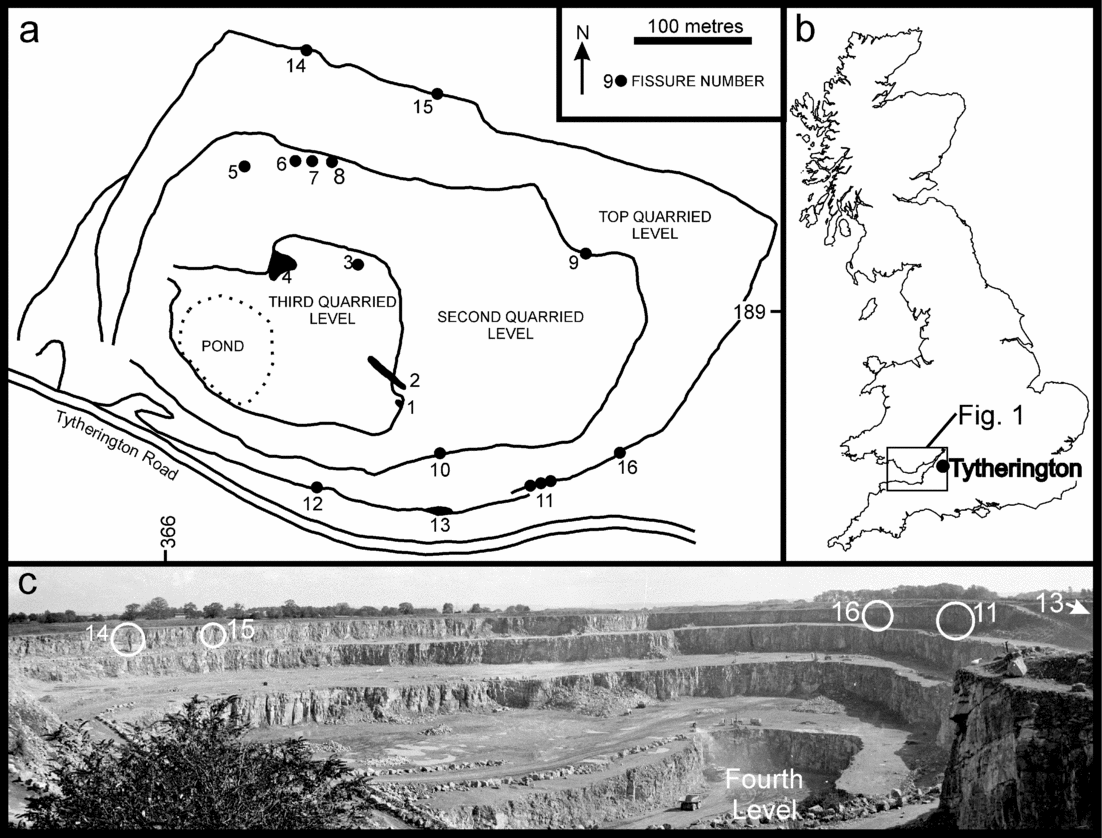
Figure 2. (a) Map of Tytherington Quarry (derived from map supplied by Amey Roadstone Corporation, 1981) showing location of the fissure localities referred to in the text. (b) Map of the UK showing the location of Tytherington and the area of Figure 1. (c) Photograph of Tytherington Quarry, looking to the east, taken in May, 1982. Only fissures 12 (not shown), 11, 13, 14, 15 and 16 (circled) in the top quarried level are still present. The other fissures have been destroyed by quarrying operations. Compared to the map of the quarry (a) there is now a fourth quarried level. No vertebrate fossils or significant fissures were found in the fourth level.

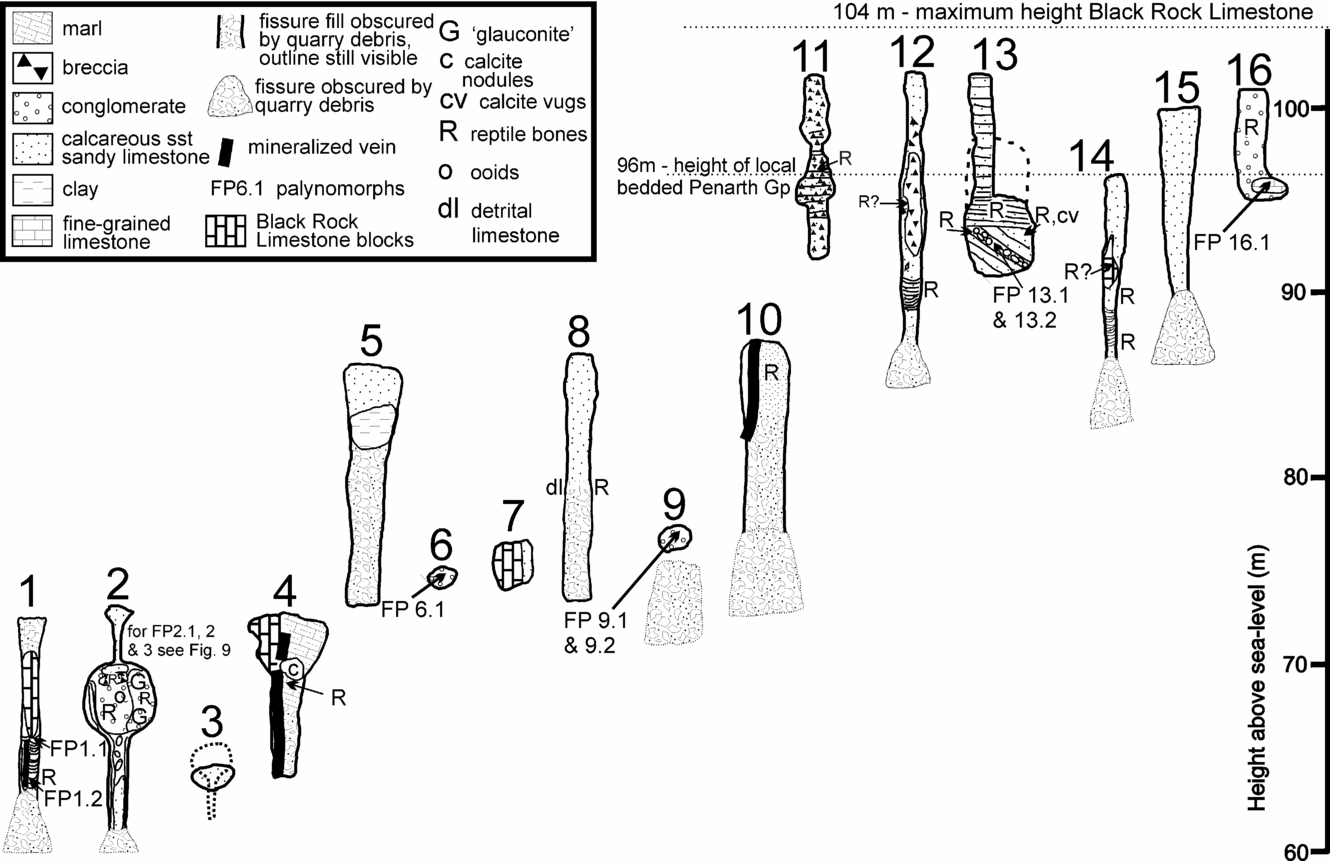
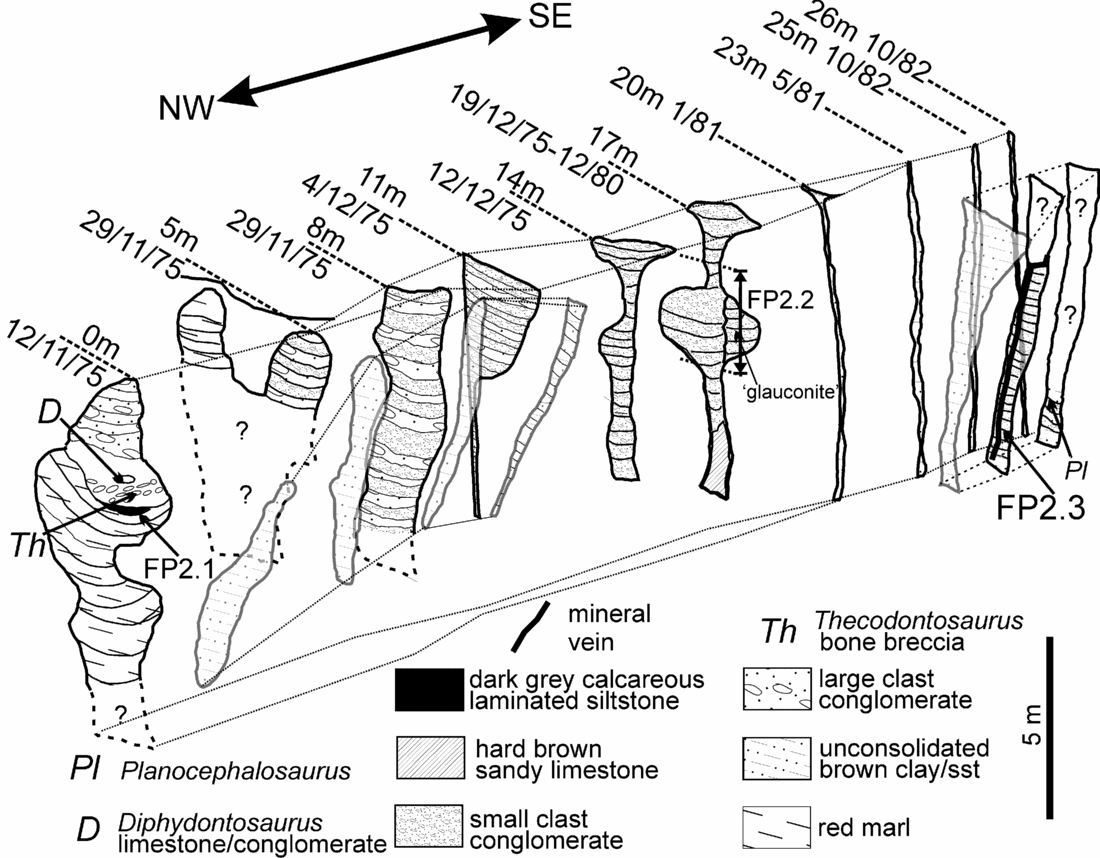
Figure 3. Cross-sections of the 16 Triassic fissure localities described in the text. Except for the top quarried level that retained the original limestone surface, the vertical extents of the fissures are generally unknown due to earlier quarrying operations or unexcavated base levels. The upper and lower parts of some fissure fills were also obscured by quarry scree. The uppermost parts of fissures 1, 5, 8, 11, 12, 14, 15 and 16 were observed through binoculars and details described from those observations, telephoto slides and hand collection of fallen rocks. The remaining sections were completed from in situ rock collections. Note the diversity of morphology and fill lithologies. The locations of the reptile bones and the productive palynological samples are shown.
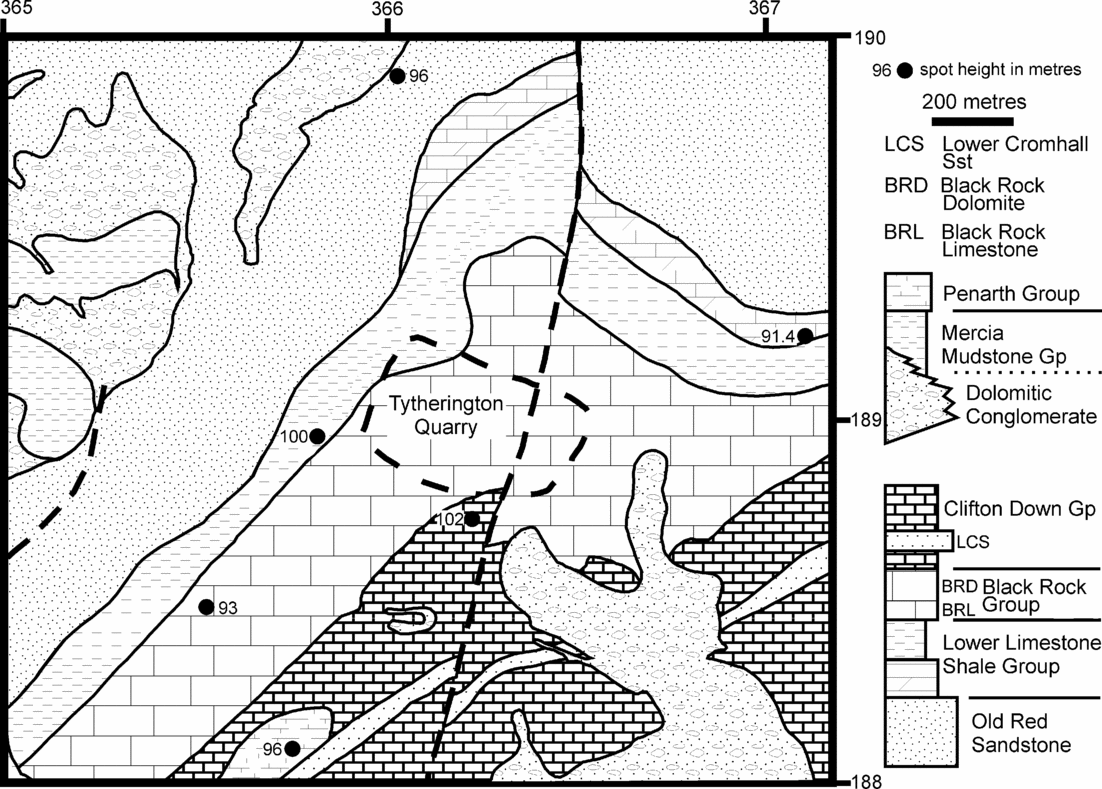
Tytherington Quarry (Figs 2, 4) lies on the western edge of the Parkfield–Coalpit Heath Basin, some 15 km NE of Bristol (Fig. 1). All the fissures are in the Black Rock Limestone Formation, which is a crinoidal limestone of Early Dinantian (Courceyan–Chadian) age (Kellaway & Welch, Reference Kellaway and Welch1993). This formation dips at about 20° to the SE and is underlain by the Lower Limestone Shale Group and the Tintern Sandstone Group (Old Red Sandstone), both of which crop out to the west, with the latter forming an elevated ridge (Fig. 4). Within 0.6 km of the quarry, various units of Triassic age (Dolomitic Conglomerate, Mercia Mudstone Group and Penarth Group) lie unconformably on the Palaeozoic strata.

Figure 4. Simplified large-scale geological map and geological succession of the area around Tytherington Quarry. The limestone was quarried from the Black Rock Group. Also note the outcrop of bedded Penarth Group at a height of 96 m, about 600 m south of the quarry. From BGS map sheet ST 68 NE, co-ordinates used are British National Grid. Geological column from Kellaway & Welch (Reference Kellaway and Welch1993).
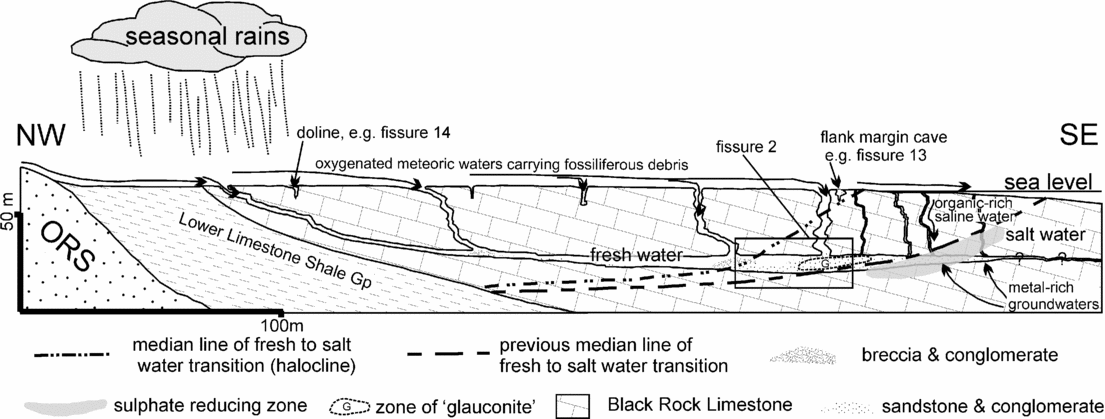
The fissures are generally orientated E–W or NW–SE along vertical joint planes, and are therefore aligned with the local joint pattern in the Black Rock Limestone. Although the time of origin of this joint system is unknown, the joints would have undergone dilation during the Mid- to Late Triassic episode of tectonic extension and basin development (Miliorizos & Ruffell, Reference Miliorizos, Ruffell and Underhill1998; Ruffell & Shelton, Reference Ruffell and Shelton1999) that affected the southern UK. Following extension, these joint systems were then modified by solution processes. However, some Tytherington fissures (for example, fissure 14) are true palaeo-dolines, with funnel-shaped entrances that narrow downwards.
The sixteen fissures known from Tytherington Quarry are shown in plan view in Figure 2. They can only be recognized in cross-section in the back-face of each excavated level and therefore their complete vertical extents are unknown. However, those fissures on the top level have the original pre-quarrying limestone surface intact. The morphology, infill and fossil content of the fissures are described below. Figure 3 shows their topographical relationship and a diagrammatic representation of the infilling lithology.
2.b. General fissure morphologies (Fig. 3)
Fissures 1, 2, 4 and 10 are aligned along a major vertical NW–SE-trending joint plane and are probably exposures of the same fissure infill. However, solution has taken place along this joint, and the cross-section varies from a 1 to 4 m wide vertical-sided slot to circular or simply a joint with no significant gap and little or no infill. Hydrothermal activity occurred in parts of this fissure system, as demonstrated by the presence of minerals such as barytes, galena and sphalerite. Fissures 6 and 8 were formed by solution of the Carboniferous Limestone, producing, in the case of fissure 6, a sub-horizontal tube or in fissure 8 a doline-like form along an E–W joint-plane that may have extended to fissure 9. Fissures 12, 14 and 16 are vertical tubes formed by solution and are here identified as true palaeo-dolines. The three exposures of fissure 11 have doline-like characteristics and trend along an E–W joint; fissure 13 shows a cavernous development that is also related to an E–W joint. The unfossiliferous fissures 3, 5, 7 and 15 are, respectively, a solution-produced tube with a vertical notch cut into the bottom in some cross-sections, a doline-like form produced by solution-widening of a joint, a minor clay-filled, solution-produced tube with pronounced metasomatic changes in the surrounding limestone and a doline-like fissure on the top quarrying level.
Most of the fissure infills (except one type in fissure 2 and part of the fissure 1 deposits) are horizontally or sub-horizontally bedded, indicating that unconsolidated sediment derived from the limestone surface accumulated under sub-aqueous or sub-aerial conditions. This type of bedding is characteristic of a surface hollow, cave or old sinkhole (old doline). In contrast, infills that show crude bedding parallel to the host limestone walls of fissures are derived from bedded rocks overlying the limestone surface that were pushed/sucked into the fissures during subsidence/extension episodes (Walsh et al. Reference Walsh, Boulter, Ijtaba and Urbani1972; Wall & Jenkyns, Reference Wall and Jenkyns2004).
3. The fossil-bearing deposits
The fauna and palynomorphs found in the Tytherington fissures are shown in Figures 5 and 6. The infill of fissure 1 (fig. 7a–d) consisted of a repeated sequence of upward-fining black conglomeratic sandy mudstones that yielded a small fragment of the dentary of Diphydontosaurus avonis, and a worn Clevosaurus tooth (fig. 5c, d), together with Gyrolepis albertii scales, a Severnichthys tooth (see fig. 5a), fish gill rakers and internal casts referable to Penarth Group gastropods. The marine fish remains and the lower jaw of Diphydontosaurus avonis are black, whereas those of the Clevosaurus are yellowish-brown. A palynomorph assemblage, FP1.1 (Fig. 6, Table 2), was discovered in the same matrix as the vertebrates and another palynomorph assemblage, FP1.2, was found below them. These assemblages include small amounts of fusain (charcoal) up to 2 mm long (fig. 5b). Similar black conglomeratic lithologies in fissures 6 and 9 have produced palynomorph assemblages (FP6.1, FP9.1, FP9.2; Fig. 3, Table 2) in the same matrix as blackened fragments of Gyrolepis albertii scales, gill rakers, other ‘Rhaetic’ fish, and unblackened pieces of reptile bones. The lithology of fissure 9 consisted mainly of conglomeratic black clay comprising palynomorph-bearing clasts and matrix; some clasts of black clay were also found in large nodules of galena and sphalerite in this fissure.
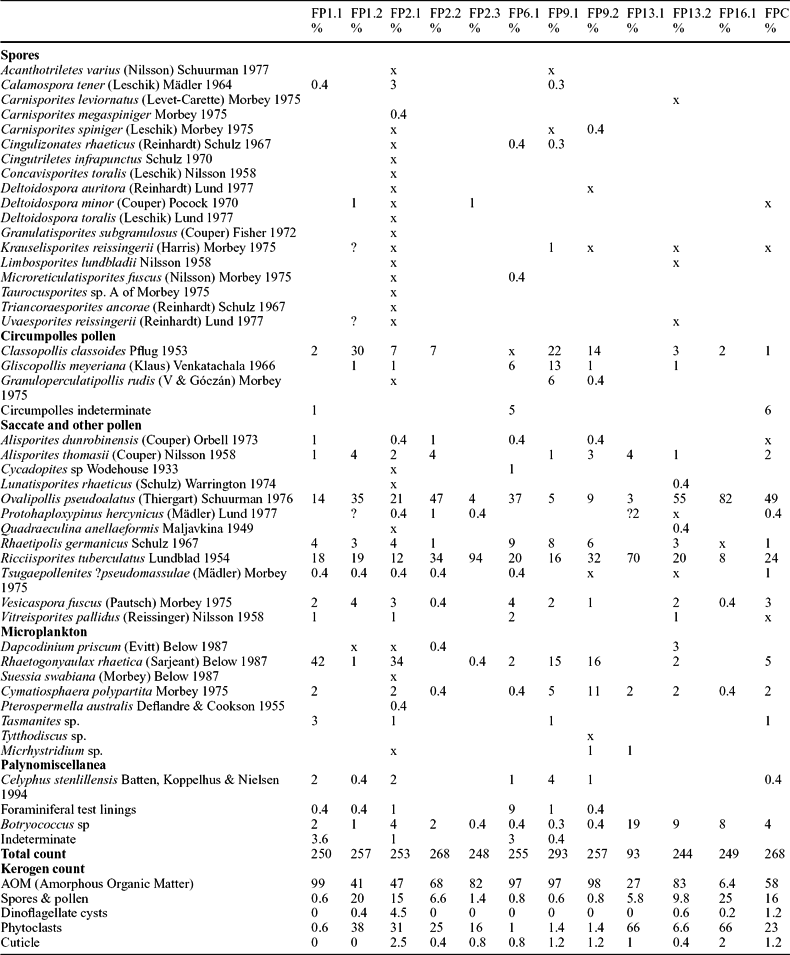
Table 2. Numbers and percentages of the palynomorph taxa recorded from the fissure fills of Tytherington Quarry and the locality near the Clifton Suspension Bridge, Bristol (FPC)
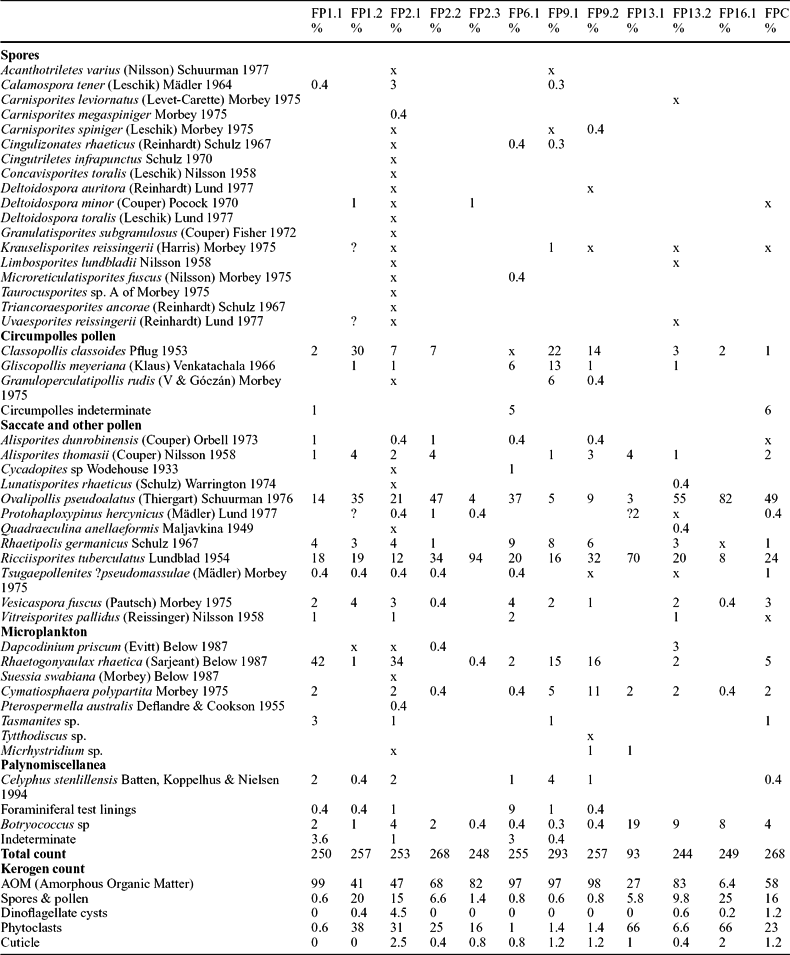
All numbers are % except where indicated as x when the species was out of count (250 palynomorphs). FP13.1 had a very sparse assemblage and <100 palynomorphs were present. Also included are the % kerogen counts for all the samples (500 points). Taxa that are cited but are not in the references can be found in Morbey (Reference Morbey1975), Lund (Reference Lund1977) or Achilles (Reference Achilles1981). The correct taxonomic assignment for both Corollina and Circulina is now Classopollis, the type species being C. classoides (Traverse, Reference Traverse2004; Skog, Reference Skog2005). Celyphus stenillensis is interpreted here as a foraminiferal test lining.

Figure 5. Vertebrates from the Tytherington fissures. All vertebrate material was prepared in 10% acetic acid following the method of Rixon (Reference Rixon1976). (a) Severnichthys. BRSUG specimen 23730. Blackened cusp of tooth from palynomorph-bearing rock; fissure 1, ×15, scale bar 500 μm. (b) Fossil charcoal. BRSUG specimen 23880. Fissure 1, ×20, scale bar 250 μm. (c, d) Clevosaurus. BRSUG specimen 23731. A worn tooth, (c) occlusal and (d) view of tooth base. Found in the same palynomorph-bearing rock specimen as BRSUG specimen 23730. Fissure 1, ×20, scale bar 250 μm. (e) Gyrolepis. BRSUG specimen 23735. Tooth, fissure 2, ×15, scale bar 500 μm. (f, g) Unknown chondrichthyan. BRSUG specimen 23729. Unidentified fossil, possibly a shark (hybodont?) denticle, fissure 2, ×15, scale bar 500 μm. (h) ‘Palaeosaurus.’ BRSUG specimen 23601. Tooth type, fissure 2, ×5, scale bar 1 mm. (i) Thecodontosaurus. BRSUG specimen 23600. Tooth, fissure 2, ×5, scale bar 1 mm. (j) Diphydontosaurus. BRSUG specimen 23725. Worn bone, showing corrosion, fissure 2, ×10, scale bar 500 μm. (k) Glauconitic clay-mineral BRSUG 23726. Fissure 2, ×20, scale bar 250 μm. (l) Ooids in aggregated cluster, BRSUG 23727. Fissure 2, ×10, scale bar 500 μm. (m) Section through aggregated ooid showing concentric calcite rings (arrowed) around a haematitic core, BRSUG 23736. Fissure 2, ×15, scale bar 500 μm. (n) Diphydontosaurus avonis. BRSUG specimen 23987. Part of left dentary, medial view, showing pleurodont teeth, fissure 2, ×10, scale bar 500 μm. (o) Diphydontosaurus avonis. BRSUG specimen 23988, Part of left dentary in lateral view, showing flaking of outer bone (arrowed) caused by sub-aerial exposure, fissure 2, ×10, scale bar 500 μm. (p) Diphydontosaurus avonis, BRSUG specimen 23989. Anterior part of right dentary in medial view, showing wear due to post mortem transport, fissure 2, ×10, scale bar 500 μm. (q) Planocephalosaurus robinsonae. BRSUG 23755. Part of right maxilla, lateral view, from palynomorph-bearing deposit, fissure 2, ×5, scale bar 1 mm. (r) Steinkern of ‘Rhaetic’ gastropod. BRSUG 23728. Fissure 2, ×20, scale bar 250 μm. (s) Diphydontosaurus avonis. BRSUG specimen 23990. Part of left maxilla, lateral view showing wear due to post-mortem transport, fissure 2, ×15, scale bar 500 μm. (t, u) Clevosaurus. BRSUG specimen 23700. Left dentary, (t) lateral and (u) medial view, fissure 2, ×2.5, scale bar 2 mm. (v) Unidentified Lepidosaur. BRSUG specimen 23737. Limb bone with metallic sulphide coating (arrowed), fissure 2, ×15, scale bar 500 μm. (w, x) Pholidophorus. BRSUG specimen 23738. White-coloured scale found in the rock that produced the holotype of Diphydontosaurus avonis, (w) internal and (x) external view, fissure 2, ×15, scale bar 500 μm. (y) Euestheria minuta. BRSUG specimen 23640. Fissure 4, ×5, scale bar 1 mm. (z) Collection of bones in a typical assemblage of white, yellow and brown bones (and bone fragments) from the rock that produced the type specimens of D. avonis, WB, white bones and YB, yellow bones of Diphydontosaurus. BRSUG specimen 23739. Fissure 2, ×5, scale bar 1 mm. (aa, bb) Lepidosaur ‘B’ of Whiteside (D. I. Whiteside, unpub. Ph.D. thesis, Univ. Bristol, 1984). BRSUG specimen 23670. Anterior of blackened left maxilla of sphenodontian related to Diphydontosaurus and Gephyrosaurus in (aa) lateral and (bb) medial view, fissure 12, ×10, scale bar 500 μm. (cc) Euestheria minuta var. brodeiana. BRSUG specimen 23641. Two individuals, fissure 12, ×5, scale bar 1 mm. (dd) Pelecymala sp. BRSUG specimen 23897. Part of left dentary in lateral view. The tooth-wear facets are caused by orthal shear rather than propalinal movement found in the similar Clevosaurus latidens Fraser, Reference Fraser, Lucas and Morales1993, fissure 14, ×3.5, scale bar 1 mm. (ee) Clevosaurus. BRSUG specimen 23890. Part of left dentary of a juvenile in lateral view, fissure 14, ×10, scale bar 500 μm. (ff) Planocephalosaurus. BRSUG specimen 23891. Scanning electron micrograph; the posterior view of the right premaxilla of a juvenile showing (?replacement) pits at the base of the teeth, fissure 14, ×25, scale bar 200 μm. (gg) Clevosaurus. BRSUG specimen 23691. Lateral view of toothed part of blackened right dentary, fissure 13, ×10, scale bar 500 μm. (hh) Pholidophorus. BRSUG specimen 23692. External view of brown coloured scale showing growth rings (arrowed), fissure 13, ×10, scale bar 500 μm. (ii, jj) Pholidophorus. BRSUG specimen 23690. (ii) External and (jj) internal view of brown coloured scale, fissure 13, ×15, scale bar 500 μm. (kk) Conodont. BRSUG 23732. Scanning electron micrograph of reworked Carboniferous conodont, probably Bisphathodus, fissure 14, ×25, scale bar 200 μm. (ll) Gyrolepis. BRSUG 23733. Scanning electron micrograph of white tooth found in the same rock with Planocephalosaurus, Tricuspisaurus and Pelecymala preserved in the same manner, fissure 14, ×25, scale bar 200 μm. (mm, nn) Diphydontosaurus. BRSUG specimen 23892. Part of blackened right dentary showing alternating sized-teeth, (mm) medial and (nn) lateral view, fissure 16, ×15, scale bar 500 μm. (oo, pp) Tricuspisaurus. BRSUG specimen 23893. Teeth, coloured grey, in (oo) lateral and (pp) occlusal view, fissure 14, ×10, scale bar 500 μm. (qq) Planocephalosaurus. BRSUG specimen 23894. Part of right maxilla brown coloured in lateral view, fissure 14, ×10, scale bar 500 μm. (rr) Planocephalosaurus. BRSUG specimen 23895. Left grey coloured dentary in lateral view, fissure 14, ×5, scale bar 1 mm. (ss) Part of rock hand specimen BRSUG 23896 showing possible associated whitish/grey-coloured Clevosaurus bones including a maxilla ‘M’ and a dentary ‘D’, fissure 16, ×4, scale bar 2 mm.

Figure 6. Palynomorphs recorded from the Tytherington fissures. Processing involved successive HCl and HF treatments followed by sieving at 20 μm and a further treatment in hot HCl to remove neoformed fluorides. No oxidation was required. The palynomorphs were mounted in Elvacite 2044 and the slides are curated in the collections of Bristol University Geology Museum. Each palynomorph name is followed by a palynological sample number (e.g. FP2.1), a BRSUG specimen number and England Finder co-ordinates (e.g. M29–3). In addition, all specimens are ringed. All ×600 unless indicated; appropriate scale bars are given. (a) Calamospora tener, FP1.1, BRSUG 23860, N45–1/3. (b) Concavisporites toralis, FP2.1, BRSUG 23861, N23. (c) Deltoidospora auritora, FP2.1, BRSUG 23861, H26–1. (d) Granulatisporites subgranulosus, FP2.1, BRSUG 23862, F41–1. (e) Carnisporites megaspiniger, FP2.1, BRSUG 23863, K22–1. (f) Carnisporites spiniger, FP2.1, BRSUG 23864, L26. (g) Microreticulatisporites fuscus, FP2.1, BRSUG 23865, U40. (h) Uvaesporites reissingerii, FP1.1, BRSUG 23860, M39–4. (i) Deltoidospora minor, FP2.1, BRSUG 23866, L43–4. (j) Taurocusporites sp A, FP2.1, BRSUG 23867, M29–3. (k) Acanthotriletes varius, FP2.1, BRSUG 23868, S36–3. (l) Cycadopites, FP2.1, BRSUG 23861, P15. (m) Cingutriletes infrapunctatus, FP2.1, BRSUG 23865, W36–1. (n) Cingulizonates rhaeticus, FP2.1, BRSUG 23862, T44. (o) Carnisporites leviornatus, FP13.2, BRSUG 23869, J36–4, ×400. (p) Limbosporites lundbladii, FP2.1, BRSUG 23864, L26–4. (q) Rhaetipollis germanicus, FP2.1, BRSUG 23861, N22–1. (r) Triancoraesporites ancorae, FP2.1, BRSUG 23868, Q43–3. (s) Krauselisporites reissingerii, FP2.1, BRSUG 23864, Q42–4. (t) Ricciisporites tuberculatus, FP2.1, BRSUG 23870, Q36, ×400. (u) Protohaploxypinus hercynicus, FP2.1, BRSUG 23868, M31–3. (v) Lunatisporites rhaeticus, FP2.1, BRSUG 23865, D36–3. (w) Tsugaepollenites ?pseudomassulae, FP2.1, BRSUG 23861, F26. (x) Ovalipollis pseudoalatus, FP2.1, BRSUG 23865, P38–4. (y) Vesicaspora fuscus, FP2.1, BRSUG 23865, Q36–4. (z) Alisporites thomasii, FP2.1, BRSUG 23861, P26. (aa) Alisporites dunrobinensis, FP2.1, BRSUG 23865, Q38–1. (bb) Granuloperculatipollis rudis, FP2.1, BRSUG 23868, J42. (cc) Quadraeculina anellaeformis, FP2.1, BRSUG 23871, K36. (dd) Classopollis classoides, FP2.1, BRSUG 23865, S34. (ee) Vitreisporites pallidus, FP2.1, BRSUG 23861, G14. (ff) Gliscopollis meyeriana, FP2.1, BRSUG 23872, O31–2. (gg) Suessia schwabiana, BRSUG 23873, N19. (hh) Rhaetogonyaulax rhaetica, FP2.1, BRSUG 23861, E29–3. (ii) Rhaetogonyaulax rhaetica, FP2.1, BRSUG 23866, L36. (jj) Micrhystridium sp, FP2.1, BRSUG 23861, Q18–1. (kk) Celyphus stenlillensis, FP6.1, BRSUG 23874, E42–4. (ll) Dapcodinium priscum, FP2.1, BRSUG 23871, K36–1. (mm) Pterospermella australis, FP2.1, BRSUG 23861, R23. (nn) Tytthodiscus, FP9.2, BRSUG 23875, N37, ×400. (oo) Cymatiosphaera polypartita, FP2.1, BRSUG 23870, L35–1, ×400. (pp) Botryococcus, FP2.1, BRSUG 23865, G39–1, ×400. (qq) foraminiferal test lining, FP2.1, BRSUG 23876, V45–3. (rr) foraminiferal test lining, FP2.1, BRSUG 23865, Q39.
Fissure 2 was the site of the original discovery of palynomorphs associated with terrestrial reptiles. In the first five exposures excavated by quarrying (Figs 8, 9, 0–14 m), the sediments yielded reworked Carboniferous fossils and numerous Triassic terrestrial reptile remains. They include a coelurosaur and the main Tytherington occurrence of Thecodontosaurus bones and teeth (Figs 5i, 7e–g) in a breccia (fig. 8a, b, d; at Th on Fig. 9, 0 m) which lay above a blackish-grey silty sandstone (Figs 7h, i; 8b, f, g; 9, 0 m) that produced the Rhaetian palynomorph assemblage FP2.1 (Fig. 6, Table 2) reported by Marshall & Whiteside (Reference Marshall and Whiteside1980). The presence of dolomite crystals, diagenetically altered from calcite, shown in the palynomorph-bearing rock FP2.1 (fig. 8g), indicate that dolomitizing fluids were active in the Rhaetian and are likely to have produced the dolomitized limestone later deposited as clasts in the Thecodontosaurus breccia. The fluids would also have contributed to the dolomitized matrix which constitutes part of the breccia.

Figure 7. (a) Fissure 1 showing convoluted dark-grey palynomorph-bearing (FP1.1) sandy mudstone of typical Westbury-type lithology ‘WL’ with fractured calcite band. The Carboniferous Limestone block ‘CLB’ has slipped and caused the disruption of the bed. (b) Fissure 1; the lower quarry scree has been removed (fig. 7a is outlined) and a more complete exposure below the slipped Carboniferous Limestone block of (a) is shown. There is dark-grey convoluted sandy mudstone to the left of the fractured calcite band (‘fc’ in fig. 7c) and a near-horizontal layered alternation of mudstone and conglomerate sequence on the right. The slumped sequence will have been related to earth movements and was formed after deposition, which is clearly demonstrated by the broken calcite and mineral vein. Such seismic movements have been identified by Mayall (Reference Mayall1983) at the beginning of the Cotham Member, and Simms (Reference Simms2006) regards the cause as bolide impact. The palynomorph assemblage FP1.2 (Table 2) was taken from the right-hand side and the sample point is shown in close-up in Figure 7c. The scale-figure (DIW) is 1.7 m tall. (c) Fissure 1. A close-up of the sequence described in (b); there are repeating conglomeratic bands that fine upwards (graded bedding) and have fine laminated mudstones between them. Note location of FP1.2 and ‘fc’ the fractured calcite vein. (d) Fissure 1. A close-up of (c) that shows two conglomeratic bands (a clast is circled). (e) Thecodontosaurus. BRSUG specimen 23613. Lateral view of left ilium showing characteristic short anterior process ‘AP’. The bone is whitish coloured which is typical of many of the Thecodontosaurus bones of the original discovery at Tytherington. Fissure 2, ×0.75, scale bar 2 cm. (f) Thecodontosaurus. BRSUG specimen 23613. Medial view of same left ilium as (e) with dark-grey silty sandstone ‘GSS’ matrix attached. This matrix is lithologically the same as the rock abutting the Thecodontosaurus bones that produced palynomorph assemblage FP2.1. Fissure 2, ×0.75, scale bar 2 cm. (g) Thecodontosaurus. BRSUG specimen 23642. A postero-medial view of part of a white and yellow coloured right femur showing well-developed trochanter ‘tr’ with muscle scar ‘ms’. Fissure 2, ×0.75, scale bar 2 cm. (h) Acid-etched rock showing brown- and white-coloured bones of Thecodontosaurus ‘Th’ in a breccia above finely laminated cross-bedded calcareous silty sandstone, including a layer of the grey lithology that elsewhere, and below the Thecodontosaurus breccia, yielded the palynomorph assemblage FP2.1. Fissure 2, ×0.6, scale bar 2 cm. (i) Opposite side of (h) showing angular white dolomitized Carboniferous limestone ‘DCL’ breccia with clasts above, and cutting into, finely laminated grey silty sandstone. Fissure 2, ×0.6, scale bar 2 cm.

Figure 8. (a) Fissure 2, first exposure (mid-November, 1975) showing the locality of the original find of the breccia and conglomerates (outlined as ‘b’) that yielded Thecodontosaurus, Diphydontosaurus and other reptiles. Palynomorph-bearing siltstone laminae occur below this original find with a similar but non-laminated lithology in the same horizon as the conglomerate. (b) Fissure 2, close-up of the Thecodontosaurus breccia ‘Th’ and conglomerate band in situ. The location of the limestone that yielded the holotype of Diphydontosaurus is marked ‘D’, as is the palynomorph-bearing rock FP2.1, the original assemblage described by Marshall & Whiteside (Reference Marshall and Whiteside1980). (c) Later exposure of fissure 2 (mid-December, 1975), about 17 m to the southeast of the first exposure, showing fissure formation on a vertical joint ‘VJ’, with an upper fissure fill ‘UFF’ lying above ‘keyhole’ fissure fill. The two sedimentary fills were undoubtedly continuous and the intermediate section was removed as a result of quarrying. (d) Two blocks of Thecodontosaurus breccia and conglomerate from fissure 2 showing bones (circled) and yellow dolomitized limestone. (e) Rock specimen from fissure 2 showing bands of red, that is, a ‘Keuper’ type lithology that yielded Planocephalosaurus ‘Pl’ with bands of green calcareous sediment, plus thin laminae of black silt that contained the Rhaetic palynomorph assemblage FP2.2, ×1.1, scale bar 2 cm. (f, g) Thin-section of rock that yielded palynomorph assemblage FP2.1; (g) shows crystals of baroque dolomite ‘d’ that are diagenetically replaced calcite. This dolomite implies dolomitization in Penarth Group times which may have produced dolomitized Carboniferous Limestone that later was deposited as clasts in the breccia layer that lay directly above (see fig. 8b, d). Organic matter ‘om’ is present as thin orange-coloured bands which contain AOM and represent a short-lived algal bloom that formed as a discrete event. The fine silty layers are characterized by larger phytoclasts and represent terrestrial input. The palynomorphs are discrete entities and not introduced as clasts by reworking or slumps. The R. tuberculatus specimen shown in (g) is circled, ×50, scale bar 300 μm. (f) Close up of thin-section (g) showing in situ specimen of Ricciisporites tuberculatus as circled in Figure 8b; (compare Fig. 12 t), ×150, scale bar 50 μm.
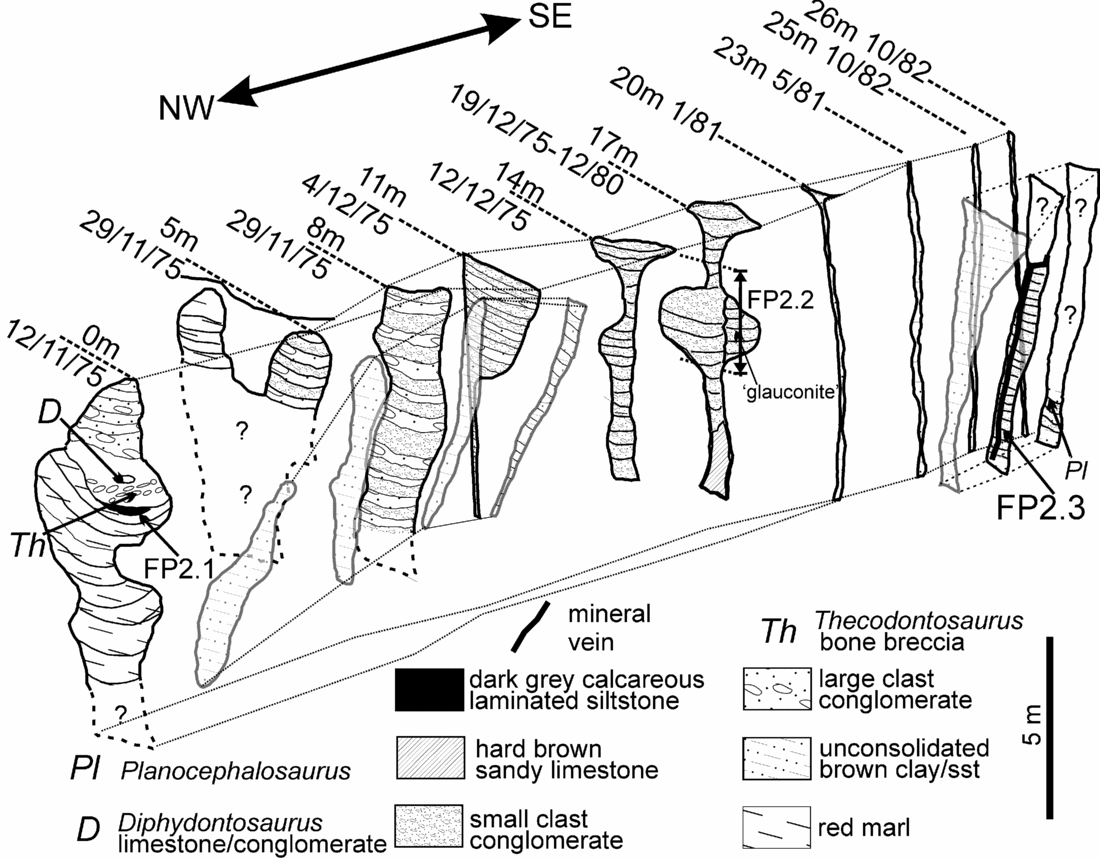
Figure 9. Fissure 2 showing the morphological evolution of fissure 2 as it was quarried from NW to SE. Overall there is a change from large clast breccias and conglomerates at 0 m to smaller clast reddish conglomerates in the middle section and to grey-black conglomerates and siltstones in the last sections at 23–26 m. The main fissure is accompanied by a brown sand and mud-filled satellite fissure development which can be followed from 0 m to the section at 11 m. The first (discovery) exposure of the fissure at 0 m is a complex fill with thick laminated red sandy marls succeeded by grey laminated Westbury-type sediments that yielded palynomorph assemblage FP2.1. This sequence continued upwards with fining-up breccia and conglomeratic sediments that included Westbury-type grey sandy mudstones; this is the rock that yielded the original find of Thecodontosaurus at Tytherington. At 5 m, the exposure shows a similar fill (where exposed) with the upper part of the fill separated into two. This separation is lost in the next exposure (8 m), which is a repeating sequence of conglomerates and finer reddish sediments. Some of the conglomerates, which are identical to ‘dolomitic conglomerate’, have large rounded clasts of sandstone and limestone including dolomitized limestone. At 11 m the lower part of the fissure has become simply a vertical joint but the upper part is much wider and a continuation of the section at 8 m. The upper infilling is also similar to that at 8 m with alternating beds of large and small clast conglomerates. The large clast conglomerates then disappear by 14 m where the fill is solely composed of red small-clast conglomerates. At 14 m and 17 m there is the complex development of a short phreatic tube. The section at 17 m has been interpreted by some (e.g. Simms, Reference Simms1990) as a phreatic tube with a vadose notch cut into the base. It is more likely to have been formed by tectonic and solutional activity along a vertical joint entirely below the water table. The very localized circular structure would have been in direct receipt of surface waters that swirled around at the freshwater lens/saline water boundary to create the feature (see Fig. 16 and Whiteside & Robinson, Reference Whiteside and Robinson1983). A comparison with the previous cross-section at 14 m demonstrates that the right hand side of the ‘tube’ at 17 m was produced later than the left hand side and is strong evidence for void formation during the Penarth Group. The void was then filled by ‘glauconitic clay’-rich conglomerates on the right hand side of the ‘tube’ at 17 m. The fissure rapidly closes to an enlarged joint by 20 m and remains so until the final section at 26 m. In parallel with this closure, the satellite fissure has opened. The satellite fissure seen at 23 m is likely to be an extension of fissure 1 and implies that there was an inter-connectivity of the two systems. The locations of the three fissure 2 samples are shown. FP2.2 can only be positioned approximately between 17 m to 18 m. These series of sections were compiled from in situ collection and measurements, photos and field notes made between successive quarry blasts. The top of the section is hung on the top of the third quarried level which is shown as the ‘horizontal’ lines on the second exposure at 5 m (29/11/75). The base of the fissure remains unknown, but as these fissures were not located during the quarrying of the fourth level it may lie in the upper part of this deepest level (at approximately 60 m OD; about 44 m below the Carboniferous Limestone surface).

Figure 10. (a) Fissure 3, first exposure showing strongly developed ochreous mineralization with fine-grained limestone ‘FGL’ to bottom right. Although barren of palynomorphs this limestone is similar to the palyniferous fine-grained limestone of fissure 16 that produced FP16.1. (b) Fissure 3, second exposure about 5 m behind (a) with a vertical notch ‘VN’ cut into its base and with ochreous layers filling the cavity. (c) Fissure 11 labelled to show the three exposures of this fissure. Scale figure is 1.75 m tall. The extent of the red sandy breccia ‘RSB’ that contained the fossils is arrowed. (d) Fissure 14 showing the extent of the funnel shape opening ‘o’ of a doline. The vertical extent of the main fossil-bearing ‘FB’ green calcareous sandy rock is arrowed. (e) Fissure 14, this is a close-up of the lower section (marked in fig. 10d) and is filled with red cross-bedded sandy material that contained a few black lepidosaur bones. There are green reduction spots in this deposit and these are frequently associated with the bones. Note the smooth limestone wall on the left of the red sediments that is due to solution. (f) Fissure 12, the layers included the cross-bedded sandstone with water drop marks ‘WDM’ and thin red sandstones. Some white bone fragments were found in situ in these rocks. However, the main fossil-bearing rock is above this horizon, and although the fossils collected were not found in situ, their lithologies were recognized using binoculars and therefore their position can be accurately located. Sample BLB (position estimated from observation through binoculars) contained black lepidosaur bones; WLB, white lepidosaur bones and Eu, Euestheria (estimated position where the lithology is uniquely identical to the Euestheria-bearing rock collected from the scree below).
Counting the number of recognizable tibias, we have recorded a minimum number of eleven individuals of Thecodontosaurus from one breccia horizon. Other reptile remains included the tooth fossil ‘Palaeosaurus’ (fig. 5h), possible suchian bones, Clevosaurus, and the holotype of Diphydontosaurus avonis (at D on Fig. 9, 0 m), which was found in the same matrix as scales of the osteichthyan fish Pholidophorus (fig. 5w, x). The reptile bones are generally unblackened and frequently white, particularly the Thecodontosaurus fossils (Figs 7e, f, 5n–p), but a few sphenodontian bones found in conglomerates of the original collection of rocks, such as a palatine of Planocephalosaurus, are black or dark brown with a sheen of metallic sulphide. The limb bone of a lepidosaur from these conglomerates (fig. 5v) has the metallic sulphide coating. The conglomerates included specimens of Clevosaurus (fig. 5t, u), together with an unblackened tooth of the palaeoniscid Gyrolepis albertii (fig. 5e), a ctenacanthoid scale and some internal casts identified as Penarth Group gastropods.
In a later exposure of fissure 2 (Fig. 9 at 17 m and Whiteside & Robinson, Reference Whiteside and Robinson1983), the southern side of the circular cross-section contained a conglomeratic rock with a green glauconitic clay found in a variety of morphologies (including pellets, fig. 5k), together with the unblackened bones and teeth of Planocephalosaurus, Clevosaurus and Thecodontosaurus. A well-preserved shark denticle, possibly from an unknown hybodontoid (fig. 5f, g), and internal casts referable to Penarth Group gastropods (fig. 5r), were also found in this conglomerate. In the same exposure of fissure 2, but not collected in situ, is a mainly reddish coloured rock (fig. 8e), with subsidiary green areas, that contains well-defined fine bands of black sandy limestone. These black laminae have yielded palynomorphs including rare dinocysts plus abundant amorphous organic matter (AOM; sample FP2.2, Table 2), while the reddish coloured rock has yielded terrestrial sphenodontians including Planocephalosaurus, and at least one Gyrolepis tooth. This rock is significant, as the thin ‘organic-rich’ layers of ‘Westbury-type’ lithology are within a predominantly terrestrial ‘Keuper-type’ lithology.
Further quarrying of fissure 2 (Fig. 9, 23–26 m) revealed 7 m of black upward-fining conglomerates containing clasts of grey weathered limestone and brown and olive green clay above a 0.3 m bed of finely laminated unfossiliferous brown, muddy sandstones. In this section the sequence was cut by a vertical development of hydrothermal mineral veins (mainly galena and barytes) and was overlain by a horizontal vein of these minerals. The black conglomerate yielded palynomorphs (sample FP2.3), a fragment of an unblackened mandible of Diphydontosaurus avonis, a black Clevosaurus tooth and a few fragmentary Rhaetic fish remains. Further excavation of this conglomerate has produced a well-preserved right maxilla of Planocephalosaurus (fig. 5q; at Pl on Fig. 9, 23 m).
Fissure 8 produced a small fauna of Clevosaurus and Diphydontosaurus, consisting of whitish bones, from a clastic limestone, whereas fissure 10 yielded the mainly black bones of Clevosaurus, Planocephalosaurus and an archosaur tooth (probably Thecodontosaurus). Clasts of black clay were found in the limestone matrix of fissure 10.
The infill of fissure 11 (fig. 10c) consists mainly of bands of red marls interbedded with a sequence of fining-upward breccias containing a fauna of Diphydontosaurus avonis, Clevosaurus, another probable although unidentifiable sphenodontian, a Pholidophorus scale, and the skull bones of a terrestrial crocodile referable to both the Terrestrisuchus described from Pant-y-ffynon by Crush (Reference Crush1984) and another from Cromhall Quarry held in the British Museum of Natural History. Other skeletal elements such as parts of a femur found in this fissure are probably from this crocodile.
In fissure 12 (fig. 10f), but not collected in situ, a bed of thinly laminated pale cream mudstones yielded the branchiopod crustacean Euestheria minuta var. brodeiana (fig. 5cc). Among other black fragmentary skeletal elements and two osteichthyan fish teeth, one of which is probably Severnichthys, a green breccia yielded the anterior part of a left maxilla (fig. 5aa, bb) of lepidosaur ‘B’ with strong affinities to Diphydontosaurus avonis and Gephyrosaurus bridensis Evans, Reference Evans1980.
The Carboniferous Limestone walls of fissure 13 have pronounced horizontal grooves and large ‘cusps’ produced by solution. There is a keyhole-shaped out-pocket (fig. 11a, KSP) 4 m to the east of the main infill of limonite-rich limestones and conglomerates, comprising lower, mainly thick, beds and upper thinner beds. The limestones contain voids, some of which are partly filled by sparry calcite and others with finely laminated limestone (fig. 11c) that resemble the caymanite described by Jones (Reference Jones1992a,b) from the Cayman islands. Caymanite is a dolostone but originated as a limestone (Jones, Reference Jones1992a), and it is this original fabric that is preserved in fissure 13. The lower beds contain palynomorphs (FP13.1, Table 2) in thin broken bands of finely laminated pale grey limestones and also an assemblage (FP13.2, Table 2) in one black silty limestone clast. Both the palynomorph-bearing rocks were found between beds containing the terrestrial reptiles Clevosaurus (fig. 5gg), Thecodontosaurus, a smaller unidentified archosaur (possibly Terrestrisuchus) and the poorly preserved bones and numerous scales of fish (fig. 5hh–jj, Pholidophorus; confirmed by C. Patterson, pers. comm.). Although some fish scales and bones are found associated, and a few bones are white, much of the abundant vertebrate material is poorly preserved, very fragile, and grey, brown or black.
The lower, red sediments of fissure 14 (fig. 10e) have yielded one in situ blackened lepidosaur bone. Above this red rock, the lithology changes to poorly consolidated yellow sands with green patches and hard green sandy conglomeratic limestones. This conglomeratic limestone yielded skeletal elements of Clevosaurus (fig. 5ee), including one reworked tooth surrounded by a matrix similar to the yellow infills of fissures 13 and 16, and specimens of, probably, Agnosphitys Fraser et al. Reference Fraser, Padian, Walkden and Davis2002. It also produced many well-preserved bones of Planocephalosaurus (fig. 5ff, qq, rr), Tricuspisaurus Robinson,1957b (fig. 5oo, pp), Pelecymala sp. Fraser, Reference Fraser1986b (fig. 5dd) and Sigmala sp. Fraser, Reference Fraser1986b, the latter two only previously known from Cromhall. One very white Gyrolepis tooth (fig. 5ll) and a significant quantity of fossil wood are found in the same matrix as this diverse reptile fauna, together with numerous reworked Carboniferous conodonts (fig. 5kk), Carboniferous fish teeth, and crinoid ossicles. A Planocephalosaurus-dominated fauna, although from a much smaller sample, was also found within a green limestone lithology in fissure 4.
The highly fossiliferous deposit of fissure 16 consists of a basal palynomorph-bearing (FP16.1) pale white and grey finely laminated limestone bordered and overlain by firstly unconsolidated, limonite-rich soft yellow sandy limestones and then by harder conglomerates. The conglomerates contain abundant, mainly black or brownish-black, bones and teeth of Clevosaurus, Diphydontosaurus avonis (fig. 5mm, nn) and reworked ‘bradyodont’ Carboniferous fish. Skull bones, possibly associated, comprising two dentaries, one maxilla and other elements, of a single Clevosaurus, were found (fig. 5ss), which is reminiscent of deposits in Cromhall Quarry where complete articulated skeletal material is recorded by Fraser (Reference Fraser1988a).
4. Dating the Tytherington fissure infills and their faunas
4.a. Dating based on the tetrapods
Comparing the Tytherington reptiles with those from the British Triassic and Jurassic strata yields some scant evidence about the age of the Tytherington reptiles.
A specimen of the tooth fossil ‘Palaeosaurus’ is recorded from the Ceratodus bone bed (Lower Westbury) at Aust (Davis, Reference Davis1881), although that specimen is larger than the Tytherington tooth found in fissure 2 (fig. 5h). Moore (Reference Moore1881) reported ‘Rhaetic’ ‘Palaeosaurus’ and Thecodontosaurus teeth from Holwell associated with Penarth Group fish. However, in the same paper Moore stated that Thecodontosaurus teeth collected from the North Curry Sandstone (Carnian) at Ruishton, near Taunton, were the most similar to the Durdham Down Thecodontosaurus antiquus that we regard as conspecific with the Tytherington prosauropod. Dating based on a few teeth alone is dubious, so Moore's evidence is tentative at best. The lepidosaur specimens collected by Moore at Holwell have affinities to Diphydontosaurus, but no Holwell specimens are certainly ascribable to any of the sphenodontian genera from Tytherington. Therefore the reptiles at Tytherington cannot be unequivocally dated by comparing them with fossils from British sequences of known age.
4.b. Dating suggested by the fish and invertebrate fauna
The marine fish fauna found in the same matrix as the tetrapods in fissures 1, 2, 6, 9, 12 and 14 (Table 3) comprised fragmentary remains of either Gyrolepis albertii and Severnichthys or both. Although Gyrolepis is known from the North Curry Sandstone Member (Carnian) of the Mercia Mudstone Group at Ruishton (C. J. Duffin, unpub. Ph.D. thesis, Univ. London, 1980), these fish are typically found in the Westbury Formation of the Bristol area (e.g. Richardson, Reference Richardson1911; and see Swift & Martill, Reference Swift and Martill1999).
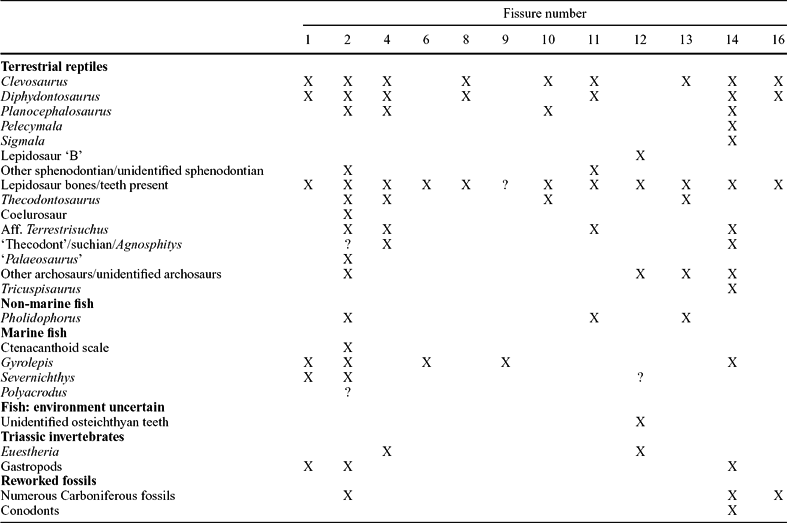
Table 3. Distribution of terrestrial reptiles, marine and non-marine fish, invertebrates and reworked Carboniferous fauna in the Tytherington fissures
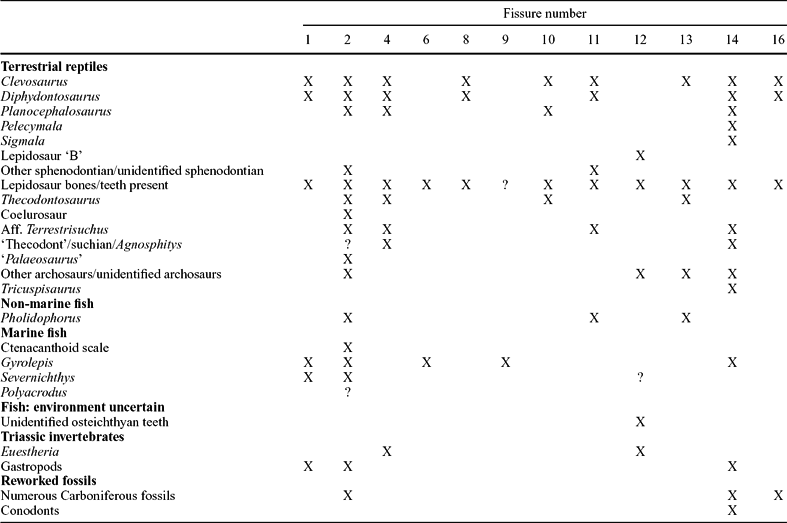
X – confirmed presence; ? – unconfirmed presence; Aff. – affinity
Fissure 13 is unusual in possessing bones and numerous scales of the non-marine fish, Pholidophorus, preserved in the same manner as bones of Clevosaurus, Thecodontosaurus and other reptiles. The scales, although larger, resemble closely those of P. higginsi known from the Cotham Member of the Lilstock Formation. However, Pholidophorus is also known from the ‘paper shales’ (Westbury) of Leicestershire (e.g. Harrison, Reference Harrison1876), so the evidence supports a general Penarth group age assignment rather than a more specific stratum level. Pholidophorus is also found in the same matrix with reptiles in fissures 2 (Whiteside, Reference Whiteside1986) and 11.
Gastropod casts found in fissures 1, 2 and 14 (fig. 5r) appear identical in form and preservation to specimens from the Westbury Formation at Patchway, Bristol (Bristol City Museum and Art Gallery, registration number CC479) and in the substantial collection from bed 9 at Hampstead Farm Quarry made by M. Curtis (pers. obs.).
Fissure 12 has yielded Euestheria, ascribable to the dwarf form E. m. brodeiana and lepidosaur B, which has affinities to Diphydontosaurus avonis and to the younger Gephyrosaurus bridensis from the Welsh Rhaeto-Liassic fissures. Lepidosaur B was found in a breccia that appears to be above the Euestheria bed, but this is uncertain, as neither have been found in situ, and the face was inaccessible. E. m. brodeiana occurs, typically, in rocks of the Cotham Member, Lilstock Formation, but has been found in some pre-Rhaetian beds by Warrington (Reference Warrington1963).
The single Euestheria minuta specimen of fissure 4 (fig. 5y), found in the same matrix as unidentifiable reptiles, provides some debatable evidence that sediments were deposited before the Westbury Formation. As far as we are aware, no specimen of a similar size to the E. minuta of fissure 4 has been found in any British Penarth Group sequence, but it does fall within the size range of specimens from the Arden Sandstone Member (Carnian) and equivalents in Worcestershire, Somerset and Warwickshire. This evidence is equivocal, as the living equivalents of Euestheria fluctuate in size with salinity, the largest specimens being found in fresh water (Jones, Reference Jones1862), and their final size is determined by temperature and food supply with low temperature and food shortages retarding the growth of the carapace (Olempska, Reference Olempska2004). It may be simply that the E. minuta in fissure 4 was inhabiting a temporary warm, freshwater pool environment with an abundant food supply in the otherwise generally brackish water conditions persisting on the Rhaetian near-shore limestone surface.
The age of some sediment in fissure 4 is debatable but the evidence from the fish and invertebrate fossils in the deposits of fissures 1,2, 6, 9, 11, 12 and 14 clearly points towards a stratigraphical level equivalent to the Penarth Group, especially the Westbury Formation, although it is possible that the fauna of fissure 12 may be equivalent to the younger Cotham Member of the Lilstock Formation.
4.c. Dating based on palynomorphs
4.c.1. The palynomorph assemblages
Following the discovery of palynomorphs in fissure 2, further samples were routinely and regularly collected from all the fissures as they were quarried. All samples were palynologically processed by standard techniques and then reprocessed at different times, in different laboratories and in different batches but gave the same results, hence ruling out the possibility of laboratory contamination. Samples were also collected at 0.1 m intervals from the Penarth Group in Hampstead Farm Quarry, Chipping Sodbury (ST 726839; locality details are given in Donovan, Curtis & Curtis, 1989), to provide comparative Penarth Group palynological assemblages in the vicinity of Tytherington. In addition to the Tytherington deposits, we sampled other fissure systems for palynomorphs and recorded a new assemblage from a fissure close to the Clifton Suspension Bridge, Clifton, Bristol (ST 566733). This fissure had been previously reported by Moore (Reference Moore1881) and relocated by Copp (pers. comm.), both of whom ascribed it to the Rhaetic (see also Benton et al. Reference Benton, Juul, Storrs and Galton2000). In common with Warrington (Reference Warrington1977), Fraser & Walkden (Reference Fraser and Walkden1983) and Walkden & Fraser (Reference Fraser, Lucas and Morales1993), no palynomorphs were found in the fissures at Cromhall.
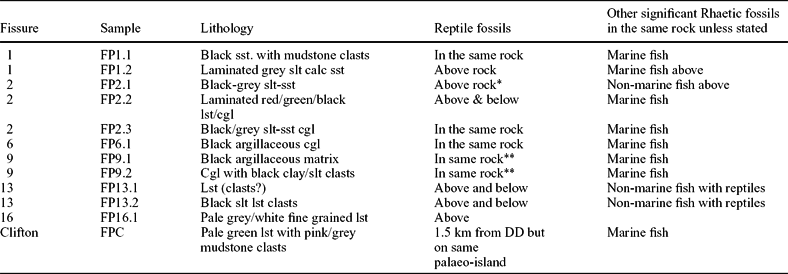
Palynomorph assemblages have been discovered in fissures 1, 2, 6, 9, 13 and 16 (Table 2). Table 4 summarizes both their lithologies and their relationship to the vertebrate fossils. In fissures 1, 2, 6 and 9, the palynomorphs were, with the exception of the black and red laminated rock from fissure 2 (fig. 8e), all found in black calcareous siltstones. This lithology frequently contains significant quantities of larger grains within the matrix, ranging from sand-sized to conglomeratic. It is, therefore, a heterolithic lithology representing an environment with variable mixed inputs of organic-rich mudstones and much coarser material resulting from localized cave collapse. There is a further assemblage from a black rock in fissure 13 (FP13.2), but the other palynomorph-bearing rocks FP13.1 and FP16.1 are distinctive fine-grained, pale coloured (white to grey) limestones.
Table 4. Lithology and faunal associations of the palynomorph-bearing rocks

The palynological assemblages are very clearly present above, below and co-bedded with the reptile fossils.
*Same lithology as palynomorph bearing rock is found in the bone horizon.
**Reptile fossils are in the same rock but it is unknown whether matrix or clasts or both.
calc – calcareous; cgl – conglomerate; lst – limestone; slt – siltstone/silty; sst, sandstone; DD, Durdham Down
The palynomorph-bearing rock FP2.1 was examined in a polished block and a thin-section including observation in incident UV light. Figure 8g shows a laminated sediment comprising dolomitized carbonate layers, silt layers with palynomorphs and phytoclasts and discrete AOM-rich layers. The content of phytoclasts including charcoal can be significant and individual palynomorphs can be recognized within these silt layers (e.g. the Ricciisporites tuberculatus in fig. 8f, g), proving that the palynomorphs are present as discrete sedimentary particles. Palynomorph preservation was excellent in the black siltstones but poor in the limestones where many of the microfossils had degraded and thinned walls, giving them a ‘ghost’ appearance. The assemblage from fissure 2 appears more diverse (Table 2) because this sample has been studied in the most detail. Apart from FP13.1, all samples contained enough organic residue for a count of 250 palynomorphs, with most samples containing several thousand individual palynomorphs. In common with other similar assemblages, Ricciisporites tuberculatus tends to disintegrate but gives very distinctive fragments of sculptured surface wall or the equally distinctive internal core of the specimen (e.g. pl. 2, fig. 8 of Orbell, Reference Orbell1973). All fragments of R. tuberculatus were counted so the relative abundance of this palynomorph is an overestimate. In addition, a 500 point kerogen count (as area coverage) was made for all the slides using a Swift point counter. Results are also shown on Table 2.
4.c.2. General age assignment based on the palynological assemblages
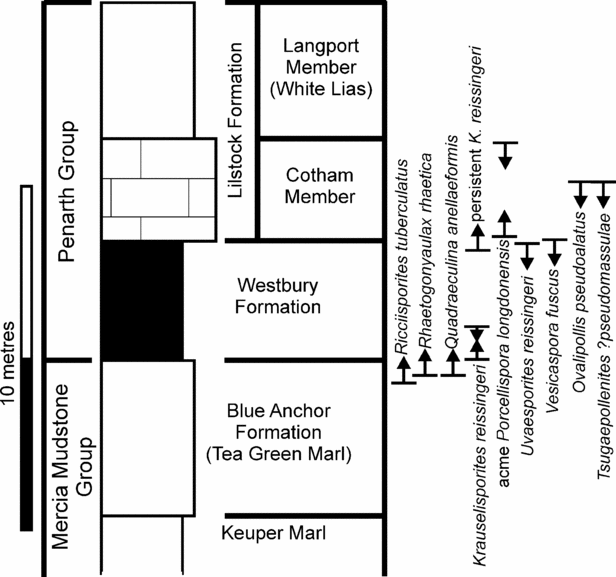
Taking the fissure assemblages as a group, the miospores, pollen and organic walled microplankton from these samples (Fig. 6, Table 2) are unequivocally characteristic of the Penarth Group and well known from the Rhaetian Stage. They contain an assemblage dominated by Ovalipollis pseudoalatus, Ricciisporites tuberculatus and Rhaetogonyaulax rhaetica, together with persistent Rhaetipollis germanicus, Classopollis classoides, Vesicaspora fuscus and Cymatiosphaera polypartita. This Rhaetian microflora is well documented, having been described from many sections in Western Europe (as reviewed in Warrington, Reference Warrington, Jansonius and McGregor1996), including the UK (e.g. Orbell, Reference Orbell1973; Warrington et al. Reference Warrington, Ivimey-Cook, Edwards and Whittaker1995; Warrington in Edwards, Reference Edwards1999; Hounslow, Posen & Warrington, 2004). Significant sources used for comparison and palynomorph identification include Morbey (Reference Morbey1975), Schuurman (Reference Schuurman1977, Reference Schuurman1979) and Lund (Reference Lund1977).
The age can be constrained more specifically (in terms of British lithostratigraphy, Fig. 12) through comparison with known palynomorph ranges (Orbell, Reference Orbell1973; Hounslow, Posen & Warrington, 2004, plus sources in Fig. 1). Using these inceptions, acme and extinctions, the earliest possible age must be equivalent to the Williton Member (very uppermost Blue Anchor Formation or younger), based on the first occurrences of R. tuberculatus (which occurs in abundance in all the fissure palynomorph assemblages), Quadraeculina annellaeformis and Krauselisporites reissingerii. The age is further constrained as older than the Langport Member from the presence of O. pseudoalatus, Tsugaepollenites ?pseudomassulae and V. fuscus. The presence of Uvaesporites reissiingerii (in fissures 2 and 9) is also significant in that it has not been found above the Westbury Formation. A further restriction can be placed using the abundances of selected palynomorphs. K. reissingerii is very rare within the fissure assemblages but common in the Cotham Member where it has been used to define the Heliosporites Zone of Orbell (Reference Orbell1973). Porcellispora is also similarly common in Cotham and younger strata but absent from the fissures. Therefore it is clear that there is a general age equivalence to the Westbury Formation.
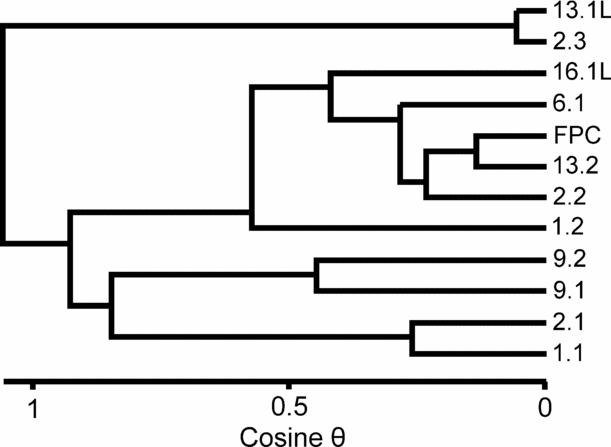
A more sophisticated analysis of the Tytherington palynological assemblage data (Table 2) has revealed some quite distinct differences in assemblage composition, coupled with the near-absence of microplankton from some samples. Significantly, this difference in assemblage composition can occur within a single fissure (contrast FP2.1 with FP2.3). Qualitatively these differences are seen in FP1.2, 2.2, 6.1, 13.2 and 16.1, where the assemblages are dominated by Ovalipollis with a lesser abundance of Ricciisporites, plus a locally high but very variable content of Circumpolles pollen (Classopollis, Gliscopollis and Granulatoperculatipollis in the fissures). In addition, the relative abundance of marine microplankton is very low or entirely absent. In contrast, FP2.3 and 13.1 have very high abundances of Ricciisporites with subordinate Ovalipollis, Circumpolles pollen is absent and marine plankton are very rare. The third group (FP1.1, 2.1, 9.1 and 9.2) has a more diverse palynological assemblage that is not dominated by a single pollen component (that is, Ovalipollis, Ricciisporites or the Circumpolles group) and contains a substantial component of organic walled marine microplankton (Rhaetogonyaulax rhaetica with Cymatiosphaera polypartita). These differences can be expressed quantitatively (Fig. 13) using cluster analysis. These results show a clear parallel to the three groups determined by visual inspection of the palynomorph count data. Note the very clear separation of the group comprising FP2.3 and 13.1. Although FP1.2 clusters with the Ovalipollis-dominated samples, it is close to the third group that is characterized by a diverse palynological assemblage. Some analytical combinations will, in fact, cause it to switch to this group. However, the similarity is probably driven by their shared relative abundance of Circumpolles pollen rather than the more diagnostic abundance of marine microplankton.
4.d. Age comparisons with the Hampstead Farm Quarry palynological profile
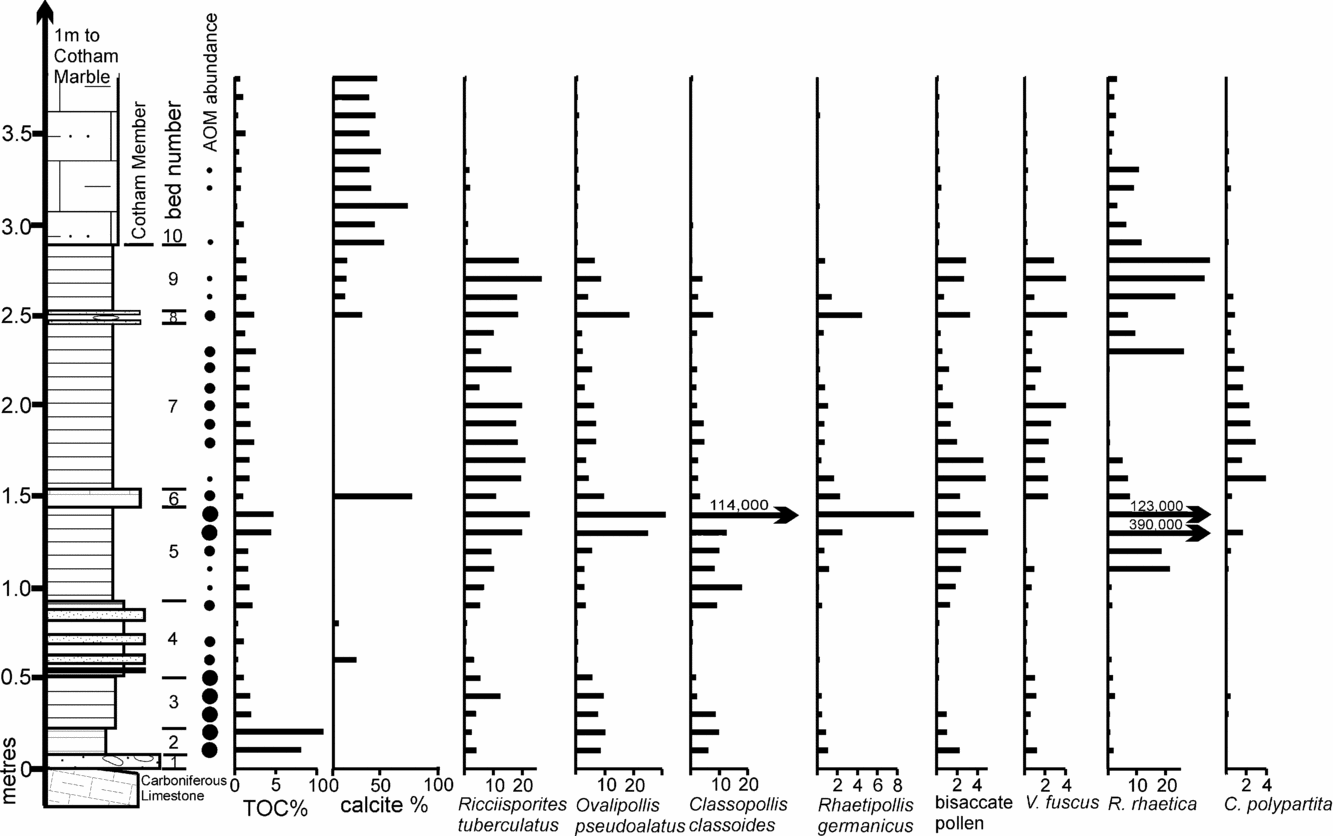
In order to secure a more direct understanding of how these Tytherington assemblages correlate within the Westbury Formation of the Penarth Group, a comparative quantitative palynological profile was produced from a normal bedded sequence at Hampstead Farm Quarry, Chipping Sodbury. This profile (Fig. 14) includes the Westbury and the lower part (Cotham Member) of the Lilstock Formation. This location is only 8 km from Tytherington and located on the same Carboniferous Limestone massif. Hence, like Tytherington, it is similarly ‘on-structure’, in contrast to the available palynological profiles that are generally more basinal (Fig. 1, e.g. Lavernock), and therefore it has been subjected to the same sequence of Late Triassic transgressions and regressions. This profile has also been constructed quantitatively to avoid the whole sum problem that arises inevitably when discussing such data. The Hampstead Farm Quarry section also emphasizes the composite nature of the Westbury Formation which contains a number of internal cycles. These are variable in number at different sites. For example, three were recognized from Filton, Bristol (Hamilton, Reference Hamilton1962), two from Chilcompton (Duffin, Reference Duffin1980), four at Hampstead Farm Quarry (Fig. 14) and six in South Wales (Warrington & Ivimey-Cook, Reference Warrington, Ivimey-Cook and Taylor1995). The cycles are defined by thin intervals of limestone or sandstone that often occur above minor erosion surfaces and represent times when the sea bed became current-swept and water column stratification broke down. As such, they could represent times when the system became tidally influenced (Allison & Wright, Reference Allison and Wright2005). The top of bed 9 at Hampstead Farm Quarry contains a significant vertebrate and diverse invertebrate marine fauna including echinoids and a single psiloceratid ammonite (Donovan, Curtis & Curtis, 1989) and clearly represents an interval where open marine conditions were prevalent and therefore a likely hiatus in deposition.
The results from Hampstead Farm Quarry can also be used to correlate with the more basinal bedded sequence at St Audrie's Bay, Somerset, where palynological work on that section (Warrington, Reference Warrington, Neale and Brasier1981) and correlatives (Warrington et al. Reference Warrington, Ivimey-Cook, Edwards and Whittaker1995) shows the same distinctive double peak of R. rhaetica at the top and base of the Westbury Formation. Bed 7 at Hampstead Farm Quarry lies between these abundances of R. rhaetica, and the palynological assemblages are relatively dominated by terrestrial spores and pollen, presumably because the dinoflagellates are not encysting. However, this part of bed 7 has an elevated total organic carbon (TOC) and a significant abundance of AOM. The source of this AOM is assumed to be from non-cysting dinoflagellates since elsewhere in the Penarth Group, biomarker studies (Thomas et al. Reference Thomas, Marshall, Mann, Summons and Maxwell1993) of AOM show that, where it occurs in high abundances, there are coincident peaks of dinosterol, a dinoflagellate specific biomarker, reflecting the presence of dinoflagellates. When compared to the St Audrie's Bay section there is a TOC high between these two R. rhaetica acme (Hesselbo, Robinson & Surlyk, 2004) that is coincident with a relative abundance of terrestrial sourced palynomorphs. This means that bed 7 from Hampstead Farm Quarry correlates with beds 10–12 of Hesselbo, Robinson & Surlyk (2004), with bed 6 (Hampstead Farm Quarry) possibly being equivalent to bed 13 and the influx of clastic sediments of bed 16 being equivalent to bed 4.
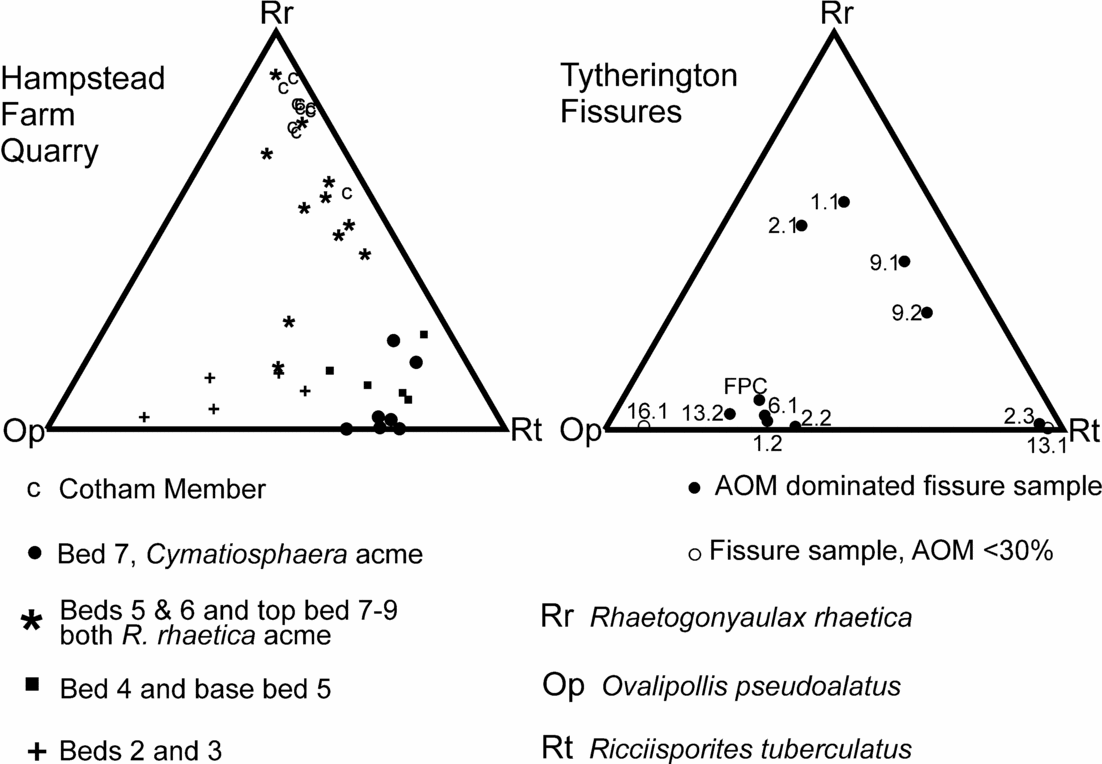
The assemblages from Hampstead Farm Quarry have also provided a quantitative sequence of palynological events that can be compared to the out-of-sequence samples from the Tytherington fissures. The three major palynological elements (R. rhaetica, R. tuberculatus and O. pseudoalatus) from both localities were recalculated and plotted on a pair of ternary diagrams (Fig. 15). The diagram for Hampstead Farm Quarry shows how the relative abundance of these three elements can be used to discriminate the groups of different beds.
4.d.1. Fissure 1
Sample FP1.1 contains abundant O. pseudoalatus with R. tuberculatus and persistent R. germanicus and V. fuscus, together with a significant percentage of R. rhaetica. The Circumpolles group is less abundant, as are specimens of C. polypartita. When contrasted with the Hampstead Farm Quarry section (Fig. 15), the abundance of R. rhaetica but rarity of C. polypartita indicates a correlation with upper bed 5 to lower bed 7 or upper bed 7 to 9. However, the paucity of the Circumpolles group suggests that a correlation within the upper part of the Westbury Formation (upper bed 7 to 9) is more likely. This is supported by the abundance of AOM in the sample. Although not age diagnostic, it demonstrates either stratification within the water column or dysoxia/anoxia within the top layer of sediments. However, when compared to the possible correlative levels in the Hampstead Farm Quarry section, AOM is only abundant within the middle part of bed 7, therefore further constraining the correlation to this interval.
The FP1.2 assemblage is quite different in being dominated by O. pseudoalatus, C. classoides with R. tuberculatus and an almost complete absence of marine palynomorphs. Figure 15 shows that the sample plots in the area of beds 2 and 3 from Hampstead Farm Quarry. Both these levels are also rich in AOM and have high TOCs, which support the correlation. These very high TOCs in laminated sediments implied stagnant bottom conditions with excellent preservation of organic matter. Previous studies of Westbury Formation palynofacies (Tuweni & Tyson, Reference Tuweni and Tyson1994) suggest that variations in the AOM quality are caused by preservational differences in this largely dysoxic facies.
The location of both samples is shown on Figures 3 and 7a–c. The relative age of the samples within a bedded shale pocket in fissure 1 demonstrates that this is an in situ normal bedded sequence of Westbury Formation. The reptile bones occur within the same matrix and are bracketed by the palynological dates. Figure 7d shows the meniscus fill to the fissure. This meniscus diminishes up-section and is therefore compaction related. It also shows the layers of locally sourced conglomerate. These contain concentrations of vertebrate bones and are interpreted as marking regressions. As such, they probably relate to the internal pattern of cycles known from the Westbury Formation.
4.d.2. Fissure 2
FP2.1 contains the same assemblage as that found in FP1.1 (Figs 13, 15) and similarly can be attributed to the upper part (upper bed 7 to bed 9) of the Westbury Formation.
The FP2.2 assemblage is similar to FP1.2 in being dominated by Ovalipollis with Ricciisporites and Classopollis but without any significant marine component. AOM is also abundant. Therefore a position in the lower part of the Westbury (beds 2 and 3) is most likely.
FP2.3 is characterized by the third type of assemblage, that is, one dominated almost entirely by Ricciisporites (94%) with only a minor content of Ovalipollis and very rare marine phytoplankton. However, the AOM content is high. There is no obvious equivalent to this type of assemblage in the bedded Westbury Formation from Hampstead Farm Quarry. Figure 15 does show some assemblages that are apparently very rich in Ricciisporites but these are from bed 7 and are also characterized by a high abundance of Cymatiosphaera which is absent in FP2.3. The closest match is with bed 4 and the base of bed 5, where the section becomes very impoverished and probably regressive as shown by the influx of sandstone horizons. The other possibility is that this assemblage does not relate directly with assemblage compositions in the local Westbury Formation. Spore-dominated assemblages are known from the British Rhaetic such as those from marginal sediments at The Cross, Cowbridge, South Wales (Fig. 1; Orbell, Reference Orbell1973). They are also known from further north in the UK (Benfield & Warrington, Reference Benfield and Warrington1988; Warrington in Frost, Reference Frost1998; Warrington, Reference Warrington2005), towards the probable margins of the marine-influenced Penarth Group (Warrington & Ivimey-Cook, Reference Warrington, Ivimey-Cook, Cope, Ingham and Rawson1992). Although generally dominated by Ovalipollis, the palynological assemblages from Northallerton (Winton Manor Borehole) do have a persistent abundance of over 40% Ricciisporites in the Westbury Formation. There is also a Ricciisporites-dominated assemblage (>60%) in the Westbury Formation at Chilcompton, Somerset (Warrington, Reference Warrington1984), but this sample is from the top of the formation, contains Cymatiosphaera sp., and is equivalent to bed 7 at Hampstead Farm Quarry. Therefore, on palynological evidence it would appear that FP2.3 is probably very locally sourced from plants growing in the immediate vicinity and it is not an assemblage that has been formed by palynomorph mixing during transport. Given that the characteristic compositional changes in palynological assemblage through the Westbury Formation are geographically widespread and reflected in the other fissure samples, then FP2.3 probably pre-dates this influence. However, it cannot correlate with any interval older than the uppermost Williton Member (Blue Anchor Formation), since R. rhaetica is restricted to the top of this 2 m thick interval. In addition, Ricciisporites has a first occurrence at its base. Therefore this assemblage, despite these differences, cannot be cited as evidence for a much earlier Norian phase of fissure deposits.
This (provisional) attribution of FP2.3 to the lower (but not lowest) part of the Westbury Formation is in accord with its position in the fissure 2 system (Fig. 9 at 17–18 m). FP2.3 was recovered from a side branch to the main fissure (possibly an extension of fissure 1) and is below both FP2.2 and FP2.1, which are respectively from the lower and upper parts of the Westbury Formation. These two more secure dates also again demonstrate an internal stratigraphy to the fissure infill.
4.d.3. Fissure 6
This contains one productive palynological sample (FP6.1) that is characterized by a high abundance of Ovalipollis with Ricciisporites and Circumpolles pollen, with only a minor component of marine microplankton. However, the marine influence is present, as demonstrated by a very high AOM content and the presence of a significant percentage (10%) of foraminiferal test linings. Therefore this assemblage can be correlated with beds 2 and 3 from the lower part of the Westbury Formation.
4.d.4. Fissure 9
This fissure contains two palynological assemblages. One (FP9.1) is from a black argillaceous sandstone, while FP9.2 is from black clay clasts contained within this matrix. These black clay clasts contain a fairly typical Penarth Group assemblage of R. tuberculatus, O. pseudoalatus and R. germanicus with a somewhat increased relative abundance of Circumpolles pollen. However, what is significant are the nearly equal abundances of both R. rhaetica and C. polypartita. These show a very striking distribution in both the Hampstead Farm Quarry section and in fact, the Penarth Group generally, with the characteristic double spike of R. rhaetica in the upper Westbury separated by an interval where C. polypartita is the dominant element in the phytoplankton. The near-equal relative abundance of the two elements suggests a position at either the base or top of the C. polypartita peak, that is, base or top of bed 7. The relative paucity of bisaccate pollen in fissure 9 suggests that the upper position in bed 7 is perhaps more likely.
The matrix to the clasts (FP9.1) differs in having a predominance of R. rhaetica, albeit still with an elevated content of C. polypartita. Given that this sample is the matrix to the FP9.1 clast, it would appear to be from the upper part of bed 7 or bed 8 and hence near-contemporaneous. This position is supported by the elevated content of Circumpolles pollen in contrast to the palynological assemblage from the clasts.
4.d.5. Fissure 13
FP13.1 is a laminated limestone sample that has a palynological assemblage which is dominated by Ricciisporites (70%) with only a very minor content of Ovalipollis, while the marine component is represented only by rare specimens of Cymatiosphaera. The palynological content of this sample was also very sparse in containing just under 100 individual palynomorphs. Hence, the assemblage is both distinctive and different from all the others at Tytherington. It also differs in palynofacies through having a low content of AOM coupled with a relative abundance of phytoclasts and particularly long (>500 μm) opaque laths of inertinite, that is, charcoal. The other significant difference in this FP13.1 assemblage is the abundance (19%) of Botryococcus. This alga is only sporadic in most other Tytherington samples. Botryococcus is an alga that is typical of quiet, clear fresh water (Batten & Grenfell, Reference Batten, Grenfell, Jansonius and McGregor1996). As such, it can thrive in temporary ponds and its presence in the fissures confirms that the Tytherington island was large enough to support such freshwater ponds. The lithology of the sediment is also fairly unusual in comprising clasts of finely laminated grey limestone. Hence, given the very high abundance of probably locally sourced Ricciisporites (recalculated at 87% without the Botryococcus) and the low AOM but high inertinite content, it seems most likely that these are reworked ‘pond’ sediments that perhaps accumulated in a temporary karst depression prior to its collapse. Although FP2.3 has a similar Ricciisporites-dominated palynomorph assemblage, it differs markedly in lithology and palynofacies. Hence there is probably little that can be deduced about the age of FP13.1 except by association with FP13.2. The abundance and large size of the charcoal fragments (e.g. Ohlson & Tryterud, Reference Ohlson and Tryterud2000) does indicate the presence of local wildfires in the island vegetation.
FP13.2 is a black calcareous siltstone clast. It contains a more typical assemblage that is dominated by Ovalipollis with a lesser content of Ricciisporites with the marine plankton represented by low abundances of Rhaetogonyaulax and Cymatiosphaera. Hence, its assemblage is similar to that of FP1.2, 2.2 and 6.1 (Figs 13, 15), with which it shares a high AOM content, and it can therefore also be correlated with beds 2 and 3 from Hampstead Farm Quarry, the only difference being the presence of Krauselisporites reissingerii and a somewhat elevated abundance (9%) of Botryococcus. Although K. reissingerii is regarded as characteristic of the uppermost Westbury through to the pre-planorbis beds of the Blue Lias, it does have an isolated occurrence in the basal Westbury Formation (Fig. 12 and Hounslow, Posen & Warrington, 2004) and similarly also occurs at Hampstead Farm Quarry in two consecutive samples from the upper part of bed 3. This is regarded as confirming the correlation to Hampstead Farm Quarry. The Botryococcus is more problematic but could be locally sourced (cf. FP13.1).
4.d.6. Fissure 16
FP16.1 contains an assemblage that like FP13.1 is in a fine-grained light-coloured limestone. The assemblage has a very high content of Ovalipollis (89%) with subsidiary Ricciisporites and only rare Cymatiosphaera specimens as the sole marine microplankton. The palynofacies contains very little AOM and there is a relatively high abundance of Botryococcus (8%). Hence this sample can be distinguished from the more typical AOM-dominated Ovalipollis plus Ricciisporites assemblage (FP1.2, 2.2 and 6.1), as shown by its separation from the main group in Figure 15. Again this may indicate an environment somewhat different from the Westbury Formation and perhaps, like FP13.1, a shallow temporary lime-rich pond containing colonies of Botryococcus, but different in that it was surrounded by a vegetation dominated by the, as yet still unknown, Ovalipollis-producing plant.
5. Overall summary of the dating of the infills and their fauna
Our dating of the fissure sediments permits, for the first time, a definitive age estimate for the terrestrial reptiles of the sauropsid fissures. Marshall & Whiteside (Reference Marshall and Whiteside1980) gave an ‘age’ assignment of Westbury or possibly lower Cotham Beds for the palynomorph-bearing dark grey laminated calcareous sandy siltstone from fissure 2, now called FP2.1, associated with rocks containing Thecodontosaurus antiquus and Clevosaurus. Those reptiIes, together with Diphydontosaurus avonis and a coelurosaur have been found above FP2.1 (fig. 8b). Clevosaurus and Diphydontosaurus avonis are present in the same matrix as the palynomorph assemblage FP1.1 of fissure 1. Planocephalosaurus robinsonae Fraser, Reference Fraser1982 has been found in the palynomorph-bearing rocks (FP2.2 and FP2.3) of fissure 2. In fissure 13, Clevosaurus and Thecodontosaurus are found surrounding FP13.2. All dates from these palynomorph assemblages give a Rhaetian age assignment at the stratigraphical level equivalent to beds 2 to 3, 4 to 5 or top bed 7 to 9 of the Westbury Formation at Hampstead Farm Quarry.
This Westbury Formation assignment is in accord with the marine fish fossils found in fissures 1, 2, 6, 9, 12 and 14. The Pholidophorus remains found in the sediments of fissures 2, 11 and 13 and gastropods of fissures 1, 2 and 14 are also consistent with a correlation to the Westbury Formation.
The tetrapods are found in the matrix, not reworked and not in discrete clasts within the palynomorph and Penarth Group fish-bearing rocks. Since the palynomorphs in most of the assemblages are preserved in excellent condition (Fig. 6) and were not introduced in discrete reworked clasts (fig. 8g), except for sample FP9.2 from fissure 9 and the ‘Westbury clast’ FP13.2 of fissure 13, we conclude that the Westbury Formation assignment ascribed to the palynomorph-bearing rocks also applies to the reptiles Thecodontosaurus antiquus, Clevosaurus sp., Planocephalosaurus robinsonae and Diphydontosaurus avonis. Therefore, although there are examples of crumpled, collapsed and fallen rocks, we refute the assertion of Walkden & Fraser (Reference Fraser, Lucas and Morales1993) that the Tytherington deposits give a misleading date because of disturbance in some deposits.
No definitive fauna from normal bedded Triassic rocks has been found associated with the tetrapods in fissures 4, 8 and 10, but Clevosaurus is known from fissures 4, 8 and 10, Planocephalosaurus from fissures 4 and10 and Diphydontosaurus avonis from fissures 4 and 8. Based on these faunal associations, we conclude that these infills are also of Rhaetian age. Hence, all identifiable vertebrate faunas (excepting Carboniferous remanié fossils) from the Tytherington fissures can be surely assigned to the Westbury Formation of the Penarth Group. It is possible that the full stratigraphical range of these vertebrates may be pre- or post-Penarth Group, but no indisputible evidence has been found that would support such a speculation. The considerable diversity of the herpetofauna and the great number of small lepidosaur individuals would suggest a productive habitat in a more humid climate than the semi-aridity proposed by Robinson (Reference Robinson1957a) and Halstead & Nicoll (Reference Halstead and Nicoll1971). Based on clay analysis, Mayall (Reference Mayall1979) suggested that such relatively humid conditions would be seen in the Rhaetian times rather than in the immediately preceding stage. Sufficient supplies of freshwater to maintain the ecosystem would be positioned near to the current limestone surface, at the top of a freshwater lens supported by the saline waters of the high sea level of the Westbury Formation (see Fig. 16).
6. Formation of the Tytherington fissures
The Tytherington fissures trend along E–W or NW–SE vertical joint planes. The direction of these fissures is within that recorded for the Ladinian–Rhaetian fill-type 1 (E–W and NNW–SSE) and Rhaetian–early Hettangian fill-type 2.1 (NNW–SSE) of the Mendip Hills by Wall & Jenkyns (Reference Wall and Jenkyns2004). In the case of fissures 1 and 10, the parallel walls suggest that the main cause of fissure widening was probably tectonic although later modified by water, since solutional effects can be observed on the enclosing Carboniferous Limestone wall. Similar minor solution conditions are reported in the Q-fissures of Wendt (Reference Wendt1971), which are submarine gapes formed by tectonic movements. The exposure of fissure 2 shown in Figure 8b also has evidence of some tectonic activity (widening) on a vertical joint.
In addition to the tectonic generation of the voids, the cavern formation in fissure 2 may well be produced, in part, by acidified waters resulting from the action of sulphate-reducing bacteria at the halocline. The presence of these bacteria is indicated by the sulphides that encrust some of the bones found in fissure 2 (see fig. 5v). Simms & Ruffell (Reference Simms and Ruffell1990) argued that because the water catchment area of the Tytherington aquifer is small (about 2 km2), the rainfall needed to produce the ‘conduits’ such as the fissure 2 cavern must have been high. However, it is the juxtaposed saline/fresh water which is more likely to be responsible for the circular voids at Tytherington such as that figured by Whiteside & Robinson (Reference Whiteside and Robinson1983). The development of fissure 2 is considered in more detail in Figure 9.
Some of the fissures such as 5 and 8 are to a major degree solution-widened joints, but the primarily solutional fissures that provide evidence of the age of formation are now considered.
Fissure 13 (fig. 11a–e) presents an interesting case. Although formed along an E–W-trending vertical joint plane, the general morphology of the fissure is of a cavernous nature with the convex walls indicating the widened tube that forms, typically, in phreatic conditions (Sweeting, Reference Sweeting1972). The walls bear horizontal grooves, upon which are superimposed large, symmetrical, solutional cusp markings (see fig. 11b). Comparison with modern caves from Sarawak described by Wilford (Reference Wilford1964) indicates that such horizontal grooves are indicative of erosion by slow-moving streams. The ‘cusps’ are believed not to have formed by strong (vadose) currents since they occur, for example, in blind passages. This evidence, therefore, strongly suggests that this fissure was formed in mainly phreatic conditions; later, slight falls in water level caused a slow-moving stream to erode levels on the limestone walls and form the vadose notch of the keyhole-shaped pocket (KSP) of Figure 11a until finally, the conditions became totally phreatic again. Therefore, this fissure formed under phreatic conditions within a few metres of the present-day limestone surface.

Figure 11. (a) Fissure 13 looking towards the SW. The top ‘entrance’ to the fissure is indicated ‘o’. The fissure widens from this opening. At the extreme left is a keyhole-shaped pocket ‘KSP’ and at the centre are horizontal solution grooves with cusps (marked ‘b’). At the lower right is the main fossiliferous deposit ‘d’ that formed part of a cavernous structure. The whole structure is interpreted as a flank margin cave. (b) The main group of solution cusps. (c) Rock specimen from fissure 13 that shows cavity filled with ‘caymanite’; note the characteristic layering; ×0.85, scale bar 2 cm. (d) Main fossiliferous deposit with Richard Bennett collecting in situ reptile bones. Pholidophorus scales and bones were found in the same horizon. The bones collected at this position were very corroded in a halo of green sediment within the calcareous yellow limonite clay sandstone. Note thin layers of hard sandy limestone above his head. To the right of the deposit, the lower limestone bands thicken and sag ‘S’ downwards. (e) Main fossiliferous deposit, looking SE with DIW (1.70 m tall) indicating the site of palynomorph-bearing limestone (FP13.1). Note the harder, thin sandy-limestone bands ‘SL’ just above the centre of the picture.
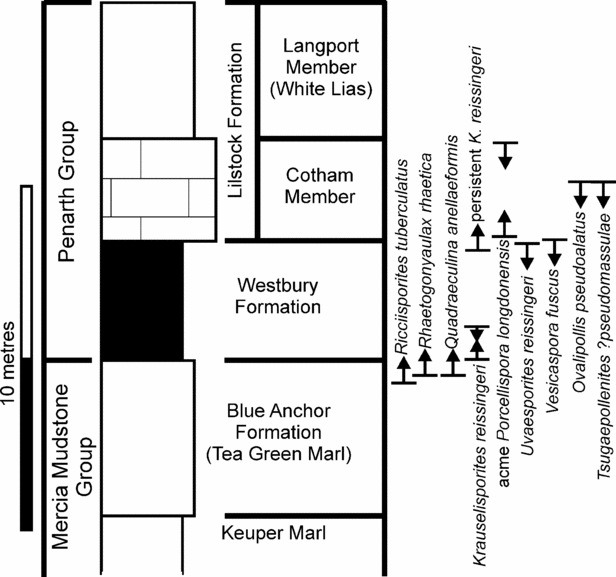
Figure 12. A representative stratigraphical section of the Penarth Group with selected palynological markers (inceptions and extinctions). Section based on Bristol district (Kellaway & Welch, Reference Kellaway and Welch1993) with palynological markers after Hounslow, Posen & Warrington (2004); Warrington & Whittaker (Reference Warrington and Whittaker1984) and Orbell (Reference Orbell1973). Note that the characteristic Penarth Group palynomorphs have inceptions in the very uppermost part of the Blue Anchor Formation.
The distribution of Triassic sediments over the top of the Carboniferous Limestone (Fig. 4) shows that the present-day land surface is essentially an exhumed Triassic topography (e.g. Robinson, Reference Robinson1957a). This topography and present-day sea levels give a water table some 67 m OD, which is about 30 m below the present altitude of the fissure 13 system, and the formation of a phreatic cave at this altitude requires a water table raised above that of the present day. The most likely way to produce this is by having a sea level higher than that of today. The first interval during which such elevated sea occurred following pre-Permian inversion and erosion was during the Rhaetian transgression. The position of the nearest ‘Rhaetic’ deposits is less than 1 km to the south (Fig. 4) and at a relative height of 6 m below the limestone surface that overlies fissure 13. This shows that marine waters would have been lapping up against the Tytherington area at this time. It is difficult to envisage this cave forming at an earlier time (contrast Simms, Ruffell & Johnston, 1994), as the British Triassic rocks were generally deposited under arid conditions and even in pluvial episodes the water table would have been too low. It is likely that marine waters intruded directly into this cavern during its formation. The cavern of fissure 13 is envisaged as an example of a large dissolution void, termed a ‘flank margin cave’, that was produced in a mixing zone, similar to those described in the Bahamas by Mylroie & Carew (Reference Mylroie, Carew, Budd, Saller and Harris1995), who reported that such voids can be produced in less than 15 ka. It is the mixing of saline and fresh waters that results in high rates of limestone dissolution. Choquette & James (Reference Choquette, James, James and Choquette1988), Smart, Dawans & Whitaker (Reference Smart, Dawans and Whitaker1988) and Palmer & Williams (Reference Palmer and Williams1984) have described such high dissolution conditions for caves under Andros Island, Bahamas.
Fissures 12, 14, 16 and to an extent fissure 11 were produced by solution and subsidence processes resulting in dolines, the cavities of which are now infilled with sediment. The depth of penetration for the doline fissures 11, 12, 14 and 16 is not certainly known, but no true doline fissure is seen below 86 m OD, suggesting that level for the water table during their formation. This indicates a high sea level at their time of formation such as that of the Westbury transgression. Fissure 16 (Fig. 3) shows an expansion at its base of 94 m OD. This would be expected where a subaerial pipe met the water table. The formation of dolines does not take a very long time. Working in Istria, Morawetz (Reference Morawetz1965) concluded that, based on a modern climate regime, a doline about 15 m deep and with a diameter of 50 m would have taken no longer than 14 ka to form. The Tytherington dolines are only about 2 m across, suggesting that there would have been plenty of time for their formation in any of the minor regressive cycles in the Westbury Formation. The doline fissures are very similar in size and shape to the ‘banana holes’ of Caribbean islands that are partly formed by decaying vegetation acting as an acidic sponge dissolving the limestone. The optimum conditions for the formation of modern dolines are massive limestones, with substantial vegetation cover and a horizontal surface or low angles of slope (Sweeting, Reference Sweeting1972), just the situation that would be present on the Tytherington Rhaetian island. The infill of fissure 16 has a sparse but clearly Rhaetian palynomorph assemblage. It is therefore deduced that the formation and infilling of fissure 16 were penecontemporaneous events. This is in strong accord with the findings of Wall & Jenkyns (Reference Wall and Jenkyns2004) for Mendip Triassic fissures, where the age of the fill-type is essentially the time of the fissure formation.
The solutional fissures could not have been mainly formed during any of the more arid periods of the Permo-Triassic, since this type of karst does not form in deserts (Jakucs, Reference Jakucs1977). It is noteworthy that in the Bristol and South Wales region no primarily solutional fissures are found underlying basal Westbury Formation rocks. Karst development is found in Barnhill Quarry (Chipping Sodbury, ST 725826) where the Rhaetic cover has been stripped off, and there, the limestone directly covered by the Rhaetic beds ‘is quite unmarked by solution channels’ (Reynolds, Reference Reynolds1938). The occurrence of a Penarth Group deposit overlying and continuous with a fissure infill at Cromhall does not contradict this, as the presence of ‘pholidophoriform’ fish in one of the Cromhall slot fissures (Walkden & Fraser, Reference Fraser, Lucas and Morales1993) and Pholidophorus in the capping sequence suggest a time equivalent to the Cotham Member (Whiteside, Reference Whiteside1986) or within the Westbury Formation, rather than basal Westbury.
There is equivocal evidence, albeit scant, that part of the 1, 2, 4 and 10 fissure systems and fissure 3 were formed in pre-Rhaetian times. The evidence of the large E. minuta specimen of fissure 4 may indicate that some infilling occurred in pre-Rhaetian times, and therefore partial fissure formation must have preceded that date. In fissure 3, the notch cut into the base of the circular cross-section (contrast fig. 10b with 10a) characteristic of vadose conditions can be interpreted as strengthening this suggestion. If vadose conditions persisted at the 64 m OD level of this fissure, then it can only be presumed that the formation must have taken place in very early (at the start of the transgression) or pre-Rhaetian times when the water table would have been at a sufficiently low level. The youngest probable pluvial episode (which would therefore account for this speleogenesis) prior to the Rhaetian, is equivalent to the North Curry Sandstone (Carnian: Warrington & Williams, Reference Warrington and Williams1984). This is the principal evidence of pre-Rhaetian solutional formation of fissures at Tytherington, but this interpretation requires hypothesizing that the notch is vadose and that water tables would not have been sufficiently low during a regressive Rhaetian phase (such as the low sea stands in the later Rhaetian postulated by Hesselbo, Robinson & Surlyk, 2004) or at the start of the major transgressive phase. ‘Keyhole’ cross-sections with apparently vadose notches are seen in Sarawak caves (Wilford, Reference Wilford1964, pl. VI), where the entire formation is believed to be phreatic. A significant pre-Rhaetian solutional formation of reptile-bearing fissures is therefore speculative and has no direct evidence. It is also possible that hydrothermal waters contributed to the formation of voids, a phenomenon described by Ford (Reference Ford, James and Choquette1988), and that these voids were infilled by fossiliferous sediments. However, much of the mineral veining in Tytherington seems to be associated with organic-rich sediments from the Westbury Formation and the dissolutional effects of hydrothermal waters may have been most significant during this interval. The significance of hydrothermal activity is considered below.
The Tytherington fissure system has a complex genealogy that includes tectonic and solutional effects and sometimes a mixture of both. It is suggested here, though, that most of the reptile-bearing fissures were formed mainly in the Rhaetian by solution processes along and down joints. With the rising sea level, all the fissures would at some time be present on the edge of a freshwater lens where the rate of solution is relatively great. This solution rate is caused by enhanced water velocity (Chidley & Lloyd, Reference Chidley and Lloyd1977) and undersaturation of aragonite and calcite in the mixing zone of fresh and saline waters resulting from bacterially derived H2SO4 that lowers the pH (Smart, Dawans & Whitaker, Reference Smart, Dawans and Whitaker1988).
6.a. Significance of the hydrothermal mineral veins
It is significant that fissures 1, 2, 9 and 10 have metal sulphide veins associated with the fossil-bearing and usually black sediments. In the case of fissure 9, black clay was found in nodules of metallic sulphide minerals. Although the mineral veins could have formed later (for example, post-Mid-Jurassic as suggested for Southfields Quarry by Curtis, Reference Curtis1981), they equally may have formed as a direct result of sulphides produced a short time after deposition or penecontemporaneously with the vertebrate fossils in organic-rich sediments. The sulphides would have been produced by sulphate-reducing bacteria using the organic material (mainly fatty acids) as a food source and the sulphate as a source of oxygen. It can be hypothesized that the blackened vertebrate fossils were derived from an anoxic environment where sulphate-reducing bacteria produced H2S that reacted with metals such as Fe, Zn and Pb in the hydrothermal groundwaters, hence causing the blackening. The black lepidosaur bones in fissures 2, 12, 13, 14 and 16 would have derived from these anoxic environments. Following the model for these fissures of Whiteside & Robinson (Reference Whiteside and Robinson1983), the anoxic environment would have been present just below the halocline and the waters above and below would be clearer and oxygen-rich (see Fig. 16). Conditions such as this occur in the inland blue holes of Andros Island, Bahamas, for example, in Stargate Hole (D. Birch, unpub. data, 2006: http://www.smallhope.com/CTSD/InlandBlueHoles.html).
7. Palaeoenvironment of the Tytherington fissure infilling
7.a. Fissure 2
The infilling of fissure 2 contains conglomerates in which the contemporaneous terrestrial to marine vertebrate fossils are in a ratio of >1000:1. There are also sequences of black argillaceous sandstone and conglomerates in the same fissure which contain the same terrestrial vertebrate components at less than 10% of the overall number of mainly marine vertebrate elements. This admixture of fauna is strongly indicative of deposition in a littoral zone or a marginal marine location. Marshall & Whiteside (Reference Marshall and Whiteside1980) originally suggested this type of palaeoenvironment for fissure 2 based on terrestrial tetrapods associated with a mixed terrestrial/marine Rhaetian palynomorph assemblage. Whiteside & Robinson (Reference Whiteside and Robinson1983) considered the known palaeogeography in the Rhaetian (Audley-Charles, Reference Audley-Charles1970) and expanded this model, postulating that fissure 2 was present in the fluctuating freshwater/saline water zone which could be found between the freshwater lens and saline water mass on small limestone islands. Their finding in fissure 2 of a conglomerate containing Clevosaurus, Planocephalosaurus and Thecodontosaurus with pellets and fine layers of glauconitic clay is commensurate with such a model.
We have accumulated further evidence of the marine influence. In the conglomerate that yielded the glauconitic clay, an unblackened shark denticle and numerous steinkerns of probable Penarth Group gastropods have been found (fig. 5r). The reptile-bearing conglomerates that comprised the bulk of the fissure infill yielded a Gyrolepis albertii tooth and ctenacanthid scale. Later exposures of the black argillaceous sandy conglomerates sequence (see Fig. 9, 25–26 m), where the clasts included orange and orange-brown clays, have also contained fragmentary fossils of Rhaetic fish. It now seems certain, therefore, that the main sedimentary mass of the fissure 2 infilling was deposited in a mixed freshwater/marine environment, confirmed by the evidence from the additional palynomorph assemblages FP2.2 and FP2.3.
The main deposit of Thecodontosaurus bones (fissure 2) was contained in a breccia and conglomeratic deposit that is interpreted as a metasomatically (dolomitized) altered host rock that collapsed at the same time or penecontemporaneously with the presence of a mixed freshwater/marine environment. Fissure 7 has the full range of metasomatism represented by the clasts and matrix observed with the Thecodontosaurus bones and clearly the fall of the collapse need only have been a few metres.
The great numerical dominance of terrestrial reptiles in the vertebrate component demonstrates that the sediments were mainly carried from the Carboniferous Limestone surface by meteoric waters. The Diphydontosaurus bones are found in proportions consistent with fluvial transport over a short distance, and the Diphydontosaurus specimens are accompanied by Pholidophorus fish scales preserved in the same manner as the reptile bones (Whiteside, Reference Whiteside1986). This indicates that non-marine waters were present on or near the limestone surface where the reptiles lived. The white Thecodontosaurus bones would have been brought in by well-oxygenated meteoric waters that ultimately caused the collapse into a lower part of the fissure. Wings’ (2004) report of low pyrite content in the Tytherington Thecodontosaurus bones is consistent with this aerated environment. The dinosaur bones, although disarticulated, are representative of much of the skeleton so that a short transport to final burial is indicated.
The fissure deposits display fluvial sedimentary features including lensing and current shadows, marked fining-upward sequences, and alternating coarse/fine laminae which are characteristic of velocity pulsations. Few definitively marine sedimentary features have been identified, although ooids, some aggregated (fig. 5l, m), are found in the cavern at 17 m (Fig. 9). These may be simply cave pearls but could have formed, probably by water swirl, in a marine zone such as those recorded from Aldabran fissures by Braithwaite, Taylor & Kennedy (Reference Braithwaite, Taylor and Kennedy1973).
7.b. Other fissures
A similar organic assemblage of mixed marine/ terrestrial vertebrates and palynomorphs to that found in fissure 2 has been recovered in the black argillaceous sandstones of fissures 6, 9 and particularly fissure 1. In the latter, a magnificent 4 m high exposure of repeated graded-bedded strata was recorded (fig. 7b–d). This was an unusual, possibly uniquely well-developed, exposure of reworked rounded black sandy mudstone clasts in a matrix of similar lithology that contained an excellent marine/freshwater palynomorph assemblage, blackened fossils of fish and a few lepidosaurs, together with lighter-coloured lepidosaur bones.
It can be hypothesized that the blackened, mainly marine, vertebrate fossils were derived from an anoxic environment where sulphate-reducing bacteria produced H2S that reacted with metals in the hydrothermal Carboniferous Limestone ground waters, causing the blackening. The lighter-coloured terrestrial bones would be decayed and transported in better-oxygenated water. This model is in accord with the mixed marine/terrestrial palynomorph assemblage and fissure 1 sediment deposition, and therefore would have taken place in a marginal marine/terrestrial environment. It can be deduced that sediment reworking is taking place as shown by the discrete scour and rip-up clasts of argillaceous sandstone in a matrix of identical lithology. Sedimentation of the infill in fissure 1 must have been rapid, since oxidation in caves generally destroys palynomorphs such as pollen (Petersen, Reference Petersen1976), which are well-preserved in this fissure infill. The graded beds of fissure 1 can therefore be explained as being formed by periodic dumping of conglomerates into the marginal location from the limestone island.
Any saline waters that covered the Black Rock Limestone surface, at the time when the black argillaceous sandstones were laid down in fissures 1, 2, 6 and 9, may well have been very shallow. This would explain the extremely tiny fragments and overall paucity of the marine fauna in these fissure infills, and their terrestrial components. The occurrence of large reworked lithoclasts in some of these fissure deposits indicates that it was not selective transport that caused the absence of larger bones of marine vetebrates, teeth or shelly invertebrates. A high concentration of Lower Limestone Shale clasts that is present in the palynomorph-bearing matrix of fissure 6 is suggestive of erosive and therefore emergent conditions at the time of deposition. The palaeoenvironmental model proposed for the infilling of fissures 1, 6, 9 and parts of 2 is, therefore, that a very thin layer of saline water intruded in pools or over the top of parts of the Black Rock Limestone, but that the region overlying the Old Red Sandstone and Lower Limestone Shale Group was low, dry land (Fig. 16).
The fissures 12, 13 and 16 that occur on the top quarried level were probably infilled by sediments in brackish water conditions. The water table in the Carboniferous host rock at that time would have been only a few metres below the present-day pre-quarrying Carboniferous Limestone surface. The abundance of Botryoccocus suggests that there were expanses of fresh water (see Sections 4.d.5 and 4.d.6), but the presence of dinocysts, the acritarch Cymatiosphaera polypartita (which is also found in fissure 16), and the non-marine fish Pholidophorus in fissure 13 suggests the presence of euryhaline conditions at infilling.
The fish teeth, including a probable Severnichthys specimen, and the dwarfed crustacean E. minuta var. brodeiana from fissure 12 is also indicative of saline influence at fissure infilling. Many of the lepidosaur (and fish) remains found in fissures 12, 13 and 16 are blackened, indicating some anoxia during deposition, which implies persistant stagnant water conditions at this topographically high level due to a very high water table. This would result in a partially flooded limestone surface allowing intrusions of salt water from a nearby sea.
The Pholidophorus scale in fissure 11 and the gastropod steinkerns and Gyrolepis tooth in fissure 14 again indicate a high water table for the infilling of these two fissures. For fissures 4, 8, 11 and 14, the deposition would have been in a largely freshwater or low salinity environment. This is deduced from the dominance of terrestrial reptiles and the great rarity of any marine fauna, flora or sediments.
For all the Tytherington deposits it is clear that stream flows of meteoric waters brought in most of the reptile material, as demonstrated in part by the sedimentary structures described above in fissure 2. In addition, the following evidence indicates the strong influence of meteoric stream flow. The fissures’ faunas show a dominance of terrestrial reptiles, including weathered bones that must have been exposed on the land surface for a short time (fig. 5o). Weathered bone occurs in all the major fossiliferous deposits except that of fissure 1 and some sediments in fissure 2. Cross-bedded sandstones (laid down by moving water) occur in fissure 12. These cross-bedded sandstones are then overlain by water droplet marks, demonstrating that the system had become dry. Channel lag deposits occur in fissure 11, similar to those depicted by Tucker (Reference Tucker1977) in the Triassic marginal facies of South Wales. The Euestheria fossils from fissures 4 and 12 provide evidence for seasonal rains, as today these crustaceans tend to live in ephemeral pools where fish are absent (Olempska, Reference Olempska2004). In addition, the large E. minuta specimen from the deep-lying fissure 4 is, by analogy with the biology of the modern Euestheridae, indicative of a freshwater environment. The reptile bones have abrasions of the type that Korth (Reference Korth1979) recognized were caused by the action of streams; these include the corrosion and rounding of the bones of Diphydontosaurus avonis (fig. 5j, p, s) from fissure 2. In fact, water-worn bones are found in all tetrapod-bearing fissures. For example, in fissure 8, the teeth, the frontal, and the nasal processes of the premaxilla have been heavily worn on juvenile Clevosaurus and from fissure 4, severe all-round abrasion has occurred on the premaxillae of a specimen identified as Planocephalosaurus. The Diphydontosaurus avonis skull bones in one limestone sample of 443 bones have a mean percentage representation of 29.8%. This is slightly less than half of the predicted percentage representation values for scatological accumulations (60%) but within the range (17% and 36%) of hydraulically sorted microvertebrate assemblages from Nebraska (Korth, Reference Korth1979).
7.c. Overall model of the palaeoenvironment of the fissure infilling
We have described how the Tytherington fissures were likely to have formed and been filled in a mixed regimen of marine and freshwater environments on a low-lying limestone island (Fig. 16). The features found in the Tytherington deposits are similar in many ways to those found by Jones (Reference Jones1992a,b) on the Cayman Islands. Jones (Reference Jones1992a) was able to ascribe the cavity-filling deposits of the Cayman Islands to sediments originating in coastal lagoons, soils, brackish-water ponds, swamps, surface weathering of the bedrock or the limestone cavities themselves. The deposits at Tytherington probably represent a similar variety of environmental origin. At Tytherington there is dolomitized limestone and breccias, dolostones, black mudstones, red rocks (possibly of Terra Rosa origin), clasts of orange-brown and green clays (in fissure 2), as well as unaltered Carboniferous limestone. Some of the laminated void fills of fissure 13 are reminiscent of the limestone precursor of the dolostone, caymanite. Jones (Reference Jones1992a) suggests that evidence favours accumulation of caymanite in a specific phase of transgressive–regressive cycles, namely the early stage of transgression. Dolomitization followed and was produced by transgressive seawater or mixed seawater–freshwater. Jones (Reference Jones1992b) further shows that storms transported terrestrial components such as animal remains and bedrock breakdown products into the voids. To explain the Cayman void-fills, Jones (Reference Jones1992a) described an experimental model, based on commercial products called ‘Sandscapes’, where sediment deposition depends on the density of the grains, the size of the passages, the quantity of sediment supplied and the presence of air. Vortices and turbidity currents may keep finer-grained sediments in suspension until the effect of the vortices is lessened. The finer-grained sediments accumulate on top of the coarser deposits and can result in graded bedding as seen in fissure 1. Overall we envisage the Tytherington fissure fills as infilling in a similar manner to that described for the Cayman Islands and in a climate that resembles the modern-day Bahamas or Cayman Islands with a rainy season and a dry season with lower rainfall. The palaeolatitude of Tytherington would have been about 25–30°N (Torsvik et al. Reference Torsvik, Van Der Voo, Meert, Mosar and Walderhaug2001), a latitude equivalent to the present-day Bahamas.
8. Spatial relationships of the sphenodontids at Tytherington
Figure 17 shows the frequency and locality of occurrences of the three main sphenodontid genera at Tytherington. The identifications were based solely on skeletal elements, that is, the tooth-bearing bones of the skull and dentary, that can be certainly ascribed to a particular genus. Plotting the abundance of the sphenodontid fossils in this manner demonstrates a clear NW–SE pattern with Planocephalosaurus dominant in the west, significant representation of all three genera in the middle and Diphydontosaurus predominant in the east. This may partly be an artefact of sampling but does indicate that there may well be a true spatial difference in occurrence that resulted from the animals living in different micro-habitats. In this regard, the catchment areas nearer the Lower Limestone Shale Group and particularly the Old Red Sandstone would have been expected to have different vegetation and terrain than the surface above the massive limestones. At the least it provides one alternative interpretation to the suggestion that the reptile assemblages in the fissure fills were solely a result of chronological differences. More work could be done to determine whether this type of distribution is found in other fissure localities.
In this respect, Walkden & Fraser (Reference Fraser, Lucas and Morales1993) show a map of the Cromhall fissure localities, and the fissures dominated by Planocephalosaurus (their sites S2, S3, S4, S5 and S7) all lie on the western side of the quarry. However, their fissure S1 that contains no Planocephalosurus but only Clevosaurus hudsoni and Diphydontosaurus is on the southern side of the quarry (Fraser, Reference Fraser, Fraser and Sues1994) and is further away from the Lower Limestone Shale Group.
The Planocephalosaurus-dominated fissures have a great diversity of fauna, particularly at Cromhall but also in fissure 14 at Tytherington, and their proximity to the Lower Limestone Shale Group and Old Red Sandstone may be significant. There, the vegetation may have been more diverse, resulting in a greater variety of food, both animal and plant, than that which grew on the massive limestones.
9. Dating of other sauropsid fissures in localities around Bristol and South Wales
The dating of the Mesozoic fissure deposits around Bristol and in South Wales is problematic. There are no specimens of the fissure reptiles indisputably recorded from normal bedded sequences of the British Triassic. The fissure tetrapods could have lived at any time when there were sufficient meteoric waters during Triassic times. However, it seems highly improbable that these reptiles, if they existed at the same time, would not have been found in the terrestrial reptile-bearing strata of, for example, the Anisian Otter Sandstone Formation, Carnian Arden Sandstone Formation or vice versa. Benton & Spencer (Reference Benton, Spencer, Wimbledon and Palmer1995) have remarked on the similarities of the fissure reptiles to those from Carnian deposits such as the Lossiemouth Sandstone Formation. The biota of the fissures is similar in composition to the Carnian fauna, but no animals are yet proven to be conspecific nor even confidently congeneric. A ‘scleromochlid’, rather than Scleromochlus itself, is reported from Pant-y-ffynon (Benton & Spencer, Reference Benton, Spencer, Wimbledon and Palmer1995), but this crocodylomorph specimen remains undescribed. Benton & Clark (Reference Benton, Clark and Benton1988) reported that Terrestrisuchus is ‘apparently synonymous’ with Saltoposuchus, although Crush (Reference Crush1984) believed it to be generically separate. Most distinctively, the gliding reptile Kuehneosaurus from Emborough, Pant-y-ffynon and Cromhall (Benton & Spencer, Reference Benton, Spencer, Wimbledon and Palmer1995) resembles Icarosaurus from the Late Carnian Lockatong Formation of North America, but the mammal Kuehneotherium, which is also found at Emborough (Fraser, Walkden & Stewart, 1985) has, in normal bedded rocks, a Rhaetian first occurrence. Clevosaurus, which is present and frequently abundant in many fissure deposits, is not recorded from any British Carnian deposit, but a typically UK Carnian genus such as the rhynchosaur Hyperodapedon is entirely absent from the fissures.
The frequently cited dating of the sauropsid fissures as Norian fails to explain the absence of any evidence (such as small bones or teeth) in any of the fissure deposits of the large reptiles well known from European normal bedded sequences at this time, such as Plateosaurus. A summary of the tetrapod assemblages found in the fissure deposits of Bristol and South Wales is given in Table 1 and their geographical location in Figure 1.
Our specific dating of the Tytherington reptiles associated with substantial palynomorph assemblages provides a guide to the dating of the other sauropsid fissure deposits of SW Britain.
Durdham Down is the fissure locality where Thecondotosaurus antiquus was discovered (Riley & Stutchbury, Reference Riley and Stutchbury1836). Moore (Reference Moore1881) regarded the Durdham Down Thecodontosaurus teeth as identical to those that he recovered from the Carnian North Curry Sandstone at Ruishton. He had originally considered the fissure infill to be ‘Rhaetic’ based on the discovery of the ‘Rhaetic bone-bed and remains of that age’ alongside the Thecodontosaurus remains. This is very similar to the field relationsip of the Tytherington deposits and the local Rhaetic, which suggests that Moore's first idea was the correct one. Also, Walker (Reference Walker1969) reported that some of the Ruishton specimens could be better ascribed to Poposaurus. The Tytherington and Durdham Down Thecodontosaurus are very similar in morphology (compare the distinctive ilium from Tytherington, fig. 7e, f, to that from Durdham Down figured by Benton et al. Reference Benton, Juul, Storrs and Galton2000), size and range, and we therefore regard them as conspecific. Clevosaurus and Diphydontosaurus have also been identified in the Durdham Down Thecodontosaurus material held at the Bristol City Museum (D. I. Whiteside, unpub. Ph.D. thesis, Univ. Bristol, 1984), paralleling the co-occurrence of these reptiles in fissure 2 at Tytherington. The Durdham Down rocks held in the Bristol City Museum are extremely similar to those containing the main Thecodontosaurus bones from Tytherington, for example, both contain the same varied lithologies of dolomitized Carboniferous Limestone clasts and would have been formed in the same manner, and they are of a similar age. Indeed the lithology is so distinctive that in 1981 one of us (DIW) recognized that the matrix containing the type specimen of Agrosaurus macgillivrayi Seeley, Reference Seeley1891 was identical to rock from Durdham Down. That matrix also contained Diphydontosaurus, which the requested acid preparation by the British Museum of Natural History confirmed, and therefore it was likely that Seeley had described Agrosaurus from a specimen of the Durdham Down Thecodontosaurus while working on the genus; he later published on Thecodontosaurus and Palaeosaurus (Seeley, Reference Seeley1895). We have discovered palynomorphs (see Table 2, FPC) in a fissure deposit at the Clifton Suspension Bridge, likely to be representative of the fissures found in the original quarry that yielded the Durdham Down Thecodontosaurus. The Clifton fissure is on the same limestone palaeo-island and is only 1.5 km from the original Durdham Down Thecodontosaurus find. The palynomorph assemblage is characterized by Ovalipollis with Ricciisporites with a minor content of both Rhaetogonyaulax and Cymatiosphaera. The palynofacies is dominated by AOM and the assemblage falls within the cluster (FP2.2, 6.1, 13.2; Figs 13, 15) attributable to the lower part of the Westbury Formation. It is therefore clear that the Durdham Down and Tytherington deposits shared a similar palaeo-island environment and Rhaetian age. Acid preparation of some of the remaining Durdham Down rocks could be expected to reveal Penarth Group fish.
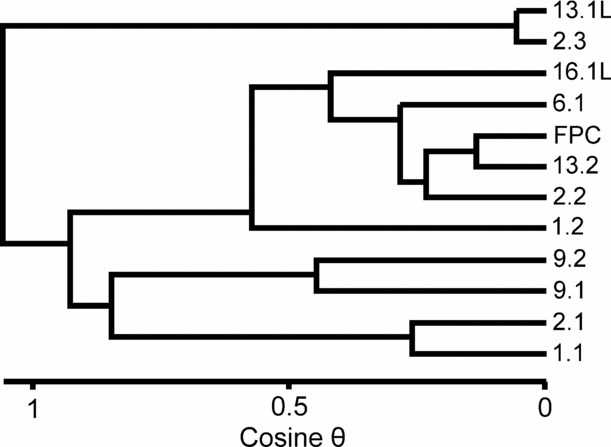
Figure 13. Cluster analysis of palynological assemblages from Tytherington plus the Clifton Suspension Bridge ‘FPC’ sample. Palynological data were recalculated to include only the major elements (Ricciisporites tuberculatus, Ovalipollis pseudoalatus, Rhaetipollis germanicus, the Circumpolles pollen, Rhaetogonyaulax rhaetica and Cymatiosphaera polypartita). A number of different methods were used to make the similarity matrix and perform the cluster analysis. These generally gave robust results.The illustrated dendrogram was clustered using UPGMA and a Cosine θ similarity measure using the MVSP © pro-gram (Kovach Computer Services). The analysis shows three clear assemblage groups in the fissure deposits. The two palynological samples that were in a limestone lithology are marked L, that is, FP13.1L and FP16.1L.
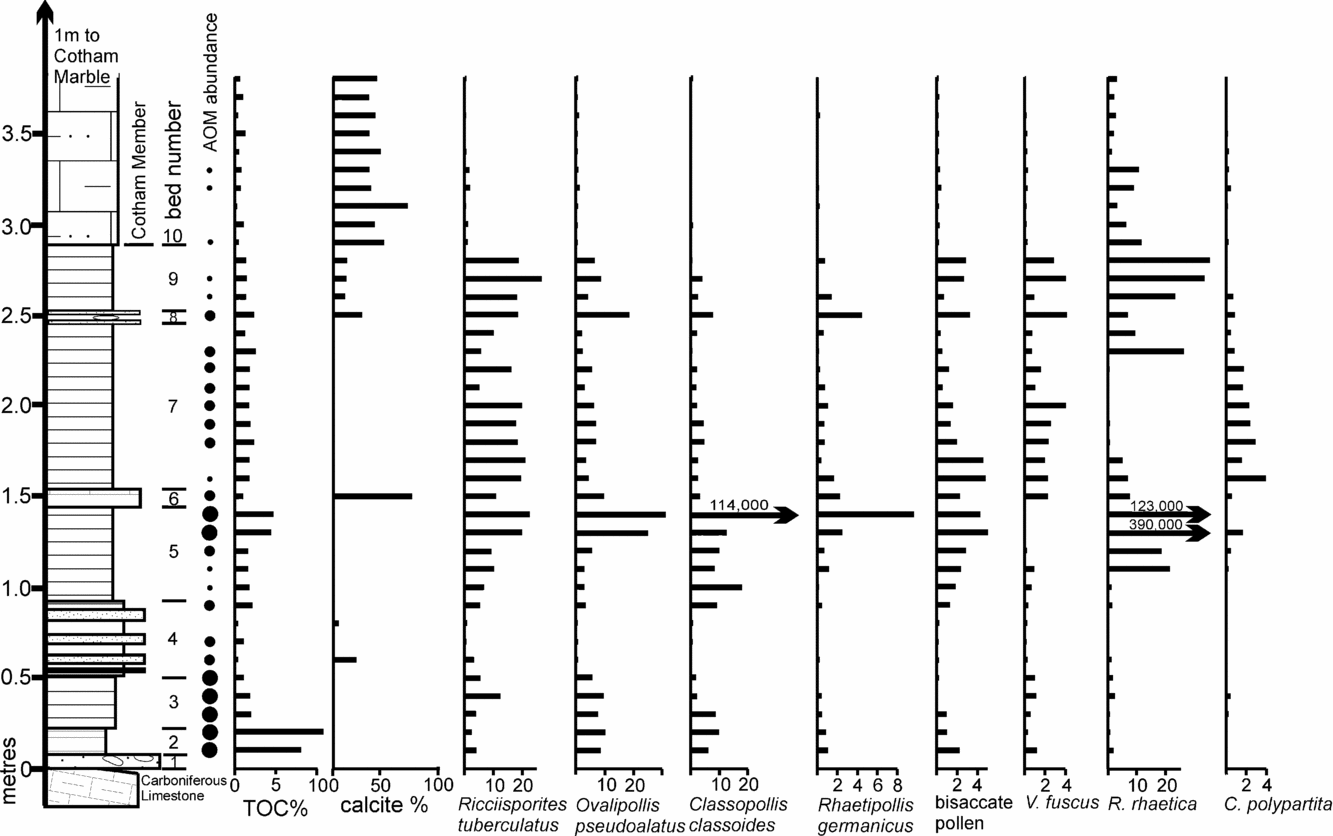
Figure 14. Quantitative palynological profile of the main terrestrial and marine palynomorphs. from the Penarth Group at Hampstead Farm Quarry. Quantitative numbers were determined using a Lycopodium spike as per Thomas et al. Reference Thomas, Marshall, Mann, Summons and Maxwell1993 and are expressed in thousands of palynomorph g−1 rock. Three numbers (114000, 123000 and 390000) are off the scales used. Also shown is the Total Organic Carbon content (TOC%), calcite % (both measured using a CE-1108 elemental analyser) and the relative abundance of AOM. Bed numbers are from Donovan, Curtis & Curtis (1989). Note the very characteristic double peak of Rhaetogonyaulax rhaetica separated by an acme of Cymatiosphaera polypartita. The abundances of all the palynomorphs declines markedly in the Cotham Member with R. rhaetica declining the least and hence becoming relatively abundant.
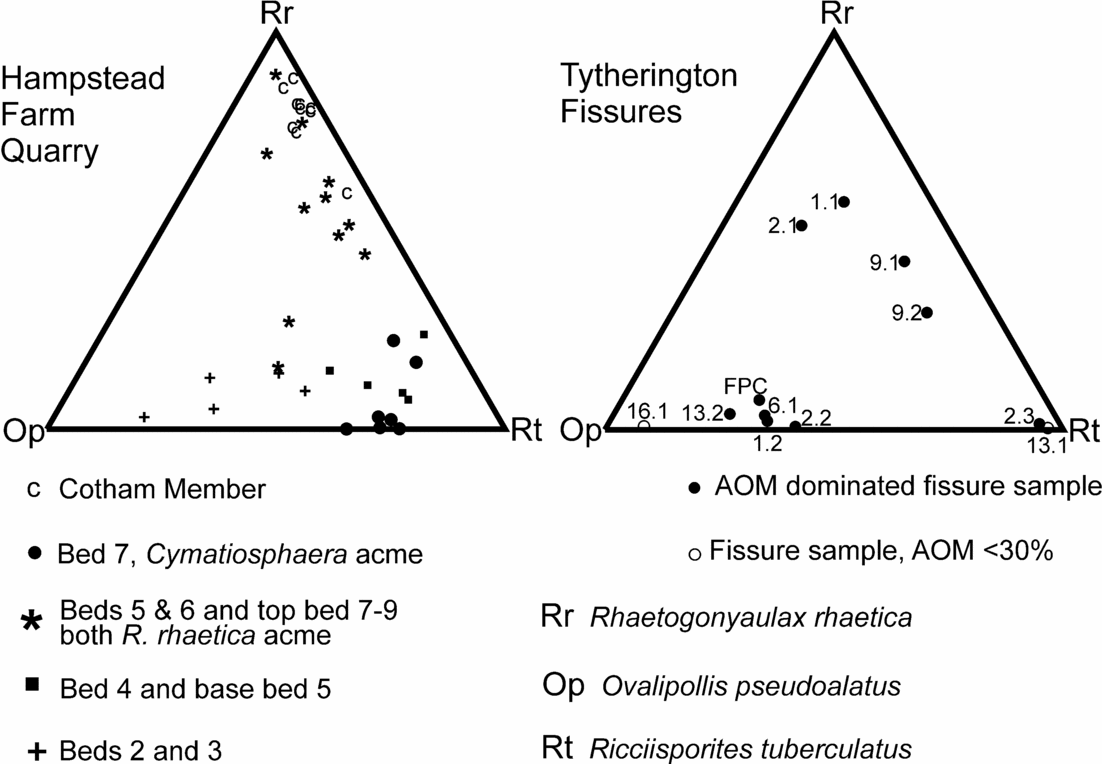
Figure 15. Ternary plots showing comparative assemblages based on the recalculated relative percentages of Rhaetogonyaulax rhaetica, Ovalipollis pseudoalatus and Ricciisporites tuberculatus from Hampstead Farm Quarry and the palynomorph-bearing fissure deposits at Tytherington and the Clifton Suspension Bridge ‘FPC’.
The deposits at Cromhall Quarry yielding the most diverse herpetofauna of the sauropsid fissures have usually been dated as Norian. In earlier papers on the Cromhall deposits, Robinson (Reference Robinson1957a) and Fraser & Walkden (Reference Fraser and Walkden1983, Reference Fraser, Walkden, Reif and Westphal1984) substantially based their dating on the presumed similarity between the red and green sediments of the fissures and those of the Mercia Mudstone Group. However, there are red and green clastic sediments, lithologically similar to the Cromhall deposits, that are recorded from the Cotham Member of South Wales (Strahan & Cantrill, Reference Strahan and Cantrill1904; Francis, Reference Francis1959). We have discovered a rock from fissure 2 at Tytherington (fig. 8e) that contains Westbury Formation palynomorphs (FP2.2) in thin black laminae between red and green sediments of ‘Keuper’ type, confirming that such lithologies occurred in the Penarth Group.
Fraser (Reference Fraser, Fraser and Sues1994) and Walkden & Fraser (Reference Fraser, Lucas and Morales1993) have compiled a suggested sequence of sediment associations which they correlate with a Carnian to Rhaetian chronostratigraphy. This is based on their suggested geomorphological and sedimentological links between the topographical heights and normal bedded sequences of Late Triassic strata. Fraser (Reference Fraser, Fraser and Sues1994) considers that many of the fissure fills are pre-Rhaetian but with no evidence of an earlier date than equivalent to the Blue Anchor Formation. He emphasized that the upper date for the Cromhall infills, including deposits with Diphydontosaurus and Clevosaurus hudsoni, can ‘be assigned with some certainty’ to the basal Penarth group, that is, the Westbury Formation. However, there is clear evidence from studies of modern day karst that contemporaneous sediments can be deposited in voids at different depths, well below the surface (Mylroie & Carew, Reference Mylroie, Carew, Budd, Saller and Harris1995). Jones (Reference Jones1992a), working in the Cayman islands, reported the same phenomenon, stating that ‘sediment from a single influx may accumulate in cavities that are at different stratigraphic levels’. Therefore, judgements of vertical relations between different fissure deposits within a locality and without intrinsic dating evidence are subject to considerable scepticism.
The Cromhall fissures believed by Walkden & Fraser (Reference Fraser, Lucas and Morales1993) to be pre-Rhaetian have openings that are topographically 6–7 m higher than the ‘capping sequence’ they accept as Rhaetian and might be expected to have been open during that time. These particular fissure faunas have an abundance of Planocephalosaurus, and M. Curtis (pers. comm.) reports the finding of 14 teeth of the typically Penarth Group Gyrolepis with a well-preserved Planocephalosaurus maxilla in Cromhall fissure S4 of Fraser (Reference Fraser1985) and Walkden & Fraser (Reference Fraser, Lucas and Morales1993). This therefore contradicts a pre-Rhaetian dating for the main reptile-bearing Cromhall fissures.
Lucas et al. (Reference Lucas, Heckert, Fraser and Hubert1999), using land-vertebrate faunachrons (LVF) as the basis for dating, considered that the presence of a single scute that they identified as Aetosaurus established the date of one Cromhall fissure fill as from the Revueltian LVF (within the Norian and pre-Rhaetian). Lucas & Tanner (Reference Lucas and Tanner2006), however, accept that Aetosaurus is also known from the younger Apachean LVF (Late Norian to Rhaetian) from the Rock Point Formation in Colorado, which they ‘tentatively’ assigned to the Late Norian. The Aetosaurus scute was only a tentative identification by Fraser (Reference Fraser1988b) and not confirmed as a member of the fauna in his later (Fraser, Reference Fraser, Fraser and Sues1994) summary paper on the Cromhall fissure fills. Lucas et al. (Reference Lucas, Heckert, Fraser and Hubert1999) confirmed the identification but Rayfield et al. (Reference Rayfield, Barrett, McDonnell and Willis2005) emphasized that there is ‘disagreement over the status of supposed ‘Aetosaurus’ remains’. For example, Sues et al. (Reference Sues, Olsen, Carter and Scott2003) pointed out that Stegomus arcuatus, referred to Aetosaurus by Lucas, Heckert & Huber (1998), was taxonomically distinct and could well have different stratigraphical ranges. There are other stagonolepidids Typothorax and Redondasuchus, identified as distinct by Heckert, Hunt & Lucas (1996), which are regarded as not distinguishable by Long & Murry (Reference Long and Murry1995). Although Benton (Reference Benton, Fraser and Sues1994) mentions aetosaur elements from the ‘Rhaetian’ of southwest England, these probably refer to the Cromhall fossils (Benton, pers. comm.) and there is a possible occurrence of a European Rhaetian stagonolepidid reported by Cuny (Reference Cuny1995) from Boisset in France. We conclude that the Aetosaurus scute identification is dubious and that the pre-Rhaetian dating of Cromhall infills on this evidence by Fraser is unfounded.
Of the SW Britain sauropsid fissure faunas, Tytherington has the greatest similarity to the diverse herpetofauna of Cromhall Quarry (see Table 1). Whiteside (Reference Whiteside1986, and previously D. I. Whiteside, unpub. Ph.D. thesis, Univ. Bristol, 1984) suggested that the Cromhall fissures are of similar Rhaetian age to those at Tytherington, based on the similarity of elements of the tetrapod fauna and to the finding of Pholidophorus in the same normal bedded horizon (that is, the capping sequence of Walkden & Fraser, Reference Fraser, Lucas and Morales1993) as Clevosaurus. In particular, our recent research has demonstrated that Tytherington fissure 14 has a number of features that characterize the Cromhall fissures, such as abundant Planocephalosaurus and the presence of Tricuspisaurus, Pelecymala and Sigmala. This has confirmed the prediction made by Whiteside that this fissure was the most similar to those at Cromhall. In parallel to M. Curtis’ discovery of Gyrolepis with Planocephalosaurus at Cromhall, a Gyrolepis tooth (fig. 5ll) has been found in fissure 14 at Tytherington in a crinoidal sandy limestone deposit that closely resembles the Cromhall lithology and ‘faunal association B’ of Walkden & Fraser (Reference Fraser, Lucas and Morales1993). Most significantly, our dating of Planocephalosaurus associated with Westbury Formation palynomorphs in fissure 2 at Tytherington demonstrates that the Cromhall deposits are most likely to be Rhaetian at a time equivalent to the Westbury Formation.
At Emborough Quarry the field relationships of the fissure deposit with the nearby ‘Rhaetic’ have been used to date the infill as pre-Rhaetic (Robinson, Reference Robinson1957a; Fraser, Walkden & Stewart, 1985). The Emborough fissure deposit contains Kuehneosaurus which is also found at Cromhall but not confirmed in the Tytherington fauna. Fraser, Walkden & Stewart (1985) claimed the first pre-Rhaetic therian mammal, Kuehneotherium, from the Emborough fissure, but Whiteside & Marshall (Reference Whiteside and Marshall1985) suggested that the Kuehneotherium should be used to date the fissure rather than vice versa. For example, a Kuehneotheriid has been found in the Rhaetian beds of Luxembourg (Godefroit et al. Reference Godefroit, Cuny, Delsate and Roche1998) and another possible occurrence in the Rhaetian of Boisset is noted by Cuny (Reference Cuny1995). To show that the Emborough deposit is ‘pre-Rhaetic’, the basal Rhaetic would have to actually cap the fissure. This has not been demonstrated, and the fissure could have been filled during the Westbury Formation before the nearby ‘Rhaetic’ sequence or during any subsequent Late Triassic transgressions. No ‘Rhaetic’ clasts in the deposit were found by Fraser, Walkden & Stewart (1985) but ‘Rhaetic’ fossils have been recognized in subsoil at the margins of the fissure deposit (Savage, Reference Savage and Savage1977, p. 90, fig. 3).
Lucas & Hunt (Reference Lucas, Hunt, Fraser and Sues1994) were also not convinced that the Emborough deposit was pre-Rhaetian in age. In fact, none of the reptile-bearing fissures at Batscombe, Cromhall, Durdham Down, Emborough and Tytherington has overlying strata of proven basal Rhaetian or ‘Rhaetic’ rocks. It seems to us that the evidence points to a dating of the Emborough deposit as Rhaetian, after the initial phase of the Westbury transgression. Kuehneosuchus that is similar to Kuehneosaurus (co-generic?) is found at Batscombe (Robinson, Reference Robinson1962) and we would therefore suggest that the infill there is Rhaetian.
Pant-y-ffynon also has a fauna of Thecodontosaurus, Terrestrisuchus, Clevosaurus and a coelurosaur, in common with Tytherington (Table 1). We judge that this similarity to the palynomorph-dated Tytherington fauna demonstrates that the Pant-y-ffynon infills are most likely to post-date the start of the transgression in the Westbury Formation. We thus conclude that all the major sauropsid fissures at Durdham Down, Cromhall, Pant-y-ffynon, Emborough, and Batscombe are, like Tytherington, infilled during the Rhaetian. If Fraser (Reference Fraser1986a, 1994) is correct in his tentative identification of Clevosaurus at Highcroft and Planocephalosaurus at Ruthin and Barnhill Quarries then, based on our dating of the genera, those deposits are also probably Rhaetian.
10. Comparison of the Tytherington reptile assemblage to other parts of the world
The dating of the Tytherington tetrapod fauna as determined here agrees well with the occurrence of the same genera in normal bedded sequences from throughout the world (Fig. 18). There are a number of sphenodontian species known from the Late Triassic to Early Jurassic interval. Clevosaurus species are known in deposits mainly of earliest Jurassic age from North America and Southern Africa (Sues, Shubin & Olsen, 1994) and China (Luo & Wu, Reference Luo, Wu, Fraser and Sues1994), although Wu (Reference Wu, Fraser and Sues1994) suggests that the stratum yielding Clevosaurus petilus is of latest Triassic (Rhaetian?) age.
Duffin (Reference Duffin1995) has ascribed some material from the Belgian Rhaetian to Clevosaurus and others as possibly Diphydontosaurus. Diphydontosaurus has also been tentatively identified in the Norian of Italy, but the description given of the Italian specimen by Renesto (Reference Renesto1995) has fewer pre-maxillary teeth and different proportions of pleurodont and pleuracrodont dentary and maxillary teeth than the D. avonis from Tytherington. Fossils of a sphenodontian with affinities to Diphydontosaurus were recorded by Sues & Olsen (Reference Sues and Olsen1990) in middle Carnian deposits of the North American Richmond Basin. However, they do not figure the specimen and a comparison cannot be made here. Evans, Prasad & Manhas (2001) described the Indian Early Jurassic Rebbanasaurus and Godavarisaurus that have similarities to, respectively, Clevosaurus and Diphydontosaurus. An Early Jurassic (Hettangian to Sinemurian) assemblage of sphenodontians, prosauropods and terrestrial crocodiles is known from the Lower Lufeng Formation of China (Luo & Wu, Reference Luo, Wu, Fraser and Sues1994) and the McCoy Brook Formation of Canada (Shubin, Olsen & Sues, 1994; Sues, Shubin & Olsen, 1994).
Benton et al. (Reference Benton, Juul, Storrs and Galton2000) found that relationships between Thecodontosaurus and other prosauropods were difficult to resolve, and the genus is distinguished mainly on the absence of derived characters. Benton et al. (Reference Benton, Juul, Storrs and Galton2000) did not include Chinese prosauropods in their comparisons. However, Whiteside (Reference Whiteside1984) had suggested that the Thecodontosaurus of Tytherington bore considerable similarities to the Early Jurassic Lufengosaurus and Gyposaurus described by Young (Reference Young1941a,b) or to the Late Triassic Plateosaurus and that it may itself be a dwarfed plateosaurid as a consequence of the population inhabiting a small island. A dwarfed species, in a restricted landmass, would be more likely to retain a minimum viable population (MVP) which, discussions in Whittaker (Reference Whittaker1998) suggest, may be about 50 but up to 500 individuals to maintain genetic variation. There were contemporaneous European (Benton, Reference Benton, Fraser and Sues1994) and nearby populations of Plateosaurus as demonstrated by the finding of a claw in the Westbury Formation of Nottinghamshire by Martill & Dawn (Reference Martill and Dawn1986). The large quantity of prosauropod bones found at Tytherington is compatible with the Apachean LVF (land vertebrate faunachron) of Lucas (Reference Lucas1999), which is primarily Rhaetian and very latest Norian. The cursorial crocodile Terrestrisuchus is a sphenosuchid which is found in the Late Triassic and Early Jurassic rocks of North America, Southern Africa and China. The coelurosaur from Tytherington has features most in common with Megapnosaurus (‘Syntarsus’), now thought to be congeneric with Coelophysis (Bristowe & Raath, Reference Bristowe and Raath2004) and therefore present in Late Triassic–Early Jurassic times.
The dating of the Tytherington reptiles as Rhaetian derived from their association with palynomorphs is therefore entirely consistent with the herpetofauna from normal bedded sequences from around the world.
11. A comment on the Triassic/Jurassic boundary extinctions
Lucas & Tanner (Reference Lucas and Tanner2004) have concluded that the tetrapods were affected by multiple extinctions through the Late Triassic rather than the single end-Triassic extinction event as supported by, for example, Colbert (Reference Colbert1958) and Olsen et al. (Reference Olsen, Kent, Sues, Koeberl, Huber, Montanari, Rainforth, Powell, Szajna and Hartline2002). Although further debate of this issue is beyond the scope of this paper, one element of criticism by Lucas & Tanner is challenged here. The list of 11 terrestrial reptile families listed by Benton (Reference Benton and Benton1993) as exterminated at the Triassic/Jurassic boundary was queried by Lucas & Tanner (Reference Lucas and Tanner2004), who believed that only two of these families had Rhaetian records: the Phytosauridae and Procolophonidae. Instead, they concluded that most of the families have Norian extinctions. Representatives of the Thecodontosauridae (Thecodontosaurus) and Saltoposuchidae (Terrestrisuchus is believed to be closely related to Saltoposuchus by Benton & Clark, Reference Benton, Clark and Benton1988, and Fraser, Reference Fraser, Fraser and Sues1994) have been found in the Rhaetian deposits at Tytherington. At Cromhall, where we believe that the evidence strongly favours a Rhaetian age assignment, there are representatives of the Kuehneosauridae (Kuehneosaurus) and Stagonolepididae (identified as Aetosaurus by Lucas et al. Reference Lucas, Heckert, Fraser and Hubert1999). If Tricuspisaurus, known from Tytherington and Cromhall, is classified as a trilophosaur (although we believe it is not), then this extends that group also into the Rhaetian.
Therefore at least four of the reptile families considered by Lucas & Tanner (Reference Lucas and Tanner2004) to be extinct at the end of the Norian are certain or likely to be present in the Rhaetian. Assuming that these families died out in the Rhaetian, then this gives further support to a significant large extinction event at the very top of the Triassic. Also, our findings cast great doubts on the validity of postulating prolonged extinctions where dating has been based on indirect evidence or where the taxonomy is highly debatable. It is also of great interest that the genus Clevosaurus spans the time before and after the postulated Late Triassic extinctions; it may well be that as dating and taxonomy continue to be refined, other genera will be recorded as extending across the extinction event(s).
12. The Tytherington Triassic reptiles as an island fauna and Quaternary analogues
We have further expanded the model of Whiteside & Robinson (Reference Whiteside and Robinson1983), who suggested that part of fissure 2 was infilled in a fluctuating marine/freshwater environment. This environment in the Tytherington fissures leads to the conclusion that, in Rhaetian times, the terrestrial tetrapods lived on a small island. The insular nature of the fissure herpetofauna is very significant in understanding the reptiles’ ecology and evolution. Also, amphibians which could colonize the palaeo-islands would have been killed by the surrounding saline waters, which explains their absence from Tytherington and the other fissure localities shown in Table 1. The sea barrier would have impeded colonization by other tetrapods such as mammals, which are apparently absent from nearly all sauropsid localities.
During the Rhaetian, South Wales and the English West Country were transgressed by the sea, and many of the rocks show evidence of repeated subaerial exposure and submergence (e.g. Warrington & Ivimey-Cook, Reference Warrington, Ivimey-Cook and Taylor1995). More recently, Hesselbo, Robinson & Surlyk (2004) have reported a sea-level lowstand within the Cotham Member (Lilstock Formation) of the southern UK.
There are Quaternary analogues of the Tytherington Rhaetian insular model of a herpetofauna living on small islands with their bones washed into solutional fissures. The fluctuating sea levels in the Quaternary, although on a much greater scale and resulting from different causes, produced a similar effect. Quaternary lepidosaur-dominated herpetofaunas are well known from solutional fissures (dolines or banana-holes) on many islands, for example, Aldabra (Arnold, Reference Arnold1976), Barbuda (Etheridge, Reference Etheridge1964) and New Providence (Etheridge, Reference Etheridge1966), and from other sediment-filled cavities on islands (e.g. Bravo, Reference Bravo1953). Some solution fissure fills contain other vertebrates, such as birds on New Providence (Brodkorb, Reference Brodkorb1959). In common with the Tytherington and Cromhall deposits, many of these Quaternary fissure fills have produced huge numbers of lepidosaur bones; over 1000 bones of squamate lizards have been found in fissure fills on Aldabra, New Providence and Barbuda. The occurrence of the fossils and the skeletal elements recovered from Aldabra is extremely similar to the lepidosaur bones at Tytherington (D. I. Whiteside, pers. obs., Aldabran collection, British Museum Natural History). In many cases, it is reported that these Quaternary assemblages are the result of regurgitated owl pellets, but our analysis of the skull and jaw bones from Barbuda taken from the data in Etheridge (Reference Etheridge1964) produces a minimum representation of 17.6%, which suggests that they are current-sorted. It is also reported that the Aldabran assemblages are suggestive of ‘concentration by flowing water’ (Taylor et al. Reference Taylor, Braithwaite, Peake and Arnold1979).
A major feature of modern small sub-tropical and tropical insular lepidosaur faunas is the very high population densities recorded compared to equivalent mainland species (Case, Reference Case1975; Turner, Reference Turner, Gans and Tinkle1977; Bennett & Gorman, Reference Bennett and Gorman1979). On some modern day islands, arboreal Anolis species reach densities of 0.97 m−2 (Schoener & Schoener, Reference Schoener and Schoener1980). It is not possible to know if some of the Tytherington lepidosaurs were arboreal, but it seems quite plausible that the very high numbers of individuals recorded could have derived from a very small area around the entrance to the fissures. Whiteside (Reference Whiteside1986) has reported a minimum number of 40 individual Diphydontosaurus in 5 kg of a detrital limestone block. It is estimated here that in the Diphydontosaurus-bearing rock collected from the same horizon in fissure 2, up to 400 individuals may have been present. A reasonable extant analogue may be the gecko Gonatodes, an animal of similar size to Diphydontosaurus that lives on the arid Caribbean island of Bonaire. Bennett & Gorman (Reference Bennett and Gorman1979) reported densities of 0.42 per m2 which they believed was ‘almost certainly an underestimate’ for Gonatodes. It is therefore quite feasible for all the Diphydontosaurus from one horizon in one fissure deposit to have been sampled from an area of about 950 m2.
Other characteristics of modern-day small sub-tropical and tropical islands are also of interest; they generally lack very large animals (cf. the absence of animals such as Plateosaurus at Tytherington), sometimes lack terrestrial mammals and amphibians, and have a low species diversity compared to an equivalent area of the mainland. Case (Reference Case1975) has shown that for small islands in the Gulf of California the number of lizard species ranges between 3 and 8, whereas the number on the nearby mainland is 15. The paucity of the modern-day insular herpetofaunas is reflected in the fossil reptile diversity of the solution cavity fills found on the same islands. For some islands the number of fossil lizard species in a single cave or doline is greater than living species on the island. Etheridge (Reference Etheridge1964) records six species of fossil lizards from a cave on Barbuda, whereas only five species currently live on the island. Two of the fossil species are still living on Barbuda, one is probably ancestral to another extant Barbudan species and one other is possibly ancestral to a fourth lizard still living on Barbuda.
Seventy-five per cent of the extant lizard genera of New Providence are found as Pleistocene fossils in one ‘banana hole’ (Etheridge, Reference Etheridge1966). In the Point Houdal fissure of Aldabra (Arnold, Reference Arnold1976), a Pleistocene terrestrial herpetofauna of a giant tortoise and six lepidosaur species reflects the extant reptile biota of one giant tortoise and three lepidosaurs (although the species are different). From these considerations it appears that the fossil reptiles have a similar biotic composition to the terrestrial herpetofauna from which they derive. By analogy it is not unreasonable to assume that the assemblages in the Tytherington fissures give excellent qualitative indications of the species content and ecosystem of the complete terrestrial herpetofauna existing at the time of deposition.
There is considerable variation in the distribution of Pleistocene reptile species between different cave deposits on small islands. For example, Etheridge (Reference Etheridge1964) recorded lizard fossils from five caves on Barbuda and found a large iguana in only one and medium-sized Anolis in two. Small Anolis species and two other lizards were each absent from one cave. The relative abundance of the species in Barbuda also varies, with one cave yielding 85.5% Anolis in the fossil lizard fauna and another giving 33.4%. This type of variation of reptile species is seen between the fissure deposits at Tytherington (Table 3), and between Tytherington, Cromhall and the other localities that are shown in Table 1. These variations can be due, in part, to sampling of different micro-habitats, but it is also clear that only a short period of geological time is needed to produce such differences. The duration for the Barbuda cave deposits is between Mid- to Late Pleistocene and pre-Columbian (Etheridge, Reference Etheridge1964) which is 1 Ma or less, and probably within a far smaller time-span.
While lacking very large animals, except for marine crocodiles, there are medium-sized herbivorous iguanids living on present-day small islands, such as Cyclura in the Bahamas, occupying a niche similar to that predicted for Thecodontosaurus on the Tytherington Rhaetian island. Although slightly smaller than Thecodontosaurus (1.5 m long as against 2 m), these iguanines are similar in order of magnitude, and populations can be supported by very small islands. The Galapagos land iguana Conolophis occurs on the island of Sur Plaza which is ‘barely 100 yds by 150 yds’ (12500 m2: Fisher, Simon & Vincent, 1969). The large predatory Varanus komoedensis, up to 3 m long (cf. the Tytherington coelurosaur), is found on small islands such as Gili Motang, where 100 Komodo dragons live (Ciofi, Reference Ciofi1999) in an area less than 10 km2.
Therefore, it can be demonstrated that the reconstructed Tytherington terrestrial herpetofaunas are paralleled by Quaternary, including recent, insular herpetofaunas. The question arises as to why these insular herpetofaunas should be so well preserved with abundant micro-vertebrate fossils. It is suggested here that the following may be some of the reasons. Firstly, the high density of the lepidosaur populations results from a low species diversity and the island tetrapods therefore expand their niche space. In the absence of ectothermic and endothermic competitors, predator intensity may be less than on the mainland and there may be a fence effect in which emigration is restricted (Case, Reference Case1975). Secondly, on islands there is a relatively short transport distance between death and final burial. This will maximize the chances of preserving delicate lepidosaur bones intact and with little significant evidence of water-wear. The rapidity of deposition is enhanced because the subterranean drainage from small catchments would take sediments to localities at, or even below, sea level (Whiteside & Robinson, Reference Whiteside and Robinson1983; Fig. 16). Reworking and transport of bones would therefore be minimal. Sea-level fluctuations are recorded in the Rhaetian of the West Country and South Wales (Hamilton, Reference Hamilton1962; Warrington & Ivimey-Cook, Reference Warrington, Ivimey-Cook and Taylor1995) and hence are similar to the repeated submergent/emergent history of many islands in the Quaternary described by Taylor et al. (Reference Taylor, Braithwaite, Peake and Arnold1979) and Black (Reference Black, Jeans and Rawson1980), for example. At times of high sea level, the deposits in the fissures would have become further consolidated and at times of emergence, new solutional pipes would form to then be filled during the next rise of sea level.
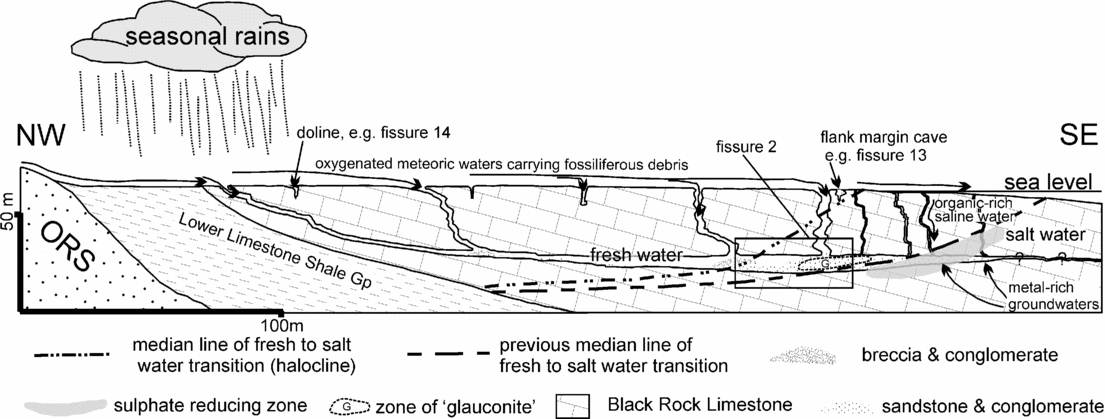
Figure 16. Palaeoenvironmental model of the Tytherington reptile-bearing fissures during their Rhaetian infilling. The cross-section runs N–W to S–E and includes fissures 2, 13 and 14. The occurrence of seasonal rainfall is predicted from the presence of Euestheria minuta, the palaeolatitude, the dolomitization of the calcite layers in the Rhaetian laminated sediments and general climatic indicators in the region at the time. The vegetation cover (not shown) would be expected to be fairly dense, especially around the entrances to the fissures, as the freshwater lens that was supported by the saline waters of the Rhaetian sea would maintain fresh water very near the limestone surface. Modern islands below about 10 ha can lack a permanent freshwater lens (Whittaker, Reference Whittaker1998), but the field relationship of local Rhaetic deposits suggest that the Tytherington paleo-island would have been much larger than this, perhaps about 700 ha.
Figure 17. Map of the fissures showing the relative abundance of the three common sphenodontids, Clevosaurus, Diphydontosaurus and Planocephalosaurus, based on skeletal elements that can be attributed with certainty (all are skull or lower jaw bones). There is a clear change in the relative abundance of these sphenodontids from west to east across the quarry. The western part of the quarry is bordered by the Lower Limestone Shale Group, and the easterly fissures are furthest away from that stratum and the Old Red Sandstone.

Figure 18. Stratigraphical comparisons between reptiles found in the Tytherington Triassic fissures and the same or closely related genera from normal bedded sequences worldwide. Data from Shubin & Sues (Reference Shubin and Sues1991), Wu (Reference Wu, Fraser and Sues1994), Shubin, Olsen & Sues (1994), Luo & Wu (Reference Luo, Wu, Fraser and Sues1994), Sues, Clark & Jenkins (1994), Benton (Reference Benton, Fraser and Sues1994), Sues & Reisz (Reference Sues and Reisz1995), Sues, Shubin & Olsen (1994), Renesto (Reference Renesto1995), Duffin (Reference Duffin1995), Lucas (Reference Lucas1998, Reference Lucas1999), Irmis (Reference Irmis2004) and Säila (Reference Säila2005). Ch is China, Eu is Europe, NAm is North America and SAfr South Africa. The Tytherington coelurosaur is most similar to Megapnosaurus which Lucas & Tanner (Reference Lucas and Tanner2006) regard as first appearing in, and defining the beginning of, the Dawan LVF (equivalent to the Sinemurian), but which Bristowe & Raath (Reference Bristowe and Raath2004) regard as congeneric with Coelophysis. In addition to the data displayed, Bonaparte & Sues (Reference Bonaparte and Sues2006) have recently described Clevosaurus from the Caturitta Formation (?Carnian–Norian) of Rio do Sul, Brazil.
Considering all of the Late Triassic and Early Jurassic fissure faunas of the Bristol and South Wales regions as originating from an archipelago of palaeo-islands means that attempts to ascribe faunal succession, except in the most general sense, may be fruitless. In this respect, the younger fissure deposits tend to contain mammals, whereas the older ones (with the exception of Emborough), as described by Robinson (Reference Robinson1957a), do not. This observation is still valid, since despite a huge amount of acid preparation of suitable rock, no Mesozoic mammals have been confirmed at Tytherington and Cromhall. However, even nearby islands are unlikely to have the same tetrapod components living contemporaneously, and some species may never reach some localities. As MacArthur & Wilson (Reference MacArthur and Wilson1963) have shown, small islands have high extinction rates and those near to landmasses have high immigration rates. This is exactly the situation pertaining to these Rhaetian and Rhaeto-Liassic islands.
13. Mode of death of the fissure tetrapods
The island model explains the presence of small-sized archosaurs and high densities of lepidosaurs. The short distance from death to final burial of the bones expected on a small island with deposition at or below sea level also explains why the bones are so well preserved. However, the means by which the reptiles met their death is more problematic.
The reptiles and mammals found in the fissure deposits are found, most frequently, in the form of numerous disarticulated skeletal elements. However, there are significant numbers of articulated remains of reptiles known from fissure localities: Clevosaurus from Cromhall (Fraser, Reference Fraser1988a), Thecodontosaurus and Terrestrisuchus from Pant-y-ffynon (Kermack, Reference Kermack1984 and Crush, Reference Crush1984, respectively), sphenodontians and unnamed archosaurs also from Pant-y-ffynon (Fraser, Reference Fraser, Fraser and Sues1994) and Kuehneosaurus from Emborough (Robinson, Reference Robinson1962). There is also an articulated limb of Diphydontosaurus (figured as Clevosaurus by Halstead & Nicoll, Reference Halstead and Nicoll1971) in the Durdham Down fissure material (D. I. Whiteside, unpub. Ph.D. thesis, Univ. Bristol, 1984; Fraser, Reference Fraser, Fraser and Sues1994).
What can explain the death of great numbers of lepidosaurs and other small tetrapods that are transported as articulated skeletons and/or numerous individual bones? Furthermore, can we also explain the presence of small dinosaurs of about 2 m length with the same cause of death? The most common explanation to identify the death of the lepidosaurs is by means of a predator that either regurgitated the bones in pellet form or in their scats. Kühne (Reference Kühne1956) concluded that the character of the skeletal element accumulations, together with the tooth marks on the bones of Oligokyphus from Windsor Hill, were the result of predation. Evans & Kermack (Reference Evans, Kermack, Fraser and Sues1994) also suggest that the original bone accumulation was via predators, either in the form of pellets by theropod dinosaurs using a bird-type digestive system, or by a mammal such as a large morganucodontid. Walkden & Fraser (Reference Fraser, Lucas and Morales1993), while accepting that predators may have played a part in originating the tetrapod bones, reported that the bones do not have features of predation such as fractures at specific points or a bias towards some elements such as dentaries.
Whiteside (Reference Whiteside1986) preferred a physical environment for the death of the large number of Diphydontosaurus represented in fissure 2 at Tytherington. He suggested that the animals lived at or near the entrances to the fissures and that night-time storms could catch the torpid animals unaware.
None of the explanations discussed so far can satisfactorily account for the associated remains of small dinosaurs such as the Thecodontosaurus assemblage common to Tytherington and Durdham Down, or the articulated skeletons of reptiles described above. Perhaps there were a number of factors both biotic and abiotic involved in the death of the tetrapods. It is very likely that mass starvation was one factor but evidence of this is difficult to discern. However, there is one environmental factor, with direct evidence, that can provide a plausible hypothesis for the death of all the tetrapods and their remains found in the Rhaetian and Rhaeto-Liassic fissure fills.
Forest fires, particularly those occurring at the end of the dry season, would kill significant numbers of reptiles and mammals, both large and small. The animals would be killed by direct heat or smoke inhalation. Small lepidosaurs hiding under broken limestone blocks would be baked alive and arboreal reptiles such as Kuehneosaurus would be engulfed by canopy fire. Larger reptiles such as dinosaurs, probably moving in groups, would be surrounded by the fire, perhaps near water at the entrance to a fissure and become asphyxiated by smoke or toxic gases. The fire would also kill larger terrestrial predators, such as coelurosaurs, that would otherwise scavenge the remains and probably destroy the bones in their digestive tracts. Some dead animals may be scavenged but the abundance of food would leave many bodies untouched. Any rains that followed the fire would wash the bones and articulated bodies into the nearest fissures. Some of the bodies would float due to gases in the decaying body produced by bacteria. These floating bodies could take bones deep into the subaerial fissure cavities, later sinking to produce the articulated remains, but others, perhaps more decayed, would break up under the forces of hydraulic transport. Depending on where they came to rest in the fissures, some of the bodies would decay in anoxic conditions and others would decompose in oxygen-rich waters. The individual bones may have been further transported and sorted before their final burial. This would result in the black, brown and white bones that are sometimes found in the same rock (see fig. 5z for a variety of coloured bones in one small sample).
The evidence that forest fires were numerous in the vegetation above fissures comes from two sources. Harris (Reference Harris1957) reports large quantities of fusain, fossil charcoal, from Cnap Twt, Ewenny and Pont Alun on St Bride's palaeo-island. Harris (Reference Harris1958) argued that seasonal storms resulted in forest fires that could kill the tetrapods without necessarily destroying them by burning. The heavy rainfall that followed would wash the dead animals, including Morganucodon and Gephyrosaurus, into the fissures. While accepting the notion of forest fire as a cause of death of the tetrapods was reasonable, Evans & Kermack (Reference Evans, Kermack, Fraser and Sues1994) suggested that the absence of invertebrates (excepting carbonized fragments of the beetle Metacupes harrisi Gardiner, Reference Gardiner1961, and the gastropod Natica oppeli recorded by Kermack, Mussett & Rigney, 1973) was not explained by the wildfire theory. We recognize that many insects would have been able to fly away from the fire, and others do not have the hard beetle elytra that would preferentially fossilize. Insect remains are likely to have been very small and fragile and will have been easily fragmented. The absence of abundant insect remains does not seem to us a significant impediment to the forest fire theory which otherwise explains the origination of the tetrapod fossils so well.
The second source of evidence comes from the semi-fusain and inertinite that we have discovered in the palynomorph-bearing fissures at Tytherington. Fossil charcoal has been discovered in fissures 1, 2 (see Fig. 5b), 6, 9 and particularly in relative abundance in 13 (FP13.1), which points to the occurrence of frequent wildfires. Therefore, there is clear evidence for forest fires in both sauropsid and mammal-bearing fissures that provides a plausible explanation for the death of the tetrapods.
14. Summary
Using evidence from well-preserved palynological samples collected from six fissures at Tytherington, we have concluded that the Tytherington sauropsid fissures were infilled by sediment in the Rhaetian at a time equivalent to the Westbury Formation, Penarth Group. This is supported by the finding of Penarth Group marine vertebrates both in the palyniferous and other fissures in the quarry. Moreover, using comprehensive and extensive evidence from the palynological samples, associated terrestrial reptiles, marine and non-marine fish and a glaucontic clay-mineral, we also conclude that the fissures were infilled in a marginal land/marine environment on a small limestone island. We have shown that there are Quaternary, including modern, analogues consistent with this palaeoenvironmental model.
The Tytherington sauropsid fissure reptiles are therefore representative of an island herpetofauna and form part of a pattern of Late Triassic/Early Jurassic island archipelago faunas known from the Bristol region and South Wales. We suggest that the evidence found in the fissures favours a forest fire cause for the death of the tetrapods and that their bones were transported to final burial by fluvial processes. Transport of many of the bones could be short and the bones would generally have been buried under sea level. We have shown that the insular model for the tetrapods found in these fissures accounts for the high abundance of lepidosaurs, the presence of small archosaurs and the absence of larger reptiles.
From a geomorphological analysis of cavern formation and the doline fissure we conclude that the Tytherington reptile-bearing fissures described in this paper were mainly formed during Rhaetian times. Although it is likely that there was some pre-Rhaetian solution of joints and cracks, there is no unequivocal evidence that supports the conclusion of substantial cave formation prior to the Rhaetian. Indeed the evidence is overwhelmingly against this notion. From our evidence, the time between formation and infilling of the fissures is not likely to be long, and this is in accord with the same conclusion for Mesozoic Mendip fissures made by Wall & Jenkyns (Reference Wall and Jenkyns2004). The model of a limestone island surrounded by marine waters has many modern analogues where mixed saline/freshwater regimens produce enlarged solution cavities similar to those at Tytherington. The palaeo-dolines at Tytherington are equivalent to the ‘banana holes’ on Caribbean islands. The continued initiation and development of fissures would also be expected at any time of emergence until the fully marine transgression in the Hettangian and Sinemurian. In this respect, one of the doline fissures (fissure 12) at Tytherington may have formed and been infilled in a timescale encompassing the later Rhaetian before inundation.
Acknowledgements
DIW would like to acknowledge the late Professor R. J. G. Savage for his strong and kindly guidance and advice for much of the work described here that derived from DIW's thesis. Dr C. J. Duffin aided in the identification of the marine fish; the late Dr Colin Patterson confirmed identification of the Pholidophorus specimens and Dr Giles Miller gave advice on the conodonts. Dr Geoff Warrington gave us continued encouragement to complete this study. We have had useful discussions with Professor John Mylroie, Charles Copp, Drs Paul Barrett, Jim Carew, Pete Smart, Fiona Whittaker and the late Rob Palmer. We thank Barry Marsh for essential photography and Shir Akbari for preparation of the palynological samples and operating the elemental analyser. Steve Kearl, Lucy Cliff and Duncan Wright made preliminary contributions to the Hampstead Farm Quarry data. David Ward is warmly thanked for his preparation of some of the vertebrate material figured and for stimulating discussions. We also acknowledge an essential discussion with Stephen Hesselbo as to the significance of the fissures and the Triassic–Jurassic mass extinction. Personal communications with Professors Susan Evans and Mike Benton and Dr Nick Fraser contributed valued evidence to this work. We appreciate the patience and comments given by our two anonymous referees. Finally we gratefully acknowledge the continued assistance of Mike Curtis for revealing the existence of the Tytherington fissure system, permitting access to Tytherington Quarry and sharing much hard-won information on the British Rhaetic and its vertebrate fauna. We wish to also extend our particular appreciation to the Amey Roadstone Corporation (now Hanson), who always permitted access to the fissures and aided in the transport of the original find of fissure material to the University of Bristol. DIW acknowledges the support of a University of Bristol Science Faculty Postgraduate Scholarship while researching some of the work presented here.