



Introduction
At the end of 2019, a total of 41 cases of pneumonia of unknown aetiology were first reported and then spread rapidly throughout the world (Ref. Reference Lu1). Further studies identified this novel zoonotic virus as an enveloped, positive-sense single-stranded RNA coronavirus belonging to the subgenus Sarbecovirus of the genus Betacoronavirus (Ref. Reference Wang2). It is named severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), and the disease caused by SARS-CoV-2 was termed corona virus disease 2019 (COVID-19) by the World Health Organization (WHO). On 31 January 2020, the WHO announced that COVID-19 is listed as Public Health Emergency of International Concern. By the end of July 2021, SARS-CoV-2 has spread rapidly and affected more than 200 countries, resulting in more than 230 million identified cases and 4.7 million confirmed deaths (Ref. 3). SARS-CoV-2 is the seventh coronavirus known to infect humans, and the third coronavirus emerged as a public health issue over the past two decades. SARS-CoV-1, MERS-CoV and SARS-CoV-2 can cause serious illness, whereas HKU1, NL63, OC43 and 229E are associated with mild symptoms (Ref. Reference Andersen4). In a previous study of 44 672 patients with COVID-19 in China, 81% of infected patients had a broad spectrum of clinical manifestations ranging from asymptomatic to cough, fever, coagulation dysfunction and metabolic acidosis. A total of 14% had severe manifestations, and 5% had critical manifestations, such as hyper-inflammation, multiple organ dysfunction syndromes and acute respiratory distress syndrome (ARDS) (Ref. 5). Most mortalities happened in elderly patients or patients with multiple comorbidities, including cardiovascular diseases, respiratory diseases, diabetes mellitus, hypertension and immune-compromised patients, such as cancer (Ref. Reference Jung6).
The pathogenesis of COVID-19 is thought to be determined by two main courses. In the early phase of infection, SARS-CoV-2 identifies host cell angiotensin-converting enzyme 2 (ACE2) receptors and invades the host cell to complete the replication cycle. The cellular damage caused during the replication cycle can lead to respiratory disease, a decrease in lymphocytes and local/systemic inflammation. As the infection progresses, immune pathologies such as cytokine storm and lymphopoenia occur (Ref. Reference Siddiqi and Mehra7).
There is currently no specific treatment available. Treatment strategies for COVID-19 infection are early detections, the quarantine of new cases and supportive therapies for the confirmed individual (Ref. 8). Immunotherapy plays an important role in inhibiting viral infection or modifying the overactivated immune response against SARS-CoV-2. In this review, we summarise the immunopathogenic features of COVID-19 and highlight current advances in immunotherapies to combat COVID-19, hoping to enlighten ideas of developing new potential therapies in the future.
Viral entry and replication
Epidemiological and virological studies suggest that SAR-CoV-2 is transmitted to others mainly by symptomatic and asymptomatic individuals through close contact via respiratory droplets or direct contact with infected individuals, sometimes through contaminated objects and surfaces (Ref. 8). The spike (S) protein of SARS-CoV-2 determines the tropism of receptors in host cells and plays a vital role in the invasion process. Figure 1 summarises the recognition and replication cycle of SARS-CoV-2. The S protein is composed of two subunits: S1 and S2. S1 is responsible for the binding of the viral receptor-binding domain (RBD) to the host cell ACE2, and S2 ensures the fusion of the virus with the host cell membrane (Ref. Reference Wu9). SARS-CoV-2 employs the cellular transmembrane serine protease 2 (TMPRSS2) for S protein priming (Ref. Reference Hoffmann10). Most recently, neuropilin 1 has been identified as an important cofactor for entry, particularly in cells with low-level ACE2 expression (Ref. Reference Cantuti-Castelvetri11). Studies also show that the interaction of the S protein with the CD26 and CD209L could be a possible way of viral entry, but the mechanism is still unclear (Refs Reference Vankadari and Wilce12, Reference Amraei13). During the fusion process, binding of the virus to ACE2 causes stabilisation of the RBD, which triggers conformational changes in the S-complex, leading to the release of the S1 subunit and activation of the fusogenic activity of S2 (Ref. Reference Song14). A study demonstrated that strains isolated from COVID-19 patients could use the host cell protease TMPRSS2 and cathepsins B/L to prime the S protein in vitro (Ref. Reference Prasad15). Structural changes in the S protein may prevent the immune system from recognising the virus. For example, the current delta variant has three mutations at the S1 subunits, which increases the affinity of RBD to bind to ACE2 and enhances its ability to escape our immune system (Ref. Reference Khateeb16).
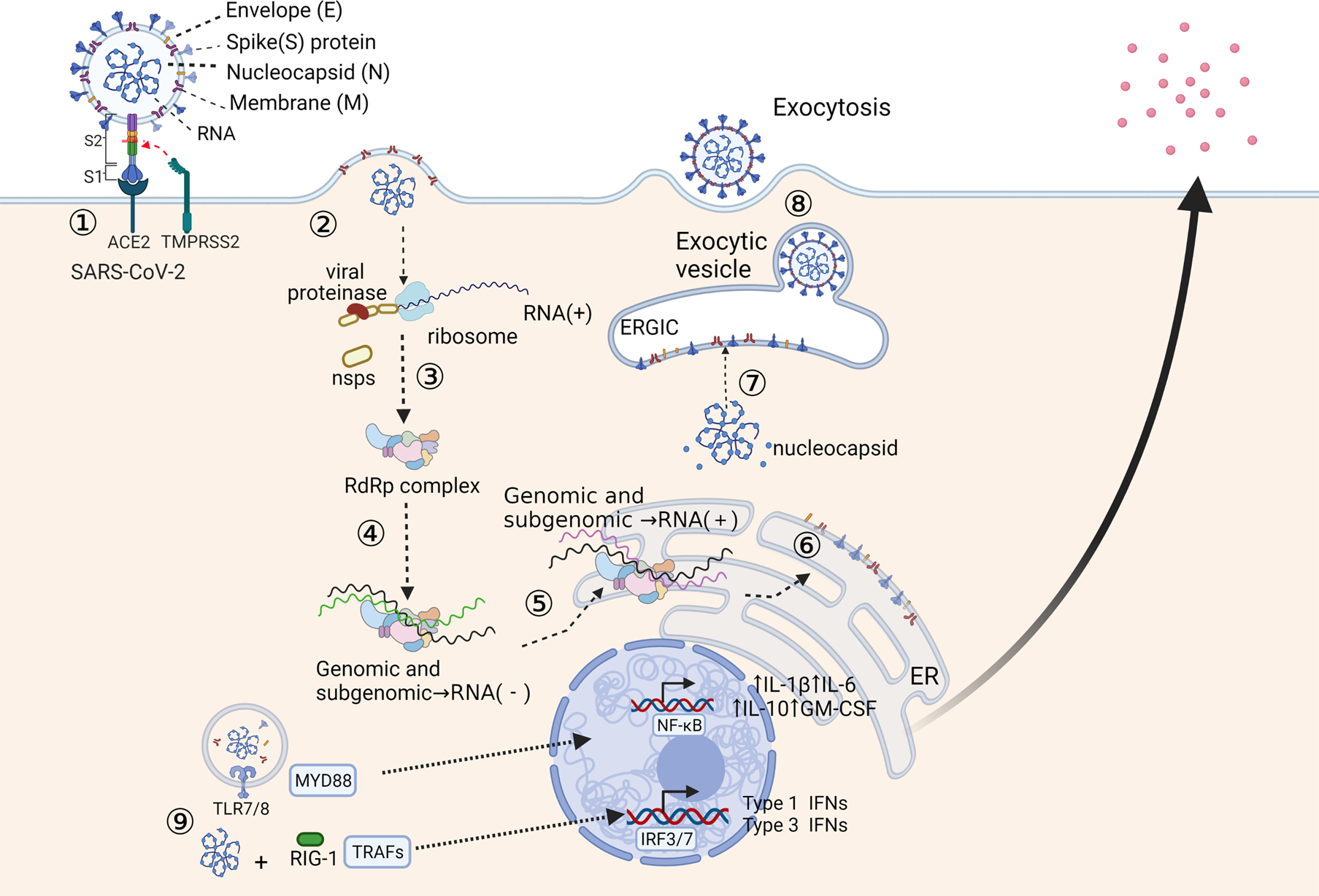
Fig. 1. Recognition and replication cycle of SARS-CoV-2. (1) The SARS-CoV-2 engages the host cell surface through the binding of spike (S) protein to ACE2. TMPRSS2 cleaves the S protein into S1 and S2 subunits, and the S2 subunit ensures the fusion of the virus with the host cell membrane. (2) The host cell ribosome translates the positive-sense RNA genome into polyproteins, which are cleaved by viral proteases to produce viral nonstructural proteins (nsps). (3) The nsps subsequently assembled into RNA-dependent RNA polymerase complex (RdRp). (4) RdRp uses the positive-sense RNA genome as a template to generate the negative-sense sub-genome. (5) Sub-genomes are used as templates for synthesising full-length positive-sense RNA genomes and sub-genomic mRNAs. (6) The sub-genomic mRNAs are translated into the endoplasmic reticulum (ER) to generate structural and accessory proteins. (7) The assembly of newly generated positive-sense RNA genome and viral structural proteins occurs in the endoplasmic-reticulum–Golgi intermediate compartment (ERGIC). (8) The enveloped virions are released through exocytosis. (9) In recognition of the SARS-CoV-2 RNA genome, TLR7/8 and RIG-1 recruit MYD88 and TRAFs. These adaptor molecules translocate transcriptions factors, NF-κB and IRFs into the nucleus leading to the production of and secretion of inflammatory cytokines.
ACE2 is an enzyme that belongs to the renin–angiotensin system and is located on the cell surface of type II alveolar epithelial cells in the lungs and other tissues (Ref. Reference South17). ACE2 plays a critical role in controlling vasoactive effects in the body and is highly co-expressed with genes related to TMPRSS in the upper airways (Refs Reference Zhang18, Reference Wruck and Adjaye19). The affinity of SARS-CoV-2 to ACE2 is 10–20 times higher than that of SARS-CoV, possibly because of the RBD of SARS-CoV-2 having a more compact conformation (Ref. Reference Wrapp20). Several residue changes in the RBD of SARS-CoV-2 stabilise two virus-binding hotspots at the RBD–ACE2 interface and increase ACE2 binding affinity (Ref. Reference Shang21).
Once the virus enters the host cell cytoplasm, translation begins using viral RNA as a template to generate virus-specific mRNAs. After the translation of virus-specific mRNAs is completed, the production of structural and nonstructural viral proteins is incorporated into the endoplasmic reticulum or Golgi membrane (Ref. Reference V'Kovski22). Finally, the vesicles containing the virus particles fuse with the plasma membrane to release the virus (Ref. Reference Li23). Another possible mechanism for SARS-CoV-2 entry is that the virus binds to its corresponding antibody to form an antigen–antibody complex and then enters the target cell through the Fc receptor (Ref. Reference Karthik24).
Innate and adaptive immunity against SARS-COV-2
Activated natural killer (NK) cells exert a major histocompatibility complex-independent immune response against SARS-CoV-2 and may limit viral pathogenesis in the early stages of infection (Ref. Reference Florindo25). The respiratory tract is the first front line of the immune system, which contains many immune effector cells such as epithelial cells, type 1 macrophages (M1) and dendritic cells (DCs). After infection, epithelial cells produce immune cytokines and express adhesive molecules, recruiting immune cells to the lung tissue to combat the virus. After virus recognition, activation of interferon regulatory factor 3 (IRF-3) in M1 leads to the production of various immune cytokines such as type I interferon (IFN-I) and phagocytosis (Ref. Reference Vabret26). IFN-I plays an essential role in the immune response against SARS-CoV. One of the mechanisms of anti-viral immunity by IFN-I is activating IFN-induced transmembrane family proteins, which inhibit virus entry into the host cell. DCs can serve as the antigen-presenting cell (APC)-mediated immune response to SARS-CoV-2 and produce IFN-I and interleukin 6 (IL-6) to inhibit viral infection. A study shows that excessive production of IFN-I by DCs leads to severe inflammation and ARDS in severe patients (Ref. Reference Chiappelli27). The APCs, such as monocytes and DCs, distinguish the viral antigen on infected cells, introduce the antigen to the helper T (Th) cells, and produce cytokines that direct the anti-viral immune response of the T cells. Serum levels of Th1-associated cytokines, including IFN-γ, tumour necrosis factor (TNF)-α and IL-2, are reported to be elevated in COVID-19 patients (Ref. Reference Liu28). These cytokines activate cytotoxic T cells (CTLs) to attack the virus-infected cells and destroy them by producing perforin and granzymes. In addition, Th2 cells present the viral antigen to B lymphocytes, which subsequently produce neutralising antibodies against the S protein of the virus. The neutralising antibodies inhibit the replication of the virus in the body and generate humoral immunity, which is one of the most important concepts for developing vaccines against SARS-CoV-2 (Ref. Reference Li23). Figure 2 briefly summarises the immunopathology of COVID-19.
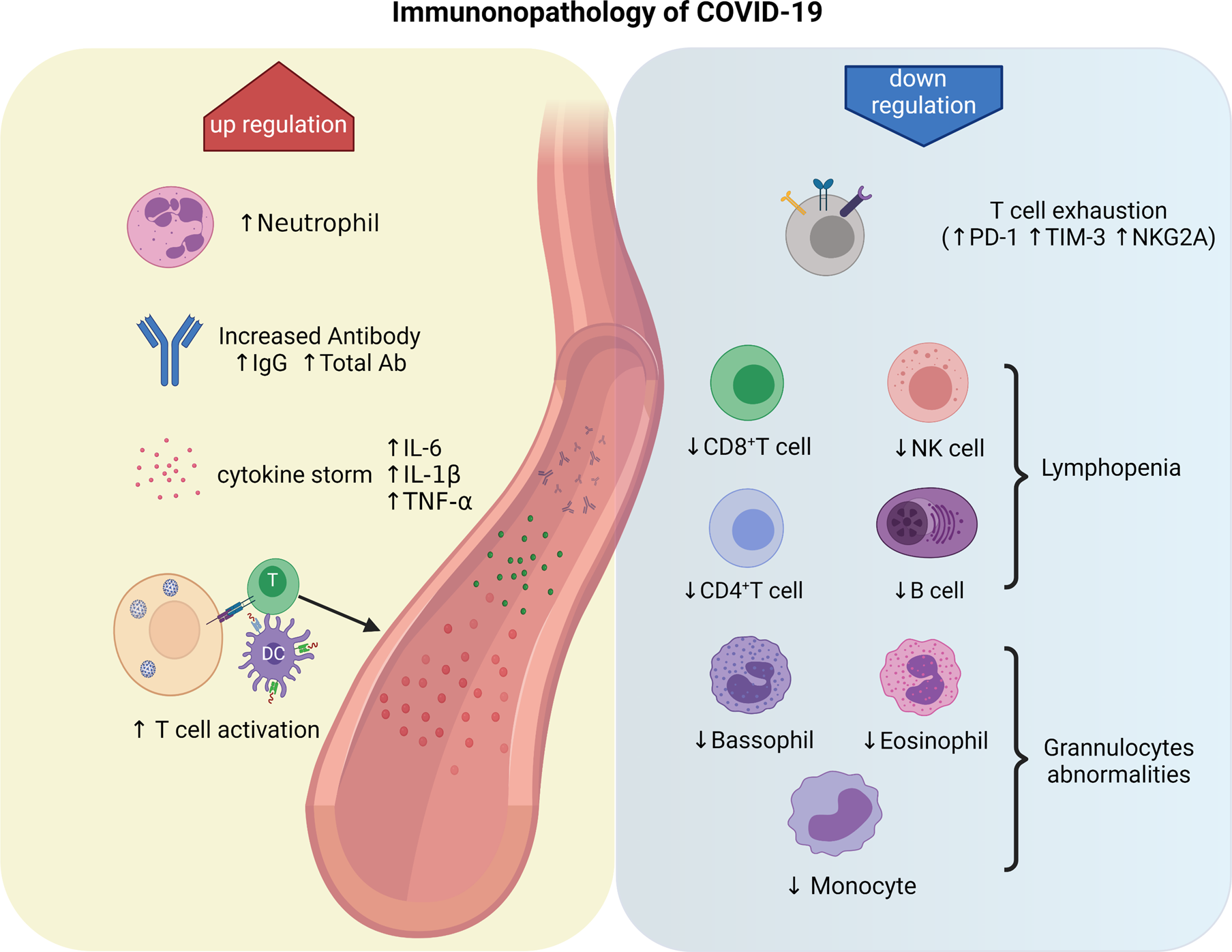
Fig. 2. Immunopathology of COVID-19. The neutrophil is significantly increased in severe cases, despite decreased eosinophil, basophil and monocyte counts. Activated B cells produce and secrete a large number of antibodies to neutralise the virus. The antigen-specific T cells are activated, and numerous cytokines, such as IL-6, IL-1β and TNF-α are secreted, leading to cytokine storm. On the other hand, lymphopoenia is an important feature of COVID-19, mainly manifested by a decreased count of CD8+ T cells, CD4+ T cells, NK cells and B cells. The increased expression of programmed cell death protein-1 (PD-1), T cell immunoglobulin domain and mucin domain-3 (TIM3), and killer cell lectin-like receptor subfamily C member 1 (NKG2A) on T cells could accelerate lymphopoenia.
Humoral immunity plays a vital role in the induction of adaptive immunity to coronaviruses. Activation of B cells and plasma cells leads to the production of neutralising antibodies that prevent further infection with the virus. The seroconversion time of Ab and immunoglobulin M (IgM) and IgG antibodies appeared consecutively, with a median seroconversion day of 11, 12 and 14, respectively (Ref. Reference Zhao29). Within 4 weeks of infection, 90–99% of individuals infected with the SARS-CoV-2 virus develop detectable neutralising antibodies (Ref. 30). The RBD domain of the SARS-CoV-2 S protein is the primary target of these viral species-specific neutralising antibodies (Ref. Reference Ju31). Appropriate anti-viral antibodies can prevent patients from reinfecting the virus. Some cases of reinfection because of insufficient serum antibody levels have been reported, questioning the humoral immunity of SARS-CoV-2 (Ref. Reference Zhou32). Further research is needed to understand the exact mechanism of the immune response to SARS-CoV-2.
Cytokine storm
As illustrated in Figure 3, cytokine storm is a state of uncontrolled systemic hyper inflammation caused by an excess of cytokines, leading to multiorgan failure and even death. Rather than direct cytopathic effects of the COVID-19, cytokine storm activates inflammatory immune cells to attack alveolar cells, causing lung tissue damage (Ref. Reference Wiersinga33). The inflamed lung tissue also stimulates fibroblast cells to form fibrotic tissue in the lungs. In addition, the leakage of fluid into the alveoli and the accumulation of the inflammatory exudate lead to the formation of hyaloid membranes and further ARDS (Ref. Reference Wang34). Damaged lung cells and M1 produce chemotactic factors that attract other immune cells and trigger an uncontrolled inflammatory response in the lung. Then the inflammatory cells begin an uncontrolled production of pro-inflammatory cytokines and chemokines contribute to the cytokine storm, including IL-1, IL-6 (Ref. Reference Conti35), IL-8, IL-10, TNF-α, IFN-γ, granulocyte-macrophage colony-stimulating factor (GM-CSF), CXCL-1, -3, -10 and CCL-2, -3 (Refs Reference Chua36–Reference Huang38). Increased IL-6 and IL-8 lead to severe inflammation in critical patients (Ref. Reference Vabret26).

Fig. 3. Schematic diagram of cytokine storm in COVID-19. (1) SARS-CoV-2 infects lung bronchial, alveolar type I and type II epithelial cells. (2) Immune cells, including macrophages, recognise viruses and produce cytokines. (3) Cytokines attract more immune cells, such as neutrophils, T cells, monocyte and DCs, which in turn produce more cytokines, creating a cycle of inflammation that damages the lung cells. (4) Persistent inflammation lead to tissue damage, leucocytes infiltration and fluid leakage from the blood vessel, causing respiratory failure. (5) Excessive deposit of fibrin in the lungs.
Currently, one article introduces the concept of dividing the cytokine storm into two phases: the first phase is a temporary state of immunodeficiency, and the second phase is an overactive state of the immune system to compensate for the failure of target clearance, resulting in a cytokine storm (Ref. Reference McGonagle39). Another report described the effects of human coronavirus on cytokines, noting delayed secretion of types I and III IFN in the early phase (within 3 days) of infection and excessive secretion of pro-inflammatory cytokines in the later phase (10–14 days) (Ref. Reference Blanco-Melo40). A study highlights a low level of IFN-I activity and downregulation of IFN-stimulated genes in COVID-19 patients and hyper-inflammatory responses represented by IL-6 and TNF-α (Ref. Reference Hadjadj41). These studies underscore that the cytokine storm results from the failure of the initial response to the type I and III IFNs of SARS-CoV-2, leading to the delayed response of the immune system the inability to clear the virus.
According to the pathogenesis of cytokine storm, appropriate treatments such as IL-1, IL-6 and TNF-α inhibitors can reduce lung tissue damage and lead to a better outcome in COVID-19 patients (Ref. Reference Ye42). Several strategies, such as injection of immunomodulatory drugs and mesenchymal stem cells (MSCs), have been used to prevent lung injury and multiple organ failure in severe patients with cytokine storm. Targeting inflammatory cytokines could also reduce the severity of cytokine storm.
Lymphopoenia, granulocytes and monocytes disorders
Lymphopoenia, the decreased number of lymphocytes in the peripheral blood, usually accompanied by an increased number of neutrophils, is one of the most important features and common symptoms in severe cases of COVID-19 (Ref. Reference Huang38). The most common cause for lymphopoenia could be apoptosis/pyroptosis of lymphocytes triggered by SARS-CoV-2 infection (Ref. Reference Tay43). Other reasons include increased production of glucocorticoids, a storm of cytokines (Ref. Reference Liao44), weakened vascular cell adhesion (Refs Reference Chen45, Reference Yao46) and FAS–FASL interaction (Ref. Reference Bellesi47). Lymphopoenia results in decreased numbers of CD8+ T cells, CD4+ T cells, B cells or NK cells and weakens patients' immunity to the virus (Refs Reference Liu28, Reference Wang48). A remarkably decreased T cell is almost always observed in severe cases, and patients admitted to the intensive care unit show significantly decreased T cells, especially CD8+ T cells (Ref. Reference Tavakolpour49). Lymphopoenia may serve as a prognostic marker for COVID-19 (Ref. Reference Huang50). Since the number of neutrophils is increased, the neutrophil-to-lymphocyte ratio can be used as a prognostic criterion for COVID-19 (Ref. Reference Zhang51). In addition to lymphopoenia, abnormalities in granulocytes and monocytes have also been observed in COVID-19 patients (Ref. Reference Zhang51).
Abnormalities in granulocytes and monocytes have also been observed in COVID-19 patients (Fig. 2). The number of neutrophils is significantly higher in severe patients than in non-severe patients (Ref. Reference Lippi and Plebani52). At the same time, a lower percentage of eosinophils, basophils and monocytes was observed in the serve patients (Ref. Reference Qin53). The mechanisms are currently unclear, and further research is needed.
Immunotherapy of COVID-19
Currently, there is no proven specific treatment modality for COVID-19. Initial treatments are mainly supportive, including oxygen therapy, ventilator support in patients with respiratory failure, antibiotics to prevent secondary bacterial infections, anti-inflammatories and fluid management (Ref. Reference Xu54). Since the COVID-19 outbreak, scientists have begun to evaluate the impact of currently available anti-viral drugs on the virus. Because the SARS-CoV-2 is an RNA virus, adenosine analogues such as remdesivir can block the viral RNA synthesis process. Remdesivir has shown clinical improvement and reduced mortality (Ref. Reference Wilt55). Other nucleoside analogues, such as ribavirin, are together with IFN beta-1b, lopinavir–ritonavir through a triple therapy in phase 2 clinical trial of treating COVID-19 (Ref. Reference Hung56).
Immunotherapy has achieved remarkable results in the treatment of many diseases such as cancer and viral infections. The use of immune-enhancing agents to boost and strengthen the immune system could have a positive impact on the treatment of COVID-19. Figure 4 summarises the immunotherapy of COVID-19, which will be discussed step by step below. The clinical trials of immunotherapy for COVID-19 are also summarised (Table 1).
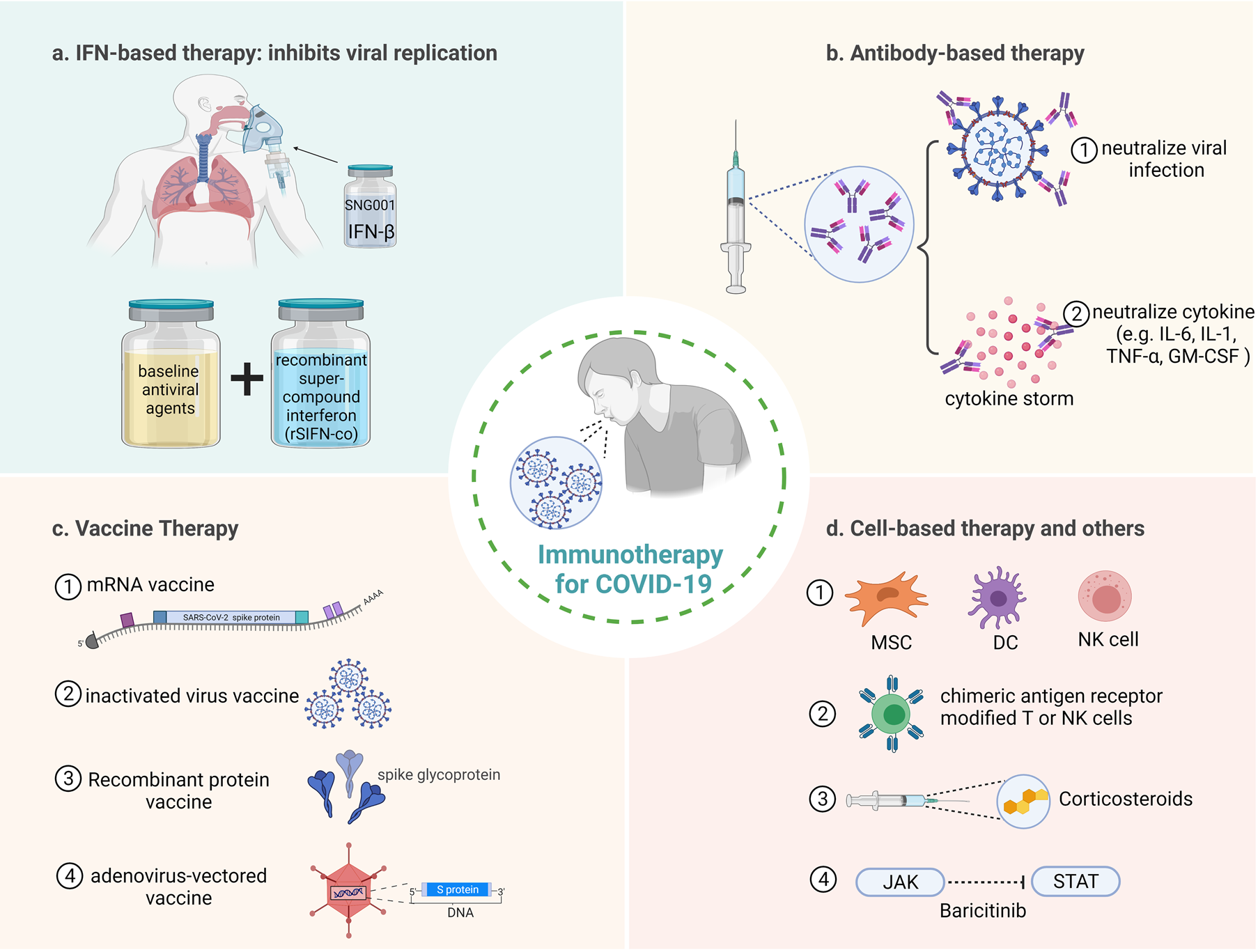
Fig. 4. Immunotherapeutic approaches for COVID-19. (a) IFN is used to inhibit virus replication, the therapy of inhaled nebulised IFN-β1a (SNG001) and baseline anti-viral agents combined with recombinant super-compound interferon (rSIFN-co) have better targeted therapeutic effect. (b) Antibody-based therapy, including (1) neutralise viral infection and (2) neutralise cytokines to prevent damage caused by cytokine storm. (c) Vaccines used to treat COVID-19, including (1) mRNA vaccine, (2) inactivated virus vaccine, (3) recombinant protein vaccine and (4) adenovirus-vectored vaccine. (d) Cell-based therapies and other treatments, including (1) therapies based on MSC, DC and NK cells, (2) CAR-modified T and NK cells have better targeted therapeutic effect, (3) corticosteroids injections act as anti-inflammatory agents and (4) blocking JAK-STAT signalling pathways to inhibit cytokine storms.
Table 1. Summary of the clinical trials of immunotherapy for COVID-19 (www.clinicaltrails.com)

NA, not available.
IFN therapy
IFN plays an essential role in host defense against SARS-CoV-2 infection. Upon viral infection, the endogenous pattern recognition receptor recognises the genetic material of the virus, which triggers a series of downstream cascade responses such as the JAK-STAT signal and stimulates the production of IFN-stimulating genes (ISGs) and IFN-I and other products (Ref. Reference Mazewski57). The downstream molecules controlled by ISGs and IFN-I can directly inhibit viral replication, recruit and activate immune cells to regulate viral infection (Refs Reference Mesev58, Reference Ribero59). It was found that binding of the SARS-CoV-2 protein nonstructural protein 1 (NSP1) to the ribosomal subunits can block the induction of IFN and ISG expression (Ref. Reference Thoms60). TBK1 can induce TLR3–IFN and TMEM173/STING-IFN signalling pathway (Ref. Reference Liyana61), and NSP13 can block the phosphorylation of TBK1 and inhibit signal transduction, which affects the production of IFN (Ref. Reference Xia62). IFN-I signal transduction can be antagonised by blocking signal transduction or blocking IRF-3 nuclear translocation (Ref. Reference Fung63).
IFN-I and IFN-III are particularly important for the anti-viral response. They express distinct homologous receptors: IFN-I receptors are expressed in most mammalian tissues, whereas IFN-III receptors are mainly found in the respiratory and gastrointestinal tracts (Refs Reference Mordstein64, Reference Lazear65). The delayed IFN response during SARS infection can cause the accumulation of various immune cells and the appearance of cytokine storm, leading to more severe pathological damage in the lung (Ref. Reference Subbian and Ramasamy66). Early administration of IFN-I will forcibly eliminate the virus (Ref. Reference Subbian67). The use of inhaled nebulised IFN-β1a (SNG001) can more effectively deliver the drug to the site of SARS-CoV-2 infection for better treatment efficacy (Ref. Reference Pdm68). IFN combination therapy can activate STAT1 to inhibit viral replication (Refs Reference Zhang69, Reference Nezhad70). Recombinant super-composite IFN (rSIFN-co) combined with baseline anti-viral therapy was found to have better clinical improvements and fewer side effects than conventional IFN anti-viral therapeutics (Ref. Reference Li71). IFN treatment also has side effects, the most common of which are flu-like symptoms, haematological toxicity, elevated transaminases, nausea, fatigue and psychiatric sequelae (Ref. Reference Van Gool72).
Antibody-based therapies
Seow et al. found that the kinetics of the neutralising antibody response against SARS-CoV-2 infection is typical of acute viral infection, with decreasing neutralising antibody titres after an initial peak (average 23.1 days post-onset of symptoms), and that the magnitude of this peak depends on disease severity (Ref. Reference Seow73). Antibody-based therapies can improve the immune response of COVID-19 patients and inhibit SARS-CoV-2 infection.
Studies have shown that the early use of convalescent plasma therapy can improve the prognosis of patients with moderate to severe COVID-19, but no significant clinical improvement was observed with the late use of convalescent plasma (Ref. Reference Briggs74). The mortality of COVID-19 patients was significantly reduced when convalescent plasma was provided in the early stages of the disease, and the use of plasma with high antibody titres did not increase the risk of death compared with unselected plasma (Refs Reference Candia75, Reference Cross76). The use of convalescent plasma not only transiently increased the level of systemic anti-SARS-CoV antibodies but also promoted the specific T-cell response (Ref. Reference Acosta-Ampudia77).
Monoclonal antibodies are used to prevent and treat various diseases, and treatment with monoclonal antibodies against coronavirus is clinically effective (Ref. Reference Blake78). REGEN-COV, a combination of two monoclonal antibodies, casirivimab and imdevimab, both specifically targeting the spike protein of SARS-CoV-2, has been shown to reduce hospitalisation rates or risk of death in individuals at high risk for COVID-19. Subcutaneous injection of REGEN-COV can prevent SARS-CoV-2 infection and reduce the duration of disease symptoms and high viral load (Ref. Reference O'Brien79). Llama-derived single-domain antibodies, also known as nanobodies, are generally more heat stable, accessible and cheaper to produce and can be better used for protein engineering than conventional antibodies. Combining nanobodies with different epitopes or complementary epitopes increases resistance to mutant escape (Ref. Reference Koenig80).
Intravenous immunoglobulin (IVIG) is widely used as an alternative therapy to treat immunodeficiency and reduce inflammatory responses (Ref. Reference Rojas81). The use of high-dose IVIG in hospitalised COVID-19 patients may improve their clinical progress by regulating their immune status. For example, IVIG may reduce the excessive inflammation observed in individuals infected with SARS-CoV-2 by reducing the levels of cytokines, chemokines and some complement factors (C5a) involved in immune activation and cell migration (Ref. Reference Concepción82). A 5% anti-COVID-19 IVIG solution contains 90% immunoglobulins (IgG, IgM and IgA), and the total anti-SARS CoV-2 antibody titre is three times higher than pooled convalescent plasma (Ref. Reference Ali83).
Vaccine therapy
The first COVID-19 vaccine urgently approved by the U.S. FDA was BNT162b2, an mRNA encoding S protein, which showed 95% efficacy (Ref. Reference Chagla84). The second vaccine, mRNA-1273, had almost the same efficacy as BNT162b2, and it remains to be investigated whether BNT162b2 can elicit a long-term anti-SARS-CoV-2 immune response. The third vaccine, CVnCoV/CV2CoV, has been shown to prevent disease and death caused by the VOCB.1.351 mutant in mouse models, and clinical trials are underway (Ref. Reference Hoffmann85). The antibody responses to SARS-CoV-2 mRNA vaccination comprise a large proportion of non-neutralising antibodies and are co-dominated by S protein N-terminal domain (NTD) and RBD antibodies. Thus, the NTD portion of the spike represents a vital vaccine target (Ref. Reference Amanat86). DNA vaccine can induce antibodies with robust neutralising ability against SARS-CoV-2 wild-type strains and against some mutants, which could open a new avenue for effective treatment and diagnosis at the point of care (Ref. Reference Chen87). The new SARS-CoV-2 mutant B.1.526 is spreading at an alarming rate and was identified by a D614G mutation and four novel point mutations in the S protein. The results show that vaccine-induced antibodies are protective against the B.1.526 mutant in the current recovery phase (Ref. Reference Zhou88).
Protein vaccines require adjuvants to enhance the immune response. Novavax's vaccine uses a genetically modified full-length S protein (NVX-CoV2373) that has higher structural stability. When administered to volunteers, the NVX-CoV2373 vaccine has a protective efficacy of 89.7% against SARS-CoV-2 infection and shows effective protection against the B.1.1.7 variant (Ref. Reference Heath89). Studies have shown that the neutralising antibody response elicited by the Ad5-nCoV vaccine against COVID-19 with two doses of the atomised adenovirus type 5 vector is similar to that elicited by one dose of the intramuscular injection, and nebulisation at 28 days after the first intramuscular injection enhanced the vaccination and induced a strong IgG and neutralising antibody response (Ref. Reference Wu90). Table 2 lists the COVID-19 vaccines within WHO Emergency Use Listing (EUL) and prequalification (PQ) evaluation process.
Table 2. COVID-19 vaccines within WHO emergency use listing (EUL) and prequalification (PQ) evaluation process

NRA, National Regulatory Authority; USFDA, The United States Food and Drug Administration; EMA, European Medicines Agency; MFDS, Ministry of Food and Drug Safety; TGA, Therapeutic Goods Administration; DCGI, Drugs Controller General of India; NMPA, National Medical Products Administration of CHINA; CECMED, Centro para el Control Estatal de Medicamentos, Equipos y Dispositivos Médicos, Cuba.
The humoral and cellular immune responses elicited by the different vaccines differed considerably. In particular, inactivated vaccines showed relatively low levels of neutralising antibodies and T-cell responses. Compared with three consecutive doses of inactivated vaccines to enhance the immune response, the use of recombinant subunits, adenoviral vectors or mRNA vaccines after two doses of inactivated vaccines can further boost neutralising antibodies and specific Th1 cell responses (Ref. Reference Zhang91). Modified vaccinia Ankara (MVA) vectors expressing membrane-anchored prefusion-stabilised spike (MVA/S) but not secreted S1 induce strong neutralising antibody responses against SARS-CoV-2 in mice. MVA/S vaccination triggered potent neutralising antibodies and CD8+ T-cell responses in macaques and protected them from SARS-CoV-2 infection, demonstrating that MVA/S is a potential vaccine candidate (Ref. Reference Routhu92).
Management of cytokine storm
Higher serum IL-6 levels are associated with the severity of COVID-19 (Ref. Reference Chen L93). Therefore, blocking IL-6 could reduce COVID-19 inflammation. Tocilizumab and sarilumab are two FDA-approved IL-6 receptor antibodies investigated in COVID-19 patients in phase III clinical trials (Ref. 94). Tocilizumab and sarilumab improved the survival of COVID-19 patients (Ref. Reference Investigators95). Administration of two doses of tocilizumab (8 mg/kg intravenously) per day reduced C-reactive protein (CRP) and cytokine storm in a man with renal carcinoma who suffered from COVID-19 (Ref. Reference Liu96). In some cases, adverse effects such as acute pancreatitis, cytopenias, elevated ferritin and lactate dehydrogenase occur with the use of tocilizumab in COVID-19 patients (Ref. Reference Morrison A97). IL-1β is a pro-inflammatory cytokine that plays an important role in the progression of airway inflammation in many viral infections (Ref. Reference Martinon98). In response to alveolar infection by SARS-CoV-2, macrophages secrete IL-1β, which induces fever and leads to fibrosis in the lungs (Ref. Reference Conti35). Inhibition of IL-1β may be a practical approach to reduce cytokine release syndrome and ARDS progression, as higher levels of IL-1β are associated with the severity of COVID-19 infection in critically ill patients (Ref. Reference Price99).
TNF-α is an inflammatory cytokine that triggers leucocyte recruitment to the site of infection in response to SARS-CoV-2 (Ref. Reference Price99). IFN-γ is a vital biomarker promoting the pathogenesis of ARDS and acute lung injury. The synergistic effect of TNF-α and IFN-γ leads to inflammatory cell death, tissue damage and mortality in SARS-CoV-2 infection (Ref. Reference Karki100). Therefore, inhibition of TNF-α and IFN-γ is a promising immunotherapy method to inhibit ARDS progression in severe COVID-19 patients. For example, XPro1595, a soluble TNF-α-neutralising protein that inhibits the interaction between soluble TNF-α and its receptor, is currently evaluated in a clinical trial to treat COVID-19 (NCT04370236). Infliximab, an approved mouse/human chimeric monoclonal antibody, is undergoing clinical trials to evaluate its therapeutic effect on COVID-19 (NCT04922827, NCT04425538 and NCT04593940). GM-CSF is an immunoregulatory cytokine that plays a central role in initiating and maintaining inflammatory diseases. Several clinical trials have considered the use of otilimab (NCT04376684), lenzilumab (NCT04351152), gimsilumab (NCT04351243), mavrilimumab (NCT04492514), TJ003234 (NCT04341116) and sargramostim (NCT04707664) to block GM-CSF as potential treatments for COVID-19.
The complement system could cause some pathophysiological aspects of COVID-19 infections, such as thrombotic microangiopathy and acute kidney injury (Ref. Reference Mahajan101). The C5 inhibitor eculizumab has be used as a potential therapeutic agent in the clinical treatment of COVID-19 (NCT04288713). Corticosteroids exert their anti-inflammatory effects by inhibiting the expression of pro-inflammatory transcription factors (Ref. Reference Ramesh102). Dexamethasone and other oral/intravenous corticosteroids are the first drugs recommended to treat cytokine storm in patients with severe COVID-19. The study shows that the use of dexamethasone resulted in lower 28-day mortality in COVID-19 patients (Ref. Reference Horby103). JAK pathway inhibitors such as baricitinib, fedratinib and ruxolitinib are effective anti-inflammatory agents against COVID-19-associated cytokine storm (Ref. Reference Richardson104). Using baricitinib in combination with anti-viral drugs such as lopinavir can reduce the inflammatory response and reduce virus recurrence in COVID-19 patients (Ref. Reference Stebbing105).
Cell therapy
MSCs can release various cytokines, such as cell growth factor, prostaglandin E2, GM-CSF, IL-6 and IL-13, to promote phagocytosis and activation of alveolar macrophages (Ref. Reference Zhou106). MSC-based therapy releases immunomodulatory factors and suppresses cytokine storm to improve the microenvironment in ARDS caused by SARS-CoV-2 infection and inhibits pulmonary fibrosis and alveolar fluid accumulation (Ref. Reference Jamshidi107). Human menstrual blood-derived MSCs were used in an exploratory study to treat severe COVID-19 patients, and the results suggested that menstrual blood-derived MSCs could reduce the mortality of COVID-19 patients (Ref. Reference Xu108). Furthermore, the potential of COVID-19 treatment can be improved by modifying the extracellular vesicles of MSCs (Ref. Reference Racchetti and Meldolesi109).
Ligands on virus-infected cells induce NK cells to activate and release IFN-γ and TNF-α, thereby lysing virus-infected cells. Ongoing clinical trials NCT04280224 and NCT04324996 in China will investigate the role of NK cells in severe pneumonia or lymphopoenia triggered by COVID-19. Recent studies have shown that chimeric antigen receptor (CAR)-NK cell therapy exhibits effective antitumor activity in haematologic cancers with minimal side effects (Ref. Reference Gang110). CAR-NK cells use the scFv of S309, a neutralising antibody that targets the highly conserved region of the SARS-CoV-2 S protein can kill target cells expressing SARS-CoV-2 S protein (Ref. Reference Ma111).
SARS-CoV-2-specific memory CD8+ T cells were detectable in SARS-CoV-2 convalescent individuals (Ref. Reference Schulien112). Furthermore, the peptide-specific memory CD8+ T cells of SARS-CoV-2 could proliferate when the antigen was re-attacked, suggesting that SARS-CoV-2-specific CD8+ T cells are not exhausted but functional (Ref. Reference Rha M113). Overexpression of inhibitory receptors CTLA4 and PD-1/PD-L1 is a typical characteristic of the exhaustion of CD8+ T cells, leading to decreased effector activity and proliferation ability. The total number of NK and CD8+ T cells was significantly decreased, and the function of NK and CD8+ T cells was exhausted in patients with SARS-CoV-2 infection (Ref. Reference Zheng114). Functional blockade of CTLA4 and PD-1/PD-L1 may unleash exhausted NK and CD8+ T cells, enhancing anti-viral immunity against SARS-CoV-2 (Refs Reference Awadasseid115, Reference Riva116).
DC is a crucial bridge between innate and adaptive immunity in anti-viral infection. COVID-19 DCs are shown to promote the secretion of cytokines and chemokines, regulate the inflammatory response and induce the differentiation of CD4+ and CD8+ T cells against pathogens (Ref. Reference Alamri117). In addition, plasma cell-like DCs inhibit viral replication by rapidly producing IFN-I (Ref. Reference Cervantes-Barragan118). It may be helpful to inhibit the functions of DC to control the excessive inflammatory process in severe COVID-19 patients.
Other immunotherapies
Fulminant activation of coagulation and consumption of clotting factors occurs in severe cases of COVID-19, leading to disseminated intravascular coagulation and patient death (Ref. Reference Tang119). On the other hand, coagulation factors can increase inflammation by acting on specific cell receptors. Considering these two aspects, inhibition of blood clotting may decrease the mortality of COVID-19 patients by reducing the risk of small intravascular thrombosis and severe inflammation. In a retrospective study, COVID-19 patients taking anticoagulants at moderate doses (7 mg/kg per 12 h) during hospitalisation were associated with a significantly lower cumulative incidence of in-hospital death compared with patients not receiving anticoagulants (Ref. Reference Meizlish120). Low-dose radiotherapy (LDRT, <100 cGy) has been used to treat a variety of chronic inflammatory diseases. LDRT is reported to have an anti-inflammatory effect by downregulating proinflammatory macrophages and upregulating anti-inflammatory macrophages and NK T cells. Thus, LDRT has the potential to counter the immune reaction incited by COVID-19 (Refs Reference Gupta121, Reference Rodel122). The latest research studies suggest that LDRT is feasible in COVID-19 patients and has shown promising results in COVID-19 pneumonia (Refs Reference Sharma123, Reference Dhawan124). IL-15 is expressed by myeloid cells to support T-cell responses, activate NK cells and modulate inflammation, and is a key immunomodulatory cytokine with antiviral properties (Ref. Reference Verbist and Klonowski125). Induction of IL-15 can elicit a robust immune response against SARS-CoV-2 (Ref. Reference Knudson126). IL-15 and IL-21 combination therapy for COVID-19 has also been performed (Ref. Reference Wilz127).
Conclusion
In this review, we have reported several aspects of the immunopathogenesis of COVID-19 and analysed the mechanism of the host immune response against this disease. Among immunotherapeutic approaches for blocking viral attachment or entry, monoclonal antibodies are preferred because of their specificity, purity, low risk of contamination with bloodborne pathogens. A monoclonal antibody cocktail or combination of different monoclonal antibodies recognising different epitopes on the viral surface may increase the efficacy of virus neutralisation.
COVID-19 is an infectious disease that causes an imbalance in the immune system and an inflammatory cytokine storm. Immunosuppressants and anti-inflammatory drugs are potential agents for the treatment of cytokine storm. For example, immunomodulators with IL-6 blockers have the therapeutic potential to specifically inhibit the status of hyperinflammation. Controlling the excessive inflammatory response is crucial to prevent the progression of ARDS in severe cases. On the other hand, enhancing host immunity is crucial to ensure that the host is sufficiently able to fight viral infections. These two aspects are essential concerns of immunotherapy against COVID-19. However, further studies are needed to evaluate the efficacy of immunotherapy in patients infected with COVID-19.
Financial support
This study was supported by the Natural Science Foundation of Guangdong Province, China (XTJ, grant number 2020A1515010981), (XL, 2020A1515110366); the Science and Technology Program of Guangzhou, China (XTJ, grant number 202102080193) and the President Foundation of Nanfang Hospital, Southern Medical University (XL, 2018C013).
Conflict of interest
None.
Ethical standards
The authors assert that this work does not involve human or animal experimental procedures.






