Introduction
In Brazil, the sugarcane crop has become increasingly important during the past 10 years, mainly because of increasing ethanol, sugar and energy demands. Water-deficit stress is recognized as a limiting factor in most of the sugarcane growing regions in Brazil (Marin et al., Reference Marin, Thornburn, Nassif and Costa2015). In some areas, irrigation is commonly needed to ensure minimum yield and to keep plants alive (Vianna and Sentelhas, Reference Vianna and Sentelhas2015). Despite the importance of this crop, few studies have focused on water use by sugarcane crops in Brazil.
Currently, irrigation management is done by using crop coefficient (Kc). However, Kc is affected by changing climate conditions and changes during growth and ripening (Allen et al., Reference Allen, Pereira, Raes and Smith1998). Under humid conditions with low wind speed, Kc is less dependent on aerodynamic variables and tends to the unity for most crops. The irrigation management for sugarcane is usually based on the FAO bulletin 56 issued to estimate Kc, which is different for early, middle and final growth (0.4, 1.25 and 0.75, respectively) stages. Some authors have reported that for maximum sugarcane crop growth, Kc values should range between 0.5 and 1.1 (Silva et al., Reference Silva, Moura, Zolnier, Soares, Vieira and Júnior2012), while Inman-Bamber and McGlinchey (Reference Inman-Bamber and McGlinchey2003) have reported Kc values between 0.5 and 1.4, depending on the radiation intercepted by the crop.
This variability arises from factors affecting the process of water loss that are not measured or considered in water consumption studies for homogeneous canopies such as sugarcane. The plant–atmosphere-related decoupling factor (Ω) (Jarvis and McNaughton, Reference Jarvis and McNaughton1986) has a strong relation to the atmospheric water demand for many crops (Marin et al., Reference Marin, Angelocci, Nassif, Costa, Vianna and Carvalho2016; Pereira, Reference Pereira2004), including sugarcane. Jarvis (Reference Jarvis1985) has explained that the decoupling factor is a numerical index of plant–atmosphere interaction and is related to the crop aerodynamic (ra) and stomatal (rs) resistances, allowing to know the variation of physiological responses to different states of atmosphere, mainly photosynthesis and water use efficiency. Aerodynamic factors are more important for defining gas exchange under coupling conditions, while the incidence of radiation becomes more important under decoupling conditions.
According to Jarvis and McNaughton (Reference Jarvis and McNaughton1986), extrapolation of experimental data on plant water consumption can result in uncertainty due to scaling up or down. For example, Eksteen et al. (Reference Eksteen, Singels and Ngxaliwe2014) have evaluated two sugarcane genotypes and found Ω to be between 0.187 and 0.968 with varying water availability. Therefore, in an attempt to reduce this source of uncertainty and to provide consistent data to improve sugarcane irrigation management, we used a set of measurement techniques at different scales for the assessment of water use in sugarcane crop.
Material and Methods
Experimental fields
Two experimental field trials were carried out at Piracicaba, Brazil (22.67ºS; 47.64ºW; 530 m a.s.l., both as Cwa Koeppen classification), with two sugarcane cultivars. In the first experimental area (EA1), we evaluated the first ratoon of CTC 12 cultivar planted in February 2010, with twin row spacing (0.5 × 1.5 m), grown in a Udox soil of 0.3 ha and harvested after 365 days (March 2012). This field received full subsurface drip irrigation without water deficit. In the second experimental area (EA2), we evaluated the plant cycle of RB867515 cultivar planted in 2012/13 growth season in an Ultisol soil (2 ha). A simple spacing (1.4 m) was used; the planting date was October 2012, and it was harvested after 365 days. This field received full sprinkling irrigation by a centre pivot, controlled by daily crop water balance with at least 80% of soil water-holding capacity.
Daily weather data (maximum and minimum air temperature, rainfall, air humidity, average wind speed, wind direction, global and net radiation) were collected every 15 min for both experiments throughout the crop cycle with the help of an Automated Weather Station (AWS) at 1 km from the experimental area. The AWS has been maintained following the World Meteorological Organization standards, and is composed of Campbell Scientific sensors: one pluviometer (TB4), one thermo-hygrometer (HMP-155), one wind speed and direction sensor (034A), one barometer (CS106), one rugged pyranometer (CM3), one net radiometer (NR-LITE2) and a quantum sensor (LI190SB).
Evapotranspiration, transpiration and decoupling factor methods and calculations
A Bowen ratio energy balance (BREB) method was used to evaluate the mass and energy exchange over fields with two forced ventilation psychrometers (Marin et al. Reference Marin, Angelocci, Coelho Filho and Villa Nova2001). In EA1, the fetch was 85 m from predominant wind direction (SE). In EA2, the fetch was 110 m from predominant wind direction (SE) and 90 m from other directions. When winds were from an unsuitable direction for this technique, with no good fetch, recorded data were discarded.
Measurements of dry and wet bulb temperatures (°C) were performed at two heights for the evaluation of latent and sensible heat fluxes from the sugarcane canopy. There was a height difference of 1 m between them, with the lower measurement maintained at least 0.5 m above canopy height and following sugarcane plant growth (Allen et al., Reference Allen, Pereira, Howell and Jensen2011). The BREB method data were collected from 30 April 2011 to 31 December 2011 at EA1 and between 14 February 2013 and 20 June 2013 at EA2. A net radiometer was installed 3 m above the canopy, and two soil heat flux sensors were also installed at 2 cm depth, on sugarcane rows and inter-rows. Crop evapotranspiration was determined according to Equation 1:
 $$LE = {{Rn - G} \over {1 + \beta }} \to ETc = {{Rn - G} \over {\lambda (1 + \beta )}}$$
(1)
$$LE = {{Rn - G} \over {1 + \beta }} \to ETc = {{Rn - G} \over {\lambda (1 + \beta )}}$$
(1)
where Rn is the net radiation (MJ m−2 d−1), G is the soil heat flux (MJ m−2 d−1), β is the Bowen ratio, LE is the latent heat flux (MJ m−2 d−1), and λ is the latent heat of evaporation.
The Bowen ratio values (β) were calculated for each 15-min interval based on the temperature gradient values (ΔT, °C), vapour pressure gradient values (Δe, kPa) and psychometric constant (γ), according to Equation 2:
 $$\beta = \gamma {{\Delta T} \over {\Delta e}}$$
(2)
$$\beta = \gamma {{\Delta T} \over {\Delta e}}$$
(2)
The BREB can show some variability in values, which were checked according to the methodology of Perez et al. (Reference Perez, Castellvi, Ibañez and Rosell1999). When measures had such variability, interpolations were done. When periods exceeded 2 h of such undesired variability, the whole day’s data were discarded. Only daytime data were used to compute evapotranspiration (ET) from the BREB method.
Sap flow (SF) measurements by the heat balance method (Sakuratani, Reference Sakuratani1981) were performed on four representative stalks (Eksteen et al., Reference Eksteen, Singels and Ngxaliwe2014) at the internodes in both areas, which were used to evaluate the water use at the plant scale. We used non-invasive sensors with a constant heat source (Dynamax Sap Flow System, SGB25-WS, Houston, Texas). SF gauges were attached 30 cm above the soil surface, at the internode portion of the stem, to avoid interference of soil heat flow, as done by Eksteen et al. (Reference Eksteen, Singels and Ngxaliwe2014). Leaves on this position were removed, and a thermal paste was used to ensure close contact between gauge and stem. Stems were changed every 10 days to reduce the chance of long-term heat damage and the sprouting of node roots. To avoid any interference of incident solar radiation, the sensors were coated with a reflective foil. SF was calculated using Equation 3 (Sakuratani and Abe, Reference Sakuratani and Abe1985):
 $$SF = {{P - Qa - Qr} \over {dT*cp}}$$
(3)
$$SF = {{P - Qa - Qr} \over {dT*cp}}$$
(3)
where SF is the sap flow (kg s−1), P is the power applied (W), Qa is the axially dissipated energy (W), Qr is the radially dissipated energy (W), dT is the upper and lower temperature difference (°C) and cp is the specific heat of water (4.186 J kg−1 °C−1).
The determination of thermal conductivity of the radial heat meter (Kr) was performed with collected data between 3:00 and 5:00 am, when SF was considered to be close to 0. In low SF conditions, the differences between temperature sensors were close to 0, which can lead to excessively high transpiration data (Marin et al., Reference Marin, Ribeiro, Angelocci and Righi2008). The 24-h integrated values of SF were considered as representative of the daily transpiration of each plant. Transpiration rates were normalized to obtain a transpiration rate on a leaf area unit basis (mm m−2 of leaf). The crop transpiration was scaled up to a ground area unit basis by multiplying the average transpiration rate of the four plants by the average leaf area index determined using the gap-fraction method (LAI-2000, Li-Cor, Inc.).
Diurnal courses of stomatal conductance (gs) were measured during 5 days in the EA2 treatment with an infrared gas analyser (LCproT Advanced Portable Photosynthesis System, ADC) on the exposed leaves (the first leaf with an apparent dewlap) between 9:00 am and sunset (local time) in 10 plants. Values of gs were manipulated to be expressed as diffusive resistance (rs), and the decoupling factor (Ω) was computed for a hypostomatous leaf as defined by Equation (4) (Jarvis and McNaughton, Reference Jarvis and McNaughton1986):
 $${\rm{\Omega }} = {\left[ {1 + \left( {{\gamma \over {\gamma + s}}} \right){{rs} \over {ra}}} \right]^{ - 1}}$$
(4)
$${\rm{\Omega }} = {\left[ {1 + \left( {{\gamma \over {\gamma + s}}} \right){{rs} \over {ra}}} \right]^{ - 1}}$$
(4)
where ra is the canopy aerodynamic resistance, rs is the stomatal resistance to vapour diffusion, γ is the psychrometric constant and s is the tangent of the vapour saturation curve.
Conceptually, the extreme values for the decoupling factor were as follows: Ω → 1 as rs/ra → 0, implying that the net radiation is the only contributor to the evapotranspiration process and that vegetation is completely decoupled from atmospheric conditions; and Ω → 0 as rs/ra → ∞, indicating complete coupling of vegetation with atmospheric vapour pressure deficit (VPD) and wind speed (Marin and Angelocci, Reference Marin and Angelocci2011).
Results
Weather conditions
At EA1 (cultivar CTC 12), the average air temperature was 21.8 °C, ranging from 16.1 to 29.0 °C, with 1456 mm total rainfall. At EA2 (cultivar RB867515), the average air temperature was 22.7 °C, ranging from 16.8 to 30.3 °C and with 1318 mm total rainfall. The predominant wind direction was SE in both areas, with 51% of the values between 90 and 180º. These data were important for the fetch used in both experiments.
Crop energy balance and evapotranspiration
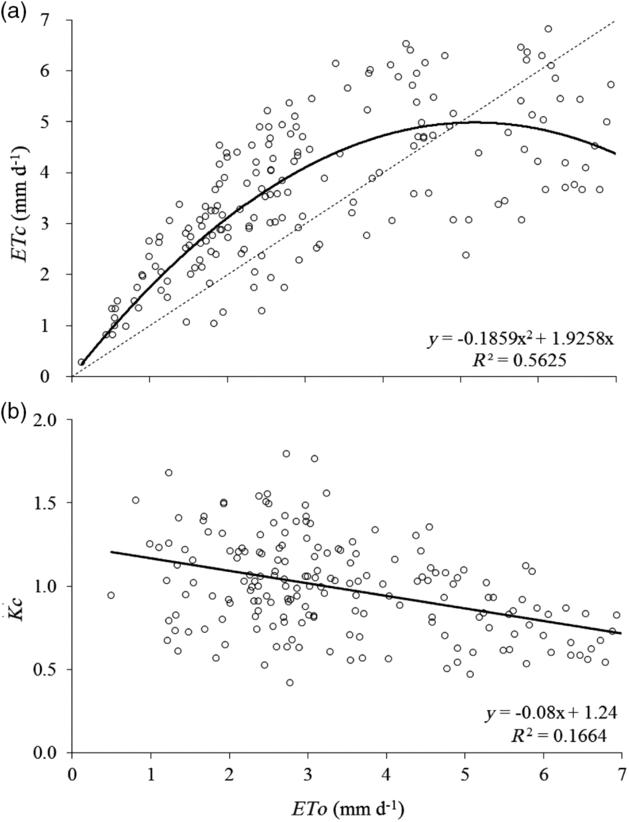
At EA1, the crop evapotranspiration (ETc) was higher than the reference evapotranspiration (ETo) during days with low ETo, and the opposite trend was detected during days with high ETo. EA1 had a mean ETc of 3.68 mm d−1 and mean ETo of 4.18 mm d−1. At EA2, ETc was similar to ETo for all days, with a mean ETc of 2.92 mm d−1 and mean ETo of 2.89 mm d−1 at the maximum crop growth period. Overall, the average crop coefficient (Kc) for the full vegetative growth stage was 0.99 ± 0.29 (Figure 1a).

Figure 1. Relationship between reference evapotranspiration (ETo) and crop evapotranspiration (ETc) (a) and crop coefficient (Kc) (b) based on the BREB method for both experimental areas (dotted line is 1:1) (p-value <0.05 for both figures).
The ETc and ETo values resulted in a variable Kc, and this variability occurred because Kc was high when ETo was low and Kc decreased when ETo increased (Figure 1b, Table 1). Thus, there is an inverse relationship between Kc and VPD. When VPD was close to 0, Kc values were close to 1.7, and when VPD was around 4.0 kPa, the mean Kc was 0.9, showing that ETc had a smaller increase under high VPD conditions.
Table 1. Relationship between reference evapotranspiration ranges (ETo), crop evapotranspiration (ETc), crop coefficient (Kc) and basal crop coefficient (Kcb) from both experimental areas with RB867515 (2013) and CTC 12 (2011) cultivars

Sap flow and transpiration
The thermal conductivity of radial heat meter (Kr) was similar in both experimental areas. At EA1, Kr varied from 0.050 to 0.098 W C−1, and this variation was caused by differences in stem diameter from 28 to 31 mm. At EA2, the Kr pattern was similar in all gauges and ranged between 0.035 and 0.045 W C−1. These values were lower than those reported by Marin et al. (Reference Marin, Ribeiro, Angelocci and Righi2008) for coffee with Kr reaching 0.180 W C−1. These authors, however, also reported (in addition to long-term Kr variation) an influence of the size of sensors used in the study, that is, larger sensors have higher Kr. In the present study, we used gauges with 25 mm diameter at both experimental sites.
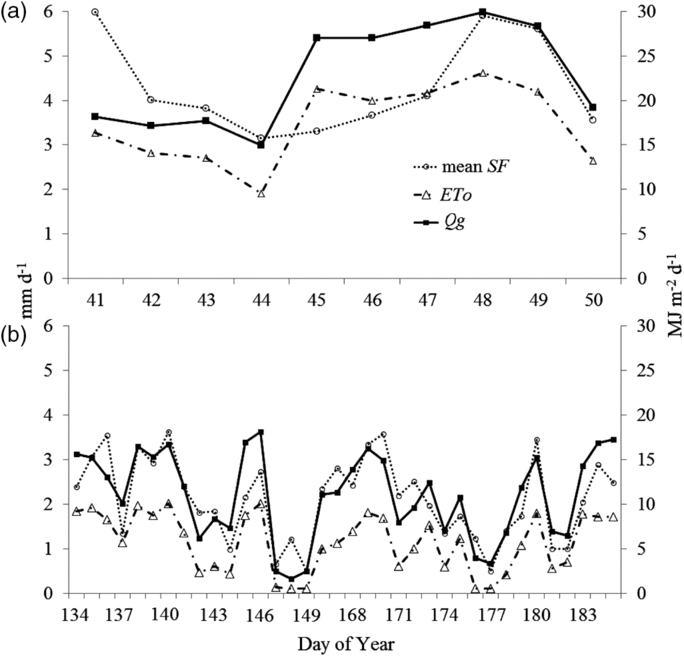
The four SF sensors installed at EA1 showed similar rates of water loss over the days, ranging from 0.23 to 0.50 L per stem for the 44th and 49th days of the year, respectively (Figure 2), representing 2.16 and 4.71 mm d−1 of transpiration. These values were higher than those detected at EA2, where water loss rates varied from 0.06 to 0.41 L per stem for the 177 and 140 days of the year (DOY), respectively, resulting in transpiration rates of 0.50 and 3.62 mm d−1. This difference was because of differences in the environment energy availability: measurements in EA1 and EA2 were taken in February 2012 (higher air temperature) and between May and June 2013 (lower air temperature), respectively.

Figure 2. Temporal variation of mean sap flow (SF, mm d−1), reference evapotranspiration (ETo, mm d−1) and global radiation (Qg, MJ m−2 d−1) for the analysed days on sugarcane crop in EA1 (a) and EA2 (b).
At EA1, there were higher rates of water loss for the 48th and 49th DOY: SF = 4.14 and 4.71 mm d−1, and Qg = 29.9 and 28.4 MJ m−2 d−1, respectively. Low water loss rates were found on 43 and 44 DOY: SF = 2.64 and 2.16 mm d−1, and Qg = 17.7 and 14.9 MJ m−2 d−1, respectively. For EA2 (Figure 2b), water loss was relatively higher on 136, 140, 170 and 180 DOY (Qg varying between 13.0 and 16.7 MJ m−2 d−1 and SF between 3.46 and 3.62 mm d−1) than on 147, 149 and 177 DOY (Qg between 2.4 and 3.3 MJ m−2 d−1 and SF between 0.49 and 0.64 mm d−1).
Assuming that the daily-integrated SF was equivalent of crop transpiration (T), crop T values were higher than ETo values on most of the EA1 season (Figure 2a), except for few cases between 45 and 47 DOY, which will be discussed later. Comparing T (2.46 ± 1.04 mm) and ETo (2.39 ± 1.19 mm), we found that T was 3% higher than ETo (Figure 3a). The SF measurement campaigns were done mostly during low ETo periods (<3 mm d−1). We also observed a trend for a reduction in the basal crop coefficient (Kcb) when ETo increased (Figure 3a), resulting in 95% of water loss from transpiration and 5% from soil water evaporation at both experimental sites.

Figure 3. Relationship between sap flow transpiration (T) and reference evapotranspiration (ETo) (a) and basal crop coefficient (Kcb) (b) and ETo for both experimental fields (p-value <0.05 for both figures).
Stomatal conductance and decoupling factor
A rapid increase in gs was observed in days with high incident radiation (28 February and 19 April). The peak of gs (0.39 mol m−2 s−1) occurred by noon, and there was a softer and constant decline in the afternoon, reaching values close to 0 (Figure 4a). This was also noticed on cloudy days with low insolation (19 March, 14 May and 4 June). However, gs had a small variation, and for some of the days the maximum gs values were considerably low (close to 0.20 mol m−2 s−1).

Figure 4. Daily variation of stomatal conductance (gs) (a) and decoupling factor (Ω) (b) for 28 February, 19 March, 9 April, 14 May and 4 June 2013 for EA2.
On 28 February and 4 June 2013 (at EA2), the sugarcane canopy was decoupled from the atmosphere (Ω = 0.65) in the morning (9:00 am) and showed increasing plant–atmosphere coupling (Ω = 0.1) until sunset. On 9 April and 14 May, the sugarcane crop was coupled at 9:00 am (Ω = 0.2), with such coupling decreasing at noon (Ω = 0.5) and increasing towards sunset (Ω = 0.1) due to high wind speed. On 14 May, for example, the sugarcane crop was coupled all-day-long (0.1 < Ω < 0.45) due to high wind speed throughout the day (Figure 4b). The mean Ω values were between 0.19 (14 May 2013) and 0.52 (28 February 2013), showing that the sugarcane crop canopy was coupled to atmosphere.
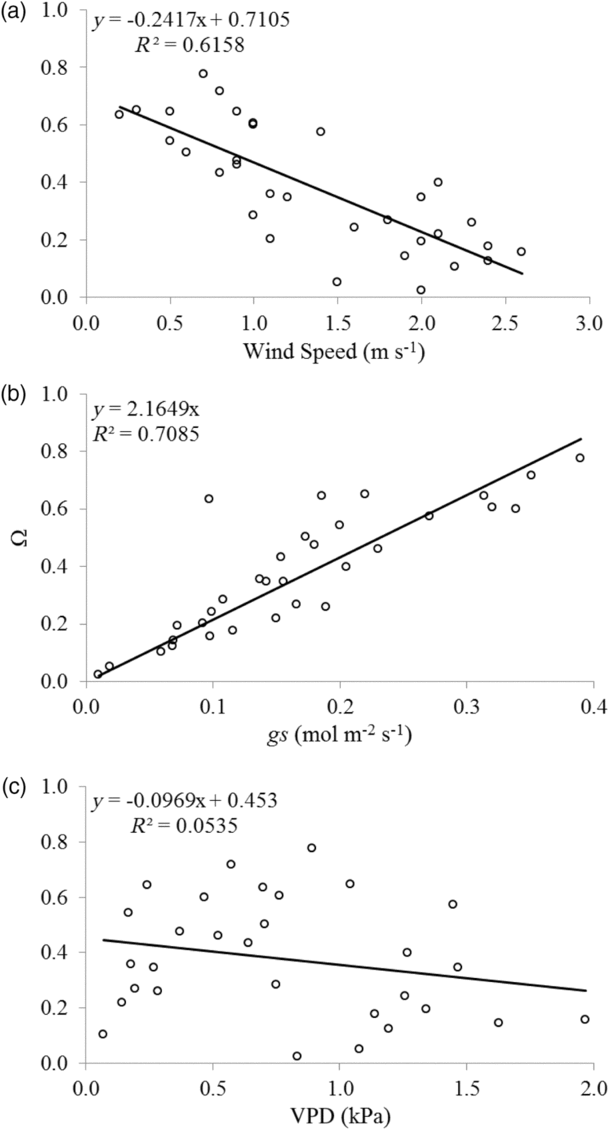
Under low wind speed, the sugarcane crop tended to decouple from the atmosphere, resulting in water loss related mainly to the incident radiation. We found smaller Ω (R 2 = 0.62) under high wind speed (u) conditions (Figure 5a). Increases in gs also affected the relationship between the canopy and aerodynamic components, with high gs causing further decoupling (R 2 = 0.7) (Figure 5b). VPD probably had short interference on crop–atmosphere coupling when assessed individually, as shown in Figure 5c (R 2 = 0.05).

Figure 5. Relationship between decoupling factor (Ω), wind speed (a), stomatal conductance (b) and VPD (c) for EA2 (p-value <0.05 for all figures).
Discussion
During both crop cycles, when the BREB method was used, it was observed that the mean sensible heat flux (H) was 10% of the available energy (Rn-G) and that the mean latent heat flux was 85% of Rn-G. This high difference between H and LE was due to high water availability in the environment, resulting in a lower H flux on the sugarcane crop. In maize, Zeggaf et al. (Reference Zeggaf, Takeuchi, Dehghanisanij, Anyyoji and Yano2008) have detected a crop H flux of about 8% (and always <10% of available energy), and the remaining energy was used to LE flux. For a full irrigated coffee plantation, Righi (2004) has reported LE representing 64% and 80% of Rn and a mean H value about 29%. For the sugarcane crop, Cabral et al. (Reference Cabral, Rocha, Gash, Ligo, Tatsch, Freitas and Brasilio2012) have measured LE and H fluxes on an irrigated field in northern Brazil using the Eddy correlation method and reported values of 97% for the sum of H and LE.
According to the FAO 56 bulletin (Allen et al., Reference Allen, Pereira, Raes and Smith1998), the Kc value for the maximum growth period of sugarcane is 1.25. However, here we estimated a lower mean Kc (0.99) compared with the FAO 56 bulletin, and this varied on daily basis, depending on ETo (Table 1). Our findings differ from those of Inman-Bamber and McGlinchey (Reference Inman-Bamber and McGlinchey2003) for irrigated sugarcane crop in Australia and Swaziland (for maximum growth period) and of Olivier and Singels (Reference Olivier and Singels2012) for South Africa. Nevertheless, these authors did not mention the Ω values for experimental fields. For a Brazilian cultivar (RB92579), Silva et al. (Reference Silva, Moura, Zolnier, Soares, Vieira and Júnior2012) have reported an average ETc of 4.7 mm d−1 for a semi-arid area in northern Brazil, also using the BREB method and resulting in a constant Kc of 1.1. This crop response could be due to the same condition observed under higher ETo, when the atmospheric demand was high. According to Marin et al. (Reference Marin, Angelocci, Righi and Sentelhas2005), differences in microclimatic conditions, especially atmospheric demand, are important for Kc determination, with Kc varying as function of VPD in a coffee plantation. The same relation was reported by Marin and Angelocci (Reference Marin and Angelocci2011) in a citrus orchard, where the crop had low water loss by transpiration under conditions of high atmospheric demand. Marin et al. (Reference Marin, Angelocci, Nassif, Costa, Vianna and Carvalho2016) have found that Kc decreases when ETo increases as a consequence of high plant–atmosphere coupling and high crop inner resistance, which limits the amount of water that plants can supply to the atmosphere. Even for sugarcane plantation (after full soil cover), Kc decreased with increasing ETo, highlighting that this trend might not be exclusive of tall sparse crops and for plants well coupled to the atmosphere.
Transpiration values were lower than observed by Chabot et al. (Reference Chabot, Bouarfa, Zimmer, Chaumont and Moreau2005) in Morocco (Mediterranean semi-arid climate) where sugarcane grown in lysimeters had an average transpiration rate of 8.0 mm d−1. However, SF measurements can result in an overestimation of the crop transpiration rate by up to 35%. Boehringer et al. (Reference Boehringer, Zolnier, Ribeiro and Steidle Neto2013) have used the same method with resistors introduced into stems and reported up to 140 g per stem water loss and that SF might underestimate water loss by up to 5% when compared with gravimetric measurements.
There was a decreasing trend in the T/ETo ratio when ETo was higher than 3 mm d−1 (Figure 3a), resulting in a constant decrease in Kcb values (Figure 3b). A similar result was reported by Marin and Angelocci (Reference Marin and Angelocci2011) for acid lime trees, where transpiration did not follow ETo but stabilized when ETo reached 4 mm d−1. This relationship between water loss and net radiation was also described by Marin et al. (Reference Marin, Ribeiro, Angelocci and Righi2008) for a coffee plantation. Sakuratani and Abe (Reference Sakuratani and Abe1985) have found that low radiation leads to a partial stomatal closure in sugarcane plants, and they also detected a decrease in transpiration due to decreasing direct radiation incidence from noon to sunset and consequent increase in diffuse radiation.
Although without practical applications, the time course of Kcb follows the Kc pattern (Table 1), providing an estimative of the soil component on sugarcane crop evapotranspiration. Because the measurements of Kcb and Kc were performed with two independent methods, it can be concluded that measurement errors were an issue.
Under water-deficit stress conditions, sugarcane has gs values between 0.15 and 0.25 mol m−2 s−1 (Machado et al., Reference Machado, Ribeiro, Marchiori, Machado, Machado and Landell2009). Under irrigated conditions, sugarcane crops have higher gs values, decreasing in the afternoon due to increased VPD (Roberts et al. Reference Roberts, Nayamuth, Batchelor and Soopramanien1990). Recently, Eksteen et al. (Reference Eksteen, Singels and Ngxaliwe2014) have reported average gs values of up to 0.10 mol m−2 s−1 for two sugarcane cultivars in South Africa, but this was not measured throughout the entire day. As described by Angelocci et al. (Reference Angelocci, Marin, Oliveira and Righi2004), gs patterns are usually influenced by many environmental factors, imposing a challenge to isolate the individual relationships between gs and VPD, leaf temperature and incident radiation. Also, no clear relationships were identified in this research between those environmental variables and gs.
Here, we found that gs affected the decoupling factor (mean Ω = 0.37), and this finding disagrees with Silva et al. (Reference Silva, Moura, Zolnier, Soares, Vieira and Júnior2012), who found higher Ω (0.74) under irrigation than under low water availability (0.6) in a furrow irrigation sugarcane crop in northern Brazil. Eksteen et al. (Reference Eksteen, Singels and Ngxaliwe2014) have reported decoupled sugarcane canopy (mean Ω = 0.81) in South Africa for the N19 and G73 cultivars, and these results suggest that radiation is the main factor controlling evapotranspiration in warm and dry regions, such as northeast Brazil and South Africa, whereas wind speed and VPD are the main factors in cold and rainy regions.
According to Jarvis and McNaughton (Reference Jarvis and McNaughton1986), the effects of gs variation in leaf transpiration can only be extrapolated to another scale if environmental conditions around this leaf and target scale are similar for Ω and VPD. Considering the known gs spatiotemporal variability (Eiksteen et al., Reference Eksteen, Singels and Ngxaliwe2014; Inman-Bamber and Smith, Reference Inman-Bamber and Smith2005; McNaughton and Jarvis, Reference McNaughton and Jarvis1991; Nassif et al., Reference Nassif, Marin and Costa2014; Smith et al., Reference Smith, Inman-Bamber and Thorburn2005) and Ω variability, extrapolation should be done with caution from leaf to plant and for different crop species. Jarvis and McNaughton (Reference Jarvis and McNaughton1986) have also noted that, as a result of individual stomatal control, leaves of the same plant might have different transpiration rates and different plants in same environmental conditions might have different water use rates. This can explain the variation of sugarcane crop Kc (Table 1) with lower Ω (highly coupled canopy), and therefore, the crop would control water vapour exchange with the atmosphere. In contrast, incident radiation is mainly responsible for water loss when the crop canopy trends to be decoupled (high Ω) (Steduto and Hsiao, Reference Steduto and Hsiao1998).
Conclusions
The RB867515 and CTC12 sugarcane cultivars were coupled to the atmosphere in southeast Brazilian climatic conditions (Ω = 0.37), with Kc and Kcb showing temporal variability due to reference evapotranspiration. These results indicate a strong relationship between sugarcane water loss and environmental variables, with a growth stage-specific Kc that often diverges from the FAO 56 bulletin recommendation. We found that almost all of the water loss was due to crop transpiration for both sugarcane cultivars, which has important implications for the irrigation management of this crop.
Author ORCIDs
Daniel Silveira Pinto Nassif 0000-0002-9780-1781; Leandro Garcia da Costa 0000-0002-4062-8877; Murilo dos Santos Vianna 0000-0003-1139-4589; Kassio dos Santos Carvalho 0000-0003-1184-3479; Fabio Ricardo Marin 0000-0003-1265-9032
Acknowledgements
We thank the State of São Paulo Research Foundation (FAPESP) grants #2011/18072-2, #2013/16511-4, #2014/12406-4, #2014/50023-0, #2014/05887-6 and the National Council for Scientific and Technological Development (CNPq) 302872/2012, 480702/2012, 403946/2013 for fellowships granted and financial support to this research.
Financial support
None.