

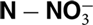


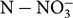
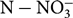
Introduction
Potato (Solanum tuberosum L.) is the fourth most important food crop in the world after corn (Zea mays L.), wheat (Triticum aestivum L.), and rice (Oryza sativa L.) (FAO, 2009; Spooner and Bamberg, Reference Spooner and Bamberg1994), with Colombia being the third country with the highest annual production of potato in Latin America (FAOSTAT, 2018). Cultivation of potato in Colombia depends on various factors, among which fertilization practices account for 20–25% of costs involved in the production (Gómez and Torres, Reference Gómez and Torres2012). To ensure an efficient use of mineral nutrients, several diagnostics tools could be used, such as soil (INIAP, 2000; Oliveira et al., Reference Oliveira, Afif and Mayor2006) and foliar analyzes that allow determining the status of nutrients at a certain moment of crop cycle (Cadahía, Reference Cadahía2008). The use of remote sensors has become an important tool permitting to access, in real time, nutrient status of the crop and, thus, provides timely decisions (Gutiérrez et al., Reference Gutiérrez, Cadet, Rodríguez and Araya2011; Zhou et al., Reference Zhou, Jabloun, Plauborg and Andersen2018).
One of these tools is a GreenSeeker™sensor used to forecast the crop growth and yield and, in turn, is associated with the development of canopy and total plant biomass (Giletto and Echeverria, Reference Giletto and Echeverria2012; Giletto et al., Reference Giletto, Zamuner, Silva and Cassino2013; Gutiérrez et al., Reference Gutiérrez, Cadet, Rodríguez and Araya2011). This sensor generates measurements in normalized difference vegetation index (NDVI) units related to plant capacity to reflect light energy of different wavelengths, which is connected to chlorophyll content in leaves (Gutierrez et al., Reference Gutiérrez, Cadet, Rodríguez and Araya2011; Schepers et al., Reference Schepers, Blackmer, Wilhelm and Resende1996). The principle of measurement consists in plant capacity to absorb energy in red (600–700 nm) spectrum of wavelengths and reflect it in the infrared one (>700 nm) (Bongiovanni et al., Reference Bongiovanni, Chartuni, Best and Roel2006). The GreenSeeker sensor emits a beam of light in red and infrared spectrum, which reaches leaves, so that reflected light is then captured by the apparatus (Govaerts et al., Reference Govaerts, Verhulst and Fuentes2010; Gutiérrez et al., Reference Gutiérrez, Cadet, Rodríguez and Araya2011; Peters et al., Reference Peters, Walter, Ji, Viña, Hayes and Svoboda2002).
The NDVI readings obtained by the GreenSeeker sensor have been correlated to many variables, such as crop nutrient deficiency, final yield in small grains, and long-term water stress (Verhulst and Govaerts, Reference Verhulst and Govaerts2010). In the Central United States, NDVI performed well-predicting soybean yield (Bolton and Friedl, Reference Bolton and Friedl2013). When corn plants were closely spaced together, GreenSeeker readings were higher and plant yield lower (Raun et al., Reference Raun, Solie, Martin, Freeman, Stone, Johnson and Mullen2005). According to Raun et al. (Reference Raun, Solie, Martin, Freeman, Stone, Johnson and Mullen2005), spatial variability of NDVI increased at the V6 growth stage of corn indicating the impact that crop phenology might have on NDVI readings. One of the practical tasks frequently assessed with NDVI is nutrient deficiency in plants, especially that of nitrogen (N) (Govaerts et al., Reference Govaerts, Verhulst and Fuentes2010; Ruíz et al., Reference Ruíz, Tijerina, Mejia, Sandoval, Sánchez, Vaquera and Colinas2011), which is directly related to photosynthetic process being an essential function of chlorophyll (Alonso et al., Reference Alonso, Rosados, Ignacio, Rozas, Lamas, Chapela and Fontúrbel2008). A deficiency of N induces a reduction in chlorophyll content affecting light absorption and, therefore, reflectance of light by N-deficient plants (Dutta et al., Reference Dutta, Singh and Panigrahy2014). Thus, the GrenSeeker sensor was successfully employed to assess the status of N in corn and spring wheat in Quebec and Ontario, Canada. It was capable of describing the N condition of the crop or variation in the stand and performed well where NDVI values exceeded 0.5 (Tremblay et al., Reference Tremblay, Wang, Ma, Belec and Vigneault2009). Different studies, mostly in N deficiency and plant reflectance, reported increases in near infrared radiation (NIR) reflected with an increased dose of N fertilizer (Feng et al., Reference Feng, Yao, Zhu, Tian and Cao2008). Giletto et al. (Reference Giletto, Zamuner, Silva and Cassino2013) found that potato plants with reduced contents of N had lower NDVI values (0.82) as compared with plants cultivated under optimal conditions of N supply (NDVI = 0.88) (Govaerts et al., Reference Govaerts, Verhulst and Fuentes2010). In cultivar Russet Burbank (Group Chilotanum), the GreenSeeker sensor was shown to accurately predict N needs through NDVI values (Bowen et al., Reference Bowen, Hopkins, Ellsworth, Cook and Funk2005).
The NDVI could also be used as an indicator of environmental stress; in potato cultivars subjected to water deficit, a positive correlation was found between NDVI and canopy size or yield, with higher NDVI readings typical for cultivars tolerant to water deficit, thus proving the effectiveness of this tool to forecast yields in potato (Giletto and Echeverria, Reference Giletto and Echeverria2012; Schafleitner et al., Reference Schafleitner, Gutierrez, Espino, Gaudin, Pérez, Martínez, Domínguez, Tincopa, Alvarado, Numberto and Bonierbale2007).
Potato is one of the major crops affecting human nutrition worldwide (George et al., Reference George, Taylor, Dodd and White2018). At the same time, in potato Group Andigenum, which is considered one of the principal agricultural products in Colombia (Gómez et al., Reference Gómez, Magnitskiy, Rodríguez and Darghan2017), almost no studies were done in field diagnostics of crop nutrition, such as through NDVI and
 ${\rm NO}_3^ - $
or K+ concentrations in stem sap. These tools might permit rapid adjustments to the current practices of N and K fertilization of potato in producing regions. The objective of this study was to determine the relationships between NDVI and yield as well as concentrations of
${\rm NO}_3^ - $
or K+ concentrations in stem sap. These tools might permit rapid adjustments to the current practices of N and K fertilization of potato in producing regions. The objective of this study was to determine the relationships between NDVI and yield as well as concentrations of
 ${\rm N - NO}_3^ - $
and K+ in stem sap of potato plants in relation to different fertilizer rates.
${\rm N - NO}_3^ - $
and K+ in stem sap of potato plants in relation to different fertilizer rates.
Materials and Methods
The study was conducted at the Center for Nutrition Research in Plants of the Tropics (CENIPLANT) in Facatativá, Cundinamarca Province, Colombia in 2015. The research site had a dry climate of the cold tropics and was located west of Bogotá, Colombia (4°49′26.9″N, 74°22′29.7″W, 2520 m a.s.l.) having an average annual precipitation of 850 mm, maximum air temperature of 21.5°C, minimum air temperature of 2°C (Figure 1), average air temperature of 12.6°C, and average relative air humidity of 85%. The soil was classified as a highly fertile Andic Eutrudepts, with loam texture, high contents of organic matter (130 g kg−1), acid reaction (pH 5.82), and adequate levels of phosphorus (70.6 mg kg−1, Bray II). The soil had high saturation of bases, high saturation of calcium (84.1%), low saturation of magnesium (11.2%), average saturation of potassium (4.5%), and adequate contents of micronutrients.

Figure 1. Daily precipitation, minimum temperature, and maximum temperature during the growing period in 2015. The tubers were harvested at 150 dap.
A repeated-measures design was implemented with two factors: two cultivars and four doses of fertilizers. The measurements over time were associated with five principal phenological stages of potato growth, adapted from Segura et al. (Reference Segura, Santos and Ñústez2006) and Valbuena et al. (Reference Valbuena, Roveda, Bolaños, Zapata, Medina, Almanza and Porras2010): development of primary stems and start of vegetative growth (50–55 days after planting, dap); formation of secondary stems and initiation of tuberization (70–75 dap); flowering, maximum tuberization, and start of tuber filling (90–100 dap); cessation of flowering and tuber filling (120–125 dap); and maximum tuber filling and maturation (150–160 dap).
In the field, a split plots design was established with three replications, where the main plot corresponded to potato cultivars (Group Andigenum) Diacol Capiro (Capiro) and Pastusa Suprema (Suprema) and the subplots were represented by four treatments: three levels of balanced edaphic fertilization in doses of 1450, 1900, and 2375 kg fertilizer ha−1 and a control without fertilizer application. Fertilizers were applied manually, and each dose was split into two applications according to Table 1. The first application was done at planting in the furrow with the seed and the remaining fertilizer was applied in a band 4–5 cm to the side of the plant (Table 1). The fertilizer doses were based on historical recommendations for these cultivars in the region and adjusted by soil–plant balance method (Castro and Gómez, Reference Castro, Gómez, Burbano and Silva2013).
Whole tubers (~70 g) were planted manually on June 30, 2015 and harvested on November 30, 2015. The planting was carried out in 24 experimental units of 50 m2 each (135 plants unit−1), with a distance of 1 m between the rows and 0.37 m between the plants, a harvest area of 36 m2 (three central rows), and a density of 27 000 plants ha−1. The plants were irrigated to maintain soil moisture at field capacity. Field management, such as weed and phytosanitary controls, was carried out according to local commercial practices. At each phenological stage, dry weight (DW) was evaluated in leaves, stems, and tubers employing five plants per unit. The DW was determined after drying plant material to constant weight at 70°C for 72 h.
NDVI was defined as
 $${\rm NDVI} = \left({\rm NIR} - {\rm RED} \right)/\left({\rm NIR} + {\rm RED}\right),$$
$${\rm NDVI} = \left({\rm NIR} - {\rm RED} \right)/\left({\rm NIR} + {\rm RED}\right),$$
where
RED (µmol m−2 s−1) is radiation in red wavelengths, incident on plant surface;
Table 1. Contribution of mineral nutrients (kg ha−1) in treatments with edaphic fertilization

† Splitting of fertilizer dose and moments of application for N: 55% at planting and 45% 45–50 dap; P: 80% at planting and 20% 45–50 dap; K: 12% at planting and 88% 45–50 dap; Mg, 63% at planting and 37% 45–50 dap; micronutrients, 65% at planting and 35% 45–50 dap. Granular fertilizer sources were used: N and P, Diammonium phosphate (18–46–0), Nitrax-S®(28–4–0); K, KCl (0–0–60); Mg, Kieserite (25% MgO); B, Zn, Cu, Mn, and Fe (Nutricomplet®, Ingeplant SAS, Bogotá, a complex source of micronutrients based on sulfates).
NIR (µmol m−2 s−1) is NIR reflected by plant surface.
The NDVI was measured in each phenological stage using a Trimble GreenSeeker Handheld Crop Sensor (Trimble, Inc., Sunnyvale, CA) positioned 60 cm above the plants in each of the subplots. Four NDVI data points were taken from each subplot to make up the average NDVI value for that plot at each moment. A preliminary calibration of NDVI was carried out, due to the effect of time (Supplementary Material Figure S1), where the most appropriate moments were determined to evaluate this index. These were obtained from measurements executed every 30 min during the day in Capiro, thus determining that morning hours between 8:30 and 9:30 am and evening hours between 3:30 and 4:30 pm were the adequate ones. These measurements were not influenced by the excess of solar radiation that generally affects plant photosynthetic efficiency at noon (Manrique, Reference Manrique2003). Therefore, in the present study, the measurements of NDVI and
 $\rm N - NO_3^ - $
or K+ concentrations in stem sap were taken between 8:30 and 9:30 am.
$\rm N - NO_3^ - $
or K+ concentrations in stem sap were taken between 8:30 and 9:30 am.
The stem sap was extracted in field using a manual extractor taking five plants per subplot at five phenological stages. The sap was extracted from the lower one-third to one-half of the main stem (stem height at five to six basal internodes). The analysis of
 ${\rm {N - NO}}_3^ - $
and K+ concentrations in sap was carried out with ion selective electrodes (Cadahía, Reference Cadahía2008) using portable tools Horiba LAQUA twin® (Horiba Europe, Leichlingen, Germany) Ion-K+ and Ion
${\rm {N - NO}}_3^ - $
and K+ concentrations in sap was carried out with ion selective electrodes (Cadahía, Reference Cadahía2008) using portable tools Horiba LAQUA twin® (Horiba Europe, Leichlingen, Germany) Ion-K+ and Ion
 ${\rm {N - NO}}_3^ - $
with levels of precision of ±2%. For analysis, an aliquot of 0.5 mL sap extract was taken.
${\rm {N - NO}}_3^ - $
with levels of precision of ±2%. For analysis, an aliquot of 0.5 mL sap extract was taken.
The data were subjected to analysis of variance and Tukey’s multiple comparison test (p > 0.05) was applied to the means. For these analyses, SAS 9.4 software was used (SAS Institute, 2014). In addition, Pearson correlation matrix of the qualitative variables was analyzed, and regression curves were adjusted using the statistical programs InfoStat® version 2014 (National University of Cordoba, Argentina) and Statgraphics® version 2010 (StatPoint Technologies Inc., Warrenton, USA).
Results
NDVI variation by phenological stage and fertilizer rate
During the experiment, rainfall followed a typical seasonal regime, with a low precipitation period (July–September, 0–90 dap) and a high precipitation period (October–November, 90–150 dap) (Figure 1). The variation of monthly air temperatures during the growing season of 2015 had the typical pattern registered in the area (IDEAM, 2017).
For Capiro, the NDVI had an ascending behavior, with significant differences (p < 0.05) obtained among the phenological stages starting from 75 dap (Figure 2a), which coincided with the start of tuberization and increased photosynthetic activity according to Segura et al. (Reference Segura, Santos and Ñústez2006) and Valbuena et al. (Reference Valbuena, Roveda, Bolaños, Zapata, Medina, Almanza and Porras2010). The highest NDVI values of 0.867–0.875 were obtained during maximum tuber filling (150 dap) for the highest fertilizer rates (1900 and 2375 kg ha−1) and corresponded to the highest yields (Figure 2a). In addition, significant differences (p < 0.05) in NDVI were found between the lowest fertilizer rate and the control, which proves that NDVI could be used as an indicator of plant response to fertilization for this cultivar and on this type of soil. These results were similar to the ones reported by Giletto and Echeverria (Reference Giletto and Echeverria2012) and Giletto et al. (Reference Giletto, Zamuner, Silva and Cassino2013), who employed different fertilizer rates and application moments in potato of Group Chilotanum.

Figure 2. Variation of NDVI in cvs. Capiro (a) and Suprema (b) according to crop phenology and in response to four fertilizer rates (kg ha−1): 0 (circle), 1450 (square), 1900 (diamond), and 2375 (triangle). Averages with different letters indicate significant differences between the fertilizer doses at each stage of crop growth according to the Tukey’s test (p < 0.05).
The NDVI values differed according to the cultivar and the stage of growth. Suprema presented a significant increase in NDVI after 125 dap at the start of tuber filling (Figure 2b), with NDVI varying from 0.855 to 0.891. On the other hand, no significant differences were detected in NDVI between the fertilizer doses for this cultivar.
On fertile soils, Capiro responded differently to the different doses of fertilizers, with 2375 kg ha−1 being the optimum fertilizer rate, since this treatment resulted in the highest NDVI (Figure 2a). In Suprema, fertilizer rates had no effect on NDVI values (Figure 2b). On the other hand, the highest increase in NDVI for these cultivars varied according to the growth stage. For Capiro, the maximum increase in NDVI coincided with the highest accumulation of leaf and total dry matter (data not shown), which occurs toward the 120 dap according to Ñústez et al. (Reference Ñústez, Santos and Segura2009). For Suprema, the maximum peak of NDVI coincided with the end of the cycle, with the highest accumulation of total dry matter taking place before 140 dap (Ñústez et al., Reference Ñústez, Santos and Segura2009). Capiro stabilized values of NDVI starting from 100 dap (Figure 2a).
The behavior of NDVI (Figure 2) revealed the presence of significant differences between the cultivars (p < 0.05), indicating the influence of not only edaphoclimatic factors but also genotypic characteristics on NDVI. This was also observed in potato by Schafleitner et al. (Reference Schafleitner, Gutierrez, Espino, Gaudin, Pérez, Martínez, Domínguez, Tincopa, Alvarado, Numberto and Bonierbale2007), who reported the differences in NDVI between cultivars of Group Chilotanum.
Relationship between NDVI and plant yield
A positive correlation was found between yield per plant and NDVI, both in Capiro (r 2 = 0.90) (Figure 3a) and Suprema (r 2 = 0.89) (Figure 3b). However, the NDVI values were higher in Suprema than in Capiro and the highest indices of NDVI corresponded to the highest yields.

Figure 3. Variation of NDVI as related to crop yield (tuber fresh weight, g plant−1) in cvs. Capiro (a) and Suprema (b). The NDVI was measured from the start of main stem development up to the tuber maturation in soils of high fertility, n = 48.
Relationship of NDVI with K+ and
 ${\bf N} - {\bf NO}_3^ - $
concentrations in stem sap
${\bf N} - {\bf NO}_3^ - $
concentrations in stem sap

The moments of highest allocation of assimilates to the tubers were illustrated for each fertilizer rate, since positive correlations (r 2 = 0.83 for Capiro and r 2 = 0.87 for Suprema) were established between K+ concentration in stem sap and NDVI (Figure 4a). The
 ${\rm N - NO}_3^ - $
concentration in stem sap correlated with NDVI only in Suprema (Figure 4b). Although both cultivars had a similar behavior in the concentration of
${\rm N - NO}_3^ - $
concentration in stem sap correlated with NDVI only in Suprema (Figure 4b). Although both cultivars had a similar behavior in the concentration of
 ${\rm N - NO}_3^ - $
in stem sap throughout the growth cycle, ascending to 100 dap and then descending (data not shown), the NDVI in Suprema increased with a decrease in
${\rm N - NO}_3^ - $
in stem sap throughout the growth cycle, ascending to 100 dap and then descending (data not shown), the NDVI in Suprema increased with a decrease in
 ${\rm N - NO}_3^ - $
concentration, which favors the growth of tubers in cultivars susceptible to the excess of N, such as Suprema (Gómez et al., Reference Gómez, Magnitskiy, Rodríguez and Darghan2017). Under the excess of N, a luxury uptake of this element could occur, while tuberization and tuber growth would be reduced (Ruza et al., Reference Ruza, Skrabule and Vaivode2013).
${\rm N - NO}_3^ - $
concentration, which favors the growth of tubers in cultivars susceptible to the excess of N, such as Suprema (Gómez et al., Reference Gómez, Magnitskiy, Rodríguez and Darghan2017). Under the excess of N, a luxury uptake of this element could occur, while tuberization and tuber growth would be reduced (Ruza et al., Reference Ruza, Skrabule and Vaivode2013).

Figure 4. Variation of K+ (a) and
 ${\rm {N - NO}}_3^ - $
(b) concentrations in stem sap as a function of NDVI in cvs. Capiro (black diamonds) and Suprema (white diamonds). The K+ and
${\rm {N - NO}}_3^ - $
(b) concentrations in stem sap as a function of NDVI in cvs. Capiro (black diamonds) and Suprema (white diamonds). The K+ and
 ${\rm {N - NO}}_3^ - $
concentrations were measured from the start of main stem development up to the tuber maturation in soils of high fertility, n = 28.
${\rm {N - NO}}_3^ - $
concentrations were measured from the start of main stem development up to the tuber maturation in soils of high fertility, n = 28.
Discussion
In Suprema, no significant differences were detected in NDVI between the fertilizer doses, possibly, due to an excess of N resulting from initial high level of soil N and N fertilizers, which was reflected in the excessive vegetative growth of Suprema. In fact, Suprema had a low harvest index of 55% as compared to the one of 91% in Capiro. This indicates a reduced efficiency of photoassimilate partitioning in Suprema with a yield of 30 Mg ha−1 that was below its potential yield of 40 Mg ha−1 reported by Ñústez (Reference Ñústez2011).
The increases in NDVI during crop cycle were also reported for other cultivars, where NDVI reaches its peak at flowering, from which it begins to decrease due to the onset of senescence (Govaerts et al., Reference Govaerts, Verhulst and Fuentes2010). However, in the current study, no evidence was found for a reduction in NDVI as the cultivars did not reach leaf senescence at 150 dap. The values of NDVI stabilized in Capiro starting from 100 dap, which could be attributed to the maximum photosynthetic activity of this cultivar coinciding with the stages of highest demand of photoassimilates for tuber filling (Valbuena et al., Reference Valbuena, Roveda, Bolaños, Zapata, Medina, Almanza and Porras2010). These data are likely to be associated with the growth habit of Capiro characterized by determined growth and shorter stems and stolons as compared to Suprema (Ñústez et al., Reference Ñústez, Santos and Segura2009). On the other hand, Suprema progressively increased NDVI starting from 125 dap (Figure 2b) and, apparently, reached its maximum photosynthetic activity at 150 dap, which would indicate that the maximum tuber filling was not obtained at 125 dap due to the indeterminate growth in this cultivar.
As for relationship between NDVI and yield, the major gain in potato dry matter and yield is related to increases in leaf area index up to 100 dap (Castellanos et al., Reference Castellanos, Segura and Ñústez2010), which reaches values of 7.04 for Capiro and 7.89 for Suprema (data not shown), and also in chlorophyll contents and photosynthetic rates. The highest gain in tuber dry matter and yield, in turn, were related to increases in NDVI as evidenced in Figure 3. However, when observing behavior of NDVI in Suprema, its major increase was found toward the end of the cycle (Figure 2b) suggesting that the plants had a capacity to maintain the photosynthetic activity for a longer period of time. This would guarantee the presence of a functional source of photoassimilate production for tuber growth as it presents indeterminate growth, which coincides with stages of tuber filling and higher rate of carbon assimilation (Ñústez et al., Reference Ñústez, Santos and Segura2009). In potato of Group Chilotanum (Giletto et al., Reference Giletto, Zamuner, Silva and Cassino2013), an increase in yield correlated with both leaf dry matter (r 2 = 0.45) and NDVI (r 2 = 0.94), indicating variation in NDVI with increasing doses of N, P, and K rates. NDVI reached 0.85 from 40 dap with variations toward the end of the cycle, whereas NDVI in treatments with lower doses of N and P corresponded to 0.79 and 0.81, respectively (Delgado et al., Reference Delgado, Núñez and Velásquez2004; Giletto et al., Reference Giletto, Zamuner, Silva and Cassino2013). At the same time, a variation in dry matter accumulation depends not only on the fertilizer rates but also on the type of soil employed in the experiment (Graciano et al., Reference Graciano, Goya and Caldiz2004). Then, it might be important to evaluate NDVI in soils of low fertility for cultivars with better adaptation and production potential under these conditions, such as Suprema.
Positive correlations were established between K+ concentration in stem sap and NDVI (Figure 4a); however, for the same value of NDVI, the concentration of K+ tended to be lower and significantly different in Suprema with respect to Capiro. Our results implied that, in both cultivars, K+ concentration in stem sap increased proportionally to the yield (Figures 3 and 4a). This might be also explained as a response to an increase in photosynthesis and respiration that favored the transport of assimilates at tuber filling and the role of K in transport and transformation of sugars as reported in potato (Perrenoud, Reference Perrenoud1993; Westermann et al., Reference Westermann, James, Tindall and Hurst1994).
The decrease in
 ${\rm N - NO}_3^ - $
concentration accompanied by increased NDVI values (later stages of crop cycle) (Figure 4b) could be explained as a flow of
${\rm N - NO}_3^ - $
concentration accompanied by increased NDVI values (later stages of crop cycle) (Figure 4b) could be explained as a flow of
 ${\rm N - NO}_3^ - $
from roots to leaves for further conversion into organic molecules as reported by Coraspe et al. (Reference Coraspe, Muraoka, Ide, Contreras and Ocheuze2009) and Waterer (Reference Waterer2000), with marked reductions in
${\rm N - NO}_3^ - $
from roots to leaves for further conversion into organic molecules as reported by Coraspe et al. (Reference Coraspe, Muraoka, Ide, Contreras and Ocheuze2009) and Waterer (Reference Waterer2000), with marked reductions in
 ${\rm N - NO}_3^ - $
concentrations in potato stems observed toward the end of the cycle. For Capiro, the presence of high
${\rm N - NO}_3^ - $
concentrations in potato stems observed toward the end of the cycle. For Capiro, the presence of high
 ${\rm N - NO}_3^ - $
concentration in stem sap, apparently, was not indicative of the optimal photoassimilate production in plant and could be due to higher N assimilation of Capiro in relation to Suprema. However,
${\rm N - NO}_3^ - $
concentration in stem sap, apparently, was not indicative of the optimal photoassimilate production in plant and could be due to higher N assimilation of Capiro in relation to Suprema. However,
 ${\rm N - NO}_3^ - $
levels in stem sap might serve as indicators of other processes in Capiro, given the fact that nitrate constitutes one of the principal inorganic forms of N absorption in plants, which is rapidly incorporated into molecules of higher molecular weight (McCauley et al., Reference McCauley, Jones and Jacobsen2009) and subsequently transported toward the sinks, mainly tubers (Haddad et al., Reference Haddad, Bani-Hani, Al-Tabbal and Al-Fraihat2016).
${\rm N - NO}_3^ - $
levels in stem sap might serve as indicators of other processes in Capiro, given the fact that nitrate constitutes one of the principal inorganic forms of N absorption in plants, which is rapidly incorporated into molecules of higher molecular weight (McCauley et al., Reference McCauley, Jones and Jacobsen2009) and subsequently transported toward the sinks, mainly tubers (Haddad et al., Reference Haddad, Bani-Hani, Al-Tabbal and Al-Fraihat2016).
The increase in K+ concentration in stem sap was, possibly, due to allocation of photoassimilates to the tubers. This allocation increases during initiation of tuber filling (Ñústez et al., Reference Ñústez, Santos and Segura2009) when K+ is mobilized from leaves and transported through stem toward tubers (Gómez, Reference Gómez2013). A positive correlation between K+ concentration in stem sap and NDVI (Figure 4a) indicates that a variation in NDVI could predict an alteration in K+ transport in potato plants of Group Andigenum. This was reflected in differences in K+ concentrations in stem sap among fertilization doses for Capiro. Capiro is a cultivar of high demand in mineral nutrients and could reach high yields exceeding 50 Mg ha−1 on high fertility soils (Gómez, Reference Gómez2013). As conditions of soil fertility can influence variations in NDVI, it would be important to evaluate NDVI for Capiro on low fertility soils.
Conclusion
This study allowed characterizing NDVI in cultivars Capiro and Suprema on soils of high fertility in potato production region of Bogotá plateau, Colombia. The plants presented a variation in NDVI ranging from 0.836 to 0.875 (Capiro) and from 0.855 to 0.888 (Suprema), with differences obtained between the cultivars due to their disparities in growth habit. The NDVI values were established per phenological stages of growth, thus forecasting productivity and nutrient status in plants in relation to concentrations of K+ and
 ${\rm N - NO}_3^ - $
in stem sap. Such results indicate NDVI as a practical tool for decision making in the region. In addition to finding differences in NDVI behavior between the cultivars, Capiro responded differently to increasing fertilizer rates than Suprema, thus highlighting the genotypic variation in response to rates of mineral nutrients supplied. The behavior of NDVI in potato of Group Andigenum should be further investigated on low fertility soils and in environments for which Suprema is better adapted.
${\rm N - NO}_3^ - $
in stem sap. Such results indicate NDVI as a practical tool for decision making in the region. In addition to finding differences in NDVI behavior between the cultivars, Capiro responded differently to increasing fertilizer rates than Suprema, thus highlighting the genotypic variation in response to rates of mineral nutrients supplied. The behavior of NDVI in potato of Group Andigenum should be further investigated on low fertility soils and in environments for which Suprema is better adapted.
Supplementary materials
For supplementary material for this article, please visit https://doi.org/10.1017/S001447971900005X.
Author ORCIDs
Stanislav Magnitskiy 0000-0002-3715-1932
Acknowledgements
The authors express their gratitude to Ingeplant SAS and the Faculty of Agricultural Sciences, National University of Colombia, Bogotá for research funding and field support. The authors greatly acknowledge the technical support provided by Mr. Elias Silva, Mr. Jonathan Patiño, and MSc. Mauricio Oliveros of the Ingeplant SAS. The authors are also grateful to the grower Mr. Ricardo Rojas for his valuable help in the field during the development of the research.







