


INTRODUCTION
Enset [Ensete ventricosum (Welw.) Cheesman (Musaceae)] has been appointed ‘the tree against hunger’ and is a straightforward way to facilitate for people to achieve food and livelihood security in parts of Africa (Brandt et al., Reference Brandt, Spring, Hiebsch, McCabe, Tabogie, Diro, Wolde-Michael, Yntiso, Shiegeta and Tesfaye1997; Negash and Niehof, Reference Negash and Niehof2004). Ensete is close to, but distinctly separated from, the genus Musa (Bekele and Shigeta, Reference Bekele and Shigeta2011). Enset is an important multi-purpose and drought-tolerant crop, used for food (e.g. Tsegaye and Struik, Reference Tsegaye and Struik2001), fodder (e.g. Funte et al., Reference Funte, Negesse and Legesse2010), fibre (Tsehaye and Kebebew, Reference Tsehaye and Kebebew2006) and traditional medicine (Nyunja et al., Reference Nyunja, Onyango and Erwin2009). The food is rich in starch, is a good source of calcium and iron (Abebe et al., Reference Abebe, Bogale, Hambidge, Stoecker, Bailey and Gibson2007; Atlabachew and Chandravanchi, Reference Atlabachew and Chandravanchi2008) and has overall nutritive values similar to potato (Mohammed et al., Reference Mohammed, Gabel and Karlsson2013). In terms of edible dry weight and energy, enset gives higher yield than other crops cultivated in Ethiopia (Tsegaye and Struik, Reference Tsegaye and Struik2001). Enset is most commonly grown in home gardens, frequently intercropped with peas or beans (Abebe et al., Reference Abebe, Wiersum and Bongers2010). Enset is suitable for agroforestry (Negash et al., Reference Negash, Yirdaw and Luukkanen2012; Tesfaye, Reference Tesfaye2008), which is suggested for land improvement (Lahmar et al., Reference Lahmar, Bationo, Lamso, Guéro and Tittonell2012). Taro, a crop frequently intercropped with enset in Ethiopia, performs well in shadow (Sanou et al., Reference Sanou, Bayala, Brazié and Teklehaimanot2012); this combination could be a suitable option in areas where taro, but not enset, is currently cultivated.
The vegetative growth habit of enset is similar to banana (Musa spp.) plants, but enset is not grown for the fruits; these contain mostly large and very hard seeds (Karlsson et al., Reference Karlsson, Tamado, Dalbato and Mikias2013a). Plants for food production are usually harvested at the onset of flowering, and thereafter the large pseudostem and corm are decorticated for further processing (Pijls et al., Reference Pijls, Timmer, Wolde-Gebriel and West1995). Enset has been cultivated since ancient time and there are hundreds of landraces (Bizuayehu, Reference Bizuayehu2008). Areka Research Centre (ARC), the national centre for enset research in Ethiopia, has 122 landraces (Bekele et al., Reference Bekele, Diro and Yeshitla2013) and in 2011 six cultivars were registered, certified and nationally released: Endale, Gewada, Kelisa, Mesena, Yanbule and Zerita (ARC database by Mikias Yeshitla).
Enset contributes positively to the local environment by improving the nutrient balance and increasing the fraction of organic matter in soil (Elias et al., Reference Elias, Morse and Belshaw1998; Tensaye et al., Reference Tensaye, Lindén and Ohlander1998). It is a part of farming systems with high biodiversity (Tesfaye, Reference Tesfaye2008) which is environmentally sustainable. Each plant requires some years to reach harvest stage, and enset is therefore usually grown by having generations of plants mixed, thus being a reliable food source over time (Brandt et al., Reference Brandt, Spring, Hiebsch, McCabe, Tabogie, Diro, Wolde-Michael, Yntiso, Shiegeta and Tesfaye1997; Dalbato, Reference Dalbato and Lagibo2000). Since climate changes will probably make droughts even more unpredictable in sub-Saharan Africa, measure should be taken in rural communities to prepare for the future (Cooper and Coe, Reference Cooper and Coe2011; Meadovs, Reference Meadovs2011). As enset performs well in areas where the natural environment is steppe (Funte et al., Reference Funte, Negesse and Legesse2010; Negash et al., Reference Negash, Yirdaw and Luukkanen2012) and naturally occurs in forests and in humid areas (Baker and Simmonds, Reference Baker and Simmonds1953) it is suitable to grow in a wide range of climates, thus in large geographical areas.
Enset is propagated by burying the corm (underground stem) after removing the apical meristem. In addition, there are different traditional treatments of the corm: keep it entire or split it to various extents; all farmers claim that they use the best method, although the used methods vary (personal observation). A study of one landrace indicates that splitting gives more but smaller suckers (Diro et al., Reference Diro, Gebremariam, Zelleke and van Staden2003) and Diro (Reference Diro1997) suggested watering on newly planted corms; an action that is believed to cause rotting. It is very important that recommendations are fact-based, reliable and beneficial for the user (Blackstock et al., Reference Blackstock, Ingram, Burton, Brown and Slee2010); therefore, there is a need of controlled studies of traditional enset propagation methods.
The purposes of this study were to increase the knowledge on enset propagation by (i) comparing traditional corm pre-treatments, (ii) comparing the six released cultivars and (iii) investigating the effect of watering on newly buried corms, using emergence rate and cormling number and characteristics as response variables. We define ‘cormling’ as a new sprout from a corm, still attached to the parent corm, thus a more narrow definition than ‘sprout’, ‘sucker’ or ‘sapling’, c.f. ‘seedling’.
MATERIAL AND METHODS
In October 2010, an experimental plot was prepared at Wolaita Sodo University campus (N 06°50′00″, E 37°45′07″, 1882 m asl). Wolaita Sodo is about 300 km south of Addis Ababa in Ethiopia, in a region where enset is grown traditionally. The plot was circa 25 × 25 m, located in a flat part of hilly environment. The area was used for extensive grazing until the university was established in 2006. In the experimental plot, soil was cultivated by hand, using traditional equipment. The plot was surrounded by a fence being 180 cm of which circa 30 cm was dug down in soil and a ditch being 50 cm wide and 30 cm deep.
On January 12, 2011, corms from 63 enset plants (the plants had grown for two years after being detached from parent corms) were uprooted at ARC (N 07°04′02″, E 37°41′22″, 1785 m a.s.l.) in Ethiopia. The plants belonged to the six certified cultivars of enset and had been reproduced vegetatively at ARC for many generations. Nine corms of each cultivar plus nine extra of Zerita were dug up after selecting equal sized plants (average pseudostem base circumference was 89.5 [SD 11.0] cm). In addition, a number of extra corms were collected to use as border plants. Roots were cut off close to the corm and the pseudostem was cut off horizontally 18 cm over the lowest attachment point to the corm; this pseudostem piece was kept and later buried still attached on the corms, following the practice at ARC. Corms, width attached 18 cm pseudostem, weighted on average 8.4 (SD 2.4) kg, and were 58.5 (SD 6.6) cm in circumference, measured on the widest part of the corm. They were individually packed in aerated plastic sacks and placed, in upright position, in the experimental plot overnight.
On January 13, corms were randomly allocated to one of three treatments: (i) keeping entire, (ii) split in two pieces, (iii) split in four pieces. The nine extra corms of Zerita were used to investigate the effect of watering in a separate part of the experimental plot. There were three replicating corms of each cultivar to each treatment. Treatments of corms followed traditional practice, using a large knife, sharp in the point and along both edges (Pijls et al., Reference Pijls, Timmer, Wolde-Gebriel and West1995). Corms to keep entire had the apical meristem (i.e. the point of growth) removed using the point of knife from top of the remaining pseudostem part, while those to split were first cut in pieces and then the apical meristem was cut away.
On January 14, corms were buried in holes dug to 40 cm deep and 50 cm wide; 150 cm between the centres of two neighbour holes. Bottom of holes was refilled with 10 cm softened top soil and watered with 5 L per hole (soil was very dry at this time). The corms were placed thereon and on them 15 L of 50/50% mix of dry cow dung (manure) and soil. All pieces of a split corm were placed in the same hole (not in contact with each other). Soil and manure characteristics (Table 1) were analysed by Sodo Soil Testing Laboratory. Weather conditions (Figure 1) were recorded, by the National Meteorology Agency of Ethiopia, from a nearby weather station (N 06°49′16″, E 37°44′56″, 1863 m asl), i.e. in the same valley as the experimental plot, SSW of Sodo town.
Table 1. Characteristics of local soil and dry cow dung used to enset (Ensete ventricosum) corm sprouting experiment in southern Ethiopia.
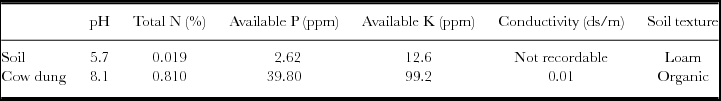

Figure 1. Daily weather in Wolaita Sodo, Ethiopia, January–October 2011; occasions for enset corm burial, emergence period of first sprout (from each of 63 corms) and harvest indicated. Weather data recorded and provided by the National Meteorology Agency.
From January 14 and onward, the corms allocated to watering were given 5 L per corm every day if not equal amount of rain came. For not watered corms, it was decided to give 5 L water per corm twice a week during the expected belg rain period (‘belg rain’ is the small rainy period that often comes in February–March in this area) in case there was no natural rain. This watering was applied one week (from March 6th) before rain begun. The experimental plot was visited daily, weeding was done when necessary and sprouting was recorded every day.
Cormlings were harvested and recorded in October, nine months after corm burial. Harvesting was done by uprooting the original corm and lay the group of cormlings down. Thereafter, each cormling was carefully detached from the parent corm and recorded. Recorded characteristics were: (1) height of main part of pseudostem (from corm connection until the point where the stalk of the lowest green, at least 50% green, leaf detached from the other), (2) pseudostem circumference, i.e. circumference in the middle of the height measured in 1, (3) number of developed leaves being more than 50% green and number of developed leaves being less green, (4) leaf height of the longest leaf, (5) blade width of the longest leaf and (6) corm circumference and height, which were used to calculate corm volume by approximating the corm to be a half globe downward and a cone upward.
Records 1–6 were done for all aboveground, green cormlings. There were also underground cormlings, for these, only leaf height (=plant height) and corm size were measured. The average of records for aboveground cormlings was calculated separately for the seven largest cormlings and for the remaining smaller from each parent corm. For measurement of overall production, (1) and (2) were used to calculate pseudostem volume (approximated to a cylinder), these values were summarized as total volume per parent corm.
For each parent corm, fresh weight was recorded from the cormling with the fourth largest pseudostem circumference, for the one being closest to the average and for the one closest to the median pseudostem circumference.
The performance records (described above) were analysed with two-way ANOVA: (i) 6 cultivars × 3 corm pre-treatments and (ii) watered or not × 3 pre-treatments. Specific comparisons were done with Tukey HSD Post-Hoc test with p < 0.05 as limit for significance. Correlation analyses of total pseudostem volume and (1) parent corm diameter and (2) time to first emergence were performed for not watered parent corms. All analyses were done with Statistica (StatSoft, 2011).
In addition, 30 randomly selected cormlings were used to record dry weight of corms, leaves (blade and stalk together) and the lowest 15 cm of pseudostem. After recording the fresh weight, these plant parts were cut in smaller pieces, air dried in a ventilated room for about two weeks and then dried in oven at 80 °C for at least 24 h, checked for constant weight before the dry weight was recorded with a precision of 0.01 g.
RESULTS
Time to emergence was 50 days or more regardless of watering, corm splitting or cultivar (Figure 2A). Time was generally longer for entire corms than for split corms (Figure 2A, Table 2). Watering gave the shortest average time to emergence and levelled out the difference otherwise present due to splitting of corms (Figure 2A, Table 2). Within 112 days from burial, emergence occurred from all 63 buried corms (Figure 1).
Figure 2. Time to first emergence and records of new sprouts (‘cormlings’) from corms (N = 3) of Ensete ventricosum, nine months after corm burial, in southern Ethiopia. Corms to bury were handled following different traditional techniques shown with symbols: circle – kept entire, quadrate – split in two pieces, and triangle – split in four pieces. Six cultivars were included and one, Zerita*, was also watered. Symbol colours: (B) white – aboveground, black – belowground, (C) white – 4th largest sprout, grey – average sprout, black – median sprout.
Table 2. ANOVA results (F-values and p significance notation from 38 analyses: ns p > 0.05, *p < 0.05, **p < 0.01, ***p < 0.001) when six cultivars1 of enset (Ensete ventricosum) were subjected to three different pre-treatments of parent corms in a full-factorial experiment in southern Ethiopia. Emergence and performance of sprouts (‘cormlings’) were recorded. One cultivar (Zerita) was used to investigate the effect of watering on buried corms subjected to the three pre-treatments.
1Endale, Gewada, Kelisa, Mesena, Yanbule and Zerita.
There were totally 3669 aboveground and 736 belowground cormlings (Figure 2B). The total number of shoots was higher from split than entire parent corms, and Endale produced more cormlings, above as well as below ground, than any other cultivar (Figure 2B, Table 2). The number of aboveground shoots did not differ between watered or not, but there were more belowground shoots when watered (Figure 2B, Table 2). From all cultivars, there were aboveground individuals that reached over 4 m leaf length and corm size of 0.5 dm3 (Figure 3D, F). Belowground cormlings were on average 21.8 cm (SD 14.3) in leaf height and had 0.0016 dm3 (SD 0.0033) corms.
Figure 3. Recorded characteristics of new Ensete ventricosum sprouts (‘cormlings’), nine months after corm burial, in southern Ethiopia. Corms to bury were handled following different traditional techniques shown with symbols: circle – kept entire, quadrate – split in two pieces, and triangle – split in four pieces (N = 3 corms, totally 3669 cormlings). Six cultivars were included and one, Zerita*, was also watered. Symbol colours: white – average of the seven largest cormlings per original corm, black – average of remaining cormlings from each corm.
Fresh weight of individual cormlings differed between cultivars, with Yanbule and Endale generally being the heaviest and lightest, respectively (Figure 2C, Table 2). Weight of the fourth largest cormling was less for entire parent corms than for split corms and less for watered than for not watered (Figure 2C, Table 2). The fresh weight of an average cormling was larger for entire corms than for split corms, as for median cormlings in the watering treatment (Figure 2C, Table 2). Water content was high: corms 89.8% (SD 3.2), leaves 94.9% (SD 0.7) and the lowest 15 cm of pseudostem 93.8% (SD 1.2).
The total pseudostem volume per parent corm was largest for Yanbule and least for Mesena, and entire parent corms produced less than split ones (Figure 2D, Table 2). Watering gave on average less volume but also levelled out the differences between corm pre-treatments (Figure 2D). There was no correlation between diameter of parent corm and produced total pseudostem volume (p > 0.05, R 2 = 0.07), while there was a correlation between time to emergence and produced total pseudostem volume (p < 0.05, R 2 = 0.390).
The pseudostem circumferences of the 189 weighted cormlings were linearly correlated to the square root of fresh weights (R 2 = 0.933), and therefore the record of pseudostem circumference was used to rank the cormlings, for each original corm, from the largest to the smallest, before analysing the seven largest and the smaller separately.
In addition to the new, developing, leaf in the centre, cormlings had 0–16 developed leaves (Figure 3C), of which 45.6% (SD 16) were green to more than 50% of the blade. The general performance of individual cormlings was cultivar-specific, with significant effects frequent due to cultivar but rarely to splitting (Figure 3, Table 2). Of the six characteristics in Figure 3, splitting affected pseudostem height and leaf height; in both cases split parent corms gave the higher result (Figures 3B and 3D, Table 2).
DISCUSSION
Emergence from buried corms was 100%, which is not expected among farmers nor by ARC where about 75% is common (Diro, Reference Diro1997). All corms were buried using the establish advice to dig a large hole and re-fill part of it with softened top-soil before placing the corm and cover with soil. Since this is a hard work, the recommendation may not always be followed, which may partly explain why less than 100% emergence is common. The cormlings became larger than expected within the allocated time period (c.f. Diro et al., Reference Diro, Gebremariam, Zelleke and van Staden2003 and Figures 2 and 3). Farmers usually apply manure by putting it on the soil surface, not directly on the corm, at burial. The belief is that manure in contact with the corm will cause rotting. In this study, manure was applied directly on corms, to give good access to nutrients and to limit weed growth. Obviously, dry cow dung mixed with soil on the corm at burial does not have negative effect on emergence or cormling development (Figure 2); instead, it may explain the unusual successful emergence and vigorous growth.
All cormlings from each parent corm were measured, thus avoiding the selection bias that most probably occurs if trying to draw a random sample from dense groups with 20–100 sprouts per original corm (Figure 2). The record for the seven largest cormlings per original corm was calculated separately since the number of smaller differed (Table 2) and could blur the result of the larger individuals. The use of cormlings with median and average weight of each original corm gives an impression of the weight distribution; if the median is smaller than the average there are many small and few large and vice versa (Figure 2C).
Time to emergence and watering effect
The first sprouts, from not watered as well as from watered corms, came 50 days after burial and soon after onset of belg rain. Thus, the moisture stored in corms was enough to prevent drying out until emergence. When main rain begun, in early May as expected, all parent corms had sprouted (Figure 1). Watering generally decreased time to emergence and levelled out the differences between entire and split parent corms, while splitting of corms stimulated emergence for not watered corms (Figure 2A, Table 2). This is contradictory to the local belief telling ‘it is impossible to water on newly buried corm, because the added water cause rotting’. Our results underline the need, in addition to gather indigenous knowledge to learn about agricultural practice, to investigate the accuracy of the information provided by users.
The belg rain came just before cormlings emergence begun (Figure 1). Therefore, the following growth should not be used for far-reaching conclusions about growth in response to watering. The growth differences between cormlings from watered and not watered corms are probably mainly a result of intra-clone competition (Benomar et al., Reference Benomar, DesRochers and Larocque2012). Watering after corm burial until emergence was successful by leading to equal emergence and therefore to production of even cormlings (Figures 2 and 3), while effect of watering after emergence and during growth remains to be studied.
Corm pre-treatment techniques and early growth of suckers
Time to emergence was longer for entire than for split corms (Figure 2A) and time to emergence was correlated to total pseudostem production per parent corm. Therefore, the fact that the fourth largest cormling and the average of the seven largest per parent corm was slightly less for entire than for split corms (Figures 2C, 3B and 3D, Table 2) may be partly due to emergence time and the following competition between sprouts from different original corms. We used the traditional spacing of 1.5 m between centres of holes for corm burial, but due to unexpected large cormlings, the stand became relatively dense. However, the size differences can be understood by one leading cormling per corm piece, not per parent corm. For pseudostem circumference (correlated to weight) there was no significant effect of corm pre-treatment for any of the two size groups (Figure 3A, Table 2), but the weight of average cormlings was larger from entire than from split corms (Figure 2C, Table 2). We conclude that a number of large cormlings will arise regardless of corm pre-treatment, and splitting of parent corms leads to a larger number of cormlings in the smaller sizes. The total production from split corms was therefore larger than from entire corms (Figure 2D).
Generally, splitting of corms was beneficial for production. However, in areas where extended droughts are common and watering is impossible, it may be preferable to bury entire corms, in order to utilize the corm water holding capacity as much as possible.
Cultivars
As the six certified and released cultivars have went through tests for growth at different environments (under the control of ARC), it is not surprising that they all performed well at Wolaita Sodo; an area where enset is grown traditionally. The cultivars responded similarly to splitting and had similar distribution of cormling sizes (Figure 3). However, the number of cormlings per parent corm differed substantially (Table 2), with Endale having most and Mesena fewest (Figure 2B). The number of cormlings emerged of the landrace Halla (Diro et al., Reference Diro, Gebremariam, Zelleke and van Staden2003) was similar to Mesena and Kelisa (Figure 2B). Yanbule, having intermediate number of cormlings (Figure 2B), had the largest total production in terms of pseudostem volume (Figure 2D).
The number and sizes of cormlings are one factor to be taken into account when selecting cultivar, while the specific performance of a plant depends on growth conditions (Tsegaye and Struik, Reference Tsegaye and Struik2003; Zipper, Reference Zipper, Huber, Sinclair and Lampe2005). When planting, farmers’ decision of landrace is based on intended use and taste preference. However, when enset is introduced to groups of people not used to it, cultivars can be chosen to be most suitable to the local need and environment.
CONCLUSIONS
Propagation of enset in the field was successful for all included cultivars, with size and number of cormlings being correlated to cultivar, corm-pre-treatment method and watering. Extended and intensified use of enset requires efficient productions of cormlings for distribution and planting. Our study showed that vegetative propagation, using traditional techniques and good agricultural practice, was efficient. The techniques we used should be clearly communicated to enset growers, ensuring successful local further propagation. Micropropagation, sometimes suggested for enset, requires special laboratory equipment and advanced controlled growth conditions during extended time after initiation of propagation (Birmeta and Welander, Reference Birmeta and Welander2004). On the contrary, vegetative propagation from buried corms in the field resulted in healthy and strong cormlings, ideal for re-planting, nine months after burial (Figures 2 and 3). These results were achieved without any need for advanced equipment or controlled environment.
Despite the many landraces with different characteristics (Bekele et al., Reference Bekele, Diro and Yeshitla2013) there are requests of future improvement of cultivars. Research is ongoing on seed germination (Karlsson et al., Reference Karlsson, Tamado, Dalbato and Mikias2013a) and seedlings can flower within two years from germination (Karlsson et al., Reference Karlsson, Tamado, Dalbato and Mikias2013b). Combined with vegetative production from corms (Figures 2 and 3), this is promising for conventional breeding through crossing and selection; a low-cost and simple technique not yet applied on enset.
Within the small-scale kind of cropping systems where enset directly can contribute to food and livelihood security, propagation can efficiently be done in fields (Figures 2 and 3). Use of well-known clones, with desired characteristics, results in suitable cormlings for distribution. We advocate further studies of cultivation methods, as the 100% emergence recorded is unusual and the achieved cormlings were unexpected large. The most efficient techniques should be revealed to further improve the enset production.
Acknowledgements
We are thankful to Yacob Gota, Bergene Ade, Zemach Chelo and Getachew Kefita for contributing with their skills and practical work. We thank Formas (The Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning) for funding this project, and Areka Research Centre for generously providing corms.





