


INTRODUCTION
The extraction of forest products is an important livelihood strategy in developing countries (Stoian Reference Stoian2005; Angelsen et al. Reference Angelsen, Jagger, Babigumira, Belcher, Hogarth, Bauch, Borner, Smith-Hall and Wunder2014), where hunting forest vertebrates is widespread and bushmeat is an important source of both animal protein and cash income (Milner-Gulland et al. Reference Milner-Gulland and Bennet2003; de Merode et al. Reference de Merode, Homewood and Cowlishaw2004). However, hunting can threaten the population viability of game species, particularly large-bodied mammals and birds, driving widespread depletion and local and regional extinctions of the most vulnerable taxa (Peres & Palacios Reference Peres and Palacios2007). Since these animals play key roles in ecological processes, including seed dispersal and herbivory, overhunting can not only compromise the livelihoods of local people, but also affect the integrity of tropical forests and the long-term persistence of non-hunted biodiversity (Stoner et al. Reference Stoner, Vulinec, Wright and Peres2007; Jorge et al. Reference Jorge, Galetti, Ribeiro and Ferraz2013). Understanding the drivers of both hunting and bushmeat consumption can help us to identify interventions to reduce unsustainable hunting whilst also seeking to avoid negative impacts on the well-being of those most dependent on bushmeat for nutrition.
Bushmeat consumption and hunting decisions are in part determined by the perceived returns compared to other activities or alternatives (Behrens Reference Behrens1992; Ling & Milner-Gulland Reference Ling and Milner-Gulland2006). These returns can be highly variable across space because they depend on landscape characteristics (i.e. spatial attributes that vary among locations) that determine either bushmeat supply, by influencing the availability of game species (i.e. forest cover), or bushmeat demand, by influencing the availability of domesticated meat (i.e. remoteness). Hence, landscape context surrounding households may be at least as important as cultural (i.e. ethnicity) and socioeconomic (i.e. wealth, income and education) characteristics of individuals and households in driving bushmeat consumption and hunting (Nyahongo et al. Reference Nyahongo, Holmern, Kaltenborn and Roskaft2009; Brashares et al. Reference Brashares, Golden, Weinbaum, Barret and Okello2011). However, most studies on bushmeat consumption and hunting have focused on individual preferences or the interplay of demographic (i.e. household size), socioeconomic (i.e. wealth and income) and cultural (i.e. ethnicity) characteristics of individuals or households (e.g. Godoy et al. Reference Godoy, Undurraga, Wilkie, Reyes-García, Huanca, Leonard, NcDade, Tanner and Vadez2010; Rentsch & Damon Reference Rentsch and Damon2013). Only recently have researchers begun to examine the importance of landscape characteristics associated with bushmeat supply and demand (Brashares et al. Reference Brashares, Golden, Weinbaum, Barret and Okello2011; Foerster et al. Reference Foerster, Wilkie, Morelli, Demmer, Starkey, Telfer, Steil and Lewbel2012; Mgawe et al. Reference Mgawe, Mulder, Caro, Martin and Kiffner2012). Indeed, unaccounted variation in landscape context may partially explain the distinct results regarding the effects of household wealth and income on bushmeat consumption and hunting among previous studies (e.g. Wilkie & Godoy Reference Wilkie and Godoy2001; Wilkie et al. Reference Wilkie, Starkey, Abernethy, Effa, Telfer and Godoy2005; Fa et al. Reference Fa, Albrechtsen, Johnson and Macdonald2009).
Landscape context is expected to be particularly important in dynamic and heterogeneous regions such as those found along deforestation frontiers and in relatively recent post-frontier areas in the tropics; that is, former agricultural frontier areas where deforestation rates are presently lower (Rodrigues et al. Reference Rodrigues, Ewers, Parry, Souza, Veríssimo and Balmford2009). In this study, we evaluate the influence of two landscape characteristics, namely forest cover and remoteness (distance to urban centres), on bushmeat supply and demand, in driving bushmeat consumption and hunting within households across a heterogeneous post-frontier tropical region in eastern Amazonia.
On the one hand, as predicted by a large body of theoretical work on the effects of habitat loss (Andrén Reference Andrén1994; Fahrig Reference Fahrig2003), higher forest cover remaining in the landscape positively affects the diversity and abundance of game species (Chiarello Reference Chiarello1999; Peres Reference Peres2001; Michalski & Peres Reference Michalski and Peres2007; Sampaio et al. Reference Sampaio, Lima, Magnusson and Peres2010; Prist et al. Reference Prist, Michalski and Metzger2012; Ochoa-Quintero et al. Reference Ochoa-Quintero, Gardner, Rosa, Ferraz and Sutherland2015). Although hunting can transform preserved continuous forest in ‘empty forests’ (Redford Reference Redford1992), forest loss and fragmentation increase both the access to forests and the effects of hunting (Peres Reference Peres2001), making populations in smaller forest patches particularly vulnerable to hunting. In summary, forest cover is associated with a higher availability of game species, thus increasing bushmeat supply to hunters (Fig. 1). On the other hand, landscape remoteness, measured by the distance from urban centres, is related to a decrease in the availability of domesticated meat, potentially increasing bushmeat demand (Fig. 1). In more remote areas, the availability of domesticated meat is frequently reduced given the distance from markets, which increases the cost and time of transportation, leading to less frequent trips to urban centres (Parry et al. Reference Parry, Day, Amaral and Peres2010a). In addition, bushmeat tends to be cheaper (Parry et al. Reference Parry, Day, Amaral and Peres2010a; Brashares et al. Reference Brashares, Golden, Weinbaum, Barret and Okello2011) and domesticated meat more expensive (Wilkie & Godoy Reference Wilkie and Godoy2001; Brashares et al. Reference Brashares, Golden, Weinbaum, Barret and Okello2011), making access to the latter limited in remote areas. We therefore hypothesize that both bushmeat consumption and hunting are influenced by landscape context and are more likely in landscapes (i) with higher forest cover and (ii) further from urban centres (Fig. 1).

Figure 1 Expected relationships of landscape context and bushmeat consumption and hunting.
To test these hypotheses, we focused on a heterogeneous, c. 1 million-ha area south of the city of Santarém in the Brazilian Amazon and conducted interviews in 262 households across 16 hydrological catchments (c. 5000 ha each) that captured the variation in forest cover, human population density and distance to urban centres resulting from rapid agricultural expansion across the region (Moran Reference Moran1993; Fearnside Reference Fearnside2005). Landscape contexts of sampled households were thus extremely variable and well suited to teasing apart the relative importance of different landscape correlates of bushmeat consumption and hunting activity. Specifically, we examined the association of bushmeat consumption and hunting with two landscape characteristics: (1) forest cover (total and per household), measured at distinct spatial scales and considering different types of forest; and (2) remoteness, measured by either distance to or the time spent to get to urban centres.
METHODS
Study area
We conducted this study in the eastern Brazilian Amazonia, Pará state, in a region of c. 1 million ha (Fig. 2), encompassing the rural areas of the municipalities of Santarém, Belterra and Mojuí dos Campos. The study region is home to both recent and long-term in-migrants from various regions of Brazil and encompasses rural properties that range from small-scale farms based on subsistence agriculture to large-scale soy farms and cattle ranches (Appendix S1, available online), meaning that the importance of bushmeat consumption and hunting should be extremely variable across the region. The different types of properties are scattered in the region, with no correlation between property size and distance to the city of Santarém (Person's correlation = 0.10).
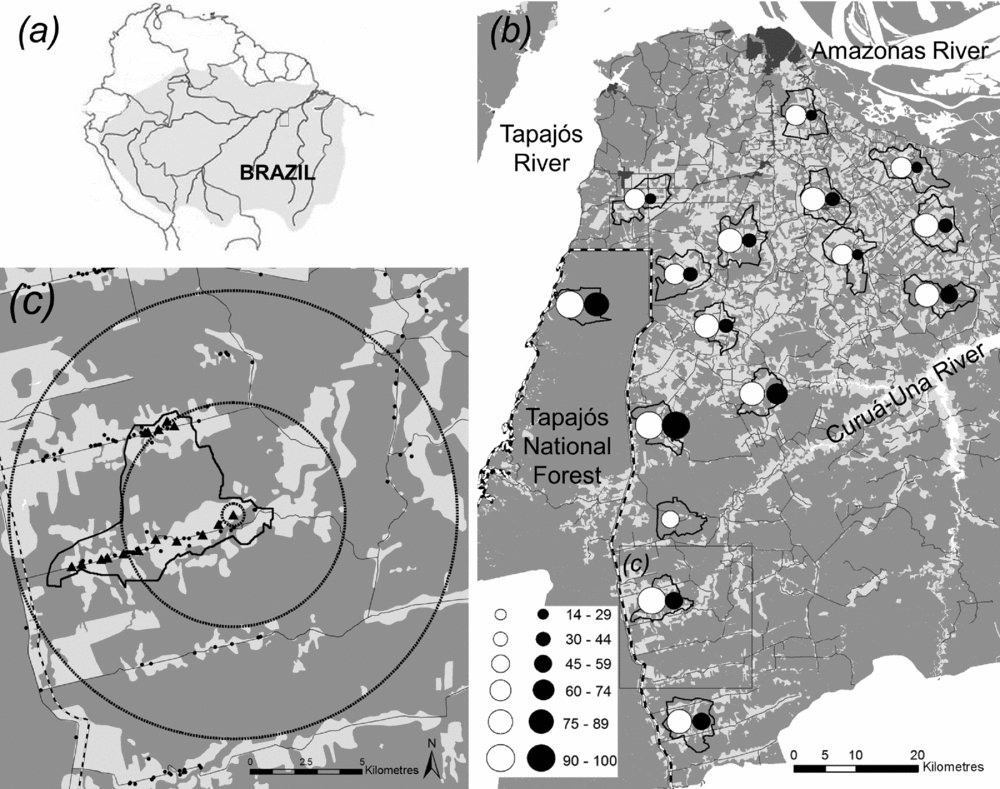
Figure 2 Land-cover map of the study region, with percentages of households where bushmeat was consumed and that had a hunter. (a) Location of the study region in Amazonia. (b) Land-cover map of the study region indicating the 16 sampled hydrological catchments (black polygons). The sizes of circles represent the percentages of sampled households in a catchment where bushmeat was consumed (white) and that had a hunter (black). Urban areas are in dark grey, forests are in grey, converted land is in light grey and water bodies are in white. (c) Distribution of households (sampled – triangles; not sampled – dots) within a catchment (solid black line) and the indication of the three buffers for calculating forest cover for a given household. Solid grey lines are roads.
Sampling design
This study is part the Rede Amazônia Sustentável (RAS; the Sustainable Amazon Network), which aims to assess social and ecological dimensions of land-use sustainability in eastern Brazilian Amazonia (Gardner et al. Reference Gardner2013). The sampling design was hierarchical, first selecting hydrological catchments that captured the variability in forest cover within the study region and then a stratified random selection of rural properties (and households) within them. In total, we sampled 16 catchments comparable in size (c. 5000 ha each), distributed across a gradient of current forest cover (24–98%), population density (0.25–34 households/km2) and straight-line distance to the nearest urban centre (5–92 km) (Fig. 2). There were 262 households within them. For more details on household selection, see Appendix S1.
Bushmeat consumption and hunting
We used an interview-based survey with the heads of sampled households (see Appendix S1). Prior to the beginning of interviews, participants had the research aims explained to them, as well as the fact that their participation was voluntary, that they could withdraw at any time and that information would be used anonymously. We then obtained verbal consent from each individual who was willing to participate. We did not ask for written consent because most people were illiterate.
The complete RAS survey included a large set of questions regarding different aspects of land ownership, land use and agricultural production. Questions about bushmeat consumption and hunting were included in a section on forest use and were designed to: (1) be straightforward, given their inclusion in a longer questionnaire; and (2) avoid sensitive topics on bushmeat trade as, although hunting for subsistence is not illegal in Brazil, trading bushmeat is. The data used here are derived from two questions: (1) in which month bushmeat was last consumed in the household; and (2) if there was at least one hunter in the household (even if only hunting occasionally). Bushmeat consumption and hunting were equal to 1 when, respectively, bushmeat was consumed at least once in the previous 12 months and there was at least one hunter in the household.
The recall period of 12 months for bushmeat consumption aimed to minimize the influence of seasonal variation in both hunting activity and bushmeat consumption (Golden et al. Reference Golden, Wrangham and Brashares2013). However, we also calculated if bushmeat was consumed at least once in the previous 30 days and results did not change (Table S1). The presence of at least one hunter in the household was adequate for capturing the variability in hunting, given that this activity was not widespread in the study region (with c. 60% of households not having a hunter; see ‘Results’ section). Indeed, hunting frequency (number of hunting trips per month), as assessed by one of the questions in the survey, was low (with a median value of two).
Landscape characteristics
We quantified the percentage of forest cover, which has been associated with the presence and abundance of game species in the Santarém region (Sampaio et al. Reference Sampaio, Lima, Magnusson and Peres2010) and across the studied catchments (Moraes Reference Moraes2016). Forest cover was calculated considering either: only non-degraded primary forest; all primary forests (degraded and non-degraded); or both primary forests and secondary forest older than 10 years. We accounted for different types of forest because the abundance of ungulate browsers and some rodent species can be higher in secondary forests in the Amazon (Parry et al. Reference Parry, Barlow and Peres2007; Parry et al. Reference Parry, Barlow and Peres2009a), while the total supply of game meat (total biomass) is usually higher in primary forests (Parry et al. Reference Parry, Barlow and Peres2009a). For each sampled household, we computed forest cover at three spatial scales: (1) a smaller scale of a 500-m radius buffer (0.78 km2), representing the immediate surroundings of the household within the property and neighbouring properties (median of property size in the region was 0.25 km2); and at two wider spatial scales of (2) a 5-km radius (78.5 km2) buffer; and (3) a 10-km radius (314 km2) buffer, given that local species persistence and abundance depend on the wider landscape context (Andrén Reference Andrén1994) (Fig. 2). Because we are interested in the association between forest cover and game populations, we also calculated forest cover per household (percentage of forest cover divided by the number of households in the landscape) to take into account human population size and associated hunting pressure from local residents. We calculated this only at the two wider spatial scales, because human population density varied little at the smaller spatial scale.
We calculated the cover of each type of forest through a time series of Landsat images from 1990 to 2010 using a decision tree classification procedure that separated primary and secondary forest, as well as primary forest with a signal of degradation from past logging and fire (Gardner et al. Reference Gardner2013). To calculate the number of households, we used a statistical grid based on the 2010 census data from the Brazilian Institute of Geography and Statistics (IBGE 2016). This grid is divided in 1 × 1-km cells in rural areas and 200 × 200-m cells in urban areas, with the number of households in each cell calculated based on households’ addresses or coordinates. When missing, households’ locations were estimated based on the distribution of roads and types of land use (IBGE 2016) (see Appendix S1 for more information).
We quantified the distance to urban centres using five different measures: straight-line distance of households to the (1) largest, (2) nearest and (3) most visited urban centre (the latter being reported in the interviews) and time spent to get from the household to the most visited urban centre (4) in the dry season and (5) in the wet season (both reported in the interviews). Cattle ranches in the study region supply local urban markets or export to other municipalities, states and countries (Appendix S1). Hence, most of the consumed domesticated meat in the study region is bought in markets and stores (Torres Reference Torres2014), which are uncommon in rural areas and offer more expensive prices far from urban centres. Distance to large urban centres should thus shape the access to domesticated meat. The largest urban centre is the city of Santarém, with two other smaller urban centres from the municipalities of Belterra and Mojuí dos Campos also present within the study region (Appendix S1).
Data analysis
Our analyses considered three landscape predictors: forest cover within the property and nearby properties; forest cover within the wider landscape; and remoteness. Our first step was to select the measure of each of the three landscape predictors that best explained bushmeat consumption and hunting using a model selection approach based on the Akaike information criterion modified for small samples (AICc). For each dependent variable (bushmeat consumption (0/1) and hunting (0/1)), we compared a candidate set of simple models, each containing a different measure for a given landscape predictor, and selected the measure contained in the first-ranked model (Table S2). To account for the hierarchical nature of the sampling design, we used generalized linear mixed-effects models (GLMMs), considering the 16 hydrological catchments as a random factor and modelling the dependent variables as binary variables using logit as the link function. Fixed factors (i.e. the different measures of each landscape predictor) were standardized so that each had a mean of zero and a standard deviation of one (Zuur et al. Reference Zuur, Ieno, Wlaker, Saveliev and Smith2009).
After selecting the best measure for each landscape predictor, we compared a set of candidate GLMMs (considering the hydrological catchments as a random factor) for each of the two dependent binary variables. Each set contained six models: an intercept-only model for reference (no fixed factors); three simple models with each of the three landscape predictors on their own; and models combining one of the two forest cover predictors (smaller and wider spatial scales) with the remoteness predictor. We tested for collinearity between the landscape predictors included in the same model using Pearson's correlation test and variance inflation factor (VIF). Although some predictors were correlated (Pearson's correlation >0.6), VIFs were below 3 in all cases (Table S3), indicating that collinearity was not a concern (Zuur et al. Reference Zuur, Ieno, Wlaker, Saveliev and Smith2009). Alternative models in each set were compared using the difference in their AICc values in relation to the first-ranked model (∆AICc) (Burnham & Anderson Reference Burnham and Anderson2002). A value of ∆AICc ≤2 indicates equally plausible models. All analyses were implemented in R 3.0.3 (R Core Team 2014) using the lme4 package (Bates et al. Reference Bates, Maechler and Bolker2011).
RESULTS
Bushmeat consumption was far more common than hunting. In 80.5% of sampled households (range = 50–100%, mean = 79.7 and SD = 14.4 across hydrological catchments), members reported having consumed bushmeat at least once in the previous 12 months (Fig. 2, Table S4). Amongst bushmeat-consuming households, 59% stated that the last bushmeat they ate was a gift, 35% stated that it was hunted by one of the members of the household and 6% stated that it was purchased. In 85% of households that consumed bushmeat in the last year, interviewees reported having last consumed either paca (Cuniculus paca) or an armadillo species (Fig. S1(a)). The presence of a hunter in the household was reported in only 40.8% of the sampled households, and varied between 0 and 100% (mean = 43, SD = 25.8) across hydrological catchments (Fig. 2), with the number of hunting events per month also being low (median = 2, mean = 3.5, SD = 3.3). The most frequently hunted species reported by hunters were either paca (C. paca) or armadillos (Fig. S1(b)).
Landscape correlates of bushmeat consumption and hunting
For bushmeat consumption, the best measure for both forest cover within the property and forest cover within the wider landscape included all forest types together. A 10-km radius buffer (without accounting for the number of households) was the best buffer size for computing forest cover within the wider landscape (Table S2). The best measure for remoteness was the distance to the largest urban centre (Table S2). These predictors were then included in the model selection procedure for analysing the landscape correlates of bushmeat consumption. Three models were selected (Table 1). The model including only forest cover within the wider landscape was the first-ranked model, followed by the model including only remoteness and then the model including both of these variables. The results did not change when considering if bushmeat was consumed at least once in the previous 30 days (rather than in the previous 12 months; Table S1).
Table 1 Model selection results for bushmeat consumption in the last 12 months and hunting as a function of landscape predictors. Selected models (∆AICc <2) are in bold. K = Number of parameters; logLik = Log-likelihood of the model; AICc = Akaike information criterion modified for small samples; ∆AICc = Difference in AICc value compared to the first-ranked model; ωi = Akaike weight; Coefficient 1 = Coefficient for the first variable in the model; Coefficient 2 = Coefficient for the second variable in the model when the model has two variables. In parenthesis = Standard errors for the coefficients.

As expected, higher total forest cover within the wider landscape was positively associated with bushmeat consumption probability (Table 1). In heavily deforested areas (first quintile; ≤27% forest cover in surrounding landscape), 70% of households consumed bushmeat compared to 92% of households within more forested landscapes (fifth quintile; 71–94% forest cover) (Fig. 3(a)). Living in remote landscapes was also associated with increased probability of bushmeat consumption (Table 1). Within 29.5 km of the largest urban centres (first quintile of the data), 70% of households had consumed bushmeat, whereas for the most remote households (between 55 and 117.5 km; fifth quintile), 90% had consumed bushmeat (Fig. 3(b)). Both variables are likely to be important for explaining the likelihood of having consumed bushmeat, as the model that included both was also selected (Table 1).

Figure 3 Relationships between landscape characteristics and bushmeat consumption and hunting. Probability of bushmeat consumption as a function of (a) forest cover within a 5-km radius buffer and (b) remoteness as predicted by the first- and second-ranked models in Table 1 (black line). (c) Probability of hunting as a function of forest cover within a 5-km radius buffer as predicted by the first-ranked model in Table 1 (black line). Bars represent the proportions of households where bushmeat was consumed in the last 12 months or that harbour at least one hunter.
For bushmeat hunting, the best measure for forest cover within the property was the total primary forest cover (degraded plus non-degraded), while total forest cover (total primary and secondary forest) in a 5-km radius buffer, without accounting for the number of households, was the best measure for forest cover within the wider landscape (Table S2). The best measure for remoteness was again the distance to the largest urban centre (Table S2). These predictors were then included in the model selection procedure for analysing the landscape correlates of hunting. Two models were selected (Table 1). The model including only forest cover within the wider landscape was the first-ranked model, followed by the model including both forest cover within the wider landscape and remoteness.
As expected, higher forest cover within the wider landscape was positively associated with hunting (Table 1). Only 23% of the households in heavily deforested areas (first quintile; ≤23.5% forest cover in the surrounding landscape) had at least one hunter, compared to 66% of household within more forested landscapes (fifth quintile; 71.5–92% forest cover) (Fig. 3(c)). In contrast to our observations for bushmeat consumption, when accounting for forest cover within the wider landscape, households near urban centres were more likely to have a hunter (Table 1). However, the negative effect of remoteness on hunting probability may not be strong because the coefficient standard error encompasses zero (Table 1).
DISCUSSION
By investigating a large set of households distributed across a wide and heterogeneous region, we demonstrate that landscape context is important for determining both bushmeat consumption and hunting. Forest cover and remoteness explained the variation in bushmeat consumption probability equally well and bushmeat consumption increased with forest cover and remoteness as predicted. By contrast, forest cover was the best single predictor of hunting probability, presenting the predicted positive effect, while remoteness explained hunting probability only when accounting for forest cover, presenting an unexpected negative effect. In the following paragraphs, we discuss in detail the effects of these landscape characteristics and their implications for wildlife conservation and rural livelihoods.
Our results support the idea that bushmeat consumption remains widespread in post-frontier tropical regions, even in deforested, densely populated landscapes near urban centres. Thus, our results support assertions that bushmeat can continue to play an important role in rural livelihoods in altered areas (Schulte-Herbrüggen et al. Reference Schulte-Herbrüggen, Cowlishaw, Homewood and Rowcliffe2013). Moreover, although consumption was more likely in remote, forested landscapes, total demand for bushmeat is likely to be higher in less remote, deforested landscapes, given the higher human population densities in these areas. Hunting was less widespread than consumption and may be under-reported. Gifts and trade, which were often declared by interviewees in our study, provide important means of acquiring bushmeat other than hunting, so that many killed animals are eventually consumed in households other than those of the hunters (de Merode et al. Reference de Merode, Homewood and Cowlishaw2004).
Bushmeat consumption was more likely in more remote and forested areas, as expected, with forest cover within the wider landscape and remoteness being equally important drivers. Our finding is consistent with the fact that people living in remote, forested areas have less access to domestic marketed meat (higher demand for bushmeat; Parry et al. Reference Parry, Day, Amaral and Peres2010a), but are exposed to a higher availability of game (higher supply of bushmeat; Sampaio et al. Reference Sampaio, Lima, Magnusson and Peres2010; Moraes Reference Moraes2016), making bushmeat more important for household food security in those areas. Indeed, studies in Africa have shown that poor access to other types of meat (because of their higher price compared to bushmeat) can drive bushmeat consumption, which is also more common in places close to game sources (Brashares et al. Reference Brashares, Golden, Weinbaum, Barret and Okello2011; Foerster et al. Reference Foerster, Wilkie, Morelli, Demmer, Starkey, Telfer, Steil and Lewbel2012; Mgawe et al. Reference Mgawe, Mulder, Caro, Martin and Kiffner2012). Remoteness has also been linked to greater dependence on hunting for subsistence (Pangau-Adams et al. Reference Pangau-Adams, Noske and Muehlenberg2012). Importantly, bushmeat consumption may be more variable and even more strongly related to landscape context, especially remoteness, within still intact pre-frontier regions, where it can take many days for a riverine household to reach an urban centre (Parry et al. Reference Parry, Day, Amaral and Peres2010a) and when quantified more precisely. Future studies should thus focus on a finer quantification of bushmeat consumption, as well as encompass and compare regions with distinct deforestation dynamics.
Forest cover within the wider landscape was the main driver of hunting, with remoteness being important only when also accounting for the variability in forest cover. This is again consistent with the idea that forested areas support larger populations of game species (Chiarello Reference Chiarello1999; Peres Reference Peres2001; Michalski & Peres Reference Michalski and Peres2007; Sampaio et al. Reference Sampaio, Lima, Magnusson and Peres2010; Prist et al. Reference Prist, Michalski and Metzger2012; Ochoa-Quintero et al. Reference Ochoa-Quintero, Gardner, Rosa, Ferraz and Sutherland2015) and thus the supply of game to hunters is higher in those areas. Indeed, the proximity of game sources (such as protected areas) has been associated with higher hunting rates (Brashares et al. Reference Brashares, Golden, Weinbaum, Barret and Okello2011; Nuno et al. Reference Nuno, Bunnefeld, Naiman and Milner-Gulland2013). However, we did not find that remote households were more likely to have a hunter. In contrast, we found some evidence that the opposite might be happening.
The literature suggests that the higher human population density – and thus higher total demand for bushmeat – near urban centres may support a stronger informal bushmeat trade, stimulating hunting. Bushmeat trade is frequently reported as supporting higher profits near urban centres, especially in Africa, where trade is well developed (Brashares et al. Reference Brashares, Golden, Weinbaum, Barret and Okello2011). Similarly, people living closer to major markets are often reported as engaging more in commercial bushmeat trade (Espinoza Reference Espinosa2009; Pangau-Adams et al. Reference Pangau-Adams, Noske and Muehlenberg2012) and higher purchasing was associated with greater monetary power by urban consumers (Fa et al. Reference Fa, Albrechtsen, Johnson and Macdonald2009). While there are few studies on bushmeat trade in the Brazilian Amazon (Nasi et al. Reference Nasi, Taber and van Vliet2011), it occurs even in open markets (Chaves Baía Júnior et al. Reference Chaves Baía Júnior, Guimarães and Le Pendu2010) and may involve large volumes in urban settings, with hundreds of tonnes of bushmeat traded per year in three Amazonian cities alone (van Vliet et al. Reference van Vliet, Quiceno-Mesa, Cruz-Antia, Aquino, Moreno and Nasi2014), indicating its importance. Although our dataset is not adequate for testing hypotheses on bushmeat trade and commercial hunting, our results suggest that hunting probability does not decline close to urban centres as we first hypothesized. Future studies in the Amazon should further investigate the effects of distance to urban centres as a means of clarifying the role of bushmeat trade in hunting probability and frequency.
By considering different measures of forest cover and remoteness, our study also indicates the best landscape predictors of bushmeat consumption and hunting decisions. Forest cover per household was not associated with either bushmeat consumption or hunting, indicating that human population density may not always be important in determining local hunting pressure (sensu Urquiza-Haas et al. Reference Urquiza-Haas, Peres and Dolman2009) or that forest cover alone is, at least in our study region, a better predictor of game availability. Given past deforestation, leading to the high variation in forest cover across the region, forest loss and fragmentation should indeed be the major determinant of game availability, as found in other highly disturbed Amazon regions (Michalski & Peres Reference Michalski and Peres2007). In more pristine regions, though, human population density may be more important (Parry & Peres Reference Parry and Peres2015). Importantly, our results highlight that forest cover should be accessed within the wider landscape in accordance with the idea that the persistence and local abundance of game species depend on ecological processes that occur at larger spatial scales (Andrén Reference Andrén1994). In addition, in post-frontier regions such as our study area, degraded and secondary forests are prevalent and probably important determinants of game availability. Habitat disturbance was also shown to have a positive effect on bushmeat supply in Africa (McNamara et al. Reference McNamara, Kusimi, Rowcliffe, Cowlishaw, Brenyah and Milner-Gulland2015). Within relatively ‘pristine’ pre-frontier regions, however, wildlife abundance within non-degraded primary forest is probably a more important determinant of bushmeat consumption and hunting (Parry et al. Reference Parry, Barlow and Peres2009b). We also found that distance to the largest urban centre was the best predictor of remoteness in the case of both bushmeat consumption and hunting. Distance to the largest city may be more strongly related to limited access to alternative sources of protein, affecting bushmeat consumption because food is usually cheaper in these larger cities compared to smaller cities and rural populations may prefer buying food there. Moreover, large cities should present a stronger association with bushmeat trade, positively affecting hunting probability.
Implications for conservation and rural livelihoods
Strategies and policies to conserve biodiversity and maintain food security should take into account the spatial variation in hunting and bushmeat consumption associated with forest cover and remoteness. People from more remote, forested areas are more likely to consume bushmeat and thus to be the most dependent on bushmeat for subsistence and the most vulnerable to law enforcement policies on hunting. Nevertheless, people living near urban centres might be the actors contributing the most to bushmeat hunting because of the higher population density and the higher probability of having a hunter in the household.
Our study also suggests that bushmeat consumption is prevalent even in heavily disturbed regions and that hunting is unlikely to decrease with the migration of rural residents to urban or peri-urban areas (Parry et al. Reference Parry, Peres, Day and Amaral2010b; Wilkie et al. Reference Wilkie, Bennett, Peres and Cunningham2011). The increased in-migration to urban and peri-urban areas, which is occurring in many areas of Amazonia, may not result in reduced hunting of game species.
Finally, given the importance of landscape context to hunting and bushmeat consumption, future studies should focus on the interactions between these and socioeconomic and cultural factors, which are frequently considered to be the main drivers of the extraction and use of this important forest product, but are usually studied in isolation (e.g. Godoy et al. Reference Godoy, Undurraga, Wilkie, Reyes-García, Huanca, Leonard, NcDade, Tanner and Vadez2010; Rentsch & Damon Reference Rentsch and Damon2013).
ACKNOWLEDGEMENTS
We thank K.S. Gonçalves, A.E.M. Ferreira, C.D.F. Costa, A. Alves and A. Gomes for helping with data collection; S. Nunes and J.V. Siqueira for mapping forest cover; and all the collaborating landowners and workers unions of Santarém and Belterra for their support. This is publication number 57 from the Sustainable Amazon Network series (www.redeamazoniasustentavel.org).
FINANCIAL SUPPORT
This work was supported by INCT – Biodiversidade e Uso da Terra na Amazônia (CNPq 574008/2008-0), Embrapa (SEG: 02.08.06.005.00), the UK government Darwin Initiative (17-023), the Nature Conservancy, NERC (NE/F01614X/1 and NE/G000816/1) and FAPESP (2011/19108-0). P.C.T. had doctoral scholarships from CAPES and FAPESP (2011/19606-0) and R.P. had a research fellowship from CNPq (308205/2014-6).
CONFLICT OF INTEREST
None.
ETHICAL STANDARDS
None.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0376892917000510






