

Some 50 years ago, Alwyn Williams published a benchmark paper on the Barr and Lower Ardmillan stratigraphy and brachiopod faunas. Williams' (Reference Williams1962) paper is seminal in that: (a) it provided detailed, complete monographic and numerical descriptions of the middle to upper Ordovician brachiopod faunas of the Girvan area, which had not been updated since the works by Davidson (Reference Davidson1866–1871, Reference Davidson1883) and Reed (Reference Reed1917), while promoting cutting-edge statistical techniques; (b) it proposed a revised stratigraphical framework for the district and correlation with Ordovician successions of the North American south Appalachians; (c) it compared the brachiopod faunas with those south Appalachian faunas (Cooper Reference Cooper1956), emphasising the strong biogeographic similarities of these faunas from localities now on opposite sides of an ocean, at a time when modern plate tectonic theory was in its infancy and just beginning to gain acceptance by the scientific community; and (d) it presented a model in which fault-controlled sedimentation explained the juxtaposition of deep and shallow-water facies, thus explaining the close proximity of graptolite and shelly biofacies (see Lapworth Reference Lapworth1882). However, since this classic paper, there has been relatively little work on the Barr and Lower Ardmillan brachiopods of the Girvan area, Scotland, whereas the Upper Ardmillan has been more thoroughly investigated (see for example Harper Reference Harper1984–2006, Reference Harper2001). Recently, Harper & Stewart (Reference Harper and Stewart2008) have synthesised faunal changes during the Barr throughout the Ardmillan groups, tracking brachiopod biodiversity through the Ordovician succession at Girvan. Following recent advances in the understanding of the stratigraphy of the cover succession in the Girvan area (Ingham Reference Ingham, Fortey, Harper, Ingham, Owen, Parkes, Rushton and Woodcock2000) and in the taxonomy of the Brachiopoda (Kaesler Reference Kaesler2000–2007), Williams' (Reference Williams1962) data are in need of revision. We provide here an updated taxonomy of the Girvan brachiopod faunas (see Appendix 1, Supplementary Material (SM)) from the Barr Group and Ardwell Subgroup. This fundamental revision, used in conjunction with data from coeval North American southern Appalachian brachiopod faunas in multivariate statistical analyses, provides a more modern and updated interpretation of depositional environments throughout the middle to late Ordovician in the Girvan area and its palaeogeographical setting.
1. Stratigraphy and brachiopod biofacies
1.1. Barr Group
This is the lower of Lapworth's (Reference Lapworth1882) two great divisions of the Ordovician cover succession above the Ballantrae Complex. This succession is most complete in the Penwhapple Burn–Stinchar Valley areas and comprises, in ascending stratigraphical order (Fig. 1), the fossiliferous horizons of the Confinis, Stinchar Limestone and Superstes formations deposited from the late Darriwilian to the early Sandbian. The Auchensoul Bridge Member is the lateral equivalent of the Confinis Formation. Deposition of these two units records the gradual abandonment of the southward prograding Kirkland fan-delta (Ince Reference Ince1984) across a confined, channelised area of the shelf into the deep shelf. Moreover, the Confinis Formation is interpreted to have developed in a position distal to the presumed nearshore transitional sandstone of the Minuntion Member of the Confinis Formation.
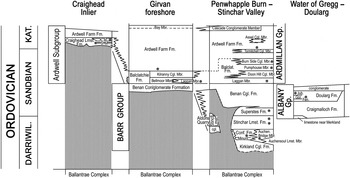
Figure 1 Stratigraphy of the Barr and Lower Ardmillan groups in the Girvan district, SW Scotland. Adapted from Ingham (Reference Ingham, Fortey, Harper, Ingham, Owen, Parkes, Rushton and Woodcock2000) and Mohibullah et al. (Reference Mohibullah, Williams, Vandenbroucke, Sabbe and Zalasiewicz2012). Abbreviations: DARRIWIL.=Darriwilian; KAT.=Katian; Balclat.=Balclatchie; Conf.=Confinis; Fen.=Fence; Gor.=Gorse; Jub.=Jubilation; K. M.=Kiln Mudstone Member; Minunt.=Minuntion; S. M.=Sericoidea Mudstone Member; Stinch.=Stinchar; Cgl.=Conglomerate; Lmst.=Limestone; Fm.=Formation; Gp.=Group; Mbr.=Member. Fossiliferous shelly horizons indicated by ❄.
These units are characterised by diverse brachiopod faunas (see Appendix 2 (SM)) in which some taxa, for example species of Leptellina, Hesperorthis, Sulevorthis or Valcourea, are recurrent throughout the Barr Group. The Confinis Formation and the Auchensoul Bridge Member share almost half of their faunal content (see Appendix 2 (SM), [CONF] and [AUCHN]), notably Sulevorthis parvicrassicostatus (Cooper), Valcourea confinis (Salter), Leptellina (L.) semilunata Williams and Hesperorthis australis exitis Williams. The Minuntion Member of the Confinis Formation (‘Transitional Sandstone’ of Ince Reference Ince1984), has a low brachiopod diversity and the assemblage is dominated by Valcourea confinis (Salter), representing 50% of the total assemblage ([MIN]). More than 65% of the taxa recorded in the Minuntion Member are present in the Confinis Formation (namely Colaptomena macallumi (Reed), Multicostella aff. plena Cooper, Triplesia craigensis (Reed) and Valcourea confinis (Reed)). All these are typical of shallow-water environments.
The Stinchar Limestone Formation is interpreted as a fan-delta abandonment phase, and is characterised by fine-grained calcareous horizons. The brachiopod content of the lower part of the formation ([lowST]) is less diverse, but over 80% are also recorded from the underlying Confinis Formation and over 40% in the Auchensoul Bridge Member. The upper part of the Stinchar Limestone is more diverse ([upST]), but about 15% of the fauna is present in the lower part of the Stinchar Formation. The faunal content of the top of the Stinchar Limestone Formation is similar to those of the underlying Auchensoul Bridge Member and Confinis Formation, with Bimuria, Christiania, Leptellina, Sulevorthis and Protozyga in common; however, the presence of taxa such as Ptychoglyptus, Cyrtonotella and Taphrorthis only in the top part of the Stinchar Limestone Formation, indicates a deepening, as shown by the associated trilobite fauna also (Tripp Reference Tripp1993).
Deposition of the Stinchar Formation ceased abruptly and the limestone unit passed into the finely graded alternation of siltstones and mudstones of the Superstes Formation (Williams Reference Williams1962; Ince Reference Ince1984). The sequence gradually disappears (eastwards), only to re-appear about 300 m north-north-east of Minuntion as dark-grey nodular mudstones (Williams Reference Williams1962). Williams (Reference Williams1962) considered the entire Superstes Formation to have been excised in two broad channels at the base of the Benan Conglomerate Formation (Ince Reference Ince1984), “not more than one mile wide” (Williams Reference Williams1962, p. 9). The mudstones are sparsely fossiliferous but contain a trilobite assemblage of the Nileid community (Tripp Reference Tripp1976, Reference Tripp1993; Ince Reference Ince1984; Ingham & Tripp Reference Ingham and Tripp1991) indicative of outer-shelf water depths. Ince (Reference Ince1984) showed evidence that deposition of the Superstes Formation (and the lowermost horizons of the Benan Conglomerate) took place within a submarine fan complex off-shore from a fan delta, the Superstes Formation being deposited in interlobe areas. Tripp et al. (Reference Tripp, Williams and Paul1981, p. 21) documented an exposure of the Superstes Formation near Colmonell (Fig. 2), along the west bank of an unnamed tributary of the Stinchar River, 10 km south-south-west of Brockloch, where Williams (Reference Williams1962) collected his fauna. Tripp et al. (Reference Tripp, Williams and Paul1981) collected a less diverse assemblage, but 80% of their fauna is in common with Williams' (Reference Williams1962) assemblage ([SUP62]). Tripp et al. (Reference Tripp, Williams and Paul1981) collected two additional genera of ‘non articulate’ brachiopods ([SUP81]), Conotreta and Eodinobolus (listed originally as Obolellina). This assemblage is also very similar to that from the top of the Stinchar Limestone Formation, with 75% of the brachiopod taxa in common. Williams' (Reference Williams1962) assemblage comprises almost 50% of the taxa described in the underlying top part of the Stinchar Limestone Formation.

Figure 2 Geographical distribution of the assemblages mentioned in the study. 1=[AUCHN]; 2=[CONF]; 3=[CRAIGM]; 4=[GORSE] and [JUB]; 5=[MIN]; 6=[lowST], [upST] and [PINMY]; 7=[SUP62] and [MEIKL]; 8=[SUP81]; 9=[BELL] and [SHIEL]; 10=[GLEND]; 11=[PINM1]; 12=[LAGG]; 13=[DALF]; 14=[DOWH]; 15=[PUMPH], [BURNS], [ASSL], [CASC] and [TORM1]; 16=[ARDB]; 17=[BARBH]; 18=[PINM2]; 19=[REAG]; 20=[TORM2]; 21=[CRAIGH], [KILN], [SERIC] and [CRAIGLIM].
1.2. Albany Group
The Albany Group (Fig. 1) is the off-shelf equivalent to the Barr Group (Rushton et al. Reference Rushton, Tunnicliff and Tripp1996). The lower part is characterised by the Craigmalloch Formation, consisting of a thick greywacke, conglomerate and siltstone unit, south of the Stinchar Valley Fault (Ince Reference Ince1984; Ingham Reference Ingham, Fortey, Harper, Ingham, Owen, Parkes, Rushton and Woodcock2000), in the vicinity of Barr (Fig. 2). Rushton et al. (Reference Rushton, Tunnicliff and Tripp1996) described a graptolite assemblage, while Ince (Reference Ince1984) listed two trilobite taxa characteristic of deep basinal settings, and two brachiopod taxa, ?Eodinobolus sp. and Sericoidea sp. ([CRAIGM]), also typical of deep-water environments (equivalent to or deeper than Boucot's (Reference Boucot1975) Benthic Assemblage 5).
The upper part of the group largely comprises the Doularg Formation, which was defined by Ingham & Tripp (Reference Ingham and Tripp1991). It consists of the unfossiliferous Fence Member at the base, succeeded by the Gorse Member, the Separation Sandstone Bed and the Jubilation Member. The Separation Sandstone Bed, however, was not formally recognised in the revision of the stratigraphy of the formation by Ingham (Reference Ingham, Fortey, Harper, Ingham, Owen, Parkes, Rushton and Woodcock2000). Both the Gorse and Jubilation members have yielded interesting assemblages of trilobites and brachiopods ([GORSE] and [JUB]). Williams (Reference Williams1962, p. 47) listed and described a diverse fauna of brachiopods from the Gorse Member ([GORSE]), his “Albany mudstones with nodular limestones”, characterised by a similar assemblage to the Superstes Formation at Brockloch (Fig. 2), with 80% of the taxa found in the underlying unit. About half of the assemblage is in common with the assemblage from the Superstes Formation collected at Colmonell by Tripp et al. (Reference Tripp, Williams and Paul1981). The Separation Sandstone has, to date, only yielded a single large specimen of Macrocoelia (Ingham & Tripp Reference Ingham and Tripp1991).
Ingham & Tripp (Reference Ingham and Tripp1991) described a diverse fauna of trilobites from the Jubilation Member, characteristic of the deep-water environment Nileid biofacies. Although not particularly abundant in the Jubilation Member (Ingham & Tripp Reference Ingham and Tripp1991, p. 29), the brachiopod assemblage ([JUB]) includes common Ptychoglyptus cf. virginiensis Willard and some post-Barr Group taxa. Most of the assemblage is constituted by linguliform brachiopods such as Conotreta cf. davidsoni (Reed), Lingulella cf. angustior (Reed), Lingulella? sp., Palaeoglossa? maccullochi (Reed) and Schizotreta medioradiata (Reed), as well as the small plectambonitoid Sericoidea sp. Ingham & Tripp (Reference Ingham and Tripp1991) listed Schizotreta mediostriata (Reed) from the Jubilation Member. However, this species, now reassigned to Lingulella, is characteristic of the younger Lady Burn and South Threave formations (uppermost Katian) of the Drummuck Subgroup in Girvan. Ingham & Tripp (Reference Ingham and Tripp1991) possibly misspelled the species name. The species medioradiata is typically found in the Ardwell Subgroup and therefore this is probably the species that should be listed in the Jubilation Member. The composition of the brachiopod assemblage of the Jubilation Member is very different from those of the other assemblages of the Barr Group, but very similar (although less diverse) to the brachiopod assemblage of the Laggan Member of the Balclatchie Formation. Three-quarters of the genera in the Jubilation Member are found in the younger Laggan Member, and two-thirds are only found in post-Barr Group horizons.
1.3. Lower Ardmillan Group
The Ardmillan Group was introduced by Lapworth (Reference Lapworth1882) as one of his major lithological divisions. It originally included all Ordovician strata as then known above the Benan Conglomerate. The Lower Ardmillan Group (Fig. 1) is the Ardwell Subgroup, and south of Girvan it is composed of, in ascending stratigraphical order, the Balclatchie Formation (upper Sandbian), which rests conformably above the Benan Conglomerate Formation, and the Ardwell Farm Formation (upper Sandbian–lower Katian), which diachronously overlies the Balclatchie Formation. In the Craighead Inlier, north of Girvan, the Ardwell Subgroup consists of the Craighead Formation and the Ardwell Farm Formation. On the Girvan foreshore and adjacent areas, the Balclatchie Formation is divided into the Bellmoor Member and the Kilranny Conglomerate Member (Ingham Reference Ingham, Fortey, Harper, Ingham, Owen, Parkes, Rushton and Woodcock2000), both substantial units but locally developed. In the Penwhapple Burn–Stinchar Valley belt, the Balclatchie Formation consists of the Laggan, the Doon Hill Conglomerate, the Pumphouse and the Burn Side Conglomerate members. The Ardwell Farm Formation includes locally the Tormitchell Conglomerate Member, the Assell Conglomerate Member and the Cascade Conglomerate Member. In the Craighead Inlier, the Craighead Formation comprises a massive limestone facies and includes the Kiln Mudstone Member towards the top and the Sericoidea Mudstone Member lower down (Ingham Reference Ingham, Fortey, Harper, Ingham, Owen, Parkes, Rushton and Woodcock2000). The Ardwell Farm Formation includes the Plantinhead Member.
The Laggan Member brachiopod assemblage from Williams' (Reference Williams1962) Laggan Burn locality ([LAGG]) is relatively diverse (see Appendix 3 (SM)) and consists of a mixture of small ‘non-articulates’, and small plectambonitoids (such as Anisopleurella and Sericoidea), together with orthides (such as Glyptorthis, Skenidioides and Dalmanella). Other assemblages were collected by Williams (Reference Williams1962) from close-by localities (Fig. 2), at Dalfask Hill ([DALF]), Pinmacher Farm ([PINM1]), Glendrissaig Farm ([GLEND]) and Woodland Farm, Dow Hill ([DOWH]). These assemblages become less diverse northwestward, but they are constituted by the same taxa, with variants, including the presence of additional ‘non-articulates’ (Dalfask Hill) and Paucicrura (Pinmacher Farm) (see Appendix 3 (SM)). Harper & Owen (Reference Harper and Owen1986) described a shelly fauna from the Laggan Member, about 100m west of Williams' (Reference Williams1962) Laggan Burn locality. It consisted of the brachiopod Onniella and the trilobite Diacanthaspis. It represents a rare occurrence of an indigenous deep-water biofacies within the outer shelf or slope environment of the early Late Ordovician of the Girvan area. Moreover, Stewart & Owen (Reference Stewart and Owen2008) documented the presence of a small strophomenoid, possibly Foliomena. Williams (Reference Williams1962, p. 31) also described two assemblages from the lateral equivalent of the Laggan Member, the Bellmoor Member (formerly known as ‘Infra-Kilranny greywackes’) of the Balclatchie Formation, in the vicinity of Kennedy's Pass [BELL] and east of Shiel Hill [SHIEL]. They are not very diverse and only contain Sulevorthis in common. The former is composed of Dolerorthis, Eoplectodonta, Leptellina, Lingulella, Sulevorthis and Rostricellula; whilst the latter is characterised by Bilobia, Bimuria, Dactylogonia, Leptaena, Plectorthis, Sulevorthis and Titanambonites. There are some taxonomic similarities between these and the assemblages collected from the Laggan Member, whilst the Bellmoor Member assemblages contain taxa such as Dolerorthis and Leptaena, better documented in post-Balclatchie horizons. The younger, fossiliferous Pumphouse and Burn Side Conglomerate members are developed in the Penwhapple Burn–Stinchar Valley area and brachiopod assemblages were described by Williams (Reference Williams1962), notably from around Penwhapple Burn (Fig. 2). The assemblages ([PUMPH] and [BURNS]) are diverse and the Pumphouse Member comprises about a quarter of the taxa described from the older Laggan and Bellmoor members, whereas the Burn Side Conglomerate Member has yielded a totally different assemblage, with only one genus in common with the Bellmoor Member. The Pumphouse Member assemblage ([PUMPH]) is characterised by a quarter of ‘non-articulates’ (Glossella, Lingulella, Apatobolus, Paterula, Multispinula and Schizotreta) and a quarter of Plectambonitoidea (including genera such as Anisopleurella, Eoplectodonta and Isophragma). The assemblage from the Burn Side Conglomerate Member is very different from any other assemblage and is dominated by rhynchonellate taxa (orthides and rhynchonellides). Only three genera (Christiania, Oxoplecia and Taphrorthis) are found in common with the Pumphouse Member. Nevertheless, the assemblage from the Burn Side Conglomerate Member is composed, at a higher taxonomic level, of similar fauna characterised by some ‘non-articulates’ (such as Eodinobolus) and plectambonitoids (including Leangella and Leptellina).
Williams (Reference Williams1962) described several assemblages (see Appendix 3 (SM)), of varied diversity, from the Ardwell Farm Formation. The most diverse was collected from Ardmillan Braes ([ARDB]), about 650 m south of Ardmillan House and Pinmore ([PINM2]), about 6 km south of Girvan (Fig. 2). Both assemblages are characterised by similar taxa (small plectambonitoids, strophomenoids and orthoids), but the Ardmillan Braes' assemblage has yielded a range of ‘non-articulates’ (Elliptoglossa, Glossella, Glyptoglossella, Lingulella and Paterula) not present in Pinmore. Conotreta is present in both, whereas Petrocrania is present in Pinmore solely. A fossiliferous mudstone above the Tormitchell Conglomerate Member, at Barbae Hill, has yielded a low diversity assemblage ([BARBH]), but 80% of its taxa have not been collected from the other Ardwell Farm Formation localities. It comprises the small dalmanelloid Paucicrura, the plectambonitoid Sericoidea and the skenidioid Skenidioides (see Appendix 3 (SM)).
In the Penwhapple Burn–Stinchar Valley area, the section includes a number of conglomeratic members, from which Williams (Reference Williams1962, pp 35–36) has described low diversity assemblages. In ascending stratigraphical order, they are the Tormitchell Conglomerate, the Assel Conglomerate and the Cascade Conglomerate members. The assemblage of the Tormitchell Conglomerate ([TORM2]) was collected from the mudstones and siltstones intercalated within the pebbly sandstone component. The assemblages from the other conglomerates ([ASSL] and [CASC]) were collected from the bedded grits. There is a gradient from the deeper-water taxa of the Tormitchell Conglomerate, such as Paucicrura, Leptellina and Bimuria, to a mix of taxa representing shallower, quiet-water environments (Assel Conglomerate), to shallow, subtidal ramp environments (Cascade Conglomerate), characterised by Hesperorthis and Platystrophia.
In the Craighead Inlier, the Lower Ardmillan Group succession is coeval with the upper Ardwell Farm Formation further south. The Craighead Limestone Formation includes the Kiln Mudstone Member towards the top and the Sericoidea Mudstone Member lower down (Fig. 1). Although mentioned by Williams (Reference Williams1962, p. 53), the faunal assemblage of the ‘Sericoidea Mudstones’ was not properly differentiated from the assemblage of the Craighead Limestone Formation as a whole and was listed with ‘the Limestones and Mudstones’. Nevertheless, Williams (Reference Williams1962) noted two genera recovered from ‘mudstones overlying the Limestones and Mudstones’, namely Cyclospira perplexa (Williams) and Sericoidea aff. restricta (Hadding), which both, without a doubt, belong to the Sericoidea Mudstone Member. The brachiopod assemblage from the Craighead Limestone Formation [CRAIGH] (sensu Williams Reference Williams1962, p. 258) is characterised by a high diversity (over 50 genera), 15% of ‘non-articulates’ and a combination of key taxa such as Sericoidea, Anisopleurella and Paucicrura, which indicate deep-water depositional environments of deep ramp to slope and basin. However, many shallow-water taxa were also listed by Williams (Reference Williams1962), which indicates that the fauna from the limestone beds and fauna from the mudstone lenses (Sericoidea Mudstone Member) within the limestone had not been discriminated. The Kiln Mudstone Member was introduced by Tripp (Reference Tripp1954). Its brachiopod fauna is not very diverse and contains a majority of rhynchonellate brachiopods ([KILN]), with rarer ‘non-articulates’ and plectambonitoids.
2. Analysis of brachiopod biofacies and environments
2.1. Barr Group
The data, compiled from various sources (Williams Reference Williams1962; Tripp et al. Reference Tripp, Williams and Paul1981; Ince Reference Ince1984; Ingham & Tripp Reference Ingham and Tripp1991), were integrated into a larger database (Appendix 2 (SM)) that includes brachiopod faunas from Potter & Boucot (Reference Potter, Boucot, Webby and Laurie1992, table 1a), from the late Darriwilian to early Sandbian of central and southern Appalachians, coeval with those of the Girvan district. The assemblage of the Blackford Formation in Virginia (Potter & Boucot Reference Potter, Boucot, Webby and Laurie1992, p. 312, table 1a) contains only one genus (Camarella); therefore these data were removed from the study. The database was investigated using the PAST software (Hammer et al. Reference Hammer, Harper and Ryan2001). The data were weighted with the value of the BA each genus is associated with, using Potter & Boucot (Reference Potter, Boucot, Webby and Laurie1992) and Patzkowsky (Reference Patzkowsky1995). Detrended correspondence analysis (DCA) was performed in order to identify any underlying gradients in the data. Axis 1 (Fig. 3) corresponds to an onshore to offshore gradient with increasing values of axis 1. The eastern North American assemblages are plotted (Fig. 3) according to brachiopod biofacies as established by Patzkowsky (Reference Patzkowsky1995). These biofacies are defined along a depth-related gradient from intertidal–shallow subtidal to offshore–deep ramp environments (see Patzkowsky (Reference Patzkowsky1995) for details), namely Rostricellula–Doleroides [Mosh, Tmbz, NMkt], Strophomena [Lnrl, Mrat, Witn, Wrdl, Elwy, WslC], Sowerbyella [Bnbt, Lnsr, WrdC] and Paucicrura–plectambonitacean [PrtF, Athn, LnrIV, LlOk, Btet, CmHl, LtzM, Efna, RchV, LbyH] biofacies. The biofacies are generally well constrained along the depth-related axis 1 and show little overlap. There is greater spread along axis 2, which accounts for the variations of the taxonomic content within each biofacies; in particular, within the Strophomena and Paucicrura–plectambonitacean biofacies. The Strophomena Biofacies is composed of a mix of assemblages typical of BAs 2–3 [Witn, Wrdl, Elwy] and 3 [Lnrl, Mrat, WslC]; variations that account for the vertical spread of the biofacies. Within the Sowerbyella Biofacies, [WrdC] and [Bnbt] represent deeper variants of the biofacies, as the assemblage is also composed of Leptellina and Skenidioides, which are more typical of the deeper-water Paucicrura–plectambonitacean Biofacies. Lastly, [LbyH] may represent a deeper–water variant of the biofacies, as it includes the small plectambonitoid Sericoidea.

Figure 3 Formation scores on axis 1 and 2 of detrended correspondence analysis (DCA), based on presence–absence of genera for the late Darriwilian and early Sandbian of brachiopod faunas from SW Scotland and central and southern Appalachian Mountains. EV1=0.6998; EV2=0.3385; EV3=0.1901; EV4=0.1483. Data from Ince (Reference Ince1984), Ingham & Tripp (Reference Ingham and Tripp1991), Potter & Boucot (Reference Potter, Boucot, Webby and Laurie1992), Tripp et al. (Reference Tripp, Williams and Paul1981) and Williams (Reference Williams1962); see Appendix 2 (SM). Appalachian faunas in lower case; Girvan faunas in capital letters.
The assemblage from the Minuntion Member [MIN] plots at the same level as formations representing a shallow, quiet-water, subtidal shelf (Potter & Boucot Reference Potter, Boucot, Webby and Laurie1992) belonging to BA 3 (Fig. 3) and typical of the Strophomena Biofacies. It is characterised by brachiopod genera such as Colaptomena, Multicostella, Valcourea and Multispinula. The assemblage from the Confinis Formation [CONF] is more diverse and the latter genera are also present here. Moreover, the Confinis Formation assemblage also yielded taxa typical of deeper BA 4, such as Leptellina, Isophragma and Gacella. This assemblage plots with the Appalachian formations consisting of a combination of sources typical of BA 3+4, and occupies an intermediate position between the Sowerbyella and Paucicrura–plectambonitacean biofacies. The assemblages from the Auchensoul Bridge Member [AUCHN] and the lower part of the Stinchar Formation [lowST] plot next to each other within the Paucicrura–plectambonitacean Biofacies, where Appalachian assemblages typical of BA 4 cluster. These are assemblages characterised by a sub-wave base deep ramp environment, in excess of 50 m deep. Although Harper & Stewart (Reference Harper and Stewart2008, p. 283) considered the “brachiopod faunas of the Auchensoul, Confinis and lower Stinchar unit” to be broadly similar, there is a marked difference in the taxonomic composition of [AUCHN] that accounts for the environmental interpretation of the fauna. Genera such as Bimuria, Christiania, Skenidioides and Xenambonites, which are typical of BA 4, are absent from the other two assemblages. The assemblage at the top of the Stinchar Limestone Formation [upST] shows a deepening in the depositional environment within the Paucicrura-plectambonitacean Biofacies, yielding taxa such as Bimuria, Christiania, Leptellina and Xenambonites, that are regarded as typical of BA 4, but also the arrival of new genera such as Craspedelia, Nothorthis, Phragmorthis and Taphrorthis, that occupy similar deep-water facies. The overlying Superstes Mudstone Formation has yielded two similar assemblages, [SUP62] and [SUP81], collected from two localities (Brockloch and Colmonell) both in the Girvan district. The assemblages are characterised by the presence of several taxa of ‘non-articulate’ brachiopods, which were poorly represented in the underlying formations, including Conotreta and Eodinobolus. These genera represent two new occurrences in the area. The ‘non-articulates’ are found in association with Glyptomena, Bimuria, Christiania and Isophragma, which are typical of deep-water facies. Both [SUP62] and [SUP81] plot closely together on Figure 3 and also closely to Appalachian assemblages typical of BA 4 to 4–5, indicating deep slope to ramp environments (Potter & Boucot Reference Potter, Boucot, Webby and Laurie1992). [SUP62] and [SUP81] are reminiscent of Patzkowsky's (Reference Patzkowsky1995) Paucicrura–plectambonitacean Biofacies, which is characterised by the presence of Paucicrura and numerous plectambonitoidean brachiopods, including Sowerbyella, Eoplectodonta and Bilobia, but also Christiania, Paurorthis and Skenidioides. This biofacies is typical of offshore environments below storm-wave base (Patzkowsky Reference Patzkowsky1995).
The Albany Group is the lateral equivalent of the upper Barr Group, including the Superstes Mudstone Formation and the unfossiliferous Benan Conglomerate Formation. The Craigmalloch Formation has yielded two brachiopod genera only, ?Eodinobolus and Sericoidea ([CRAIGM]). The assemblage plots on Figure 3, quite separate from any other assemblage, at the end range of the onshore–offshore gradient. This is due, on the one hand, to the taxonomic composition of the assemblage (both genera are typical of deeper-water environments, BA>5) and, on the other hand, to the low diversity of the assemblage. Ince (Reference Ince1984) listed a fauna from the fine-grained, hemipelagic tops of turbidites cropping out in the Water of Gregg near Changue House [NX 2892 9401], less than 2 km east of Barr. In addition to the brachiopods cited above, it comprised a diverse assemblage of graptolites and two trilobites, the small cyclopygid Microparia sp. and Homalopteon aff. portlocki Salter, both typical of deep-basinal settings. The overlying Doularg Formation has yielded two faunal assemblages, from the Gorse Member [GORSE] and the Jubilation Member [JUB], in ascending stratigraphical order. The assemblage from the Gorse Member [GORSE] is relatively diverse, with 22 species listed by Williams (Reference Williams1962, p. 47). This assemblage plots close to the assemblages typical of BA 4 to 4–5 from the central and southern Appalachian (Potter & Boucot Reference Potter, Boucot, Webby and Laurie1992) characterising the Paucicrura–plectambonitacean Biofacies. [GORSE] is characterised by rarer ‘non-articulates’ (sensu Patzkowsky Reference Patzkowsky1995) and by the absence of Sericoidea, a combination which defines deep-water slope to basin settings. [GORSE] possesses about 75% of its faunal content in common with the Superstes Mudstone Member and about 50% with the upper part of the Stinchar Limestone Formation. Such taxa include Bimuria, Christiania, Isophragma, Phragmorthis and Skenidioides, which are all typical of BAs at least 4–5 or deeper. The assemblage from the overlying Jubilation Member [JUB] plots clearly between a broad cluster comprising assemblages from the Gorse Member, the Paucicrura–plectambonitacean Biofacies, and the deeper-water assemblage of the Craigmalloch Formation. The assemblage from the Jubilation Member is less diverse than that of the Gorse Member, but more diverse than that of the Craigmalloch Formation. It comprises the small plectambonitoid brachiopod Sericoidea, together with some ‘non-articulate’ taxa such as Conotreta, Lingulella, Palaeoglossa and Schizotreta. These four taxa are typical of post-Barr assemblages in the Girvan area and, together with Sericoidea and Ptychoglyptus, are typical of deep ramp to slope and basin environments (Patzkowsky Reference Patzkowsky1995), below BA 5. Patzkowsky (Reference Patzkowsky1995) also showed that ‘non-articulate’ brachiopods biofacies mapped onto the slope and basin environmental framework (Patzkowsky Reference Patzkowsky1995, fig. 11), which can also explain the position of [CRAIGM] on Figure 3. This evidence indicates that [JUB] and [CRAIGM] occupy the deeper end of the gradient of the Paucicrura–plectambonitacean Biofacies, or a transitional biofacies between Patzkowsky's (Reference Patzkowsky1995) Paucicrura–plectambonitacean and inarticulate biofacies, the latter characterised solely by the presence of ‘non-articulates’.
2.2. Ardwell Subgroup
A second database (Appendix 3 (SM)) was compiled with assemblages from the Ardwell Subgroup (Williams Reference Williams1962), together with coeval faunas listed by Potter & Boucot (Reference Potter, Boucot, Webby and Laurie1992, table 2) from the Middle Ordovician (Sandbian) of North America (see also Potter & Boucot Reference Potter, Boucot, Webby and Laurie1992, fig. 1 for location map). Potter & Boucot's (Reference Potter, Boucot, Webby and Laurie1992) Locality 42 was excluded from the study because it yielded a single genus only. These faunas are interpreted as ranging between BAs 2–3 and 4–5. DCA was also used here, and the data were weighted using the same technique as above. Axis 1 (Fig. 4a) corresponds to an onshore to offshore gradient with decreasing values on axis 1. Potter & Boucot's (Reference Potter, Boucot, Webby and Laurie1992) faunas compare closely with Patzkowsky's (Reference Patzkowsky1995) brachiopod biofacies. Assemblages [Dcrh, Esba1, OpSt, Viol, Wtkr] are characteristic of the Rostricellula–Doleroides Biofacies, [Esba2, Hes, HsEo, HsSn] are typical of the Strophomena Biofacies, and [Trem, Ospl, Geis, Trip, Brmd] of the Sowerbyella Biofacies. There is great overlap between the clusters of the first two biofacies, which is a reflection of similar environmental conditions; i.e., shallow subtidal to lower transition zone. These two clusters show larger variations along axis 1 than axis 2, whereas the cluster typical of the third biofacies shows little variations along axis 1 but great variations along axis 2. This evidence shows that the Rostricellula–Doleroides and Strophomena biofacies are more taxonomically coherent than the Sowerbyella Biofacies. These are a mix of cratonic and extracratonic localities with undoubtedly some regional signature (localities are from the southern Appalachians and central North America). Therefore, these faunas show much greater taxonomic variation. Faunas interpreted as Paucicrura–plectambonitacean Biofacies [Encr, Lios, GR1, uOl, Cpnh, Mtbg, Artk] are scattered along axes 1 and 2, together with most of the Girvan assemblages.

Figure 4 Formation scores on axis 1 and 2 of detrended correspondence analysis (DCA), based on presence–absence of genera for the middle Sandbian–early Katian of brachiopod faunas from SW Scotland and North America: (a) EV1=0.6124; EV2=0.3497; EV3=0.2803; EV4=0.2087; (b) EV1=0.6083; EV2=0.3415; EV3=0.2756; EV4=0.1791. Data from Potter & Boucot (Reference Potter, Boucot, Webby and Laurie1992) and Williams (Reference Williams1962); see Appendix 3 (SM): (a) uses the database in Appendix 3.1; (b) uses the database in Appendix 3.1, in conjunction with supplementary material from the National Museums Scotland (Appendix 3.2), namely [SERIC] and [CRAIGLIM]. Appalachian faunas in lower case; Girvan faunas in capital letters.
The deep-water depositional environment trend demonstrated in the underlying horizons is a common feature of the Laggan Member of the Balclatchie Formation; these assemblages plot within the deeper end of the gradient. The assemblages [DOWH], [DALF], [LAGG], [PINM1], and [GLEND] plot with very little variation along axis 1, but greater variations along axis 2, which corresponds to inter-faunal taxonomic variations. The variation along axis 2 is due to the taxonomic composition of [DALF] the only assemblage where Apatobolus and Schizotreta were collected. Nevertheless, these assemblages are strongly coherent, yielding taxa such as Eoplectodonta semirugata (Reed), Sericoidea aff. abdita Williams and Bimuria youngiana recta Williams, typical of environments of BAs 4 to 4–5 or deeper, within the deeper end of the Paucicrura–plectambonitacean Biofacies, taxa that are present in three to all of the mentioned localities. The presence of ‘non-articulate’ brachiopods such as Conotreta and Lingulella, found in fine siltstone to mudstone beds, also supports the environmental interpretation.
The coeval assemblages from the Bellmoor Member are better developed around the present-day Girvan foreshore. Those assemblages ([BELL] and [SHIEL]) are also characterised by a range of brachiopods typical of BAs 4 or deeper, such as Bilobia, Sulevorthis, Eoplectodonta and ‘non-articulates’ such as Lingulella, indicating a position at the deeper fringe of the Paucicrura–plectambonitacean Biofacies.
There is relative shallowing of the depositional environment with the Pumphouse Member [PUMPH] and the Burn Side Conglomerate Member [BURNS], typified by the loss of many genera of ‘non-articulate’ brachiopods, but also by a drop in the relative content of taxa typical of BAs 4 or deeper, within the Paucicrura–plectambonitacean Biofacies. The increased presence of taxa typical of BA 3 and shallower, such as Mimella, Oligorhynchia, Platystrophia and Plectorthis in [BURNS] support this hypothesis. The assemblage [BURNS] plots with high values on axis 2, whereas [PUMPH] plots with the older Balclatchie Formation assemblages. Such a discrepancy indicates high inter-faunal taxonomic variation: [BURNS] only has three taxa in common with [PUMPH], whereas the latter had almost half of its taxonomic content in common with the older Ardwell assemblages in general.
The Ardwell Subgroup assemblages [REAG], [ARDB], [MEIKL], [PINM2], [BARBH], [TORM2] and [PINMY] (listed in a broad eastward direction) show substantial variation along axis 1, reflecting the variation in sediment (from greywackes to fine mudstones). Within these, the assemblages located south and southeast of Girvan, [MEIKL], [PINM2], [PINMY] and [TORM2], plot in a tight cluster which indicates similarities in faunal composition (Bilobia, Bimuria and Eoplectodonta for example). On the other hand [REAG], [ARDB] and [BARBH] are distinct from that cluster but also from each other. [BARBH] is composed of Paucicrura with other taxa, characteristic of BA 3 and also absent from [REAG], [ARDB] and that cluster. [REAG] and [ARDB] have also yielded plectambonitoid brachiopods, typical of deeper BAs 4 or 4–5, which complete the assortment. At the genus level, the Ardwell Subgroup assemblages are comparable with those described by Candela (Reference Candela, Brunton, Cocks and Long2001, Reference Candela2003) from the Bardahessiagh Formation (upper Sandbian to lower Katian) in Pomeroy, Northern Ireland. The Irish assemblages (named Q2 and M2) were also composed of taxa typical of BAs 4 or deeper (Bimuria, Isophragma and Salopina in Q2 and Diambonia, Bilobia, Anisopleurella and Sericoidea in M2). These were interpreted as local variants of the ‘Paucicrura–plectambonitacean’ biofacies. The gradient within the Scottish assemblages is that of a relative shallowing of the depositional environment southeastward.
The assemblages collected within the three conglomerate members of the Ardwell Farm Formation, [TORM1], [ASSL] and [CASC], listed in ascending stratigraphical order, are poorly diverse and show little taxonomic overlap, although [TORM1] and [ASSL] plot closer together. Figure 4a shows a shallowing trend with time from relatively deep-water [TORM1], defined by genera such as Oxoplecia, Dolerorthis and Laticrura (BA 4), to [ASSL] characterised by an input of taxa representative of BAs 2–3 to 3, both nevertheless included within the Paucicrura–plectambonitacean Biofacies, to [CASC] entirely composed of brachiopods typical of shallower environments (such as Hesperorthis and Platystrophia), which plots closer to the Sowerbyella Biofacies. The lithology coarsens and the beds thicken, indicating the shallowing of the depositional environment.
As noted above, Williams (Reference Williams1962) did not discriminate the fauna collected from the limestone beds of the Craighead Limestone Formation from the fauna specific to the mudstone lenses of the Sericoidea Mudstone Member, except in two instances when Sericoidea aff. restricta (Hadding) and Cyclospira perplexa (Williams) are listed from the “calcareous mudstones overlying the Limestones and Mudstones” (Williams Reference Williams1962, pp 188, 246). Therefore the combined assemblage [CRAIGH] loses some of its faunal and environmental signals. Nevertheless [CRAIGH] plots within the wide cluster of faunas characteristic of the Paucicrura–plectambonitacean Biofacies. As listed by Williams (Reference Williams1962, appendix X, p. 258) and as noted above, [CRAIGH] is highly diverse and is composed of an equal mix of shallow-water (BA 2–3) and deep-water (at least BA 4) taxa, such as Camerella, Rostricellula and Plectorthis on the one hand, and Sericoidea, Anisopleurella, Bilobia and Eoplectodonta on the other. [KILN], however, plots at the opposite end of axis 2, with little variation along axis 1. The variation along the taxonomic-sensitive axis is due to the low number of genera in common between [KILN] and [CRAIGH]: 16% of the taxa in [KILN] are present in [CRAIGH].
The National Museums Scotland holds a collection of brachiopods collected from the Craighead quarry, from both the Sericoidea Mudstone Member and the limestones of the Craighead Limestone Formation, information clearly indicated on the specimens' labels. The former has yielded Anoptambonites grayae (Davidson), Eoplectodonta conspicua (Reed), Bimuria youngiana (Davidson), Sericoidea aff. restricta (Hadding), Bilobia etheridgei (Davidson), Craigella grayiae (Davidson), Multispinula scotica (Davidson) and Petrocrania sp.; whereas the latter has yielded Anoptambonites grayae(?), Reuschella americana Cooper, Rostricellula lapworthi (Davidson), Palaeoglossa maccullochi (Reed), Dinorthis carrickensis Reed, Conotreta conoidea Reed, Longvillia lata (Williams), Idiospira thomsoni (Davidson), Anazyga orbis (Reed), Parastrophina balcletchiensis (Davidson), Pionodema girvaniensis (Davidson) and Hesperorthis craigensis (Reed). Inclusion of this material in the study shows that the newly differentiated [SERIC] (assemblage from the Sericoidea Mudstone Member) and [CRAIGLIM] (assemblage from the limestone beds of the Craighead Limestone Formation) plot in opposite positions within the broad Girvan cluster (Fig. 4b). [SERIC], which is composed of taxa typical of BAs 4–5 or deeper, plots close to other assemblages characteristic of offshore quiet environments, below the storm-wave base. [CRAIGLIM], on the other hand, is composed of taxa typical of BA 3 and plots closer to [BARBH] and the eastern North American faunas described by Potter & Boucot (Reference Potter, Boucot, Webby and Laurie1992), characteristic of a shallow offshore shelf environment and representing the Sowerbyella Biofacies; water-depth estimated by Potter & Boucot (Reference Potter, Boucot, Webby and Laurie1992, pp 313–314) of 10–25 metres. [KILN] plots slightly more towards the opposite end of the environmental gradient, due to the presence of shallower-water taxa, including Hesperorthis, Rostricellula, Camerella and Trigrammaria, between the core of the Paucicrura–plectambonitacean Biofacies and the Sowerbyella Biofacies, but still presents a deeper-water component with, amongst others, Leptaena and Eoplectodonta.
3. Conclusions
The Barr and Ardmillan succession contains some of the most diverse and varied Ordovician brachiopod assemblages associated with marginal Laurentia, due in part to their proximity to an active shelf margin (Ingham Reference Ingham, Fortey, Harper, Ingham, Owen, Parkes, Rushton and Woodcock2000). In particular, the Barr and Lower Ardmillan groups expose a mosaic of shallow to deep-water environments deposited in a fault-controlled basin, overstepping a dismembered ophiolite basement. Within the Barr Group, the outer shelf biofacies of the Confinis and Superstes formations represent the lowest and highest fossiliferous units within the group and are interrupted by the shallower-water, inner shelf biofacies of the Stinchar Formation. The broadly coeval, deeper-water facies of the Albany Group, south of the Stinchar Valley Fault, contain very low-diversity assemblages dominated by the minute, thin-shelled Sericoidea. In the upper part of the Lower Ardmillan Group, north of the Craighead Fault in the Craighead Inlier, the Craighead Formation contains a spectrum of deep and shallow-water biofacies. The brachiopod-dominated biofacies are comparable to those described and recently analysed from the Appalachian mountain belt and its western margin. The Girvan faunas, however, represent a much wider depth range, especially at the deeper-end of the spectrum, heralding the deep-water environments later in the Katian (Harper Reference Harper2001).
4. Acknowledgements
The authors thank Alan Owen (University of Glasgow) and Matthew Parkes (National Museums of Ireland, Dublin) for their most helpful reviews of the manuscript, and Maggie Cusack and Vicki Hammond for their efficient editorial work. YC thanks Gail Grey (BGS Library, Murchison House, Edinburgh) for access to the collection in her care. This paper is a contribution to IGCP 591 “The Early to Middle Palaeozoic Revolution”.
5. Supplementary Material
The data files for (1: List of the taxa from the Barr Group and Ardwell Subgroup; 2: Database for the early Sandbian analysis; 3: Database for the middle Sandbian–early Katian Ardwell Subgroup analyses), together with explanatory , are published as Supplementary Material with the on-line version of this paper. This is hosted by the Cambridge Journals Online (CJO) service, and can be viewed at http://journals.cambridge.org/tre






