

It has been an accepted view for over 75 years (e.g., Westoll Reference Westoll1951; Waterston Reference Waterston and Craig1965; Browne et al. Reference Browne, Smith and Aitken2002; Read et al. Reference Read, Browne, Stephenson, Upton and Trewin2002) that the uppermost part of the ‘Old Red Sandstone' (ORS; now the Kinnesswood and older formations) in Scotland and the Scottish Borders (Fig. 1) is Carboniferous in age. This is fundamental for understanding where to place the Devonian–Carboniferous boundary and its relationship to palaeoenvironmental change within this interval. Importantly, the Early Carboniferous Ballagan Formation (Fig. 2) in Scotland contains a number of globally significant fish, tetrapod and arthropod faunas (Smithson et al. Reference Smithson, Wood, Marshall and Clack2012; Clack et al. Reference Clack, Bennett, Carpenter, Davies, Fraser, Kearsey, Marshall, Millward, Otoo, Reeves, Ross, Ruta, Smithson, Smithson and Walsh2016) so understanding their time relationships is fundamental to understanding their evolutionary relationships. Following ongoing research on the Late Devonian–earliest Carboniferous rocks in Central Scotland and the Borders as part of the TW:eed Project (Tetrapod World: early evolution and diversification), coupled with new integrated palynological and fish occurrences in East Greenland, this age assignment can be questioned. This contribution will show that these hitherto earliest Carboniferous red-bed sections are demonstrably Devonian in age and that a full Tournaisian section is present in the Ballagan Formation of Scotland. A number of field sections in Central Scotland and the Borders were investigated during the TW:eed Project and here we report on those from Pease Bay, Burnmouth and along the Whiteadder Water at Preston (Fig. 1).

Figure 1 Map of the Ballagan Formation outcrop area in the Scottish Borders and NE England. The key localities of Burnmouth, Preston and Pease Bay are shown together with the Norham West Mains Farm borehole. (Green area of the Ballagan Formation is from the British Geological Survey DiGMapGB © NERC 2018. It contains Ordnance Survey data © Crown copyright and database right 2018.)
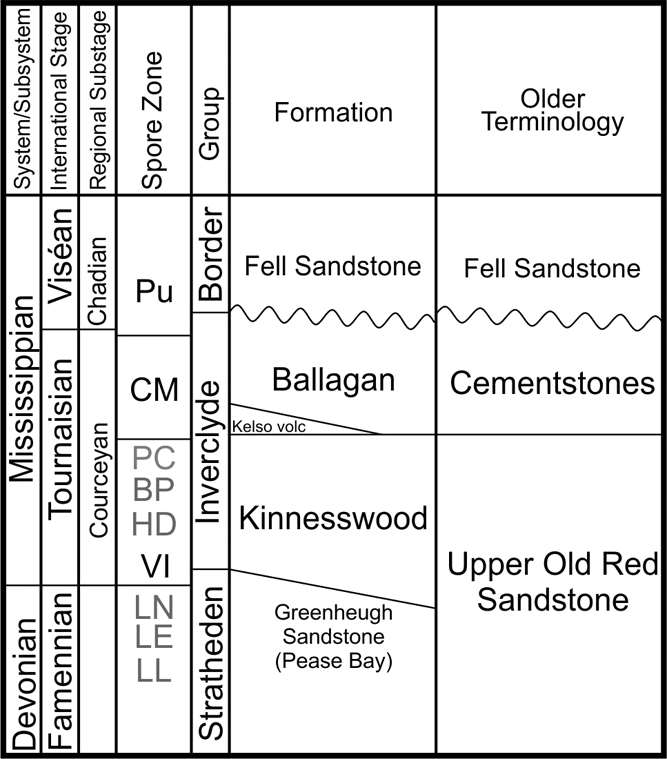
Figure 2 Lithostratigraphic units from the Borders area correlated with the established Late Devonian and Early Carboniferous chronostratigraphy. The spore zones are shown but only the Pu, CM and VI have been identified. The rest are conjectural. The current established formations are shown together with the older terminology as used in earlier literature. Largely after Waters et al. (Reference Waters, Somerville, Jones, Cleal, Collinson, Waters, Besly, Dean, Stephenson, Davies, Freshney, Jackson, Mitchell, Powell, Barclay, Browne, Leveridge, Long and McLean2011).
1. Historical context
In any major reinterpretation of this kind it is important to understand the historical roots as to how and why this Early Carboniferous consensus age for the uppermost ORS was achieved. The first documented discussion (Anon 1939) as to the possibility of a Carboniferous age for the uppermost ORS was at the 1939 British Association (BA) meeting (Dundee) when there was a scheduled discussion meeting on ‘the boundary between the Old Red Sandstone and the Carboniferous'. However, the start of this BA meeting coincided with the German invasion of Poland and, with it, a time of considerable national uncertainty. Added to this was the chaos of the evacuation of the local children, a general transport requisition and UK-wide troop mobilisation. So, the organisers decided to halt the meeting early on the 3rd September and the ORS–Carboniferous boundary discussion that was scheduled for the 5th September never took place. However, contributions were taken as read and reported without the merit of having had the scientific discussion. They were published as summaries (e.g., Westoll Reference Westoll1940) by six of the intended contributors (G. Hickling missing) who were a singularly prominent group of geologists and palaeontologists (W. Q. Kennedy, T. N. George, W. T. Gordon, T. S. Westoll, V. A. Eyles and M. Macgregor). Their views were that a practical boundary for the base of the Carboniferous was at the lowest shale-cementstone horizon, but that this, particularly in the W of Scotland, would include beds that lithologically would be regarded as Upper ORS; these W of Scotland sections (Eyles et al. Reference Eyles, Simpson and MacGregor1949) being atypical in that they show an intercalation of intervals of cementstones and ‘ORS' (red sandstone with calcrete nodules), i.e., the Kinnesswood Formation. These contributors were well aware that this was purely a lithostratigraphical definition and a practical convention. However, Westoll, as a vertebrate palaeontologist, reproduced the then new fish zonation of Säve-Söderbergh (Reference Säve-Söderbergh1934) from East Greenland and noted the common fauna to Scotland and, hence, a correlation between the Phyllolepis Series and the highest fossiliferous ORS from Dura Den (Fife) and Rosebrae (Moray) in what were apparently contiguous sequences. Westoll's argument being that since both Dura Den and Rosebrae were only a little below the accepted base of the Carboniferous then the overlying Remigolepis and Arthrodire Series in East Greenland might be contemporaneous with the lowest Carboniferous elsewhere (Fig. 3). This implied that the important, and, at that time, the earliest known tetrapod fauna from East Greenland was, in fact, Carboniferous in age. In other contributions Westoll generally referred to these tetrapods as Upper Devonian or high Upper Devonian or ? Early Carboniferous (both ages are quoted in Westoll Reference Westoll1938) or lowest Carboniferous (Westoll Reference Westoll1943). Säve-Söderbergh (Reference Säve-Söderbergh1934) had very much invited this interpretation by noting that the Phyllolepis Series was the highest Upper Devonian yet known with the Remigolepis and Arthrodire (Groenlandaspis) Series representing even younger Devonian rocks that were, as yet, unknown from elsewhere, perhaps corresponding to a gap in these other sequences. Westoll, recognising that the ORS-to-Carboniferous transition in Scotland was contiguous, simply moved the Remigolepis and Arthrodire Series into the Carboniferous. In any event, we can speculate that Westoll was probably always going to give a determined response as it was a member of the Stockholm School (of vertebrates; Schultze Reference Schultze2009) working in Greenland who had pre-empted the publication (Patterson & Fortey Reference Patterson and Fortey1999) of Westoll's own thesis on Permian fish from County Durham. A response to this reassignment of the East Greenland tetrapods to the Carboniferous was only made in 1948 by Jarvik, at the request of Säve-Söderbergh who had been debilitated by tuberculosis (Jarvik Reference Jarvik1996) from 1937 until his premature death in 1948. Jarvik (Reference Jarvik1948) gave a critical discussion of the age of the East Greenland tetrapods by systematically going through each ‘Series' and its fauna. This expressed disappointment that Westoll's supposition of a possible earliest Carboniferous age for the tetrapods had been adopted in textbooks such as Romer's Vertebrate Paleontology (Romer Reference Romer1945). Jarvik (Reference Jarvik1948) based his arguments on the occurrences of the complete fauna bar tetrapods (i.e., Remigolepis, Bothriolepis, Phyllolepis, rhizodonts and holoptychiids) and demonstrated that none of these forms were known from Carboniferous sections elsewhere in the world. This correlation was then reiterated (Jarvik Reference Jarvik1950) against the more precisely defined Devonian–Carboniferous boundary of the Heerlen Congress (Jongmans & Gothan Reference Jongmans and Gothan1937). In reality, these comparisons were always limited by fossil fish being restricted to certain facies so that the most important comparative sections were in Belgium and the Baltic. To an extent this was based on accepting that certain fish groups were restricted to the Devonian (e.g., placoderms); something that is now accepted (Sallan & Coates Reference Sallan and Coates2010) but was, at that time, not evident, with the possibility, then as now, for the existence of hold-over taxa (i.e., dead clades walking). This situation of these somewhat imprecise and ambiguous ages (e.g., Bütler Reference Bütler and Raasch1961; Jarvik Reference Jarvik and Raasch1961; Nicholson & Friend Reference Nicholson and Friend1976; Olsen & Larsen Reference Olsen and Larsen1993) for the East Greenland tetrapods continued without any additional information, such that the clarity of these initial correlations by Jarvik (Reference Jarvik1948, Reference Jarvik1950) was lost. This introduced element of uncertainty has meant that, subsequently, the tetrapods tended to be attributed to a rather ambiguous age of around the Devonian–Carboniferous boundary.

Figure 3 Stratigraphic columns showing the contrasting views of Säve-Söderbergh and Westoll of fish ranges against the Devonian–Carboniferous boundary chronology. The Greenland ranges are from Säve-Söderbergh (Reference Säve-Söderbergh1934). The strata in Greenland show the sequence of faunas from Phyllolepis, Remigolepis and Groenlandaspis, interpreted as entirely Devonian in age. At that time the Devonian–Carboniferous boundary was poorly defined, with the Strunian from Belgium being variously Devonian or Carboniferous. Subsequently, formal definition of the boundary has placed it in the Devonian, with the Famennian extending to the base Carboniferous and the Strunian relegated to a regional stage. In Scotland only Phyllolepis was recognised. Westoll (Reference Westoll1951) noted that this was from the uppermost part of the Scottish Devonian and below a conformable succession into the Carboniferous. Therefore, by correlation, the Devonian–Carboniferous boundary in Scotland was placed below the range of Remigolepis.
The dating and recognition of a defined Devonian–Carboniferous boundary was only resolved in East Greenland in 1999. This renewed interest was driven by the re-dating of the sections based on palaeomagnetism and geochronology (Hartz et al. Reference Hartz, Torsvik and Andresen1997) that indicated the tetrapods were Early Carboniferous and potentially as young as Viséan in age, which proved somewhat controversial (Hartz et al. Reference Hartz, Torsvik and Andresen1998; Stemmerik & Bendix-Almgreen Reference Stemmerik and Bendix-Almgreen1998). The problem was solved by palynology (Marshall et al. Reference Marshall, Astin and Clack1999; Streel & Marshall Reference Streel, Marshall and Wong2006; Astin et al. Reference Astin, Marshall, Blom, Berry, Vecoli, Clement and Meyer-Berthaud2010). This gave a clear position for the Devonian–Carboniferous boundary within the Obrutschew Bjerg Formation (on Stensiö Bjerg) and above the ranges of both Remigolepis (by 150m) and the in situ tetrapod Acanthostega (by 230m) that are pre-latest Famennian in age.
It is from this time that there was a divergence of view between the Stockholm School who regarded Remigolepis and related fish and tetrapod faunas as clearly Devonian in age (Fig. 3), whereas the same faunas, despite being absent from Scotland, were attributed to the early Carboniferous (Westoll Reference Westoll1951). For example, Waterston in the Geology of Scotland (1965, p. 302) simply notes that the Upper ORS persisted into the Tournaisian in Scotland, the rationale being that in Greenland the Remigolepis Zone was younger than anything known from Europe and, hence, fell into the Carboniferous. Similarly, Westoll compiled (Reference Westoll, House, Richardson, Chaloner, Allen, Holland and Westoll1977) the sections on Northern Britain for the Devonian Special Report of the British Isles. The Ballagan Formation section from Pease Bay to Cove in the Borders was regarded as particularly important as it had cementstones that were dated as being in the Schopfites claviger – Auroraspora macra (CM) spore zone (late Tournaisian) by Neves et al. (Reference Neves, Gueinn, Clayton, Ioannides, Neville and Kruszewska1973), succeeded by the Horse Road and Heathery Heugh Sandstones that were regarded as similar in facies to the Upper ORS. The Horse Road Sandstone was noted as containing lower Pa (sic) zone spores and was equivalent to Tn2 or Tn3 in Belgian nomenclature (Hance et al. Reference Hance, Poty and Devuyst2006). In fact, this is a misspelling of the Pu spore zone named after Lycospora pusilla, which has an inception at about the Tournaisian/Viséan boundary. However, it should be noted that L. pusilla had not been reported by Clayton (Reference Clayton1971, p. 597) from the Horse Road Sandstone at outcrop but only in the Birnieknowes Borehole from below the base of a ?Horse Road Sandstone correlative. More recent work by Stephenson et al. (Reference Stephenson, Williams, Monaghan, Arkley, Smith, Dean, Brown and Leng2004a) gives only a single occurrence of L. pusilla, and this is from above the Horse Road Sandstone Member. This correlation and the name error of Pa for Pu zone has been reported in subsequent accounts of the section (Grieg & Davies in Friend & Williams Reference Friend and Williams1978; Dineley & Metcalf Reference Dineley and Metcalf1999).
In part, this view of a young ‘ORS' was significantly influenced by the palynological work of Clayton (Reference Clayton1971) on the Ballagan Formation cementstone section at Pease Bay. The section was recognised as thin with the then unnamed CM spore zone spores (Schopfites claviger and Auroraspora macra) being regarded as from the upper part of the Ballagan in what was seen as a contiguous section. Clayton discussed two possibilities – firstly that the ORS beneath the cementstones was in the early Carboniferous or secondly that the ‘lower' Tournaisian was extremely condensed. This discussion was always influenced by the consensus (Westoll Reference Westoll1951) that the top of the Kinnesswood Formation was well within the Tournaisian. In addition, stratigraphic palynology was in its infancy, such that there were few independent dated Tournaisian sections with which it were compared. This placement of the CM spore zone in the later part of the Tournaisian has been generally adopted (e.g., Stephenson et al. Reference Stephenson, Williams, Monaghan, Arkley and Smith2002, Reference Stephenson, Williams, Monaghan, Arkley, Smith, Dean, Brown and Leng2004a, Reference Stephenson, Williams, Leng and Monaghanb). Independent age evidence for the inception of S. claviger comes from Ireland where in the Baunta borehole (Keegan Reference Keegan1981; Higgs et al. Reference Higgs, Clayton and Keegan1988) there is a CM assemblage in the lower part of the Polygnathus mehli conodont zone. In the Tatestown prospect boreholes of County Meath, Ireland, the base of the CM spore zone is placed (Keegan cited in Andrew & Poustie Reference Andrew, Poustie, Andrew, Crowe, Finlay, Pennel and Pyne1986) in the slightly older Lower Pale Beds from the underlying Pseudopolygnathus multistriatus conodont zone. There is a further constraint from Cumbria where Schopfites delicatus but not S. claviger occurs in the lower part of the Pinskey Gill Formation (Johnson & Marshall Reference Johnson and Marshall1971; Holliday et al. Reference Holliday, Neves and Owens1979; Welsh Reference Welsh1979) together with conodonts (Varker & Higgins Reference Varker and Higgins1979; Higgins & Varker Reference Higgins and Varker1982; Varker & Sevastopulo Reference Varker, Sevastopulo, Higgins and Austin1985) that are ascribed to Fauna A of mid-Courceyan age (i.e., mid-Tournaisian). Schopfites claviger does occur in the Stone Gill Limestone Formation that, together with the underlying Marsett Formation conglomerates, unconformably overlie the Pinskey Gill Formation (Waters et al. Reference Waters, Somerville, Jones, Cleal, Collinson, Waters, Besly, Dean, Stephenson, Davies, Freshney, Jackson, Mitchell, Powell, Barclay, Browne, Leveridge, Long and McLean2011). The conodonts in the upper part of the Stone Gill Limestone Formation are from the Chadian Taphrognathus Zone. This occurrence of the Chadian, which is the regional substage at the base of the Viséan, is confirmed by the presence of Lycospora pusilla (i.e., Pu zone) in the Stone Gill Limestone Formation. This places the delicatus to claviger transition in the mid- to late Tournaisian interval, although Welsh (Reference Welsh1979) regarded S. delicatus as a variant of S. claviger and placed the base of the CM zone at the inception of the former with its earlier species inception.
The CM spore zone has become effectively synonymous with the Ballagan Formation with its somewhat monotonous palynological assemblage. This assemblage has been reported from throughout the Ballagan Formation (Sullivan Reference Sullivan1968; Clayton Reference Clayton1971; Neves et al. Reference Neves, Gueinn, Clayton, Ioannides, Neville and Kruszewska1973; Neves & Ioannides Reference Neves and Ioannides1974) in terms of both stratigraphic and geographic range. The most detailed recent accounts are by Stephenson et al. (Reference Stephenson, Williams, Monaghan, Arkley and Smith2002, Reference Stephenson, Williams, Monaghan, Arkley, Smith, Dean, Brown and Leng2004a, Reference Stephenson, Williams, Leng and Monaghanb) where range charts detail the occurrence of Schopfites claviger throughout the formation, including the Glenrothes, Birnieknowes and East Dron boreholes (e.g., Millward et al. Reference Millward, Davies, Brand, Browne, Bennett, Kearsey, Sherwin and Marshall2019) where the succession continues down to the Kinnesswood Formation. At Burnmouth, S. claviger is reported (Scott et al. Reference Scott, Galtier and Clayton1984) from 70m above the base of the formation to 45m below its top.
The current view of stratigraphic nomenclature and spore zonation in the Late Devonian and Early Carboniferous of the Scottish Borders is shown in Figure 2. It is important to note which spore zones have actually been found in the region (bold) in contrast to the spore zones that have been recognised from sections outside Northern Britain through the same time interval.
2. Methods
All the palynological samples from Burnmouth, Pease Bay and Preston were processed by standard techniques with 5g of roughly crushed rock (grey to black or dark green mudstone) treated with 30% hydrochloric acid (HCl) to remove carbonates, followed by decant washing to neutral pH. They were then demineralised in 60% hydrofluoric acid (HF) followed by decant washing again to neutral and sieved at 15μm. The samples were then placed in glass beakers and briefly boiled in 30% HCl to solubilise neoformed fluorides that were then removed by diluting into a large volume of water and resieving. The samples were then vialed and strew slides mounted in Elvacite 2044™. Any amorphous organic matter (AOM) was removed from the samples with a 15–30s treatment using a Sonics and Materials ultrasonic probe followed by resieving at 15μm. The AOM preferentially fragments through the action of the probe and can be removed as a smaller size fraction.
3. Pease Bay
It was from the Pease Bay section (Fig. 1) that a ?Remigolepis specimen was described by Andrews (Reference Andrews1974). This had been collected by Sir Frederick Stewart in 1957 from a fallen block of ‘ORS' lithology and was only questionably assigned to the genus. However, reinvestigation in the field against the locality details given by Stewart (in Andrews Reference Andrews1974) and the rock description of Andrews (Reference Andrews1974, p. 311) gives a fairly obvious level from which the specimen originated:
… the sandstone matrix is of very irregular texture and colour, with lumps of dark calcareous matter and small grey-green clay galls. It had probably fallen from a similar bed of cornstone in the cliff above, lying about 6m below the top of the Cornstone Group from a clay gall sandstone….
We have also had the specimen re-evaluated from the illustrations and it can be described confidently as Remigolepis (Ritchie, pers. comm. 2016). The presence of this specimen was noted by Westoll (Reference Westoll, House, Richardson, Chaloner, Allen, Holland and Westoll1977) and it was speculated that it might prove significant for dating the Remigolepis Series in East Greenland. We will now show that the converse is in fact the case. Following the work of Blom et al. (Reference Blom, Clack and Ahlberg2005), it is now possible to match (Fig. 4) the exact range of Remigolepis against both the lithostratigraphy and the palynological record, all from East Greenland. These occurrences of Remigolepis spp. are from within the Aina Dal to Britta Dal Formations. Importantly, the spore Retispora lepidophyta has its first occurrence in the Upper Britta Dal Formation with an inception just 10m above Acanthostega from the in situ locality on Stensiö Bjerg. Retispora lepidophyta has a globally restricted distribution to the latest Famennian (Streel & Marshall Reference Streel, Marshall and Wong2006; Streel Reference Streel and Königshof2009), which means that in East Greenland Remigolepis becomes extinct in the early latest Famennian. Extending this correlation to the UK shows that the uppermost ORS in Pease Bay is within the age range of mid–early latest Famennian. The palynological assemblages in the Ballagan Formation from Pease Bay (Clayton Reference Clayton1971) contain many specimens of Claytonispora distincta. Our data from two long Ballagan Formation sections (e.g., Burnmouth, Fig. 5) show that C. distincta is restricted to the lowest 140m of the formation. Hence, the Pease Bay Ballagan Formation is also from the lower part of the formation. The Ballagan Formation section at Pease Bay (Grieg Reference Grieg1988) appears to be thin but is, in fact, truncated by the Cove Fault with the Pu zone absent and the overlying Perotrilites tessellatus – Schulzospora campyloptera (TC) spore zone occurring in the shale overlying the Kip Carle Sandstone. This means that the missing latest Famennian (∼2 My) should be present within the super mature Stage 6 calcrete (Eastern Hole Conglomerate) present in Pease Bay (Andrews et al. Reference Andrews, Turner, Nabi and Spiro1991; Wright et al. Reference Wright, Turner, Andrews and Spiro1993) and other Upper Kinnesswood Formation sections (Glenrothes Borehole, Wright et al. Reference Wright, Turner, Andrews and Spiro1993; Carham Limestone, Carruthers et al. Reference Carruthers, Burnett and Anderson1932). The terrestrial palaeoclimate record from the Famennian in East Greenland (Astin et al. Reference Astin, Marshall, Blom, Berry, Vecoli, Clement and Meyer-Berthaud2010) shows a very arid interval through the mid- and late Famennian Britta Dal Formation. But the latest Famennian Stensiö Bjerg Formation had greater seasonality, with wetter wet times and drier dry times, as shown by the presence of lakes and calcretes rather than a stack of vertisols. This can be attributed to the latest Famennian glaciations (as reviewed in Lakin et al. Reference Lakin, Marshall, Troth, Harding, Becker, Königshof and Brett2016), which, in the Borders, showed the cumulative far-field response of a composited supermature calcrete representing a significant arid hiatus in excess of several hundred thousand years per calcrete horizon (Wright et al. Reference Wright, Turner, Andrews and Spiro1993). Hence, the top of the Kinnesswood Formation at Pease Bay is placed at the top of the range of Remigolepis to reflect a view that the most likely condensed interval would be the latest Famennian glacial cycles.

Figure 4 New compilation of the Late Devonian and Early Carboniferous lithostratigraphy from East Greenland. This shows the range of Remigolepis spp from Blom et al. (Reference Blom, Clack and Ahlberg2005) plotted against the identified spore zones (GF to Pu). The Devonian–Carboniferous boundary is within the Obrutschew Formation (OBF). Compiled by direct section measurement from Astin et al. (Reference Astin, Marshall, Blom, Berry, Vecoli, Clement and Meyer-Berthaud2010) from Stensiö Bjerg and Nathorst Bjerg below the base Carboniferous and then from Celsius Bjerg for the Early Carboniferous. Other data from Marshall et al. (Reference Marshall, Astin and Clack1999) and Vigran et al. (Reference Vigran, Stemmerik and Piasecki1999). The presence of Remigolepis (R) within a section that runs into the CM spore zone shows that the Kinnesswood Formation is Famennian and younger than early latest Famennian in age. It is placed at the top of the range of Remigolepis based on the argument that the latest Famennian glacial–interglacials are the time gap represented by the calcrete.

Figure 5 The lowest 300m of the Ballagan Formation from Burnmouth, from Bennett et al. (Reference Bennett, Kearsey, Davies, Millward, Clack, Smithson and Marshall2016) and Kearsey et al. (Reference Kearsey, Bennett, Millward, Davies, Gowing, Kemp, Leng, Marshall and Browne2016). The occurrences of key spores enabled the VI, HD and Cl1 spore zones to be identified. Significantly, this enables application of the integrated spore and conodont zonation from Poland (Matyja et al. Reference Matyja, Turnau and Żbikowska2000) and a better correlation to the Early Carboniferous than using the more limited data from Ireland. Taxonomic citations are in Table 1.
A further stratigraphically important fish known from the Pease Bay area (Miles Reference Miles1968) is Grossilepis brandi from Hazeldean Burn, close to Siccar Point. It is related to other Grossilepis species, which, in the Baltic (Esin et al. Reference Esin, Ginter, Ivanov, Lebedev, Luksevics, Avkhimovitch, Golubtsov and Petukhova2000), are found in the Frasnian. This is the only evidence for the age of the base of the Upper ‘ORS' Greenheugh Sandstone Formation (Fig. 2) in Scotland and implies that it lies within the Frasnian.
4. Burnmouth
The longest and most complete surface section of the Ballagan Formation (Fig. 5) is in the intertidal zone at Burnmouth where it totals some 520m of near-vertical strata (Bennett et al. Reference Bennett, Kearsey, Davies, Millward, Clack, Smithson and Marshall2016; Kearsey et al. Reference Kearsey, Bennett, Millward, Davies, Gowing, Kemp, Leng, Marshall and Browne2016) from an apparently conformable contact with the Kinnesswood Formation to a truncated top beneath the Fell Sandstone. This has previously been ascribed a CM late Tournaisian spore zone age (Scott et al. Reference Scott, Galtier and Clayton1984). During the initial discovery of tetrapods from the Burnmouth section (Smithson et al. Reference Smithson, Wood, Marshall and Clack2012) we recovered a palynological residue from a tetrapod bed at the base of the section. This contained a sparse simple assemblage lacking Schopfites claviger, the spore which defines the base of the CM zone. Dominant in the assemblage were specimens of Retusotriletes incohatus and Plicatispora scolecophora (Fig. 6), which enabled us (Smithson et al. Reference Smithson, Wood, Marshall and Clack2012) to place it within the Vallatisporites verrucosus – Retusotriletes incohatus (VI) spore assemblage of earliest Carboniferous age. The lithologies in the underlying Kinnesswood Formation are dominantly red and yellow sandstones with no preserved organic matter and so it proved impossible to find palynomorphs. The only fossils from this location are the rare fish fragments collected by Stan Wood that include holoptychiid scales.

Figure 6 Illustration of important stratigraphic spores from Burnmouth (1–10) and the Whiteadder at Preston (11–21). All figured spores are in the palynology collections of the British Geological Survey, Keyworth. Microscope co-ordinates refer to Olympus BHS-313 No. 210685 in the School of Ocean and Earth Science, University of Southampton. England Finder co-ordinates (e.g., R20–22) are also provided. Scale=10 μm. 1. Plicatispora scolecophora, Burn W-10 W Hbr 14, (130, 10), S29-2 ×750. 2. Retusotriletes sp, Burn W-10 W Hbr 27, (121, 11), R20-2, ×750. 3. Cristatisporites hibernicus, Burn 13-6 W Hbr 45, (125.1, 7.4), U24-4, ×750. 4. Grandispora upensis, Burn 12-10 E Hbr 10, (125.7, 10.8), R25-4, ×750. 5. Claytonispora distincta, Burn 13-6 W Hbr 22, (126.1, 16), M25-2, ×750. 6. Neoraistrickia cymosa, Burn 12-10 E Hbr 1, (125.3, 10.2), S25-1, ×750. 7. Prolycospora claytonii, Burn 16-4 E Hbr 3, (118.2, 12.6), P17-4, ×1000. 8. Prolycospora claytonii, Burn 16-4 E Hbr 3, (116, 14.8), N15-4, ×1000. 9. Schopfites delicatus, Burn 12-10 WH 14, (129.5, 13.7), O29-1, ×750. 10. Schopfites spp approaching Schopfites claviger in size of sculpture, Burn W Hbr 40, (121.8, 14.6), N21-4, ×750. 11. Anapiculatisporites baccatus, Burn 12-10 S Cliffs 215, (125.9, 10.3), S25-2, ×500. 12. Retispora lepidophyta, Preston 17-5-4 (3), (121.2, 11), R21-1, ×750. 13. Vallatisporites pusillites, Preston 17-5-4 (4), (126, 6.7), V25-4, ×500. 14. Diducites versabilis, Preston 17-5-4 (3), (114.7, 23), D14-3, ×500. 15. Tergogobulasporites immensus, Preston 17-5-4 (3), (133.7, 19.8), ×250. 16. Rugospora radiata, Preston 17-5-5 (3), (117.8, 7.1), ×500. 17. Isolated bifurcate spine, Preston 17-5-4 (2), (114.6, 21.5), ×250. 18. Retispora macroreticulata, Preston 17-5-4 (2), (111.6, 3.7), ×500. 19. Ancyrospora with multifurcate spines, Preston 17-5-4 lc (3), (113.6, 11.8), ×250. 20. Isolated spine from megaspore sized Nikitinsporites, Preston 17-5-4lc (3), (139, 16.9), ×250. 21. Hystricosporites with bifurcate tipped spines, Preston 17-5-4 (3), (120.6, 12), ×250.
As part of the TW:eed project, we undertook detailed logging of the Ballagan Formation at Burnmouth, with particular attention to the base of the formation. The recognition that there were earliest Tornaisian VI zone palynomorphs at the base of the formation and reported late Tournaisian CM spores (Scott et al. Reference Scott, Galtier and Clayton1984) at a height of 70m in the formation implied that a significant fault or hiatus should be present. However, detailed investigation, a drone survey, LiDAR and logging has shown no evidence for any fault or hiatus along the line of our logged section. Minor faults were identified, but these do not cut the section. The critical parts of the section were studied twice a year during low spring tide. These rocks might appear entirely covered by algal growth, but at the lowest levels that are only exposed during the spring tides the dominant cover is the brown seaweed Laminaria. The upper blades of these weeds can be turned to reveal the underlying holdfast and stipe. These holdfasts are spaced apart with, in contrast to higher levels of the shore, little intervening encrusting cover such that rock and particularly the bedding becomes visible. This reveals that the section is entirely contiguous. It could still be speculated that there were significant hiatuses present within palaeosols similar to the Stage 6 calcrete at Pease Bay. However, there is now a detailed published record of the palaeosols from the Burnmouth section (Kearsey et al. Reference Kearsey, Bennett, Millward, Davies, Gowing, Kemp, Leng, Marshall and Browne2016) in addition to the much better record from the comparative section in the Norham West Mains Farm borehole. This shows that there are no identifiable palaeosols present at Burnmouth from the base of the Ballagan Formation to the inception of Schopfites delicatus. Instead, the last bed of the Kinnesswood Formation (Fig. 7) is directly overlain by a cementstone and fine sandstones with wave ripples. This indicates that there was no significant erosional unconformity at the base of the Ballagan Formation. Some 64 palaeosols have been identified higher in the section, but these are vertisols, inceptisols, gleyed inceptisols and entisols. None of these palaeosols have calcrete levels present within them and there are no thick higher-stage calcretes. These Ballagan palaeosols only represent hiatuses of 10–1000 years. The palaeosol types that are present formed relatively rapidly, and are evenly distributed in both their individual thickness and spacing throughout the formation. This shows that there is no possibility of a hidden hiatus or hiatuses within the section. In addition to the detailed logging, there was extensive focused sampling for palynomorphs from the Ballagan Formation at Burnmouth. These were located along the logged section line using differential GPS (Global Positioning System) with an accuracy of 2cm. From this sample set we processed some 275 samples for palynology (plus a further 82 rejected on lab inspection), of which 107 (39%) were productive. Compared to the spore recovery (85%) from the correlative section in the Norham West Mains Farm borehole (Fig. 1), the spore recovery is disappointing. Within the near-vertical section it proved very difficult to sample adequately the very thin mudstone laminae that are present on partings between the sandstones; these are readily altered along bedding plane surfaces and are often the preferential horizon for the extant local burrowing infauna. The best recovery was at the base of the section and through the interval to the S side of the bay. Many of the samples contain organic matter, although this is often dominated by black phytoclasts with relict spores, either the result of recent alteration or immediate post-depositional Carboniferous weathering. In general, the palynological assemblages are somewhat simple and dominated by S. delicatus and Auroraspora macra, together with simple retusoid spores. Clearly there is an element of the Ballagan Formation flora (Neves & Belt Reference Neves and Belt1971; Van der Zwan et al. Reference Van der Zwan, Boulter and Hubbard1985; Stephenson et al. Reference Stephenson, Williams, Monaghan, Arkley and Smith2002, Reference Stephenson, Williams, Monaghan, Arkley, Smith, Dean, Brown and Leng2004a) that was dry-adapted with a restricted spore assemblage. But our intensive sampling programme revealed the existence of rarer samples that contained a more diverse microflora that included key taxa used in international correlation. Figure 5 shows the sequence of inceptions of these key taxa for the lower part of the Burnmouth section. Inceptions include Cristatisporites hibernicus, (32.75m), Anaplanisporites baccatus (50.0m), Claytonispora distincta (70.75m; formerly Umbonatisporites or Dibolisporites distinctus), Grandispora upensis (96.25m) and Neoraistrickia cymosa (98.55m) with a similar sequence of inceptions as found in both southern Ireland (Higgs et al. Reference Higgs, Clayton and Keegan1988), SW Britain (Hennessy & Higgs Reference Hennessy and Higgs1999), South Wales (McNestry Reference McNestry1988) and Belgium (Higgs et al. Reference Higgs, Dreesen, Dusar and Streel1992). This enables us to recognise VI and Krauselisporites hibernicus – Claytonispora distincta (HD) spore zones. Significantly, there is the inception of Schopfites as S. delicatus at 33.75m within these ranges. Identification of this as S. claviger would lead to recognising this as the base of the CM zone (i.e., the late Tournaisian). But, this gives a conflicting result as this late Tournaisian marker would be either considerably below its normal range or the inceptions of the VI and HD zone spores were considerably delayed. However, these early Tournaisian spores still appeared in the correct sequence as the section transitions from the fluvial Kinnesswood Formation to the coastal matrix of Ballagan environments. The answer is that the spore reported as S. claviger through much of the Ballagan Formation is the species with much smaller sculpture known as S. delicatus and, as such, occurs below the range of S. claviger (Higgs Reference Higgs1975; Higgs et al. Reference Higgs, Clayton and Keegan1988). This distinction was first discussed by Keegan (Reference Keegan1981) and then reiterated in Higgs et al. (Reference Higgs, Clayton and Keegan1988), although some authors regard S. delicatus as a variant of S. claviger (Welsh Reference Welsh1979). In Burnmouth, S. delicatus is only replaced significantly up-section by forms that approach S. claviger as defined by the mean size of sculpture (e.g., Fig. 8; 257.9m). Hence, it would appear that S. claviger has been generally used within the Ballagan Formation for S. delicatus with its smaller more gracile sculpture and before the significance of this distinction was established. Realistically, this distinction is quite difficult to apply, as any method that requires averages of micron-scale measurement of spore sculpture does not fit easily into a zonation that is otherwise based on inceptions of distinctive species. The implication for time correlation within the Ballagan Formation is that many sections that have been attributed to the CM zone and, hence, a late Tournaisian age on the basis of S. claviger are, in fact, somewhat older. Another spore present is A. baccatus, which has an inception at 50m just above that of S. delicatus. It is the in situ microspore of the creeping lycopod Oxroadia (Bateman Reference Bateman1992; Stevens et al. Reference Stevens, Hilton, Rees, Rothwell and Bateman2010) that is very common in the Ballagan Formation and appears adapted to less stable environments. In Ireland, A. baccatus is not present on the clastic margin but has an inception in the higher part of the Spelaeotriletes pretiosus – Raistrickia clavata (PC) zone.

Figure 7 Logged section and annotated field photo of the contact between the Kinnesswood and Ballagan Formations, Burnmouth. The field photo is of near-vertical strata with way up to the left. The uppermost Kinnesswood Formation is a cross-bedded sandstone containing small rhizocretions and abundant rootlet structures. It has a gently undulating top surface without obvious truncation and no calcretisation or pre-Ballagan Formation weathering. The Ballagan Formation is conformably bedded on this surface with cementstones present at the contact that must represent flooding by a lagoon that infilled depressions on the top of the Kinnesswood Formation. The log shows a concentration of wave ripples inter-bedded with these first cementstones, again showing the presence of shallow standing water. There are dark-coloured mudstones at the contact that were prepared for palynology, but only residual black phytoclasts were present.

Figure 8 Measurements of the mean height and width of the sculpture of two populations of Schopfites from Burnmouth. The specimens with smaller sculpture (the mean is the six-point star) are from 71.85m and assigned to Schopfites delicatus. The larger sculpture is from 257.9m within the Cl 1 zone and approaching that of Schopfites claviger in size. The mean is the open cross. Measurement as per Higgs et al. (Reference Higgs, Clayton and Keegan1988), except that the means are from multiple sculptural elements on individual spores rather than means of many spores from multiple samples.
In the Western European spore zonation, the zones above HD are defined on the inception of various species of Spelaeotriletes and Raistrickia. However, these species are not abundant in the Borders, occurring only sporadically. At Burnmouth, the VI spore zone can be identified at the base of the Ballagan Formation, with the Pu zone occurring in other sections (Millward et al. Reference Millward, Davies, Brand, Browne, Bennett, Kearsey, Sherwin and Marshall2019) at about the level of the Fell Sandstone Formation, which shows that a complete Tournaisian section is present. However, within this section Spelaeotriletes is never abundant. This contrasts with both southern Ireland (Higgs et al. Reference Higgs, Clayton and Keegan1988) and the Bristol area of England (Hennessy & Higgs Reference Hennessy and Higgs1999) where they can make up 10% of the assemblage in a marginal shelf environment and are sufficiently diverse to define an evolutionary lineage (Brittain & Higgs Reference Brittain and Higgs2007). They are similarly abundant in the fully terrestrial Tournaisian in East Greenland (Vigran et al. Reference Vigran, Stemmerik and Piasecki1999) and Nova Scotia (Utting et al. Reference Utting, Keppie and Giles1989) where they can comprise 90% of the palynological assemblage. Clearly, there is an element of environmental control on the abundance of these taxa in the Ballagan Formation. This focuses attention on the validity of single inceptions to define zones; as key taxa increasingly appear to occur with different sequences of first appearance when investigated from different areas. For example, Claytonispora distincta occurs together with Retispora lepidophyta in Devonian rocks in western Canada (McGregor and Utting in Utting et al. Reference Utting, Keppie and Giles1989, p. 130). Other exceptions to ranges between the UK, Ireland and Eastern Europe are detailed in Clayton & Turnau (Reference Clayton and Turnau1990).
The next significant inception as regards palynological zonation (Fig. 5) is that of Prolycospora claytonii, with an inception at 189.9m and with a local acme (48%) at 257.9m. With only a few records in Western Europe (Clayton & Turnau Reference Clayton and Turnau1990), P. claytonii has not been regarded as important for the palynological subdivision in the British Isles. It is distinctly different from Prolycospora rugulosa, which has been recognised in Western Europe but is tripapillate (Butterworth & Spinner Reference Butterworth and Spinner1967; Higgs et al. Reference Higgs, Clayton and Keegan1988). Prolycospora claytonii was a major component in palynological assemblages from Eastern Europe (Turnau Reference Turnau1978; Avchimovitch & Turnau Reference Avchimovitch and Turnau1994), primarily Poland and Belarus, and forms the basis for an alternative zonation. More importantly, the zonation from Poland is tied to the international conodont scale (Matyja et al. Reference Matyja, Turnau and Żbikowska2000) with palynomorphs, conodonts and ostracods in the same sections. This contrasts with the limited conodont information available from Ireland that produces a local zonation based largely on shallow water and endemic forms. The inception of Prolycospora at Burnmouth is used to place the Cl 1 zone base at 189.9m; this is somewhat below the base of the late Tournaisian in terms of the established Tournaisian conodont zone. Applying the Polish zonation to Burnmouth also enables other approximate ties to be made to the conodont zonation and giving some element of precision for correlation into the marine sequences. In the Polish zonation (Matyja et al. Reference Matyja, Turnau and Żbikowska2000), spore zone Convolutispora major 1 (Ma1) is placed at the inception of Cristatisporites hibernicus, with Convolutispora major 2 (Ma2) and HD both coincident with the base of Claytonispora distincta. This places the base of the sandbergi conodont zone just below the inception of Ma2/HD and is also the base of the Alum Shale (Hartenfels et al. Reference Hartenfels, Hartkopf-Fröder, Herbig, Becker and Esteban Lopez2016). This is an episode of marine warming and stratification coincident with the Miller Diamictite/Soutklouf Shale in South Africa (see review in Lakin et al. Reference Lakin, Marshall, Troth, Harding, Becker, Königshof and Brett2016) and the ending of a Tournaisian glaciation. In Western Europe, this is marked by an increase in spore diversity with the inception of new species. The next conodont zone that can be placed is the typicus zone, just above the base of spore zone Cl 1.
5. Whiteadder Water, Preston
This re-zonation of the Ballagan Formation from Burnmouth with the VI spore zone at its base followed by the entire Tournaisian causes an immediate problem as, by implication, the underlying Kinnesswood Formation has to be mostly Devonian in age. This is supported by the presence of the Famennian fish Remigolepis at Pease Bay. However, to resolve this issue, what is required is direct, unqualified and independent age evidence from the Kinnesswood Formation. There have been continuing attempts to find palynomorphs in the Kinnesswood and underlying Upper ‘ORS' formations but none have succeeded. The recognition in 1999 of the Devonian–Carboniferous boundary in terrestrial sections in East Greenland gave a sedimentological motif of a distinctive prolonged climatically wet interval (Marshall et al. Reference Marshall, Astin and Clack1999) that hopefully could be identified within the Scottish sections. But despite the first author searching across Scotland, no similar motif could be identified and no spores have been found. There has been more success in the offshore hydrocarbon wells with a number of records of Retispora lepidophyta. But these are all based on drill cuttings from rather old wells in sand-rich sequences and have not yet enabled a clear definition of the boundary with respect to the lithostratigraphy.
It was from the Whiteadder Water at Preston that the first ever palynological assemblage from Upper ‘ORS' strata underlying the Ballagan Formation was discovered in a section where thin mudstones are interbedded with volcaniclastic rocks from the Kelso Volcanic Formation. This palynological assemblage is regarded as very significant and, together with the re-zonation of the Ballagan Formation and the Remigolepis specimen from Pease Bay, provides a coherent narrative. To place this in context, it is the first ever palynological assemblage from the onshore Upper ORS from Northern Britain and, as such, is reported here before being fully documented elsewhere. Diverse and well preserved Famennian palynological assemblages are rare in Western Europe and this new assemblage contains many examples of grapnel-tipped spores that are otherwise poorly described.
The main elements of the assemblage are provided in Table 2 and illustrated in Figure 6, with ranges against the established spore zonation shown in Figure 9. The assemblage contains a diversity of the three main genera of grapnel-tipped spores: Ancyrospora, Hystricosporites and Nikitinsporites. The latter is only present as an isolated spine from a megaspore. All three of these genera become extinct at the Devonian–Carboniferous boundary. Other species present within the assemblage that also terminate at the Devonian–Carboniferous boundary include Diducites versabilis, Rugospora radiata and Vallatisporites pusillites. Also present is the megaspore Tergobulasporites immensus, which is Famennian in age (Turnau Reference Turnau2002) and reportedly became extinct before the first appearance of Retispora lepidophyta. Retispora macroreticulata is present, which is again Famennian in distribution, but becomes extinct before the Devonian–Carboniferous boundary and at the base of the Retispora lepidophyta – Knoxisporites literatus (LL) (Maziane et al. Reference Maziane, Higgs and Streel1999) or Retispora lepidophyta – Verrucosisporites nitidus (LN) (Higgs et al. Reference Higgs, Clayton and Keegan1988) spore zones. Also present are rare specimens of R. lepidophyta, which has an inception in the latest Famennian and also became extinct at the Devonian–Carboniferous boundary. There are not enough specimens of R. lepidophyta to acquire meaningful data on its diameter, but those present are small in size (Fig. 6(12), 52μm diameter) and close to Retispora lepidophyta minor in diameter, which characterises the base of the species range (Maziane et al. Reference Maziane, Higgs and Streel2002). This places it in the LL spore zone or younger accepting the presence of T. immensus. Absent from the assemblage are Indotriradites explanatus and Verrucosisporites nitidus. These define the bases of the younger LE and LN spore zones, respectively (but see Prestianni et al. Reference Prestianni, Sautois and Denayer2016 for further discussion of the LN zone), and implies a position in the LL zone of early latest Famennian age. So, the assemblage is unequivocally Famennian and, more specifically, early latest Famennian in age.
Table 1 Spores from the Ballagan Formation at Burnmouth. Taxonomic citations not in the references can be found in Playford & Melo (Reference Playford and Melo2012) or Higgs et al. (Reference Higgs, Clayton and Keegan1988).

Table 2 Spores from Kinnesswood Formation locality Whiteadder Water, Preston. Taxonomic citations not in the references can be found in Playford & Melo (Reference Playford and Melo2012) or Higgs et al. (Reference Higgs, Clayton and Keegan1988).


Figure 9 Range chart of spores from the productive Kinnesswood Formation sample in the Whiteadder at Preston. The sample is clearly Devonian in age as Retispora lepidophyta, Ancyrospora spp., Hystricosporites spp., Nikitinsporites spp., Rugospora radiata, Diducites versabilis and Vallatisporites pusillites all become extinct exactly at the Devonian–Carboniferous boundary. The age is more specifically latest Devonian from the range base of R. lepidophyta. Retispora macroreticulata becomes extinct (Maziane et al. Reference Maziane, Higgs and Streel1999) in the very base of the LL zone, also constraining the age. Tergobulasporites immensus further constrains the age to early latest Famennian as it is not reported to overlap the age range of R. lepidophyta. The absence of Indotriradites explanatus and Verrucosisporites nitidus supports an assignment to the LL spore zone rather than LE or LN.
Identification of an LL palynological assemblage is very significant as it enables the key stratigraphic markers in the uppermost Kinnesswood Formation to be time-correlated in the section at Pease Bay. The Whiteadder Water samples are interbedded with volcaniclastic debris and this also demonstrates that the Kelso Lavas were, at least in part, from the LL spore zone. However, this section on the Whiteadder is of little further value for establishing stratigraphical correlations as the beds are at a steep angle and lie within a fault zone. A more tractable although now poorly exposed section is 18km to the S at Carham and was described by Carruthers et al. (Reference Carruthers, Burnett and Anderson1932). The section (Fig. 10) includes a very significant Stage 6 dolomitic calcrete, which includes a 60-cm-thick central band of pink chert, i.e., a silcrete. It is reported as up to 6m in thickness (normally 3m) and is present above the Kelso Lavas from which it is separated by a thin sedimentary intercalation. The calcrete has been extensively exploited for lime production and was well exposed at the time of the primary survey (Clough Reference Clough1888). Above the calcrete there is a poorly exposed sequence of Ballagan Formation that was formerly quarried (c.1860) in the Shidlaw Tile Works (Carruthers et al. Reference Carruthers, Burnett and Anderson1932). All these units were previously well exposed with stratigraphical continuity in cuttings along the then operational North Eastern Railway. These stratigraphic relationships have been re-examined by the current authors and are as described by Carruthers et al. (Reference Carruthers, Burnett and Anderson1932). The calcrete is a very significant stratigraphic marker in the Borders and Midland Valley of Scotland with the occurrence of a single Stage 6 calcrete in the Glenrothes borehole (Wright et al. Reference Wright, Turner, Andrews and Spiro1993), Pease Bay (Andrews & Nabi Reference Andrews and Nabi1998) and South Ayrshire (Burgess Reference Burgess1961). Wright et al. (Reference Wright, Turner, Andrews and Spiro1993) estimate that this calcrete represents a time gap of several hundred thousand years or longer. The calcrete is used as the correlation tie between Carham and Pease Bay, with the further tie being between the Kelso Lavas from Preston to Carham. These lithostratigraphic correlations demonstrate that the time gap represented by the calcrete lies above the early Latest Famennian and below the start of the Ballagan Formation that approximates the Devonian–Carboniferous boundary. In the Borders, the development of these calcretes represents the complex interplay between local uplift associated with volcanic doming (Millward et al. Reference Millward, Davies, Brand, Browne, Bennett, Kearsey, Sherwin and Marshall2019) and latest Famennian glacial drawdown. In East Greenland, this time interval is represented by the Stensiö Bjerg Formation (Astin et al. Reference Astin, Marshall, Blom, Berry, Vecoli, Clement and Meyer-Berthaud2010) that is equivalent to the latest Devonian Gondwana glaciation(s) that was represented in the southern hemisphere arid zone by an interval of sustained aridity.

Figure 10 Lithostratigraphic correlation between Carham, the poorly exposed new locality with early latest Famennian spores from the Whiteadder Water at Preston and Pease Bay. Carham log based on information in Carruthers et al. (Reference Carruthers, Burnett and Anderson1932). Pease Bay from our own data. The Carham Limestone is a thick single Stage 6 calcrete with a chert core that correlates with the Stage 6 calcrete at Pease Bay. The calcrete is present above the Kelso Lavas at Carham and Preston, where it is only represented by one flow (Tomkieff Reference Tomkieff1945). The mudstone with early latest Famennian spores from Preston contain volcaniclastic debris and are correlative with part of the Kelso Lavas.
Leeder (Reference Leeder1976) documented a further series of calcrete localities in the southern part of the Border Basin. Importantly, these occur as a pair of calcretes that are below the Birrenswark Volcanic Formation, which is the lateral correlative of the Kelso Volcanic Formation. If these can be dated palynologically they have the potential to provide further links to the Famennian palaeoclimate record. There are further thick calcretes in the western part of the Midland Valley, including two on Arran (Young & Caldwell Reference Young and Caldwell2009; Jutras et al. Reference Jutras, Young and Caldwell2011), which have a complex age relationship that now requires reinterpretation following reassignment of the Kinnesswood Formation to the Famennian.
6. Conclusions
This review of existing stratigraphical information coupled with new palynological data (Fig. 11) shows that:
the Upper ORS Kinnesswood Formation in Scotland is not Early Carboniferous in age as previously inferred, but unequivocally Late Devonian in age and, more specifically, in part Famennian;
the base of the Ballagan Formation approximates to the Devonian–Carboniferous boundary, with the oldest Tournaisian VI spore zone present at the base of the succession at Burnmouth;
the Ballagan Formation does not represent only the CM spore zone of late Tournaisian age but includes the entire Tournaisian, with at least the VI, HD, Cl 1 and CM (ss) spore zones present; and
calcretes can be correlated from key sections at Pease Bay and Carham, which enables the Pease Bay calcrete to be recognised as latest Devonian in age.

Figure 11 Revised stratigraphy of the Late Devonian to Early Carboniferous interval in the Borders. The new interpretation shows a much extended Ballagan Formation that extends through the entire Tournaisian. The base approximates to the base of the VI spore zone and above a hiatus or condensed horizon. The Kinnesswood Formation and other ‘ORS' formations are entirely Devonian in age. Spore zones proved to be present in the sections are shown in bold. LE and LN have not been identified. The previous age shows a more condensed Ballagan Formation equivalent to the CM spore zone and entirely late Tournaisian in age. In the Burnmouth section the Fell Sandstone Formation lies above an erosive gap and approximates the base of the Pu spore zone of Viséan age.
Records of typical Devonian fish such as Holoptychius and Bothriolepis characterise the Kinnesswood Formation and were regarded as range extensions into the Carboniferous. These are now shown to be Devonian in age, further emphasising the magnitude of the terrestrial Devonian–Carboniferous boundary mass extinction. They can be used as zone fossils to identify rock successions as Devonian in age.
This means that all of the Romer's Gap tetrapods and arthropods found in the Ballagan Formation are not from a brief upper interval of late Tournaisian age but from the entirety of the 12 My of Tournaisian time. This has significant implications for the construction of phylogenies for the early tetrapods. In particular, it brings the lower Ballagan Formation tetrapods close in time to the very different aquatic forms (Acanthostega and Ichthyostega) from East Greenland. This redistribution of Ballagan tetrapods through the entirety of the Tournaisian has filled the crucial lower part of Romer's Gap.
7. Acknowledgements
This research formed part of the TW:eed Project, led by Jenny Clack (University Museum of Zoology, Cambridge) and funded by the Natural Environment Research Council (NERC) consortium grants to the University of Southampton (NE/J021091/1), British Geological Survey (NEJ021067/1), University of Leicester (NE/J020729/1) and University of Cambridge (NE/J022713/1). Mike Browne, Tim Kearsey and Dave Millward publish with the permission of the Executive Director, BGS (NERC). Shir Akbari (Southampton) processed the palynological samples. Alex Ritchie and Vincent Dupret reinvestigated the Remigolepis from Pease Bay. Sasha Ivanov provided useful information on the synonymies of Grossilepis. Peter Osterloff (Shell) facilitated access to cuttings samples from offshore wells that cross the Devonian–Carboniferous boundary. Jenny Clack, our TW:eed Project Principal Investigator, given the nature of this published volume, graciously declined her right to be a co-author on this contribution. We acknowledge her contributions to the project from its inception and particularly her understanding of the importance of a new look at the low-diversity Ballagan Formation spore assemblages. They proved to be anything but boring.













