


1. Introduction
The tetrapod Diadectes sideropelicus (Diadectidae, Diadectomorpha) (Klembara et al. Reference Klembara, Hain, Ruta, Berman, Pierce and Henrici2020a, Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b) was first described by Cope (Reference Cope1878). Since Cope's publication several other species, as well as other genera of Diadectidae, have been described (for review see Kissel Reference Kissel2010; Klembara et al. Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b). To date, diadectids discovered have been described from North America spanning from the Upper Pennsylvanian Missourian (Case Reference Case1908) to the Lower Permian Leonardian (Cope Reference Cope1896) and the Lower Permian Artinskian of Europe (Berman et al. Reference Berman, Sumida and Martens1998, Reference Berman, Henrici, Kissel, Sumida and Martens2004).
Most of the reconstructions of the Diadectes skull are in dorsal view (van Huene Reference von Huene1913; Gregory Reference Gregory1946; Olson Reference Olson1947, Reference Olson1950; Watson Reference Watson1954; Lewis & Vaughn Reference Lewis and Vaughn1965; Berman et al. Reference Berman, Sumida and Lombard1992; Reisz Reference Reisz2006; Kissel Reference Kissel2010; Berman Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013). Four reconstructions are in lateral view (Gregory Reference Gregory1946; Olson Reference Olson1947; Watson Reference Watson1954; Reisz Reference Reisz2006), three in occipital view (Olson Reference Olson1947; Watson Reference Watson1954; Berman Reference Berman2000, Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013), and four in palatal view (Case & Williston Reference Case and Williston1912; Gregory Reference Gregory1946; Olson Reference Olson1947; Reisz Reference Reisz2006). Diadectes absitus was named and described in detail by Berman et al. (Reference Berman, Sumida and Martens1998), and it was reconstructed in dorsal and lateral views by Kissel (Reference Kissel2010), and a partial reconstruction of the palate was illustrated by Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013). Berman et al. (Reference Berman, Sumida and Lombard1992, fig. 8) compared five dorsal reconstructions of Diadectes skulls from previous authors. Those by van Huene (Reference von Huene1913) and Lewis & Vaughn (Reference Lewis and Vaughn1965) were reproduced by Berman et al. (Reference Berman, Sumida and Lombard1992) on the basis of the specimens described by van Huene (Reference von Huene1913) and Lewis & Vaughn (Reference Lewis and Vaughn1965). The skull reconstructions of Diadectes in occipital and dorsal views by Berman (Reference Berman2000, Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013) include anatomical data from D. absitus (Berman et al. Reference Berman, Sumida and Martens1998).
In comparing the reconstructions of the Diadectes skull in dorsal aspect in Figure 8 by Berman et al. (Reference Berman, Sumida and Lombard1992), we observed major differences in the anatomy and the general shape of the skulls presented by previous authors (van Huene Reference von Huene1913; Gregory Reference Gregory1946; Olson Reference Olson1947, Reference Olson1950; Lewis & Vaughn Reference Lewis and Vaughn1965). These differences involve the shape of the snout, the width of the postorbital region, and the anatomy of the posterodorsal and posterolateral portions of the skull. Although preservation of the skull material used for these reconstructions surely played a significant role in the resulting reconstructions, it is still very surprising that such major differences are observable within one genus.
Kissel (Reference Kissel2010) presented a phylogeny of diadectomorphs and in his cladograms D. absitus falls outside the genus Diadectes. He concluded that D. absitus (Berman et al. Reference Berman, Sumida and Martens1998) belongs to a new genus for which he introduced the name Silvadectes. However, because his thesis was not published, the name Silvadectes is rejected here on the grounds that it does not meet the requirements of the International Code of Zoological Nomenclature. Furthermore, the phylogeny presented by Klembara et al. (Reference Klembara, Hain, Ruta, Berman, Pierce and Henrici2020a) does not require the recognition of D. absitus as a new genus.
The aims of this paper are: (1) to present new reconstructions of the skull of D. absitus based on four specimens from central Germany; (2) to compare our reconstructions of the skull of D. absitus with those of Kissel (Reference Kissel2010), and those of Diadectes of preceding authors; (3) to describe and illustrate new cranial features in the skull of D. absitus; and (4) explicitly note errors in the previous reconstructions of the skull of Diadectes. The reconstruction of the lower jaw and stapes of the adult skull will be presented separately.
Institutional abbreviations: CM, Carnegie Museum of Natural Museum, Pittsburgh, USA; MNG, Museum der Natur, Gotha, Germany; UC, The Field Museum, Chicago, Illinois, USA; UCMP, University of California Museum of Paleontology, Berkeley, USA.
2. Material and methods
For the present study four skulls of D. absitus were used (Figs 1–3): most data are from MNG 8747 (paratype; subadult skull) and MNG 8853 (holotype; adult skull) (Berman et al. Reference Berman, Sumida and Martens1998) (Figs 1, 3). Additional anatomical data were obtained from two new specimens: MNG 14473 (skull length about 13.5 cm); and MNG 15101 (skull length about 13 cm) (Fig. 2). Skull length (measured in the median plane) of MNG 8747 is about 12.5 cm (Figs 1A, 3A, D) and that of MNG 8853 is only slightly longer at 13 cm (Fig 1B, C). The skull of MNG 8853 is more robustly constructed. The holotype specimen probably is an adult individual based on the well-developed, fully ossified postcrania, whereas the paratype (MNG 8747) exhibits a subadult stage of molarisation of its dentition, particularly in the lower jaw. It appears that the early attainment of skull size far outpaces that of the postcrania. This is also the case in the diadectomorph Tseajaia (Moss Reference Moss1972) in which the skull size (midline length) of the holotype of Tseajaia campi UCMP 59012 and that of the juvenile Tseajaia sp. CM 38033 are almost identical, yet the postcranium of the latter is much smaller and represents a much earlier stage of developmental ossification. Therefore, the holotype skull (MNG 8853) of D. absitus is considered to represent an adult specimen; the same is true for specimens MNG 14473 and MNG 15101.
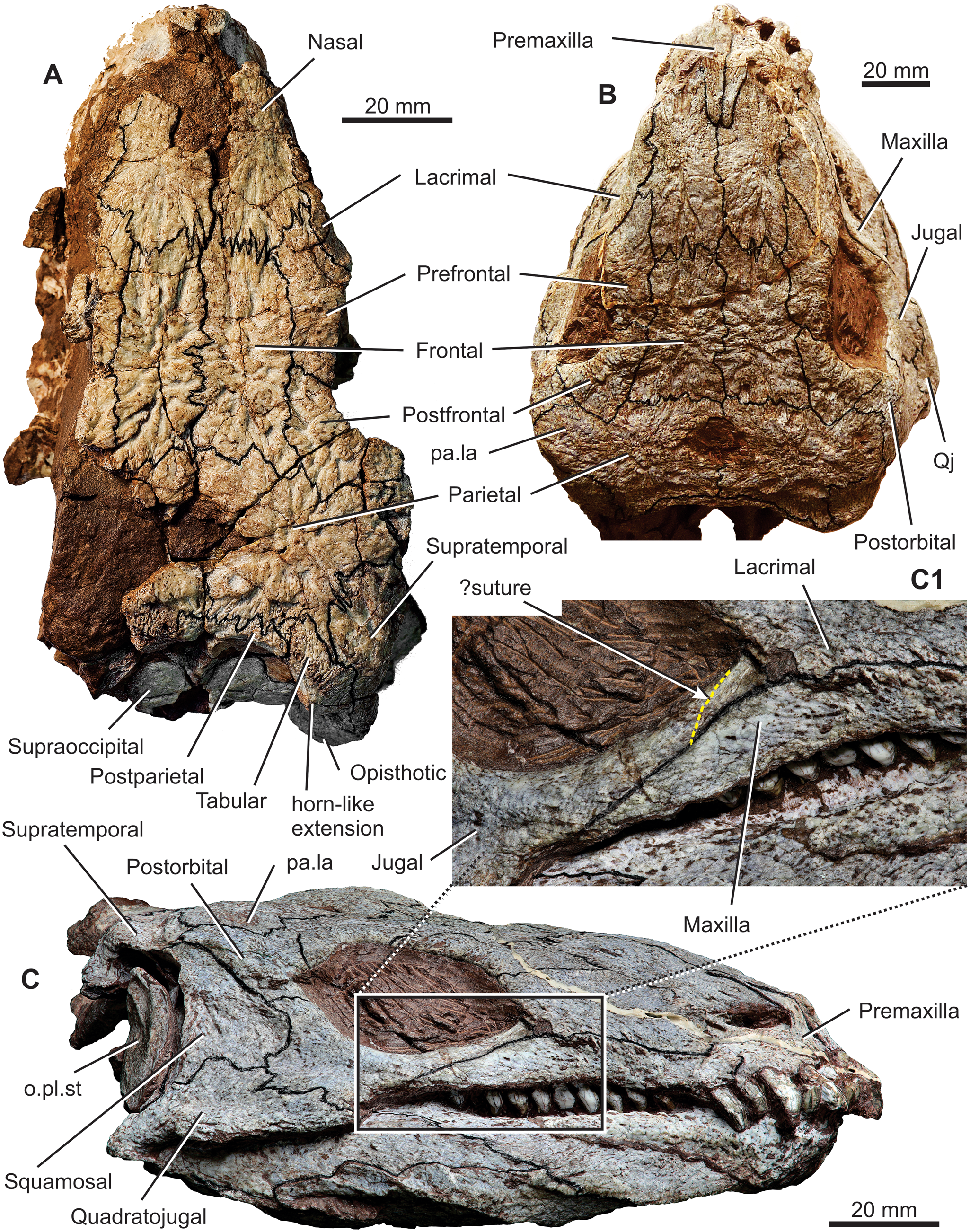
Figure 1. Diadectes absitus. (A) MNG 8747, skull in dorsal view. (B) MNG 8853, holotype skull in dorsal view. (C) MNG 8747, skull in left lateral view; (C1) close-up view of the lacrimal, jugal, and maxilla suture, with yellow dotted line indicating the hypothesised course of the lacrimal–jugal suture. Abbreviations: o.pl.st= ossified plate of stapes; pa.la= parietal lappet; Qj= quadratojual.

Figure 2. Diadectes absitus. (A) MNG 14471, skull in laterodorsal and slightly anterior view; (A1) and (A2) close-up views of the orbital portion of the skull. (B) MNG 15101, skull in dorsolateral and anterior view; (B1) and (B2) close-up views of the orbital portion of the skull.

Figure 3. Diadectes absitus. (A) MNG 8747, skull in ventral view. (B) MNG 8853, partial palate of holotype skull in ventral view. (C) Transverse computed tomography section through the pterygoid and palatine. (D) MNG 8747, skull in right lateral view. Abbreviations: Ecpt= ectopterygoid; pa.la= parietal lappet.
The skull of D. absitus is reconstructed in dorsal, lateral, posterior, and ventral views (Figs 4–7). The reconstruction of the skull roof in dorsal aspect, including the size and position of the orbits, is based mainly on MNG 8747 (Fig. 1A) and partially on MNG 8853 (Fig. 1B). The posterodorsal and occipital aspects are based on MNG 8747 (Fig. 1A) and in part on MNG 8853 (Fig. 6A). The reconstruction of the cheek in lateral aspect is based mostly on the holotype skull MNG 8853 (Fig. 1C), and partially on MNG 14471 (Fig. 2A) and MNG 15101 (Fig. 2B). The palate was reconstructed from MNG 8747; especially its right side, the parasphenoid and braincase are well preserved, although most of the ectopterygoid is missing and there is a break in the posterior portion of the palatine (Fig 3A, C). However, the right palatine and ectopterygoid are perfectly preserved in MNG 8853 (Fig. 3B; see also Berman et al. Reference Berman, Sumida and Martens1998). The marginal dentition is reconstructed mostly from MNG 8747. The proximal portion of the stapes is also perfectly preserved in MNG 8747 (Berman et al. Reference Berman, Sumida and Martens1998; Klembara et al. Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b), and the distal portion of the stapes is well preserved in MNG 8853 (Berman et al. Reference Berman, Sumida and Martens1998). To understand better the proportional interrelationships of the posterolateral and posterodorsal portions of the skull, the skull table, posterior cheek, and corresponding portion of the palate were reconstructed using plasticine models of individual bones. All bones of these skull portions were measured, modelled at two times natural size, and then joined together using metal bars.

Figure 4. Diadectes absitus. (A) Reconstruction of skull in dorsal view. (B) Line drawing of skull with coloured bones at left side.

Figure 5. Diadectes absitus. (A) Reconstruction of skull in right lateral view. (B) Line drawing of skull in right lateral view with colour-coded bones.
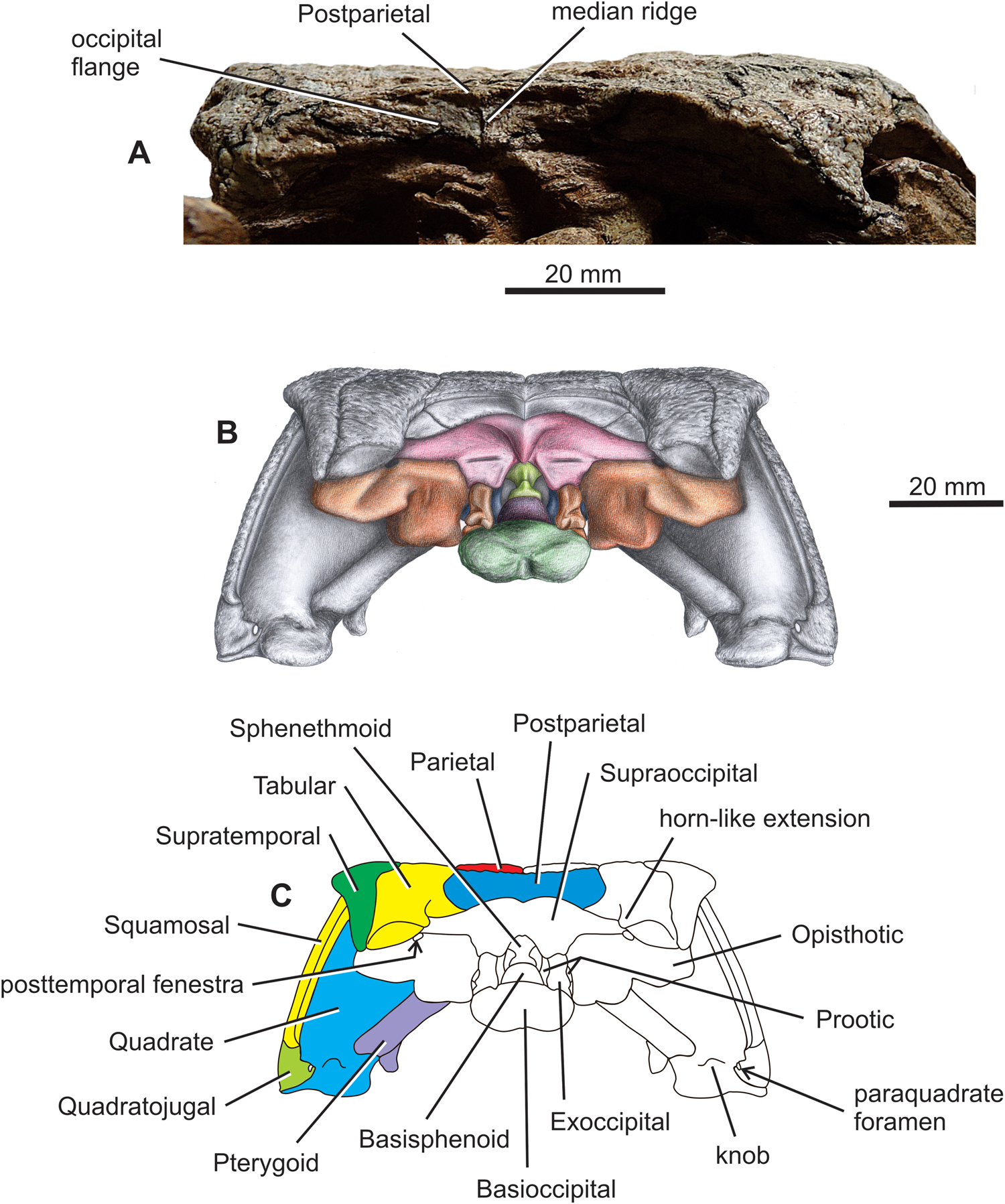
Figure 6. Diadectes absitus. (A) MNG 8853, skull in posterior view. (B) Reconstruction of skull in posterior view. (C) Line drawing of skull in posterior view with colour-coded bones on the left side.

Figure 7. Diadectes absitus. (A) Reconstruction of skull in ventral view. (B) Line drawing of skull in ventral view with colour-coded bones on the left side.
Reconstructions of the braincase, parasphenoid, and most of the stapes based on X-ray microcomputed tomography were published recently by Klembara et al. (Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b), and they were used for the reconstruction of the complete skull of D. absitus presented here. The braincase bones were colour-coded for easier identification in Klembara et al. (Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b) and similar colours are used here.
All previous reconstructions were based on one or more specimens of Diadectes, often from specimens assigned to different species. The reconstructions of the skull of Diadectes by Olson (Reference Olson1947) were based mostly on Diadectes sideropelicus and Diadectes tenuitectes. No reconstruction was ever provided that was based upon a specific species of Diadectes.
3. Description
3.1. Skull in dorsal and lateral views
The skull roof is longer than wide (Figs 1A, B, 4–7). The snout is bilaterally narrowed, reflecting the distinctly concave shape of the maxilla immediately anterior to the orbit (Figs 1B, 3A, 4, 7). The posterior margin of the skull table is embayed anteriorly, and its posterolateral corners, formed by posteriormost portions of the tabulars and supratemporals, extend posteriorly almost to the posterior margins of the opisthotics. The jaw joint lies at the level of the posterior portion of the basioccipital. The skull roof is subtriangular in dorsal view, and the sutures are distinct and mostly complicated. The sculpturing consists of high ridges and deep grooves. The centres of ossification are easily recognisable on most of the bones, with that of the parietal lying slightly medial to the parietal lappet. The parietal lappet presumably represents the intertemporal co-ossified with the parietal (Watson Reference Watson1954; Berman et al. Reference Berman, Sumida and Lombard1992). In the stem amniote seymouriamorph Discosauriscus austriacus the ossification centre of the fused parietal–intertemporal lies about halfway between the two bones (Klembara Reference Klembara, Goujet, Janvier and Poplin1995). The external naris is mediolaterally narrow with its posterior end extending slightly posterior to the premaxilla–maxilla suture. The parietal foramen is large and has an elliptical outline that is slightly wider than long. The midline suture of the parietals is shorter anterior to the parietal foramen than its length posterior to the foramen. The postparietal is unpaired in MNG 8853. In the subadult MNG 8747, a crack occurs in the median plane of the posteriormost portion of the skull table, so it is not possible to establish whether the postparietal is unpaired. The posterior margin of the ornamented surface of the postparietal extends posteriorly forming a small median process (Figs 4, 5B). The right and left portions of the unsculptured occipital flange are anteriorly concave and meet in a distinct median ridge that extends into a small but prominent process (Fig. 6).
In lateral aspect the skull roof is dorsoventrally narrowest preorbitally, but posteriorly it gradually increases in height ventrally, reaching its greatest height at the level of the jaw joint (Fig. 5). The frontal was probably slightly visible in lateral view. The premaxilla has almost a vertical position in lateral view, but its nasal ramus sharply curves ventrally extending posteriorly and slightly dorsally (cf. also Berman et al. Reference Berman, Sumida and Martens1998, fig. 4). The posterior tip of the nasal ramus lies at a level posterior to the external naris. The suborbital portion of the maxilla tapers rapidly to a point posteriorly, reaching a level slightly posterior to the mid-length of the orbit. The jugal forms most of the ventral margin of the orbit. The orbit length exceeds its depth in a lateral projection, with the maximum depth occurring slightly posterior to the orbit mid-length, which in turn lies slightly anterior to the skull mid-length, but slightly posterior to the orbit mid-length portion of the skull roof (Fig. 5).
The ventral flange of the transverse process of the pterygoid is aligned vertically with the posteroventral margin of the orbit. The orbital margin of the lacrimal is anteriorly vaulted. The anterior tip of the lacrimal process of the jugal is dorsoventrally flat (well preserved in MNG 14473 and MNG 15101) and overlaps the short, posteriorly directed jugal process of the lacrimal (Figs 2, 4, 5). The suture between the lacrimal and jugal extends in an anterodorsal to posteroventral direction, and immediately anterior to it is a wedge-shaped opening of the nasolacrimal canal. This opening is located in the posteroventral portion of the lacrimal (Figs 2A, 4, 5), with the posterior end of the canal opening directly into the orbit. The nasolacrimal canal is well preserved in MNG 14473 which opens dorsally and is divided by a longitudinal partition (Fig. 2A); this condition is often present in the seymouriamorph stem amniote Diascosauriscus (Klembara Reference Klembara1997, fig. 4-left lacrimal). The posteriormost portion of the orbit produces a small notch in the anteroventral margin of the postorbital (Figs 1B, C, 4, 5). The notch results from a posterior extension of the posterior margin of the orbit, which is accentuated by a ventrolateral extension of the postfrontal and neighbouring margin of the postorbital (Figs 1C, 5). The smooth orbital margin of the postorbital is well preserved in MNG 15101 (Fig. 2B). The postorbital portion of the skull is vertically deep, more so than the preorbital portion. The lateralmost portion of the lateral lappet of the parietal is visible in lateral view (Figs 1C, 5). Its posterior margin lies at the same level as the posterior margin of the postorbital. The suture separating the jugal, quadratojugal, and squamosal has a rather complicated pattern of deep interdigitating incisions. Posteriorly the squamosal is moderately concave, and it forms the anterior wall of the otic notch. The posterior margin of the squamosal bears a groove for the ossified plate of the stapes (Fig. 5 and Berman et al. Reference Berman, Sumida and Martens1998, figs. 5, 6).
The occipital portion of the skull roof is well preserved and exposed occipitally, as well as dorsally in the subadult MNG 7478 (Fig. 1A). However, the occiput of the larger holotype MNG 8853 (Berman et al. Reference Berman, Sumida and Martens1998) and those of MNG 14473 and MNG 15101 are not well exposed (Figs 1B, 6A). Three bones of the neural endocranium visible in dorsal aspect include the supraoccipital, opisthotic, and basioccipital (Figs 1A, 4). The supraoccipital is a wide median element, and its exposed lateral portion extends lateral to the level of the lateral extremities of the postparietal (Fig. 4). The anteromedial portion of the opisthotic is visible in dorsal view, and it contacts the supraoccipital medially in an anterolaterally–posteromedially oblique suture. The medial portion of the paroccipital process of the opisthotic is well-exposed dorsally. The posttemporal fenestra is extremely small and lies immediately ventral to the tabular horn-like extension of the ornamented surface of the tabular (Figs 1A, 4, 6B, C and see below). Only the posteriormost portion of the occipital condyle of the basioccipital is visible in dorsal view (Fig. 4).
In lateral view the paroccipital process of the opisthotic extends slightly more posteriorly than the posteroventral corner of the supratemporal (Fig. 5). A substantial portion of the posterolateral portion of the opisthotic is exposed in lateral view. In addition to the opisthotic, the posterolateral portions of the basioccipital, prootic, and supraoccipital are also exposed in lateral view (Fig. 5). A substantial portion of the sphenethmoid and the entire cultriform process of the parasphenoid are visible through the orbit (Fig. 5). The dorsalmost portions of the sphenethmoid are mediolaterally broadened and abut the ventral surfaces of the frontals (cf. Klembara et al. Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b).
3.2. Occiput
In posterior view the mid-region of the skull roof is slightly concave (Figs 4, 6B, C). The occipital portion of the skull roof is arched posteroventrally at angle of about an 45°, thus the posterior walls of the postparietal(s), tabulars, and supratemporals face posterodorsally (in the subadult MNG 8747, Figs 1A, 3D, 4–6). In the large specimens, the occiput is much steeper relative to the vertical plane (Fig 1B, C; Berman et al. Reference Berman, Sumida and Martens1998; and specimens MNG 14473 and MNG 15101). The posteroventral ends of the tabular and supratemporal substantially overlap the paroccipital process of the opisthotic (Figs 1A, 4–6). The smooth occipital flanges of the postparietal are fully exposed in occipital view (Fig. 6). In contrast, only about the medial half of the tabular occipital flange is exposed in posterior view (Fig. 6). This is a consequence of the posteromedial flexion of the posterior corner of the skull table formed by the tabular and supratemporal. About at the mid-length of the tabular, the posteromedial portion of its ornamented surface protrudes into a small but distinct horn-like extension which is oriented posteromedially (Figs 1A, 4, 6B, C). The posttemporal fenestra is located immediately ventral to this horn-like extension (Fig. 5B, C and see fig. 5B in Klembara et al. Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b). Smooth occipital flanges of the postparietal and tabular overlap the anterodorsal portions of the supraoccipital and posterodorsal portions of the opisthotic, respectively (Figs 4, 6B, C). Substantial portions of the opisthotics, exoccipitals, and basioccipital are exposed in posterior view (Fig. 6B, C).
The cheeks are inclined at about 60° to the plane of the skull roof (Fig. 6). The midline height of the skull is slightly greater than two-thirds of the absolute height of the skull in posterior aspect (Fig. 6B, C). The quadrate is a robust element contacted by the quadrate ramus of the pterygoid medially, and by the quadratojugal and squamosal laterally (Figs 3A, 5, 6B, C).
3.3. Palate
The palate narrows anteriorly, and from a point between the anterior and middle one-thirds it gradually broadens posteriorly (Figs 3A, 7). The triradiate vomer (Berman et al. Reference Berman, Sumida and Martens1998) consists of a dorsal premaxillary process, posterolateral wing, and posteromedial process bearing a row of eight teeth (in MNG 8747). There is a narrow anteroposterior slit-like opening between the premaxillary processes of the vomer (Fig. 7). The posterior (vomerine) processes of the premaxillae are dorsal to the vomers, and are partially visible in the slit between the vomers (Figs 3A, 7). As pointed out by Berman et al. (Reference Berman, Sumida and Martens1998) there is no contact between the posterolateral wing of the vomer and the palatine. Thus, the anterolateral margin of the palatal process of the pterygoid forms the posteromedial margin of the endochoanal fenestra. Further, a typical feature of the palate of Diadectes is the presence of the ventral secondary palatal shelf of the palatine, which in the distance from about its anterior and middle thirds of its length obscures from ventral view the suture between the pterygoid and dorsal portion of the palatine (Figs 3C, 7). The anterior end of the interpterygoid vacuity lies at the level of the posterior end of the maxilla. The adductor fossae are large, indicating the presence of a conspicuous adductor musculature.
Most of the posterior half of the braincase and the nearly complete parasphenoid are exposed in ventral view (Fig. 7). The posterior wall of the quadrate lies at the level of the posterior margin of the basioccipital.
4. Comparisons
4.1. Comparisons with the reconstructions of the skull of D. absitus of Kissel (Reference Kissel2010)
Kissel (Reference Kissel2010) presented dorsal and lateral reconstructions of the skull of D. absitus. Our reconstructions of the skull of D. absitus differ from those of Kissel (Reference Kissel2010) in the following points:
1) Kissel (Reference Kissel2010, fig. 34B) reconstructed the maxilla in D. absitus as slightly convex in dorsal view, giving it a substantially wider shape of the preorbital portion of the skull. The shape of the maxilla in dorsal view as presented by Kissel is evidently in strong contrast with the morphology of the maxilla in all known skulls of D. absitus. Instead, the maxillae in D. absitus are distinctly concave, and thus the snout is narrowed (Figs 1B, 3A, 4, 7).
2) In his reconstruction of the skull in dorsal view the external naris is not visible (Kissel Reference Kissel2010, fig. 34B). This interpretation is highly improbable and has never appeared in any of the skull reconstructions of Diadectes by previous authors (see below and Berman et al. Reference Berman, Sumida and Lombard1992).
3) In Kissel's (Reference Kissel2010, fig. 34B) reconstruction of the skull in dorsal view it is slightly concave along the jugal length. As we demonstrate here, the skull is convex in this area (Fig. 4A).
4) Kissel (Reference Kissel2010, fig. 34B) reconstructed the parietal lappets as being posterolaterally oriented, but they clearly are oriented anterolaterally in D. absitus (Figs 1A, B, 4).
5) In his reconstruction of the skull in lateral view the snout is formed by the premaxilla, which has an arch-like outline (Kissel Reference Kissel2010, fig. 34A). This is contrasted by a distinct obtuse angle between the anterior portion and the nasal process of the premaxilla (Figs 1B, C, 5).
6) In Kissel's (Reference Kissel2010, fig. 34) lateral view of the skull, the tip of the anterior process of the jugal is dorsoventrally broad, lies anterior to the anterior margin of the orbit, and the jugal process of the lacrimal is absent. However, the anterior end of the lacrimal process of the jugal does not quite reach the anterior margin of the orbit, and the lacrimal has a short, but distinct jugal process. There is an anterodorsally to posteroventrally oblique suture between the anterior tip of the lacrimal process of the jugal and the jugal process of the lacrimal (Figs 1C, 2, 4, 5).
7) In dorsal and lateral views, the posteriormost portion of the supratemporal lies distinctly posterior to the posterior tip of the quadratojugal (Figs 4, 5). In the reconstruction by Kissel (Reference Kissel2010, figs. 34A, B), the posterior tips of both bones lie almost in the same level in lateral view; however, in dorsal view the posterior tip of the quadratojugal lies much more posteriorly relative to that of the supratemporal.
8) In the reconstruction by Kissel (Reference Kissel2010, fig. 34A), the posterior (jaw joint) portion of the quadratojugal is not present; however, the posterior portion of the quadratojugal is visible in dorsal and lateral views (Figs 4, 5).
9) The reconstruction of the posterolateral portion of the skull (Kissel Reference Kissel2010, figs. 34A, B) is extremely simplified, and the reconstructions of the opisthotic, prootic, and basioccipital are not included; however, these three endocranial bones are visible in lateral view (Fig. 5). Thus, no comparisons are possible.
10) In several places the sutures between the skull roof bones are much more complicated than shown by Kissel (Reference Kissel2010, fig. 34). For example, they are highly interdigitating between the lacrimal and nasal and the postorbital and jugal (Figs 4, 5).
11) The anteriormost teeth are procumbent, as correctly shown in lateral view, and extend anteriorly beyond the anterior margin of the snout (Kissel Reference Kissel2010, fig. 34A). However, in dorsal view the teeth are not reconstructed as extending anteriorly beyond the snout (Kissel Reference Kissel2010, fig. 34B) (see Figs 4, 5 here).
12) In his lateral view of the skull, Kissel (Reference Kissel2010, fig. 34A) reconstructs the maxillary teeth as much smaller than the premaxillary teeth; however, the first maxillary tooth is well preserved in MNG 8853 and is of similar size to the last premaxillary tooth (Figs 1C, 5). Farther from the first maxillary tooth all other maxillary teeth become gradually lower posteriorly (Fig. 5).
4.2. Comparisons with the previous reconstructions of the skull of Diadectes
4.2.1. Reconstructions in dorsal view
Previous reconstructions of the skull of Diadectes in dorsal aspect show the common features, which are listed below with their revisions. Previous reconstructions: (1) do not exhibit the narrowing of the skull immediately anterior to the orbits; (2) the lateral portion of the premaxilla, maxilla, and jugal are not visible in dorsal view; and (3) the orbits are only narrow anteroposteriorly elongated slits. Yet, in the reconstruction of the Diadectes skull presented by Gregory (Reference Gregory1946) all the above features are present, as in D. absitus (Fig. 4). Furthermore, only Gregory (Reference Gregory1946) and Watson (Reference Watson1954) reconstructed the skull of Diadectes with a slight narrowing of the preorbital portion of the skull. This is similar to our reconstruction. Thus, in most previous reconstructions the preorbital region is always broader relative to the postorbital region, as seen in comparison with the reconstruction of the skull of D. absitus presented here. The exception is the skull of Diadectes molaris figured in dorsal view (van Huene Reference von Huene1913, fig. 15) in which the maxilla–jugal suture is visible in dorsal view, as in our reconstruction (Fig. 4). Only in the reconstruction by Gregory (Reference Gregory1946) does the premaxilla have a long, posterodorsally extended nasal process. A similar process is present in AMNH 4352 (D. molaris of van Huene Reference von Huene1913), Diadectes sanmiguelensis of Lewis & Vaughn (Reference Lewis and Vaughn1965), and Kissel (Reference Kissel2010). The same process is well developed in the skulls of D. absitus (Berman et al. Reference Berman, Sumida and Martens1998; and Figs 1B, C, 2, 4, 5 here).
In the reconstruction of the posterior half of the skull, only those described by Gregory (Reference Gregory1946), Olson (Reference Olson1947, Reference Olson1950), Berman et al. (Reference Berman, Sumida and Lombard1992), and Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013) is the quadratojugal visible in dorsal view. However, only in the reconstruction of Gregory (Reference Gregory1946) is the entire length of the quadratojugal fully exposed in dorsal view, as in D. absitus (Figs 1B, 4). As already noted above, there is no separate intertemporal ossification in Diadectes; rather, it is undoubtedly incorporated into the parietal to form the lateral lappet (Berman et al. Reference Berman, Sumida and Lombard1992). The postparietal is paired in the reconstructions of van Huene (Reference von Huene1913), Gregory (Reference Gregory1946) and a subadult specimen of D. sanmiguelensis of Lewis & Vaughn (Reference Lewis and Vaughn1965). The postparietal is absent in the reconstruction of Olson (Reference Olson1947), but it was identified and reconstructed in his later publication (Olson Reference Olson1950). The shape of the postparietal strongly varies in all skull reconstructions of Diadectes presented by the previous authors (van Huene Reference von Huene1913; Gregory Reference Gregory1946; Olson Reference Olson1947, Reference Olson1950; Watson Reference Watson1954; Lewis & Vaughn Reference Lewis and Vaughn1965; Berman et al. Reference Berman, Sumida and Lombard1992; Kissel Reference Kissel2010; Berman Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013). The unpaired postparietal in the reconstructions of Olson (Reference Olson1950) and Watson (Reference Watson1954) has a long, posterior median extension reminiscent of the sagittal crest of the supraoccipital (Figs 4A, 6B, C). As described above, in D. absitus the postparietal is an unpaired, elongated plate in the large holotype (Fig. 6A). The dorsal surface of the postparietal is ornamented, and an unornamented occipital flange is inclined posteroventrally (Figs 1A, B, 4, 6). However, the median process extending posteroventrally from the ornamented surfaces is never as large as that in the Diadectes skull figured by Olson (Reference Olson1950) and Watson (Reference Watson1954). The sizes, shapes, and interrelationships of postorbital, squamosal, tabular, and supratemporal differ among authors (see the reconstructions of the skulls of Diadectes in dorsal view in Berman et al. Reference Berman, Sumida and Lombard1992, fig. 8). The postorbital is large in dorsal view in the reconstructions by van Huene (Reference von Huene1913) and Gregory (Reference Gregory1946), relative to the very small postorbital in the reconstructions by Olson (Reference Olson1947, Reference Olson1950). The supratemporal is absent in the reconstruction by Gregory (Reference Gregory1946), and the area corresponding to it is instead occupied by a large squamosal that contacts the parietal immediately posterior to the postorbital. In the reconstruction by Lewis & Vaughn (Reference Lewis and Vaughn1965), the supratemporal overlaps the tabular at about its mid-length. This position of the supratemporal is incorrect; the supratemporal lies immediately lateral to the tabular (Berman Reference Berman2000 and Figs 1A, C, 4–6 here). In the reconstruction by Gregory (Reference Gregory1946), the quadratojugal–postorbital contact separates the squamosal from the jugal. In contrast with our reconstruction (Fig. 4) in none of the reconstructions of the Diadectes skull, is the squamosal–jugal suture visible in dorsal view.
In none of the Diadectes skull reconstructions is the supraoccipital–opisthotic suture marked. This is because in the adult skulls of Diadectes these two bones are mostly fused together. However, except for Berman et al. (Reference Berman, Sumida and Lombard1992), in reconstructions produced by all other authors (van Huene Reference von Huene1913; Gregory Reference Gregory1946; Olson Reference Olson1947, Reference Olson1950; Watson Reference Watson1954; Lewis & Vaughn Reference Lewis and Vaughn1965), the fused opisthotic–supraoccipital complex is designated as the supraoccipital. In our reconstruction, the supraoccipital–opisthotic suture runs mediolaterally and is situated medially to the posterior portion of the tabular (Figs 4, 5). In contrast to the previous reconstructions, the posteriormost portion of the occipital condyle of the basioccipital is slightly visible in dorsal view (Fig. 4).
4.2.2. Reconstructions in lateral view
All previous reconstructions of the snout of Diadectes show the premaxilla as having a gradual arch-like anterodorsal outline. However, the premaxilla of D. absitus has an almost vertical anterior portion and a sharply posterodorsally flexed nasal process (Berman et al. Reference Berman, Sumida and Martens1998). The premaxilla is preserved in an undistorted position in the subadult MNG 8748 (Fig. 3D) and adult MNG 8853 (Fig. 1C) specimens. This is true also of other diadectomorph species, such as D. sideropelicus (Kissel Reference Kissel2010, specimen UC 675), D. sanmiguelensis (Lewis & Vaughn Reference Lewis and Vaughn1965), Orobates pabsti (Berman et al. Reference Berman, Henrici, Kissel, Sumida and Martens2004), Limnoscelis paludis (Berman et al. Reference Berman, Reisz and Scott2010), and Tseajaia campi (Moss Reference Moss1972).
In most previous reconstructions of the skull of Diadectes the posteroventrally extending jugal process of the lacrimal and the lacrimal process of the jugal are present, but are dorsoventrally high on the skull and meet along an almost vertical suture. In the reconstructions of Olson (Reference Olson1947), Watson (Reference Watson1954), Reisz (Reference Reisz2006), and Kissel (Reference Kissel2010), the lacrimal–jugal suture extends anteroventrally–posterodorsally. In contrast, the suture in D. sanmiguelensis runs anterodorsally–posteroventrally (Kissel Reference Kissel2010); however, the reconstruction of the skull of Diadectes by Gregory (Reference Gregory1946) shows a long lacrimal process of the jugal reaching the anterior margin of the orbit. The anterior tip of the jugal is bifurcated and a short process of the lacrimal fits between these small processes (Gregory Reference Gregory1946, fig. 6a). Up to now, the jugal–lacrimal interrelationship has remained unclear in D. absitus. This portion of the skull is not preserved in the subadult specimen, and in the holotype specimen the sutural pattern is unclear (Berman et al. Reference Berman, Sumida and Martens1998). On the left side of the skull of MNG 8853, the lacrimal process of the jugal appears to be shorter than the right side; however, on the right side of the skull the lacrimal process appears to be anteriorly elongated, reaching the anterior margin of the orbit, but its suture with the lacrimal is not clearly recognisable (Fig. 1C). On the D. absitus skulls, MNG 14473 and MNG 15101, the lacrimal process of the jugal does not reach the anterior margin of the orbit (as it appears to do so on the left side of the holotype skull; Berman et al. Reference Berman, Sumida and Martens1998) and its anterior tip is dorsoventrally flat (Fig. 2). Furthermore, the jugal process of the lacrimal is present and extends below the anterior tip of the lacrimal process of the jugal (Figs 2, 4, 5).
In all previous reconstructions of Diadectes skulls, the posterior margin of the orbit is rounded. In D. absitus, the posterior wall of the orbit extends at about its mid-height into a posteriorly oriented slight extension in the ventral portion of the postorbital (Figs 1C, 2B). In all previous reconstructions of the adult skull of Diadectes, the anterior end of the quadratojugal does not reach the level of the posterior margin of the orbit, but it does in a subadult specimen of D. sanmiguelensis (Lewis & Vaughn Reference Lewis and Vaughn1965; Kissel Reference Kissel2010). In contrast, in D. absitus the anterior portion of the quadratojugal extends anteriorly ventral to the posterior portion of the orbit in the holotype (Figs 1C, 5).
4.2.3. Reconstructions in occipital view
Olson (Reference Olson1947) and Watson (Reference Watson1954) reconstructed the skull of Diadectes in occipital view. The reconstructions of the skull in occipital view of Diadectes by Berman (Reference Berman2000, Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013) include the morphological features present in D. absitus (Berman et al. Reference Berman, Sumida and Martens1998). The occiput of Diadectes, as reconstructed by Watson (Reference Watson1954), is more similar to our reconstruction than to that of Olson (Reference Olson1947). The angle between the cheek and skull roof is very similar in Watson's (Reference Watson1954) and our reconstruction, but this angle is smaller in the reconstructions of Olson (Reference Olson1947), Berman et al. (Reference Berman, Sumida and Martens1998), and Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013). There is a deep notch of triangular shape in the median portion of the skull roof in the occipital reconstruction by Olson (Reference Olson1947). Such a deep notch is not present in the corresponding portion of the skull in either the reconstruction by Watson (Reference Watson1954), Berman et al. (Reference Berman, Sumida and Martens1998), and Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013), or our reconstruction. The reconstruction by Watson (Reference Watson1954) shows a suture between the supraoccipital and supposed smooth occipital flanges of the postparietals and tabulars, and our reconstruction has this suture in a similar position (Fig. 6B, C). In Watson's (Reference Watson1954) reconstruction of the occiput a short, distinct split separates the posterior median process of the ornamented surface of the postparietal, probably indicating an originally paired postparietal. In most previous reconstructions, there is a distinct median elevation between the paired portions of the smooth postparietal occipital flanges (Olson Reference Olson1947; Watson Reference Watson1954). In contrast, in D. absitus there is a narrow sharp median ridge between the smooth occipital flanges of the postparietal (Figs 4A, 6B). The supratemporal has a horn-like posteroventral extension that reaches the posteroventral margin of the tabular (Berman et al. Reference Berman, Sumida and Martens1998; Figs 1A, 5B, C here). In contrast, the supratemporal in the reconstructions by Olson (Reference Olson1947) and Watson (Reference Watson1954) is shorter than the tabular and does not reach the posteroventral margin of the tabular. In the reconstruction by Olson (Reference Olson1947) the dorsal ends of the exoccipitals are fused in the medial plane. In contrast, in our reconstruction, like those of Watson (Reference Watson1954), Berman et al. (Reference Berman, Sumida and Martens1998), and Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013), the dorsal portions of the exoccipitals do not meet in the midline. The medial portion of the quadrate is much more exposed in occipital view in our reconstruction than it is in the reconstructions by Olson (Reference Olson1947), Watson (Reference Watson1954), Berman et al. (Reference Berman, Sumida and Martens1998), and Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013). In our reconstruction (Fig. 6B, C) and that of Watson's (Reference Watson1954), the quadrate ramus of the pterygoid is much more exposed in posterior view. In the reconstructions of Olson (Reference Olson1947), Berman et al. (Reference Berman, Sumida and Martens1998), and Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013), the quadrate ramus of the pterygoid forms a dorsoventrally narrow sheet attached to the ventromedial portion of the quadrate.
4.2.4. Reconstructions in palatal view
Case & Williston (Reference Case and Williston1912), Gregory (Reference Gregory1946), Olson (Reference Olson1947), and Reisz (Reference Reisz2006) illustrated the reconstructions of the skull of Diadectes in palatal view. A partial reconstruction of the right palate was illustrated by Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013). Reisz's (Reference Reisz2006) reconstruction mostly follows Olson's (Reference Olson1947) reconstruction, but several amendments were added. These modifications regard mostly the morphology of the vomer and posterior portion of the parasphenoid. According to their reconstructions of the anterior portion of the palate the posterior (vomerine) processes of the premaxillae are exposed in ventral view. Yet according to Case & Williston (Reference Case and Williston1912) and Gregory (Reference Gregory1946) the anterior processes of the vomers completely underlie the posterior processes of the premaxillae. In D. absitus the posterior processes of the premaxillae are almost completely covered ventrally by the anterior processes of vomers except for a narrow median fissure present between these processes (Berman et al. Reference Berman, Sumida and Martens1998, Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013; Figures 3A, 7 here). In D. absitus the vomer has a well-developed posterolateral process that contacts the anterior margin of the palatal ramus of the pterygoid (Berman et al. Reference Berman, Sumida and Martens1998; Figures 3A, 7 here). The process is not shown in the reconstructions by Case & Williston (Reference Case and Williston1912), Gregory (Reference Gregory1946), and Olson (Reference Olson1947); however, it is illustrated in the reconstruction of the palate of Diadectes by Reisz (Reference Reisz2006). Olson (Reference Olson1947) described a rounded fenestra bordered by the ectopterygoid laterally, palatine anteromedially, and pterygoid posteromedially. Such fenestra is absent in D. absitus (Berman et al. Reference Berman, Sumida and Martens1998); however, there is a distinct excavation limited by a ridge of the ectopterygoid laterally and distinct ridges of the anterior margin of the transverse process of the pterygoid and posterior portion of the palatine (Berman et al. Reference Berman, Sumida and Martens1998 and Figs 3A, B, 7 here). The space limited by these ridges continues freely anteriorly (Fig 3A, B). The ectopterygoid has a short anteroposteriorly directed suture with the posteriormost portion of the palatal ramus of the pterygoid (Figs 3B, 7). In no previous reconstructions of the palate of Diadectes has this feature been documented. A typical feature of the pterygoid of D. absitus is that the anterior and posterior margins of the transverse process of the pterygoid are parallel to one another (Figs 3A, B, 7). In addition, the angle between the posterior margin of the transverse process of the pterygoid and the lateral margin of the quadrate ramus of the pterygoid is acute (Figs 3A, B, 7). There is a deep U-shaped notch between the transverse process and quadrate ramus of the pterygoid in D. absitus (Berman et al. Reference Berman, Sumida and Martens1998; Figs 3A, B, 7 here), similar to that in Limnoscelis paludis (Berman et al. Reference Berman, Reisz and Scott2010). In all other species of Diadectes (e.g., Olson Reference Olson1947; Reisz Reference Reisz2006) the posterior margin of the transverse process is short and forms an obtuse angle with the medial margin of the quadrate ramus. The anteroventral margin of the transverse process bears a distinct and sharp ridge (Figs 3B, 7), which has not been previously described in any other species of Diadectes.
The morphology of the posteriormost margin of the parasphenoid as presented by Olson (Reference Olson1947) is completely different from that presented by Klembara et al. (Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b) and here (Figs 3A, 7); however, the reconstruction of the posterior margin of the parasphenoid as described by Reisz (Reference Reisz2006) is basically the same as that presented here. It consists of two posterolateral processes, two sagittal processes, and a median process; the posterolateral processes are the most robust and the longest (Figs 3A, 7).
5. Discussion and conclusion
Since the first description of the skull of Diadectes (Cope Reference Cope1878) many additional specimens have become available. Despite many being well-preserved, important differences still exist between their reconstructions. The latest reconstructions of the Diadectes skull are those in dorsal and posterior views by Berman (Reference Berman, Lucas, DiMichele, Barrick, Schneider and Spielmann2013), which are based on Diadectes sp. CM 25741 (Berman et al. Reference Berman, Sumida and Lombard1992) and D. absitus MNG 8747 and MNG 8853 (Berman et al. Reference Berman, Sumida and Martens1998). Thus, in these latest reconstructions the morphology of the tabular and supratemporal are similar to those presented here. Nevertheless, as determined above, there are many morphological and proportional differences between them (van Huene Reference von Huene1913; Gregory Reference Gregory1946, Olson Reference Olson1947, Reference Olson1950; Watson Reference Watson1954; Lewis & Vaughn Reference Lewis and Vaughn1965; Reisz Reference Reisz2006) and those of D. absitus presented here. This is probably due not only to varying degrees of preservation of the Diadectes skulls used but also reflects that previous reconstructions are based on the morphologies of two or more species. Until now, there have been no reconstruction based on a single species of the genus Diadectes.
We present here for the first time the reconstructions of the skull of one of several species of Diadectes, D. absitus, from the early Permian of central Germany (Berman et al. Reference Berman, Sumida and Martens1998). The reconstructions are in dorsal, lateral, ventral, and posterior views and include the braincase bones (described in detail by Klembara et al. Reference Klembara, Hain, Čerňanský, Berman and Henrici2020b). In addition, numerous inconsistencies between the actual anatomy of the fossils and previous reconstructions of the skull in Diadectes are pointed out and amended.
6. Acknowledgements
We thank Dr T. Hübner (Museum der Natur, Gotha, Germany) for the permission to publish the photographs of the Diadectes absitus skull. The photographs in Figs 1C and 2 were made by Mr Stephan Brauner (Museum der Natur, Gotha, Germany). The drawings of the surface of the reconstructions (Figs 4A, 5A, 6B, 7A) were executed by Ivana Koubová (Bratislava). We thank Dr M. Ruta (University of Lincoln, UK) and one anonymous reviewer for their helpful reviews. This project was supported by the Scientific Grant Agency of the Ministry of Education of the Slovak Republic and the Slovak Academy of Sciences, Grant Number 1/0228/19 (to J.K.) and by the Agency of the Ministry of Education of the Slovak Republic and Slovak Academy of Sciences, Grant/Award Number: APVV-14-0719 (to M.H.).







