Although most Cenozoic crocodyliforms closely resemble their living relatives, the ziphodont forms do not. Strictly speaking, the term “ziphodont” refers to the possession of labiolingually compressed serrated teeth (Fig. 1a; Langston Reference Langston1975), but the term has come to include a suite of derived features often found in crocodyliforms with ziphodont dentition, including a mediolaterally compressed and dorsoventrally deep (altirostral) snout (Fig. 1c). At least some had blunt, hoof-like unguals (Fig. 1b), and they are sometimes seen as an archosaurian attempt to retain the terrestrial predator ecological roles cleared by the disappearance of non-avian theropods.

Figure 1 Morphological features characteristic of ziphodont eusuchians: (A) labiolingually compressed, serrated teeth (AMNH 2090, Boverisuchus vorax); (B) hooflike unguals (GM Tr I Leo, B. magnifrons, ventral view); (C) altirostral snout (FMNH PR399, B. vorax, left lateral view). Scale bars=1 cm (A, B); 10 cm (C).
One such group is found in the Palaeogene of North America and Eurasia. These are referred to Pristichampsinae Kuhn, Reference Kuhn1968 (Rossmann Reference Rossmann1998; Brochu Reference Brochu2003) and, based on phylogenetic analyses, are part of a lineage basal within or closely related to Crocodylia (Salisbury & Willis Reference Salisbury and Willis1996; Pol et al. Reference Pol, Turner and Norell2009; Brochu et al. Reference Brochu, Njau, Blumenschine and Densmore2010). Although not known prior to the Palaeocene, the phylogenetic placement of the group implies a minimum time of origin in the Campanian.
Its phylogenetic position makes this group relevant to the controversy over higher-level crocodylian relationships. Morphological evidence draws the Indian gharial (Gavialis gangeticus) and its extinct relatives to the base of Crocodylia and supports a crocodylid affinity for the false gharial (Tomistoma schlegelii), but molecular data link Gavialis and Tomistoma together and root trees on the alligatorids (e.g., Densmore Reference Densmore, Hecht, Wallace and Prance1983; Norell Reference Norell1989; Brochu Reference Brochu2003; Gatesy et al. Reference Gatesy, Amato, Norell, DeSalle and Hayashi2003; Piras et al. Reference Piras, Colangelo, Adams, Buscalioni, Cubo, Kotsakis, Meloro and Raia2010; Oaks Reference Oaks2011). Improved sampling from early-diverging groups such as this might help determine if morphological data are being misled by incorrect character state polarity optimisations.
At present, only one Laurasian species – Pristichampsus vorax (Troxell Reference Troxell1925) – has been included in phylogenetic analyses (Salisbury & Willis Reference Salisbury and Willis1996; Brochu Reference Brochu1997, Reference Brochu1999, Reference Brochu2007; Delfino et al. Reference Delfino, Codrea, Folie, Dica, Godefroit and Smith2008a; Pol et al. Reference Pol, Turner and Norell2009; Martin Reference Martin2010). This is one of the youngest and most morphologically derived of these forms. Expansion of our sample, especially from earlier in the history of the group, could limit the impact of phylogenetic overprinting of plesiomorphic traits and improve our ancestral state reconstructions at the base of Crocodylia.
To do this, we face several challenges. The first is a clear understanding of species-level diversity. Some argue that all known Eocene ziphodont eusuchians, including P. vorax, are referable to a single species, Pristichampsus rollinati (Gray Reference Gray1831), and that differences between samples reflect intraspecific variation (Rauhe & Rossmann Reference Rauhe and Rossmann1995; Rossmann Reference Rossmann1998), but not all of the differences are within observed levels of variation among living crocodylian species. Indeed, some currently recognised species might include multiple diagnosable units.
The preservation of some holotypes complicates our assessment. Whether P. rollinati can be diagnosed is doubtful (Langston Reference Langston1975) – the holotype is limited to teeth, postcranial remains, a partial dentary and something that is part of either a snout or a mandible (Fig. 2). This renders the term Pristichampsinae itself systematically problematic, in spite of its prominence in the literature.
Figure 2 Pristichampsus rollinati, material referred by Cuvier (Reference Cuvier1824): (A) MNHN AG 8, partial dentary, dorsal view; (B–C) TMM 40741-8, cast of MNHN AG 7, fragment of skull or lower jaw in lateral (B) and medial (C) views. Scale bar=1 cm. Abbreviation: ?emf=possible external mandibular fenestra.
This paper represents an initial reevaluation of the phylogenetic relationships of these forms based on first-hand examination of specimens from the United States, France, Germany, Italy and China. It also provides a revised taxonomy for some of these species. Hopefully, it will help guide further research the origins, biogeographic history and extinction of ziphodont crocodyliforms during the Age of Mammals.
Wann Langston coined the term “ziphodont,” and his 1975 paper on Pristichampsus vorax stands as a model of descriptive morphology and taxonomic review. There isn't much hyperbole involved when I say I owe Dr. Langston my career – I may have absorbed my phylogenetic philosophy elsewhere, but much of what I am as a scientist was learned from him. Whatever good comes from this paper is dedicated to Dr. Langston. The shortcomings should be laid right on my feet.
Institutional abbreviations. AMNH, American Museum of Natural History, New York, NY; Field Museum of Natural History, Chicago, Illinois, USA; GM, Geiseltalmuseum, Martin Luther Universität, Halle, Germany; HLMD, Hessisches Landesmuseum, Darmstadt, Germany; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; LACM, Museum of Natural History of Los Angeles County, Los Angeles, CA; MGPD, Museo di Geologia e Paleontologia, Università di Padova, Padua, Italy; MNHN, Museum National d'Histoire Naturelle, Paris, France; MNHT, Musée d'Histoire Naturelle de Toulouse, Toulouse, France; NHMUK, Natural History Museum, London; PU, Princeton University, Princeton, NJ (collections now at YPM); TMM, Texas Memorial Museum, Austin, TX; UCM, University of Colorado Museum of Natural History, Boulder, CO; UCMP, University of California Museum of Paleontology, Berkeley, CA; USNM, United States National Museum of Natural History, Washington, DC; YPM, Peabody Museum of Natural History, Yale University, New Haven, CT.
1. Taxonomic review and phylogenetic sampling
1.1. Pristichampsus rollinati (Gray Reference Gray1831)
Pristichampsus rollinati (Gray Reference Gray1831) is based on the “Crocodile des marnières d'Argenton” of Cuvier (Reference Cuvier1824). It consists of some teeth and bone fragments from the middle Eocene of Argenton in western France. Gray (Reference Gray1831) established Crocodilus rollinati from one of the teeth (MNHN AG 3) and vertebrae (MNHN AG 4) figured by Cuvier (Reference Cuvier1824). This was transferred to Pristichampsus by Gervais (Reference Gervais1853).
Cuvier (Reference Cuvier1824) was struck by the flattened, serrated teeth associated with the Crocodile d'Argenton. They are sometimes compared with those of theropod dinosaurs (Langston Reference Langston1975), but nonavian theropods being poorly understood in 1824, Cuvier compared them with the teeth of a monitor lizard (as have others, e.g. D'Amore & Blumenschine Reference D'Amore and Blumenschine2009). He interpreted two cranial elements as a partial maxilla preserving strongly compressed alveoli, and a piece of the rostrum preserving the anterior margin of the orbit. Based on the maxillary fragment, Cuvier concluded that the snout was deep relative to that of a modern crocodile.
Although his assessment of comparative snout depth was eventually verified, this seems to be a case of drawing the right conclusion from the wrong evidence. What Cuvier identified as a partial maxilla is actually a partial dentary (Fig. 2a). The other skull fragment (Fig. 2b, c) does seem to preserve a sutural contact trending from the border of an opening, but there is no indication on what would be the ventral surface of either the lacrimal duct or the prefrontal pillar. The specimen is equally consistent with a partial surangular and angular posterior to the external mandibular fenestra, though this is also equally tenuous.
Fossils from elsewhere in Europe have been referred to P. rollinati (e.g. Caraven-Cachin Reference Caraven-Cachin1880; Astre Reference Astre1931; Weitzel Reference Weitzel1938; Berg Reference Berg1966; de Lapparent de Broin et al. Reference Lapparent de Broin, Merle, Fontana, Ginsburg, Hervat, Le Calvez and Riveline1993; Windolf Reference Windolf1994; Rossmann Reference Rossmann1998; Kotsakis et al. Reference Kotsakis, Delfino and Piras2004; Sachse Reference Sachse2005). These might, indeed, represent a single species. Some of them are also of approximately the same age as the type – fossils from Argenton, Messel, and the lower horizons at Geiseltal are all of early Lutetian age (European Mammal Paleogene Zone [MP] 11; Sudre & Lecomte Reference Sudre and Lecomte2000; Franzen Reference Franzen2005). Most of the Geiseltal fossils studied for this analysis are geologically younger, deriving from MP 12, 13a, or 13b, but they are morphologically indistinguishable from older Geiseltal specimens and those from Messel, and they can be referred to the same species (Berg Reference Berg1966; Rossmann Reference Rossmann1998). It would thus be reasonable to suspect that the Lutetian ziphodont eusuchians from Europe represent a single species.
However, as suggested by Langston (Reference Langston1975), there is nothing diagnostic at the species level in the type material. The syntype serrated tooth and procoelous vertebra are enough to establish the presence of a ziphodont eusuchian at the type locality, but nothing more. Even if we include all of the material figured and discussed by Cuvier (Reference Cuvier1824), we would be unable to refer it, on morphological grounds alone, to the same species as the Messel/Geiseltal form and not to P. vorax. It follows that Pristichampsus rollinati (Gray Reference Gray1831) is a nomen dubium.
1.1.1. Status of Pristichampsus
Because its type species is a nomen dubium, Pristichampsus Gervais, Reference Gervais1853 itself is undiagnosable. The oldest generic name available for the eusuchians formerly called Pristichampsus appears to be Boverisuchus Kuhn, Reference Kuhn1938. Limnosaurus Marsh, Reference Marsh1872 is based on material probably conspecific with P. vorax, but it can no longer be identified (Langston Reference Langston1975) and is, thus, a nomen nudum.
I do not say this lightly; Pristichampsus is widely used. Its elimination may lead to confusion in the literature and, in all likelihood, my own violent death at the hands of my colleagues. Taxonomic stability should be promoted, and I would rather not be killed. The ideal solution would be the discovery of a more complete and diagnosable specimen from Argenton that can serve as a neotype. In the absence of such material, we are obliged to set the name aside.
1.1.2. Clade name
The ICZN (1999) recommends, but does not mandate, the rejection of suprageneric names based on nomina dubia. The draft PhyloCode (Cantino & de Queiroz Reference Cantino and de Queiroz2007; article 11.8) requires that definitions of clade names derived from typified names include the types of these names as specifiers. It also requires that the specifier be included, either in a reference phylogeny or in a statement clarifying how the specifier relates to taxa in the phylogeny, but this does not necessarily mean that clade names become invalid should one of the specifying species be diagnostically problematic; a type species may not be diagnosable at the species level, but we may nonetheless be able to affirm a close relationship with a clade of better-known species.
The name Pristichampsinae is defined in reference to Pristichampsus rollinati and all crocodylians more closely related to it than to Gavialis gangeticus, Alligator mississippiensis and Crocodylus niloticus (Brochu Reference Brochu2003). We could maintain P. rollinati as a specifier for Pristichampsinae if we could comfortably assume it to be closely related to something like Boverisuchus magnifrons. This would have the desirable result of maintaining a widely-used name.
Nevertheless, there are reasons against following this approach. The association of ziphodont teeth with procoelous vertebrae is enough to distinguish the Argenton crocodile from any of the putative sebecids known from the European Palaeogene (Berg Reference Berg1966; Buffetaut Reference Buffetaut1982b, Reference Buffetaut1988; Vasse Reference Vasse1995; Ortega et al. Reference Ortega, Buscalioni and Gasparini1996; Rossmann et al. Reference Rossmann, Rauhe and Ortega2000), but the specimens, by themselves, are just as referable to the mekosuchine crocodyloid Quinkana as they are to Boverisuchus. A close relationship with coeval material from the same continent is somewhat more likely than with a substantially younger animal from the other side of the world, but taxonomic conclusions should be drawn on the specimens and the specimens alone. Analyses of historical biogeography and stratigraphic diversity patterns become circular if the taxa underpinning the analyses are based, in part, on biogeography or stratigraphy (Bell et al. Reference Bell, Gauthier and Bever2010). Independence of taxonomic data from nonbiotic signals (such as time and space) must outweigh taxonomic stability, however important taxonomic stability is.
The oldest suprageneric name available for these forms is Planocranidae Li Reference Li1976. The correct Latin spelling should actually be Planocraniidae, and as first reviser I amend the name as such. Planocraniidae is defined in reference to Planocrania datangensis 1976 and all eusuchians more closely related to it than to Alligator mississippiensis (Daudin Reference Daudin1802), Crocodylus niloticus Laurenti, Reference Laurenti1768, Gavialis gangeticus (Gmelin Reference Gmelin1789), Borealosuchus sternbergii (Gilmore Reference Gilmore1910), Thoracosaurus macrorhynchus (Blainville Reference Blainville1835), Allodaposuchus precedens Nopcsa, Reference Nopcsa1928, or Hylaeochampsa vectiana Owen, Reference Owen1874. The number of specifiers is increased because of the lability shown by basal crocodylians and related lineages in recent analyses (Martin & Buffetaut Reference Martin and Buffetaut2008; Pol et al. Reference Pol, Turner and Norell2009; Turner & Brochu Reference Turner and Brochu2010; Puértolas et al. Reference Puértolas, Canudo and Caruzado-Caballero2011; Brochu Reference Brochu2011; Brochu et al. Reference Brochu, Parris, Grandstaff, Denton and Gallagher2012).
The name Pristichampsinae, in the phylogenetic sense, has not actually been suppressed. Assuming the phylogenetic relationships presented in this paper are upheld, Planocraniidae would become redundant along with Pristichampsinae, should we someday be able to diagnose P. rollinati. This might happen if, for example, more complete material sufficient to create a neotype is found at the type locality. But unless that happens, we cannot provide a robust link between the Argenton sample and any particular ziphodont eusuchian without including nonbiotic information.
1.2. Boverisuchus vorax (Troxell Reference Troxell1925) and the North American Planocraniid record
Boverisuchus vorax, new combination, is based on material described by Troxell (Reference Troxell1925, as Crocodilus vorax), though now-lost material described by Marsh (Crocodilus ziphodon; Reference Marsh1871) probably belonged to this species. Langston (Reference Langston1975), following comparison with European material, referred it to Pristichampsus.
The specimens used in this analysis are all from Wyoming. Most, including the holotype (Fig. 3d), are from the Bridger Formation (and most of these are from the Grizzly Buttes area), and the exceptionally preserved skull described by Langston (Reference Langston1975; FMNH PR399, Fig. 3a, b, c) is from the correlative Lower Washakie Formation. All are thus from the later part of the Bridgerian North American Land Mammal Age (NALMA), corresponding approximately with the lower part of MP 11 in Europe.
Figure 3 Boverisuchus vorax, middle Eocene (Bridgerian), Wyoming: (A–C) FMNH PR399, skull, in dorsal (A), ventral (B) and left lateral (C) views; (D) YPM 249 (holotype), disarticulated skull with associated postcranial material; (E–G) UCMP 170767, right dentary, in medial (E), lateral (F) and dorsal (G) views; (H–I) AMNH 29993, postdentary region of right mandibular ramus, in lateral (H) and medial (I) views. FMNH PR399 is from the lower Washakie Formation; remaining specimens are from the Bridger Formation. Scale bars=10 cm (A–D); 5 cm (E–I). Abbreviations: an=angular; art=articular; bo=basioccipital; d=dentary; ect=ectopterygoid; emf=external mandibular fenestra; en=external naris; eoa=external otic aperture; ex=exoccipitial; f=frontal; fio=foramen intermandibularis oralis; ic=internal choana; inf=incisive foramen; itf=infratemporal fenestra; j=jugal; l=lacrimal; meu=median Eustachian foramen; mx=maxilla; n=nasal; o=orbit; pa=parietal; pal=palatine; pf=prefrontal; pmx=premaxilla; po=postorbital; pos=preotic siphonium; pt=pterygoid; q=quadrate; qj=quadratojugal; sa=surangular; sof=suborbital fenestra; sp=splenial; sq=squamosal; stf=supratemporal fenestra.
1.2.1. Other North American occurrences
Boverisuchus has been reported from North American deposits ranging in age from the Uintan down to the base of the Palaeocene (Bramble & Hutchison Reference Bramble and Hutchison1971; Bartels Reference Bartels1980; Gingerich Reference Gingerich1989; Westgate Reference Westgate1989, Reference Westgate2008; Gunnell et al. Reference Gunnell, Bartels, Gingerich and Torres1992; Hanson Reference Hanson, Prothero and Emry1996; Stucky et al. Reference Stucky, Prothero, Lohr, Snyder, Prothero and Emry1996; Gunnell & Bartels Reference Gunnell and Bartels1999; Zonneveld et al. Reference Zonneveld, Gunnell and Bartels2000; Alexander & Burger Reference Alexander, Burger and Gunnell2001). Most are limited to fragmentary material, primarily teeth, and cannot be referred to Boverisuchus, or even to Planocraniidae, without making spatiotemporal assumptions.
The oldest reported occurrence is an isolated tooth, PU 17074, from the early Palaeocene (Puercan) Mantua beds of Wyoming (Bartels Reference Bartels1980). The specimen was not figured, and it can no longer be located in the PU collections (now housed at the YPM). We would predict the presence of planocraniids in rocks of this age, but without a specimen, it is difficult to substantiate.
Some have suggested that Orthogenysuchus olseni Mook, Reference Mook1924, an enigmatic crocodylian from the Wasatchian of Wyoming, is a planocraniid (Gunnell et al. Reference Gunnell, Bartels, Gingerich and Torres1992; Rauhe & Rossmann Reference Rauhe and Rossmann1995; Rossmann Reference Rossmann1998). Orthogenysuchus is known only from a poorly-preserved skull, but the teeth are neither compressed nor serrated (Brochu pers. obs.), and in spite of being dorsoventrally compressed, it does not appear to have been especially deep-snouted. It preserves several character states unique to alligatoroids, and some phylogenetic analyses support a close relationship with the outlandishly bizarre South American caimanine Mourasuchus Price, Reference Price1964 (Brochu Reference Brochu1999, Reference Brochu2010; Aguilera et al. Reference Aguilera, Riff and Bocquetin-Villanueva2006; Hill & Lucas Reference Hill and Lucas2006; Bona Reference Bona2007; Martin Reference Martin2007). This hypothesis is not universally accepted (Langston Reference Langston2008), and recent discoveries in Utah (Masters et al. Reference Masters, Sandau, Burk and Krumenacker2010) suggest a close relationship with another poorly-known alligatoroid from the Palaeogene of North America, Listrognathosuchus multidentatus (Mook Reference Mook1930). Moreover, the close relationship between Orthogenysuchus and Mourasuchus has collapsed in recent analyses, largely because of some newly-included incomplete early caimanines (Brochu Reference Brochu2011). Nonetheless, Orthogenysuchus appears to be an alligatoroid and not a planocraniid.
The oldest Cenozoic North American ziphodont crocodyliforms presently in museum collections are isolated teeth from the late Palaeocene (Tiffanian) of Moffatt County, Colorado (Fig. 4) and Park County, Montana (Gunnell et al. Reference Gunnell, Bartels, Gingerich and Torres1992). If these are planocraniid, the coarse serrations indicate a closer relationship to Boverisuchus than to Planocrania datangensis.
Figure 4 UCM 98628, ?planocraniid tooth, DeBeque Formation (Tiffanian), Moffat Co., Colorado: (A) labial view; (B) mesial view. Scale bar=1 cm.
Uintan occurrences include an isolated maxilla from San Diego County, California (Fig. 5a, b), and fragmentary material from elsewhere in California, Oregon, and southern Texas (Bramble & Hutchison Reference Bramble and Hutchison1971; Westgate Reference Westgate1989, Reference Westgate2008; Hanson Reference Hanson, Prothero and Emry1996). The most informative material is from the Devil's Graveyard Formation of Brewster County, western Texas, including an isolated left premaxilla (TMM 41747-52, Fig. 5c, d, e) that Busbey (Reference Busbey1986) tentatively referred to Pristichampsus vorax. It differs from Bridgerian Boverisuchus premaxillae in some respects – it is mediolaterally narrower, and whereas occlusal pits for the dentary teeth are lingual to the alveoli in B. vorax, they are between the alveoli in the Devil's Graveyard form. The Devil's Graveyard specimen is comparatively small and possibly immature, and Busbey (Reference Busbey1986) suggested that the differences between Bridgerian premaxillae and TMM 41747-52 might be ontogenetic. Although the shape of the premaxilla changes during ontogeny in modern crocodylians, occlusal patterns generally do not (Brochu pers. obs.), and in-line occlusion between the premaxillary and anteriormost dentary teeth is characteristic of Boverisuchus from Germany. The Devil's Graveyard form might be a distinct species, but more material is needed before it can be characterised.
Figure 5 Uintan planocraniids from North America: (A–B) LACM 21002, left maxilla, San Diego Co., California, in lateral (A) and medial (B) views; (C–E) TMM 41717-52, left premaxilla, Devil's Graveyard Formation, Brewster Co., Texas, in dorsal (C), ventral (D) and lateral (E) views. Scale bar=1 cm. Abbreviations: en=external naris; inf=incisive foramen; op=occlusal pit; p1–p5=premaxillary alveoli.
Jamaica was also home to a ziphodont crocodyliform during the middle Eocene (Vélez-Juarbe & Brochu Reference Veléz-Juarbe, Brochu, Portell and Domningin press). Other vertebrates from the locality are of North American origin (Domning et al. Reference Domning, Emry, Portell, Donovan and Schindler1997; Pregill Reference Pregill1999), which would suggest a planocraniid affinity for the tooth, but such a conclusion cannot be drawn from the specimen itself.
1.3. Boverisuchus magnifrons Kuhn, Reference Kuhn1938 and the European Planocraniid record
The holotype of B. magnifrons is from the Lagerstätte at Geiseltal near Halle, Germany (Kuhn Reference Kuhn1938). Codings in this analysis are based primarily on this and other well-preserved skeletons and skulls from Geiseltal (Fig. 6; Kuhn Reference Kuhn1938; Rossmann Reference Rossmann1998, Reference Rossmann2000a). Material from Messel (Weitzel Reference Weitzel1938; Berg Reference Berg1966) was also considered. All of these specimens are of Lutetian age.
Figure 6 Boverisuchus magnifrons, middle Eocene, Geiseltal, Germany: (A) GM LEO X 8001 (holotype, Weigeltisuchus geiseltalensis), skull, oblique ventral view ; (B) GM XXXVI 274, skull, dorsal view; (C) GM XVIII 3094, skull, ventral view ; (D) GM XXXV 216, left surangular and articular, dorsal view; (E–G) GM XXXVI 499, anterior end of right dentary, in medial (E), dorsal (F) and lateral (G) views; (H–I) GM XXXVI 245, right mandibular ramus, in lateral (H) and medial (I) views. Scale bar=5 cm. Abbreviations: an=angular; art=articular; bo=basioccipital; d=dentary; ect=ectopterygoid; emf=external mandibular fenestra; en=external naris; f=frontal; inf=incisive foramen; itf=infratemporal fenestra; j=jugal; mg=Meckelian groove; mx=maxilla; n=nasal; o=orbit; pf=prefrontal; pmx=premaxilla; po=postorbital; pt=pterygoid; q=quadrate; qj=quadratojugal; sa=surangular; sof=suborbital fenestra; sp=splenial; sq=squamosal; stf=supratemporal fenestra.
The Messel form has always been referred to P. rollinati (Weitzel Reference Weitzel1938; Berg Reference Berg1966), but at Geiseltal, the specimens were initially thought to represent two forms – a caiman-like alligatorid, Boverisuchus magnifrons, and a crocodylid, Weigeltisuchus geiseltalensis Kuhn Reference Kuhn1938. I follow Kälin (Reference Kälin and Piveteau1955) and recognise W. geiseltalensis as a junior synonym of B. magnifrons. Kuhn (Reference Kuhn1938) acknowledged similarities between them, and the caiman-like features he indicated in B. magnifrons (larger dorsal supraoccipital exposure and constricted supratemporal fenestrae) are concentrated on the skull table, which is very poorly preserved on the holotype (Brochu pers. obs.).
Rauhe & Rossmann (Reference Rauhe and Rossmann1995) and Rossmann (Reference Rossmann1998) argued that B. vorax and B. magnifrons, along with Planocrania datangensis from the Early Eocene of China, represent a single pan-Laurasian species. They referred all such material to P. rollinati. Skulls from Wyoming (Fig. 3) are consistently mediolaterally broader than those from Germany (Fig. 6a, b), but care must be taken to consider the levels of variation seen in modern species (Kälin Reference Kälin1933). Moreover, most known skulls are compressed, distorting the original shape and complicating our comparisons. Rossmann (Reference Rossmann1998, Reference Rossmann2000a) argued that the differences between American and European samples were insufficient to justify species-level separation.
Nonetheless, there are consistent differences that lie outside the observed ranges of variation for modern crocodylian species. They also appear to be unrelated to ontogenetic stage or body size. One of these is the relationship between the nasals and the external naris. Rossmann (Reference Rossmann2000a) stated that the nasals of P. rollinati contact the external naris except in one specimen from Geiseltal. One of his figures (Rossmann Reference Rossmann2000a, fig. 2) implies a similar situation in at least one specimen from North America (USNM 12957), but the surface of the rostrum is not preserved immediately behind the naris, and the nasals enter the naris in all other appropriately preserved Bridgerian specimens (Fig. 7a). Conversely, there are no known Geiseltal specimens in which the nasals contact the naris, and those specimens in which the relevant part of the skull is preserved invariably show a broad separation between them (Fig. 7b, c). The dorsal surface of the rostrum is imperfectly preserved on the most complete Messel specimen (HLMD Me-5326), but the orientation of the right nasopremaxillary suture relative to the sagittal plane strongly suggests that the premaxillae completely surrounded the naris (Brochu pers. obs.).
Figure 7 Anterior end of rostrum: (A) FMNH PR399, Boverisuchus vorax, dorsal view; (B–C) GM XXXVI 245, Boverisuchus magnifrons, dorsal view; (D) FMNH PR399, Boverisuchus vorax, ventral view; (E) GM XVIII 526, Boverisuchus magnifrons, ventral view. Scale bars=1 cm. Abbreviations: en=external naris; inf=incisive foramen; mx=maxilla; n=nasal; op=occlusal pit; pmx=premaxilla; p1–p5=premaxillary alveoli.
Dental occlusion patterns at the tip of the rostrum differ between samples. Based on occlusal pits preserved on the premaxillae, the dentary teeth occluded lingual to their premaxillary counterparts in B. vorax (Fig. 7d). The occlusal pits are between, rather than lingual to, premaxillary alveoli in B. magnifrons (Fig. 7e). And although the anterior maxillary teeth are ziphodont in both forms, the first six alveoli are circular in B. vorax (Fig. 3b) and labiolingually compressed in B. magnifrons (Fig. 6a)
The mandibular symphysis of B. magnifrons (Fig. 6i) is narrower in dorsal view than that of B. vorax (Fig. 3e). This, of course, is consistent with the relatively narrower snout in the German form, and one could argue that such differences are intraspecific in nature. But the anterior end of the dentary between the first and fourth alveoli is also more deeply concave in B. magnifrons, even taking postmortem distortion into account (Fig. 6e, h).
Snout shape, by itself, is not a robust difference between known Middle Eocene planocraniids. But taken along with the other consistent differences – whether or not the nasals contribute to the narial rim, whether occlusal pits are lingual to or between premaxillary alveoli, whether the frontoparietal suture contacts the supratemporal fenestrae, and the shape of the dentary – synonymy between B. vorax and B. magnifrons can be rejected.
1.3.1. Crocodilus bolcensis
Crocodilus bolcensis Sacco, Reference Sacco1896, based on a skull and articulated skeleton from the Lutetian Monte Purga di Bolca locality of northeastern Italy, is a possible complicating factor. Synonymy with the German material was suggested by Berg (Reference Berg1966) and accepted by later authors (e.g. Rossmann Reference Rossmann1998; Kotsakis et al. Reference Kotsakis, Delfino and Piras2004). Skull shape is constent with Messel and Geiseltal material, and fossils from that locality are correlative with Argenton, Messel and lower Geiseltal. One might expect the planocraniids from these regions to be conspecific.
Determining the status of this species bears on nomenclature. Pristichampsus rollinati (Gray Reference Gray1831) has priority over Crocodilus bolcensis Sacco, Reference Sacco1896, but C. bolcensis has priority over Boverisuchus magnifrons Kuhn, Reference Kuhn1938. The correct name for the European planocraniid would thus be Boverisuchus bolcensis (Sacco Reference Sacco1896).
Nevertheless, there are reasons to be cautious. Surface details are not readily visible on the holotype (Sacco Reference Sacco1896, fig. 5b), and derived character states diagnostic of B. magnifrons (e.g. exclusion of the nasals from the naris) are not readily apparent. The teeth of another specimen (Fig. 8) appeared to be unserrated when examined with a 10× hand lens (Brochu pers. obs.), and the mandibular symphysis appears to be craniocaudally short and mediolaterally wide compared with those of B. magnifrons or B. vorax. This specimen shows no evidence of the ventral armor preserved on articulated B. magnifrons skeletons from Geiseltal.
Figure 8 MGPD 27567, Crocodilus bolcensis, middle Eocene, Purga di Bolca, Italy: (A) skeleton. Scale bar=10 cm; ( B) closeup of skull and lower jaws.
Because so little morphological detail could be obtained from these specimens (in spite of their completeness), the species was not included in this analysis. A closer examination of this material is warranted.
1.3.2. Other European occurrences
In Europe, as in North America, fragmentary specimens might extend the range of planocraniids below the Lutetian. Berg (Reference Berg1969) cited a possible planocraniid from the Paleocene Walbeck locality of Germany, and fossils from the early Eocene might be referable to the group (Berg Reference Berg1966; Russell Reference Russell1982; Buffetaut Reference Buffetaut1985; Antunes Reference Antunes1986; Vasse Reference Vasse1992; Augé et al. Reference Augé, Duffaud, Lapparent de Broin, Rage and Vasse1997). They have also been reported from the Bartonian (Russell Reference Russell1982; Antunes Reference Antunes1986).
But while only Planocraniidae is known with certainty in the Palaeogene of North America, non-eusuchian ziphodont crocodyliforms have been found in the Eocene of Europe (Kuhn Reference Kuhn1938; Berg Reference Berg1966; Antunes Reference Antunes1975; Buffetaut Reference Buffetaut1982b, Reference Buffetaut1988; Vasse Reference Vasse1995; Ortega et al. Reference Ortega, Buscalioni and Gasparini1996; Rossmann et al. Reference Rossmann, Rauhe and Ortega2000). Amphicoelous vertebrae consistent with these forms are known from the middle Paleocene of Belgium (Groessens-Van Dyck Reference Groessens-Van Dyck1986) and the Bartonian of France (Buffetaut Reference Buffetaut1986), and they are found in some of the same deposits preserving Boverisuchus at Messel and Geiseltal (Berg Reference Berg1966; Rauhe & Rossmann Reference Rauhe and Rossmann1995). Although some have argued that the teeth of a planocraniid can be distinguished from those of a non-eusuchian ziphodont form (Antunes Reference Antunes1986), there is considerable dental variation within these groups (Legasa et al. Reference Legasa, Buscalioni and Gasparini1993; Prasad & de Lapparent de Broin Reference Prasad and Lapparent de Broin2002) and more work is needed before we can distinguish the stratigraphic ranges of planocraniids and similar-looking but unrelated groups.
Atacisaurus glareae Astre, Reference Astre1931 from the middle Eocene of Issel, France, has also sometimes been referred to Boverisuchus (Rossmann Reference Rossmann1998). Vasse (Reference Vasse1992) argued that its systematic position was not determinable. The holotype, the symphyseal part of a lower jaw, is now lost (Ortega et al. Reference Ortega, Buscalioni and Gasparini1996). As figured by Astre (Reference Astre1931), the symphysis appears to have extended back past the tenth dentary alveolus and included a more substantial splenial component than in Boverisuchus, and Astre (Reference Astre1931, p. 37) described the carinae of the teeth as “non crénelée.”
Astre (Reference Astre1931) also referred a skull (Fig. 9a) to A. glareae. It appears to share no derived states with planocraniids. It does, however, share similarities with basal tomistomines such as Megadontosuchus Mook, Reference Mook1955. The lateral squamosal groove flares anteriorly (a feature also found in gavialoids), the supratemporal fenestrae are large and separated by a narrow interfenestral bar, and the skull table is concave at the midline in posterior view. The snout is relatively slender, but the lateral margins are concavoconvex in dorsal view and not linear, as they would be in a planocraniid. The teeth are unknown. A non-eusuchian ziphodont crocodyliform is known from Issel (Ortega et al. Reference Ortega, Buscalioni and Gasparini1996), but A. glareae appears to be a tomistomine and not a planocraniid.
Figure 9 (A) unnumbered specimen at MNHT referred by Astre (Reference Astre1931) to Atacisaurus glareae, middle Eocene, France: skull, dorsal view. Scale bar=10 cm. (B) NHMUK R.4112: Vertebrata indet. tooth, Miocene, Nyanza Province, Kenya. Scale bar=1 cm.
1.4. Planocrania datangensis Li, Reference Li1976
This is based on an articulated skull and partial mandible from the Nongshan Formation of Guandong Province, China (Li Reference Li1976; Fig. 10). It is the only known specimen of the species. The Nongshan Formation is thought to be of Paleocene (Salandian) age (Ting et al. Reference Ting, Bowen, Koch, Clyde, Wang, Wang and McKenna2003, Reference Ting, Tong, Clyde, Koch, Wang, Bowen, Li and Snell2011).
Figure 10 IVPP V5016 (holotype), Planocrania datangensis, Palaeocene, China: skull and articulated dentaries in dorsal (A), ventral (B) and right lateral (C) views. Scale bar=5 cm. Abbreviations: en=external naris; eoa=external otic aperture; itf=infratemporal fenestra; meu=median Eustachian foramen; o=orbit; ?plp=possible palpebral bone; stf=supratemporal fenestra.
Planocrania datangensis was referred to Pristichampsus rollinati by Rossmann (Reference Rossmann1998). The overall shape of the snout is similar between the two forms, but there are some clear differences. Most notably, although the teeth of P. datangensis are labiolingually compressed, serrations are not apparent (Fig. 11) even with a 10× hand lens. This does not appear to be a preservational artifact. If serrations were present (Li Reference Li1976), they were very fine and unlike the more robust serrations found in both species of Boverisuchus. The holotype of P. datangensis is somewhat smaller than most Boverisuchus skulls, but smaller Boverisuchus nonetheless have serrated teeth. The nasal bones also reach the external naris and the frontoparietal suture passes through the supratemporal fenestrae in P. datangensis.
Figure 11 (A) IVPP V5016 (holotype), Planocrania datangensis; (B) closeup of teeth of right maxilla. Scale bars=1 cm.
1.5. Planocrania hengdongensis Li, Reference Li1984
Like P. datangensis, P. hengdongensis is known from only a single specimen (Li Reference Li1984; Fig. 12). It was collected from what Li (Reference Li1984) described as red beds of possible Palaeocene age in Hunan Province, China. These are now known to be part of the late Palaeocene Lower Lingcha Formation (Ting et al. Reference Ting, Bowen, Koch, Clyde, Wang, Wang and McKenna2003).
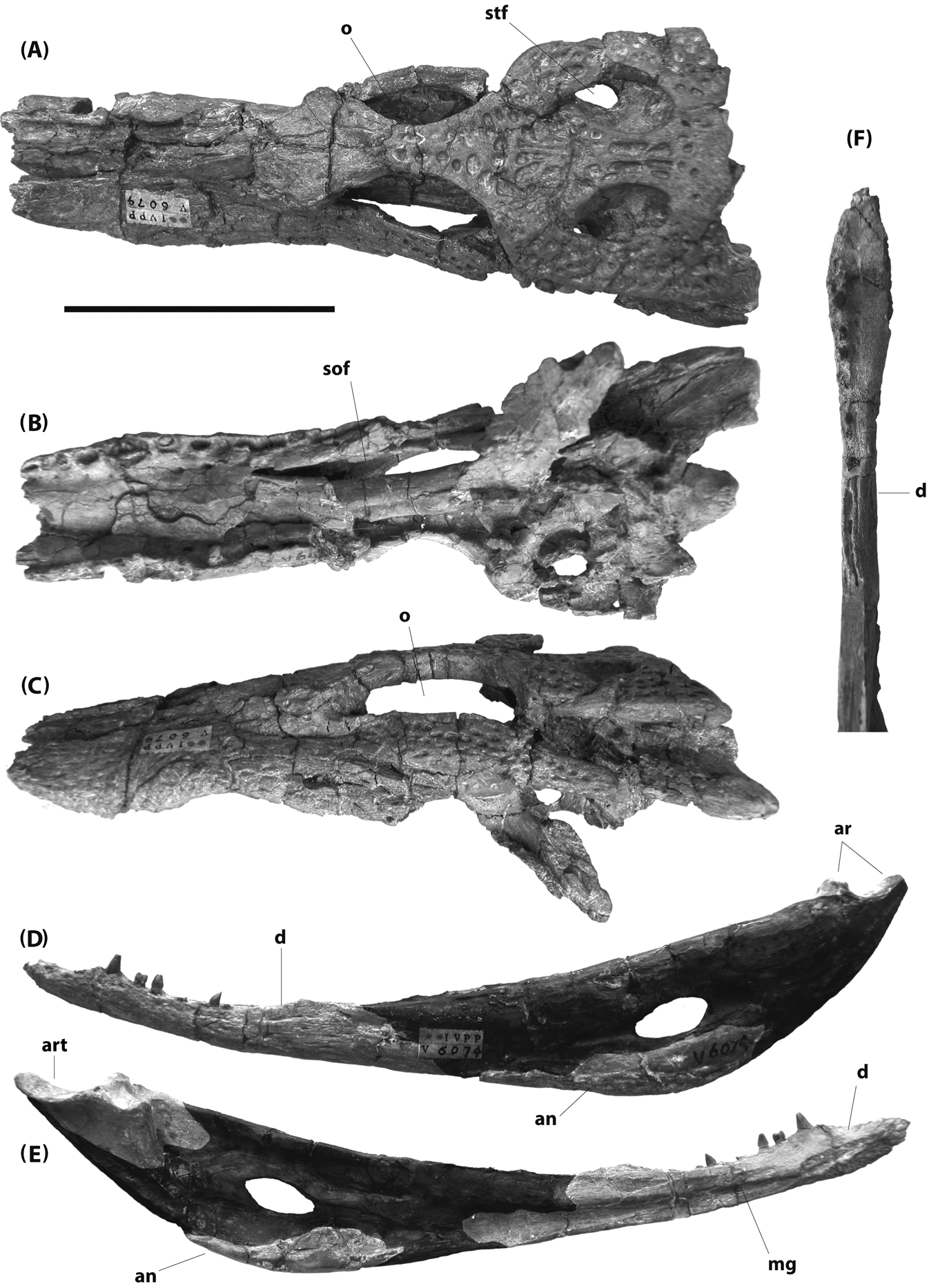
Figure 12 IVPP V6079 (holotype), Planocrania hengdongensis, Palaeocene, China: (A–C) skull in dorsal (A), ventral (B) and left lateral (C) views; (D–E) left mandible in lateral (D) and medial (E) views; (F) left dentary in dorsal view. Portions of mandibular ramus restored with epoxy digitally darkened in (D) and (E). Scale bar=5 cm. Abbreviations: an=angular; art=articular; d=dentary; mg=Meckelian groove; o=orbit; sof = suborbital fenestra; stf=supratemporal fenestra.
Li (Reference Li1984) stated that the maxillary teeth of P. hengdongensis are serrated. This is difficult to assess, as there are very few preserved tooth crowns available. The premaxilla described by Li (Reference Li1984) is no longer with the specimen. Only one maxillary tooth crown is preserved, and although compressed, it does not appear to be serrated. The same is true for teeth preserved on the left dentary.
There are several other differences between P. hengdongensis and P. datangensis. The frontal between the orbits is unusually broad in P. datangensis, as it is in Boverisuchus, but not in P. hengdongensis. The splenial does not contribute to the mandibular symphysis in P. hengdongensis, as it does in P. datangensis and Boverisuchus. Planocrania hengdongensis also appears to have a shorter maxillary toothrow than P. datangensis. Li (Reference Li1984) estimated 11 to 12 maxillary teeth in P. hengdongensis. The alveolar series on both maxillae are damaged, and the number could have been larger, but would not have exceeded 14 (and was most likely 12 or 13; Brochu pers. obs.). Planocrania datangensis, however, had 15 or 16 maxillary alveoli (Li Reference Li1976), as did Boverisuchus (Rossmann Reference Rossmann2000a).
The holotype of P. hengdongensis might be from an immature animal. Nevertheless, although some differences between P. datangensis and P. hengdongensis might be ontogenetic in nature, the number of alveoli in any of the tooth-bearing bones never increases ontogenetically in crocodylians. Indeed, the number sometimes decreases by one or two – faster-growing alveoli might crowd out their more slowly-growing neighbors. Even if P. hengdongensis is known from an immature individual, it can be distinguished from all other planocraniids.
1.6. Other Asian occurrences
Ziphodont crocodyliform teeth are known from the Eocene of the Indian Subcontinent (Sahni & Srivastava Reference Sahni and Srivastava1976; Buffetaut Reference Buffetaut1978; Sahni et al. Reference Sahni, Srivastava and D'Souza1978; Sah & Schleich Reference Sah and Schleich1990; Srivastava & Kumar Reference Srivastava and Kumar1996). A partial premaxilla with teeth from the early Eocene of Himachal Pradesh, India, was described by Panadés I Blas et al. (Reference Panadés I Blas, Loyal, Schleich and Agrasar2004).
Two putative planocraniids – Pristichampsus birjukovi Efimov, Reference Efimov1988 and Pristichampsus kuznetzovi Efimov, Reference Efimov1988 – have been described from middle Eocene deposits in the Zaysan Depression region of Kazakhstan. They have sometimes been synonymised with B. magnifrons (Rossmann Reference Rossmann1998). The holotype of P. kuznetzovi has not been figured. The skull of the holotype of P. birjukovi is limited to a skull table, braincase and left quadrate ramus but, as figured by Efimov (Reference Efimov1993), the quadrate condyle appears similar to those of B. magnifrons and B. vorax. This material should be revisited and reevaluated.
1.7. Australasia
A ziphodont eusuchian, Quinkana Molnar, Reference Molnar1981, is also known from the Late Oligocene through Quaternary of Australia and New Guinea (De Vis Reference De Vis1905; Molnar Reference Molnar1981; Megirian Reference Megirian1994; Willis & Mackness Reference Willis and Mackness1996; Willis Reference Willis1997). Phylogenetic analyses suggest that Quinkana is a mekosuchine crocodyloid and, thus, part of a diverse endemic Australasian radiation (Willis Reference Willis1993; Salisbury & Willis Reference Salisbury and Willis1996; Willis & Mackness Reference Willis and Mackness1996; Molnar et al. Reference Molnar, Worthy and Willis2002; Brochu Reference Brochu2007), but a relationship to Boverisuchus has also been proposed (Rauhe & Rossmann Reference Rauhe and Rossmann1995; Rossmann Reference Rossmann1998). This would suggest transoceanic dispersal from Laurasia and a substantial missing history for the group.
None of the described species is well known, and it was represented by a composite taxon in earlier analyses based on this matrix (e.g. Brochu Reference Brochu2007; Vélez-Juarbe & Brochu Reference Veléz-Juarbe, Brochu, Portell and Domningin press). Most data came from Q. fortirostrum Molnar, Reference Molnar1981, which was observed first-hand, and Q. timara Megirian, Reference Megirian1994, which was coded from the literature. The other two species (Q. babarra Willis & Mackness, Reference Willis and Mackness1996 and Q. meboldi Willis, Reference Willis1997) were largely redundant with Q. fortirostrum and Q. timara in the matrix. The analyses unambiguously put Quinkana among the mekosuchines, and moving the composite to Planocraniidae increases tree length by at least 12 steps.
Depending on how they are applied, composite taxa make a priori assumptions about phylogenetic relationships that should be tested. Unfortunately, although the species currently referred to Quinkana collectively support a mekosuchine affinity, they are individually incomplete enough to limit their informative value and were not included in this analysis. Additional material relevant to mekosuchine phylogeny is currently under study (Salisbury et al. Reference Salisbury, Holt, Worthy, Sand and Anderson2010), and its inclusion, along with a more detailed comparison with the known species of Quinkana, will enhance our capacity to test the relationships of these species.
1.8. Africa
The Cenozoic fossil record of ziphodont crocodyliforms in Africa is spotty at best. Non-eusuchian forms have been described from the Eocene of Algeria (Buffetaut Reference Buffetaut1982a, Reference Buffetaut1989), and an isolated tooth from the Miocene of present-day Kenya was discussed by Andrews (Reference Andrews1914), who suggested a possible referral to Planocraniidae. If correct, it would represent a substantial geographic and stratigraphic range extension for the group.
The tooth is compressed, but although the carinae are expansive, they are unserrated (Fig. 9b). The enamel is translucent, and it is not clear it had a root. It is difficult to refer the specimen to Crocodyliformes, much less Planocraniidae.
2. Phylogenetic analysis
2.1. Methods
A maximum parsimony analysis was conducted based on a matrix of 89 ingroup taxa and 189 discrete morphological characters (Appendix 1). All characters had equal weight, and multistate characters were treated as unordered. Bernissartia fagesii was the outgroup. One hundred heuristic searches were conducted using PAUP* (Swofford Reference Swofford2002) with the starting order of ingroup taxa randomised for each iteration.
A list of planocraniid specimens used for scoring the matrix is provided in Appendix 2.
2.2. Results
The analysis recovered 5044 equally optimal trees 633 steps in length (CI excluding uninformative characters=0·367, RI=0·820). A strict consensus of these trees (Fig. 13) is consistent with the results of previous morphological analyses of Crocodylia (e.g., Salisbury & Willis Reference Salisbury and Willis1996; Brochu Reference Brochu1997, Reference Brochu2004, Reference Brochu2006, Reference Brochu2011; Delfino et al. Reference Delfino, Piras and Smith2005, Reference Delfino, Codrea, Folie, Dica, Godefroit and Smith2008a, Reference Delfino, Martin and Buffetautb; Piras & Buscalioni Reference Piras and Buscalioni2006; Martin Reference Martin2007; Ösi et al. Reference Ösi, Clark and Weishampel2007; Shan et al. Reference Shan, Wu, Cheng and Sato2009; Puértolas et al. Reference Puértolas, Canudo and Caruzado-Caballero2011; Brochu & Storrs Reference Brochu and Storrs2012), albeit with diminished resolution at some levels. The closest relatives of Crocodylia in this study are the hylaeochampsids, and the root node of Crocodylia is a polytomy involving gavialoids, Borealosuchus and a lineage including Crocodyloidea, Alligatoroidea and Planocraniidae. Planocraniidae is the sister lineage to Brevirostres.
Figure 13 Strict consensus of 5044 equally optimal trees recovered from maximum parsimony analysis of 89 ingroup taxa and 189 morphological characters (length=633, CI excluding uninformative characters=0·367, RI=0·820). Trees rooted on Bernissartia fagesii.
2.3. Position of Planocraniidae within Crocodylia
A distinct convex process or lamina is present on the anterolateral corner of the paramedian osteoderms of gavialoids, Borealosuchus, and non-crocodylian eusuchians (Brochu Reference Brochu2004; Brochu et al. Reference Brochu, Parris, Grandstaff, Denton and Gallagher2012). This is absent in alligatoroids, crocodyloids and planocraniids. Some basal alligatoroids (e.g. Diplocynodon) and crocodyloids (e.g. “Crocodylus” affinis) preserve a modest concavity on the anterior margin of the paramedian osteoderms (Brochu pers. obs.); this may indicate a remnant of the ancestral lamina, but none of these forms has anything like the plesiomorphic condition, and its absence unites Planocraniidae and Brevirostres in this analysis.
The second unambiguous synapomorphy at this node comes as a surprise – a quadrate ramus with a small and prominently depressed medial hemicondyle. This was previously thought to diagnose Alligatoroidea (Fig. 14e) and to be a consequence of the dorsal shift in the position of the quadrate foramen aëreum (Brochu Reference Brochu1999). But in fact, a similar quadrate is preserved on the holotype of P. hengdongensis (Fig. 14a). The foramen aëreum is not located as far dorsally in this form, suggesting that depression of the medial hemicondyle is unrelated to changes in quadrate-articular pneumaticity.
Figure 14 Morphology of the left quadrate condyle in planocraniids (A–C) and other crocodylian groups (D–F): (A) IVPP V6079 (holotype), Planocrania hengdongensis; (B) FMNH PR479, Boverisuchus vorax; (C) GM XXII 815, Boverisuchus magnifrons; (D) FMNH 23505, Gavialis gangeticus; (E) FMNH 69871, Paleosuchus palpebrosus; (F) FMNH 17157, Crocodylus niloticus. D, E, and F are digitally-reversed right quadrates. Scale bars=1 cm. Abbreviations: fae=foramen aëreum; lhc=lateral hemicondyle; mhc=medial hemicondyle.
There is some distortion to the quadrate ramus of P. hengdongensis. I am confident that the general outline of the condylar region is close to what a fresh skull would reveal, but one could argue that modest damage to a plesiomorphic crocodylian quadrate – one in which the medial hemicondyle is small and ventrally reflected, but without the sharp demarcation – might mimic the condition seen in alligatoroids. Caution is also required because the quadrates are poorly known in P. datangensis, and neither Boverisuchus (Fig. 14b, c) nor crocodyloids (Fig. 14f) share this condition.
In previous analyses (e.g. Brochu Reference Brochu1997), keeled paramedian osteoderms were thought to diagnose this clade. This is no longer true if Borealosuchus is a basal gavialoid lineage (as is the case in some optimal trees in this analysis); the osteoderms of Bernissartia and at least some hylaeochampsids (e.g. Acynodon adriaticus) are keeled. In that case, the lack of keeled osteoderms links Borealosuchus and gavialoids.
Likewise, whether exclusion of the nasal bones from the naris diagnoses this group depends on how basal crocodylian relationships are resolved. The nasals form part of the narial border in most basal eusuchians (including hylaeochampsids), basal planocraniids and (at least ancestrally) in alligatoroids and crocodyloids, but not in gavialoids or Borealosuchus. A close relationship between Borealosuchus, Planocraniidae and Brevirostres would require loss of this contact at the root of Crocodylia and its reacquisition at this level. A closer relationship between Borealosuchus and gavialoids would be supported by this feature, making the condition seen in basal planocraniids ancestral for Crocodylia.
Another complication is the Gavialis–Tomistoma issue. Moving all putative gavialoids up with Tomistoma (and thus rendering Planocraniidae a non-crocodylian lineage) renders keeled osteoderms and nasals contacting the naris plesiomorphic for Crocodylia. If, following divergence time estimates based on molecular analyses (Harshman et al. Reference Harshman, Huddleston, Bollback, Parsons and Braun2003; Roos et al. Reference Roos, Aggarwal and Janke2007; Oaks Reference Oaks2011), we move only post-Lutetian gavialoids (Eogavialis, Gavialis, Gryposuchinae) and leave the thoracosaurs where they are, these conditions would be plesiomorphic for Crocodylia only if Borealosuchus and thoracosaurs were closely related.
Some planocraniids preserve a mosaic of crocodile-like and alligator-like features. The iliac blade has a modest indentation in Boverisuchus and, ancestrally, in Crocodyloidea. Indentations arise independently in caimans and Alligator, but the iliac blade in basal alligatoroids is smooth. Likewise, the lateral carotid foramen on the exoccipital opens dorsal to the posterolateral exposure of the basisphenoid in B. vorax and all crocodyloids. Conversely, the squamosal–quadrate suture of B. vorax extends no further than the posteroventral corner of the auditory aperture, as in most alligatoroids. These character states cannot be assessed in other planocraniids.
2.4. Relationships within Planocraniidae
Support for planocraniid monophyly in this study is comparatively thin. Only a single unambiguous synapomorphy unites the group – labiolingually compressed teeth throughout the maxillary or post-caniniform dentary toothrows. The posteriormost teeth are somewhat longer mesiodistally than wide labiolingually in most crocodylians, and strong compression can be observed in the living smooth-fronted caimans (Paleosuchus) and dwarf crocodiles (Osteolaemus), but this is true only for teeth toward the back of the mouth. Only in planocraniids (and Quinkana) do we see strong compression throughout the maxillary series and in most of the dentary teeth.
Planocrania datangensis and Boverisuchus share upturned orbital margins. This feature arose multiple times within Crocodylia, but is absent from Borealosuchus and the earliest gavialoids, crocodyloids and alligatoroids. At the same time, the frontal is very broad between the orbits in P. datangensis and Boverisuchus. Indeed, the frontal as a whole and the orbital surfaces in ventral view are comparatively broad in these forms compared with most other eusuchians.
The broad frontal may serve a similar function as the expanded compound palpebrals in living deep-snouted crocodylians such as Paleosuchus and Osteolaemus – it may help reorient the orbits. The eyes face more laterally in deep-snouted forms than they do in other crocodylians. The large palpebrals of Paleosuchus were once thought to help protect the eyes from debris in flowing streams (Medem Reference Medem1958), but neither Paleosuchus nor Osteolaemus are exclusively (or even preferentially) found in such environments (Magnusson Reference Magnusson1985; Magnusson & Lima Reference Magnusson and Lima1991; Kofron Reference Kofron1992; Ouboter Reference Ouboter1996; Luiselli et al. Reference Luiselli, Akani and Capizzi1999; Riley & Huchzermeyer Reference Riley and Huchzermeyer1999; Shirley et al. Reference Shirley, Oduro and Beibro2009). That two groups with similar modifications to the skull and osteoderms would share similar palpebral bones suggests a common cause. These structures are more or less fixed and move very little when the eyelid is closed, and they may instead move the medial margin of the orbit laterally, thereby reorienting the orbit itself. Derived planocraniids may have found a different solution – the frontals themselves expand. The holotype of P. datangensis appears to preserve articulated palpebrals (Fig. 10a), and these, too, may have helped impart a lateral orientation to the orbits, but palpebrals are unknown in Boverisuchus and are universally lacking in articulated skeletons.
In Planocrania datangensis and Boverisuchus, the splenials meet at the midline. This is not true for P. hengdongensis, but whether this is a synapomorphy at this level depends on how basal crocodylian lineages are resolved. Most basal crocodylians have splenial symphyses, and the lack of this feature could instead diagnose P. hengdongensis.
Likewise, occlusal patterns may or may not diagnose this node. Ancestrally for Eusuchia, the dentary caniniforms occluded in a notch between the maxilla and premaxilla, but dentary teeth occluded lingual to their maxillary counterparts otherwise. Derived alligatoroids lost the notch, and derived crocodyloids lost the overbite (Brochu Reference Brochu2003). Planocrania datangensis has an intermediate condition with occlusal pits between (and not lingual to) the seventh and eighth maxillary alveoli, but an overbite behind the premaxillary–maxillary notch otherwise. This is the condition found in basal species of Borealosuchus (B. sternbergii), a few alligatoroids, and in all but the basalmost crocodyloids. Other planocraniids, by contrast, retain the plesiomorphic maxillary overbite. All known gavialoids (including thoracosaurs) have completely interdigitating dentition. As a result, it is difficult to resolve the ancestral condition for this character in early crocodylians.
Boverisuchus is diagnosed by the presence of serrated teeth. The teeth of P. hengdongensis and P. datangensis are flattened, but the carinae are either smooth or bear serrations that cannot be seen with the unaided eye. In contrast, the teeth of B. vorax and B. magnifrons bear prominent serrations.
A highly modified quadrate condyle might also diagnose this node, though the absence of information about the quadrates of P. datangensis renders it ambiguous. In Boverisuchus, there is a prominent dorsal projection on the surface of the quadrate between the hemicondyles (Fig. 14b, c; Langston Reference Langston1975; Rossmann Reference Rossmann2000a). Similar quadrates are seen in some noneusuchian altirostral taxa, such as Sebecus (Langston Reference Langston1975), and there might be a correlation between this quadrate type and lateral snout flattening. Whether Quinkana had a similar quadrate is unclear.
3. Discussion
I sought to accomplish two goals with this manuscript: first, clarifiy the nomenclatural status and species-level diversity of Paleogene ziphodont eusuchians; and second, provide a preliminary estimate of their phylogenetic relationships.
The material available to Cuvier in the 1820s was visibly very different from any other crocodyliform known at the time, but subsequent discoveries in Europe, North America and Australia have diminished its diagnostic value. The purposes of precision are best served by constructing our nomenclatural framework on more diagnosable specimens. From the material studied for this analysis, at least four valid species can be recognised – Boverisuchus magnifrons, Boverisuchus vorax, Planocrania datangensis, and Planocrania hengdongensis. Crocodilus bolcensis and planocraniids of Uintan age from North America may also be distinguishable, but more information is needed.
Although it appears distinctive, phylogenetic support for planocraniid monophyly is weaker than we might like. There are two reasons for this. First, although two derived planocraniids (B. vorax and B. magnifrons) are well-known, their more basal relatives are not. Neither is the basalmost known crocodyloid (Prodiplocynodon). Secondly, relationships at the root of Crocodylia are less stable than in previous analyses. Whether Borealosuchus is closer to Brevirostres or to Gavialoidea is an open question, and we are beginning to understand just how alligator-like the closest relatives of Crocodylia were (Turner & Brochu Reference Turner and Brochu2010). Characters formerly diagnosing Alligatoroidea and subordinate clades may instead by plesiomorphic at a more inclusive level. For the moment, planocraniids seem to be linked together as animals sharing similar rostral and dental geometry and exclusion from the Alligatoroidea+Crocodyloidea group.
With only four species in the analysis, we can say little about planocraniid historical biogeography, except that the group appears to be of Laurasian origin. All known planocraniids are from North America and Eurasia, as are the earliest known members of proximate clades. This is consistent with broad phylogenetic and biogeographic patterns throughout the Palaeogene; while endemic radiations dominated crocodyliform diversity during the Neogene, lineages from earlier in the Cenozoic tended to be morphologically uniform and geographically widespread (Brochu Reference Brochu2003).
The evolutionary history suggested by our limited sample bears on previous speculation about planocraniid paleoecology. Ziphodont crocodyliforms have generally been interpreted as terrestrial predators (Molnar Reference Molnar1981; Rossmann Reference Rossmann1999, Reference Rossmann2000b, Reference Rossmannc), but as noted by Wroe (Reference Wroe2002), many of the features thought to indicate a more terrestrial habit for Boverisuchus – deep snout, laterally-directed orbits, compressed teeth, improved capacity to hold the head in an elevated position – are found in modern Paleosuchus, which is still a semiaquatic animal, even if it sometimes spends less time in the water than other sympatric crocodylians (Medem Reference Medem1958; Magnusson Reference Magnusson1985; Magnusson & Lima Reference Magnusson and Lima1991; Ouboter Reference Ouboter1996). The same is true for Osteolaemus – it has a deep snout with compressed teeth, but would not be described as a terrestrial predator (Kofron Reference Kofron1992; Luiselli et al. Reference Luiselli, Akani and Capizzi1999; Riley & Huchzermeyer Reference Riley and Huchzermeyer1999; Shirley et al. Reference Shirley, Oduro and Beibro2009). One might instead argue that smooth-fronted caimans and dwarf crocodiles are reasonable modern analogs for planocraniid ecology.
The analogy might be imperfect. Similarities to Paleosuchus and Osteolaemus are greatest with the oldest planocraniids known from more than teeth – Planocrania hengdongensis and P. datangensis. Whether the serrated teeth and specialised quadrate seen in Boverisuchus indicate an ecological difference is unclear, but their larger body size might. The two known specimens of Planocrania might be juveniles, but Boverisuchus – though not really a large crocodyliform – regularly exceeded the maximum sizes attained by Paleosuchus and Osteolaemus (Medem Reference Medem1958; Luiselli et al. Reference Luiselli, Akani and Capizzi1999; Campos et al. Reference Campos, Sanaiotti and Magnusson2010). There are also some postcranial differences between Boverisuchus and the living dwarf forms, such as the hooflike unguals (which are not a preservational artifact) and substantially more robust muscle attachment surfaces on the limb bones.
In any case, Boverisuchus appears to have been derived not directly from a more conventional semiaquatic ambush predator in the immediate aftermath of the Cretaceous–Palaeogene mass extinction, but from something more like a smooth-fronted caiman. This may have been an ecological shift away from the water, or it may merely have been enhancement of the adaptations found in smaller deep-snouted crocodylians.
This question, and the others left unanswered in this paper, can only be addressed with improved sampling. Planocraniids must have been present at least as far back as the Campanian, and some of the Asian species not included in this study – especially those from the Zaysan Depression – should be included. But we must also fortify our sample of characters relevant to this part of the tree; the earliest planocraniids were probably not altirostral, and whether current data sets are capable of recognising planocraniids with a more conventional cranial morphology is doubtful. But the effort may yield benefits beyond the planocraniids – it may shed much-needed light on the origins of Crocodylia itself.
4. Acknowledgments
For access to collections, I am indebted to P. Barrett, M. Brett-Surman, D. Brinkmann, M. Carrano, S. Chapman, T. Culver, F. de Lapparent de Broin, L. Del Favero, J. Eberle, M. Fornasiero, E. Frey, J. Gauthier, G. Gunnell, M. Hellmund, P. Holroyd, Y. Laurent, J. Li, P. Makovicky, S. McLeod, C. Mehling, R. Molnar, L. Murry, M. Norell, O. Rauhut, O. Rieppel, F. Schrenk, P. Willis, and X. Xing. I am also grateful to M. Delfino, W. Bartels, S. Salisbury, T. Rossmann, and P. Willis for discussion and information. Careful reviews by M. Delfino, B. Schubert, and members of the UI vertebrate palaeo community greatly improved this manuscript. The debt of gratitude I owe Wann Langston can never be overestimated. This work was supported by the US National Science Foundation (NSF DEB 0444133 and DEB 028648 to CAB) and the University of Iowa Obermann Center for Advanced Studies.
5. Appendices
5.1. Appendix 1. Character list and matrix used in phylogenetic analysis
The number in square brackets following the character number indicates the original character number in the analyses of Brochu (Reference Brochu1997, Reference Brochu1999).
(1) [1]: Ventral tubercle of proatlas more than one-half (0) or no more than one half (1) the width of the dorsal crest.
(2) [2]: Fused proatlas boomerang-shaped (0) strap-shaped (1) or massive and block-shaped (2).
(3) [10]: Proatlas with prominent anterior process (0) or lacks anterior process (1).
(4) [17]: Proatlas has tall dorsal keel (0) or lacks tall dorsal keel; dorsal side smooth (1).
(5) [5]: Atlas intercentrum wedge-shaped in lateral view, with insignificant parapophyseal processes (0) or plate-shaped in lateral view, with prominent parapophyseal processes at maturity (1). (Modified from Clark Reference Clark, Fraser and Sues1994, character 89.)
(6) [14]: Dorsal margin of atlantal rib generally smooth with modest dorsal process (0) or with prominent process (1).
(7) [16]: Atlantal ribs without (0) or with (1) very thin medial laminae at anterior end.
(8) [15]: Atlantal ribs lack (0) or possess (1) large articular facets at anterior ends for each other.
(9) [20]: Axial rib tuberculum wide, with broad dorsal tip (0) or narrow, with acute dorsal tip (1).
(10) [21]: Axial rib tuberculum contacts diapophysis late in ontogeny, if at all (0) or early in ontogeny (1).
(11) [11]: Anterior half of axis neural spine oriented horizontally (0) or slopes anteriorly (1).
(12) [12]: Axis neural spine crested (0) or not crested (1).
(13) [3]: Posterior half of axis neural spine wide (0) or narrow (1).
(14) [4]: Axis neural arch lacks (0) or possesses (1) a lateral process (diapophysis). (Adapted from Norell Reference Norell1989, character 7.)
(15) [6]: Axial hypapophysis located toward the center of centrum (0) or toward the anterior end of centrum (1).
(16) [19]: Axial hypapophysis without (0) or with (1) deep fork.
(17) [7]: Hypapophyseal keels present on eleventh vertebra behind atlas (0) twelfth vertebra behind atlas (1) or tenth vertebra behind atlas (2).
(18) [8]: Third cervical vertebra (first postaxial) with prominent hypapophysis (0) or lacks prominent hypapophysis (1). (Adapted from Norell Reference Norell1989, character 12; Norell & Clark Reference Norell and Clark1990, character 11; Clark Reference Clark, Fraser and Sues1994, character 91.)
(19) [9]: Neural spine on third cervical long, dorsal tip at least half the length of the centrum without the cotyle (0) or short, dorsal tip acute and less than half the length of the centrum without the cotyle (1).
(20): Cervical and anterior dorsal centra lack (0) or bear (1) deep pits on the ventral surface of the centrum.
(21) [18]: Presacral centra amphicoelous (0) or procoelous (1). (Adapted from several previous analyses, e.g. Benton & Clark Reference Benton, Clark and Benton1988; Norell & Clark Reference Norell and Clark1990, characters 8 and 10; Clark Reference Clark, Fraser and Sues1994, characters 92 and 93.)
(22) [13]: Anterior sacral rib capitulum projects far anteriorly of tuberculum and is broadly visible in dorsal view (0) or anterior margins of tuberculum and capitulum nearly in same plane, and capitulum largely obscured dorsally (1).
(23) [22]: Scapular blade flares dorsally at maturity (0) or sides of scapular blade subparallel; minimal dorsal flare at maturity (1). (Adapted from Benton & Clark Reference Benton, Clark and Benton1988.)
(24) [23]: Deltoid crest of scapula very thin at maturity, with sharp margin (0) or very wide at maturity, with broad margin (1).
(25) [24]: Scapulocoracoid synchondrosis closes very late in ontogeny (0) or relatively early in ontogeny (1).
(26) [25]: Scapulocoracoid facet anterior to glenoid fossa uniformly narrow (0) or broad immediately anterior to glenoid fossa, and tapering anteriorly (1).
(27) [26]: Proximal edge of deltopectoral crest emerges smoothly from proximal end of humerus and is not obviously concave (0) or emerges abruptly from proximal end of humerus and is obviously concave (1).
(28) [29]: M. teres major and M. dorsalis scapulae insert separately on humerus; scars can be distinguished dorsal to deltopectoral crest (0) or insert with common tendon; single insertion scar (1).
(29) [27]: Olecranon process of ulna narrow and subangular (0) or wide and rounded (1).
(30): Distal extremity of ulna expanded transversely with respect to long axis of bone; maximum width equivalent to that of proximal extremity (0) or proximal extremity considerably wider than distal extremity (1). (Salisbury et al. Reference Salisbury, Molnar, Frey and Willis2006, character 173.)
(31) [30]: Interclavicle flat along length, without dorsoventral flexure (0) or with moderate dorsoventral flexure (1) or with severe dorsoventral flexure (2).
(32) [31]: Anterior end of interclavicle flat (0) or rodlike (1).
(33) [34]: Iliac anterior process prominent (0) or virtually absent (1). (Adapted from Benton & Clark Reference Benton, Clark and Benton1988; Clark Reference Clark, Fraser and Sues1994, character 84; although transformation here is different.)
(34) [28]: Dorsal margin of iliac blade rounded with smooth border (0) or rounded, with modest dorsal indentation (1) or rounded, with strong dorsal indentation (wasp-waisted; 2) or narrow, with dorsal indentation (3) or rounded with smooth border; posterior tip of blade very deep (4).
(35) [32]: Supraacetabular crest narrow (0) or broad (1).
(36) [33]: Limb bones relatively robust, and hindlimb much longer than forelimb at maturity (0) or limb bones very long and slender (1).
(37) [160]: M. caudofemoralis with single head (0) or with double head (1).
(38) [35]: Dorsal osteoderms not keeled (0) or keeled (1). (Adapted from Buscalioni et al. Reference Buscalioni, Sanz and Casanovas1992, character 22.)
(39) [36]: Dorsal midline osteoderms rectangular (0) or nearly square (1). (Adapted from Norell & Clark Reference Norell and Clark1990, character 16; Clark Reference Clark, Fraser and Sues1994, character 95.)
(40) [37]: Four (0) six (1) eight (2) or ten (3) contiguous dorsal osteoderms per row at maturity (adapted from Norell & Clark Reference Norell and Clark1990, character 12; Clark Reference Clark, Fraser and Sues1994, character 97.)
(41) [38]: Nuchal shield grades continuously into dorsal shield (0) or differentiated from dorsal shield; four nuchal osteoderms (1) or differentiated from dorsal shield; six nuchal osteoderms with four central and two lateral (2) or differentiated from dorsal shield; eight nuchal osteoderms in two parallel rows (3).
(42) [39]: Ventral armor absent (0) or single ventral osteoderms (1) or paired ventral ossifications that suture together (2). (Adapted from Buscalioni et al. Reference Buscalioni, Sanz and Casanovas1992, character 21.)
(43) [40]: Anterior margin of dorsal midline osteoderms with anterior process (0) or smooth, without process (1). (Adapted from Norell & Clark Reference Norell and Clark1990, character 13; Clark Reference Clark, Fraser and Sues1994, character 96.)
(44) [155]: Ventral scales have (0) or lack (1) follicle gland pores (Poe Reference Bona1997.)
(45) [156]: Ventral collar scales not enlarged relative to other ventral scales (0) or in a single enlarged row (1) or in two parallel enlarged rows (2). (Poe Reference Bona1997.)
(46) [157]: Median pelvic keel scales form two parallel rows along most of tail length (0) or form single row along tail (1) or merge with lateral keel scales (2). (Poe Reference Bona1997.)
(47) [52]: Alveoli for dentary teeth 3 and 4 nearly same size and confluent (0) or fourth alveolus larger than third and alveoli are separated (1).
(48) [53]: Anterior dentary teeth strongly procumbent (0) or project anterodorsally (1).
(49): Dentary symphysis extends to fourth or fifth alveolus (0) or sixth through eighth alveolus (1) or behind eighth alveolus (2). (Modified from Brochu Reference Brochu2004, character 166.)
(50) [68]: Dentary gently curved (0), deeply curved (1), or linear (2) between fourth and tenth alveoli.
(51): Largest dentary alveolus immediately caudal to fourth is (0) 13 or 14, (1) 13 or 14 and a series behind it, (2) 11 or 12, or (3) no differentiation, or (4) behind 14. (Modified from Brochu Reference Brochu2004, character 167.)
(52) [41]: Splenial with anterior perforation for mandibular ramus of cranial nerve V (0) or lacks anterior perforation for mandibular ramus of cranial nerve V (1). (Adapted in part from Norell Reference Norell1988, character 15; Norell Reference Norell1989, character 8.)
(53) [42]: Mandibular ramus of cranial nerve V exits splenial anteriorly only (0) or splenial has singular perforation for mandibular ramus of cranial nerve V posteriorly (1) or splenial has double perforation for mandibular ramus of cranial nerve V posteriorly (2). (Adapted in part from Norell Reference Norell1988, character 15; Norell Reference Norell1989, character 8.)
(54) [43]: Splenial participates in mandibular symphysis; splenial symphysis adjacent to no more than five dentary alveoli (0) or splenial excluded from mandibular symphysis; anterior tip of splenial passes ventral to Meckelian groove (1) or splenial excluded from mandibular symphysis; anterior tip of splenial passes dorsal to Meckelian groove (2) or deep splenial symphysis, longer than five dentary alveoli; splenial forms wide V within symphysis (3) or deep splenial symphysis, longer than five dentary alveoli; splenial constricted within symphysis and forms narrow V (4). (Adapted from Clark Reference Clark, Fraser and Sues1994, character 77.)
(55) [46]: Coronoid bounds posterior half of foramen intermandibularis medius (0) or completely surrounds foramen intermandibularis medius at maturity (1) or obliterates foramen intermandibularis medius at maturity (2). (Adapted from Norell Reference Norell1988, character 12.)
(56) [54]: Superior edge of coronoid slopes strongly anteriorly (0) or almost horizontal (1).
(57) [55]: Inferior process of coronoid laps strongly over inner surface of Meckelian fossa (0) or remains largely on medial surface of mandible (1).
(58) [56]: Coronoid imperforate (0) or with perforation posterior to foramen intermandibularis medius (1).
(59) [59]: Process of splenial separates angular and coronoid (0) or no splenial process between angular and coronoid (1).
(60) [47]: Angular–surangular suture contacts external mandibular fenestra at posterior angle at maturity (0) or passes broadly along ventral margin of external mandibular fenestra late in ontogeny (1). (Adapted from Norell Reference Norell1988, character 40.)
(61) [48]: Anterior processes of surangular unequal (0) or subequal to equal (1).
(62) [61]: Surangular with spur bordering the dentary toothrow lingually for at least one alveolus length (0) or lacking such spur (1).
(63) [62]: External mandibular fenestra absent (0) or present as narrow slit, no discrete fenestral concavity on angular dorsal margin (1) or present with discrete concavity on angular dorsal argin (2) or present and very large; most of foramen intermandibularis caudalis visible in lateral view (3). (Clark Reference Clark, Fraser and Sues1994, character 75; incorporates information from Norell Reference Norell1988, character 14.)
(64) [65]: Surangular–dentary suture intersects external mandibular fenestra anterior to posterodorsal corner (0) or at posterodorsal corner (1).
(65) [66]: Angular extends dorsally toward or beyond anterior end of foramen intermandibularis caudalis; anterior tip acute (0) or does not extend dorsally beyond anterior end of foramen intermandibularis caudalis; anterior tip very blunt (1).
(66) [67]: Surangular–angular suture lingually meets articular at ventral tip (0) or dorsal to tip (1).
(67) [106]: Surangular continues to dorsal tip of lateral wall of glenoid fossa (0) or truncated and not continuing dorsally (1).
(68) [44]: Articular-surangular suture simple (0) or articular bears anterior lamina dorsal to lingual foramen (1) or articular bears anterior lamina ventral to lingual foramen (2) or bears laminae above and below foramen (3).
(69) [45]: Lingual foramen for articular artery and alveolar nerve perforates surangular entirely (0) or perforates surangular/angular suture (1).
(70) [49]: Foramen aerum at extreme lingual margin of retroarticular process (0) or set in from margin of retroarticular process (1). (Adapted from Norell Reference Norell1988, character 16.)
(71) [50]: Retroarticular process projects posteriorly (0) or projects posterodorsally (1). (Adapted from Benton & Clark Reference Benton, Clark and Benton1988; Clark Reference Clark, Fraser and Sues1994, character 71; Norell & Clark Reference Norell and Clark1990, character 7.)
(72) [51]: Surangular extends to posterior end of retroarticular process (0) or pinched off anterior to tip of retroarticular process (1). (Adapted from Norell Reference Norell1988, character 42.)
(73) [162]: Surangular–articular suture oriented anteroposteriorly (0) or bowed strongly laterally (1) within glenoid fossa.
(74) [60]: Sulcus between articular and surangular (0) or articular flush against surangular (1).
(75) [57]: Dorsal projection of hyoid cornu flat (0) or rodlike (1).
(76) [58]: Dorsal projection of hyoid cornu narrow, with parallel sides (0) or flared (1).
(77) [158]: Lingual osmoregulatory pores small (0) or large (1).
(78) [159]: Tongue with (0) or without (1) keratinised surface.
(79) [166]: Teeth and alveoli of maxilla and/or dentary circular in cross-section (0), or posterior teeth laterally compressed (1), or all teeth compressed (2). (Modified from Brochu Reference Brochu2004, character 165.)
(80): Maxillary and dentary teeth with smooth carinae (0) or serrated (1).
(81) [79]: Naris projects anterodorsally (0) or dorsally (1).
(82) [95]: External naris bisected by nasals (0) or nasals contact external naris, but do not bisect it (1) or nasals excluded, at least externally, from naris; nasals and premaxillae still in contact (2) or nasals and premaxillae not in contact (3). (Adapted from Norell Reference Norell1988, character 3; Clark Reference Clark, Fraser and Sues1994, characters 13 and 14.)
(83) [161]: Naris circular or keyhole-shaped (0) or wider than long (1) or anteroposteriorly long and prominently teardrop-shaped (2).
(84): External naris of reproductively mature males (0) remains similar to that of females or (1) develops bony excrescence (ghara).
(85): External naris (0) opens flush with dorsal surface of premaxillae or (1) circumscribed by thin crest.
(86) [142]: Premaxillary surface lateral to naris smooth (0) or with deep notch lateral to naris (1).
(87) [97]: Premaxilla has five teeth (0) or four teeth (1) early in posthatching ontogeny (Norell Reference Norell1988, character 17.)
(88) [124]: Incisive foramen small, less than half the greatest width of premaxillae (0) or large, more than half the greatest width of premaxillae (1) or large, and intersects premaxillary–maxillary suture (2).
(89) [153]: Incisive foramen completely situated far from premaxillary toothrow, at the level of the second or third alveolus (0) or abuts premaxillary toothrow (1) or projects between first premaxillary teeth (2).
(90) [145]: Dorsal premaxillary processes short, not extending beyond third maxillary alveolus (0) or long, extending beyond third maxillary alveolus (1).
(91) [77]: Dentary tooth 4 occludes in notch between premaxilla and maxilla early in ontogeny (0) or occludes in a pit between premaxilla and maxilla; no notch early in ontogeny (1). (Norell Reference Norell1988, character 29.)
(92) [78]: All dentary teeth occlude lingual to maxillary teeth (0) or occlusion pit between 7th and 8th maxillary teeth; all other dentary teeth occlude lingually (1) or dentary teeth occlude in line with maxillary toothrow (2). (Adapted from Norell Reference Norell1988, character 5; Willis Reference Willis1993, character 1; states modified from original.)
(93) [89]: Largest maxillary alveolus is #3 (0) #5 (1) #4 (2) #4 and #5 are same size (3) #6 (4) or maxillary teeth homodont (5), or maxillary alveoli gradually increase in diameter posteriorly toward penultimate alveolus (6). (Adapted from Norell Reference Norell1988, character 1.)
(94) [135]: Maxillary toothrow curved medially or linear (0) or curves laterally broadly (1) posterior to first six maxillary alveoli. (Adapted from Clark Reference Clark, Fraser and Sues1994, character 79.)
(95) [101]: Dorsal surface of rostrum curves smoothly (0) or bears medial dorsal boss (1).
(96) [143]: Canthi rostralii absent or very modest (0) or very prominent (1) at maturity (Norell Reference Norell1988, character 34.)
(97) [144]: Preorbital ridges absent or very modest (0) or very prominent (1) at maturity.
(98): Antorbital fenestra present (0) or absent (1). (Norell & Clark Reference Norell and Clark1990, character 2; Salisbury et al. Reference Salisbury, Molnar, Frey and Willis2006, character 176.)
(99) [125]: Vomer entirely obscured by premaxilla and maxilla (0) or exposed on palate at premaxillary–maxillary suture (1). (Adapted from Norell Reference Norell1988, character 22.)
(100) [126]: Vomer entirely obscured by maxillae and palatines (0) or exposed on palate between palatines (1).
(101) [148]: Surface of maxilla within narial canal imperforate (0) or with a linear array of pits (1).
(102) [120]: Medial jugal foramen small (0) or very large (1).
(103) [111]: Maxillary foramen for palatine ramus of cranial nerve V small or not present (0) or very large (1).
(104) [91]: Ectopterygoid abuts maxillary tooth row (0) or maxilla broadly separates ectopterygoid from maxillary tooth row (1). (Norell Reference Norell1988, character 19.)
(105): Maxilla terminates in palatal view anterior to lower temporal bar (0) or comprises part of the lower temporal bar (1).
(106): Penultimate maxillary alveolus less than (0) or more than (1) twice the diameter of the last maxillary alveolus.
(107): Prefrontal dorsal surface smooth adjacent to orbital rim (0) or bearing discrete knoblike processes (1).
(108) [137]: Dorsal half of prefrontal pillar narrow (0) or expanded anteroposteriorly (1). (Adapted from Norell Reference Norell1988, character 41.)
(109) [136]: Medial process of prefrontal pillar expanded dorsoventrally (0) or anteroposteriorly (1).
(110) [99]: Prefrontal pillar solid (0) or with large pneumatic recess (1). (State 1 describes prefrontal recess of Witmer Reference Witmer1997.)
(111) [138]: Medial process of prefrontal pillar wide (0) or constricted (1) at base.
(112) [105]: Maxilla has linear medial margin adjacent to suborbital fenestra (0) or bears broad shelf extending into fenestra, making lateral margin concave (1).
(113) [108]: Anterior face of palatine process rounded or pointed anteriorly (0) or notched anteriorly (1).
(114) [109]: Anterior ectopterygoid process tapers to a point (0) or forked (1).
(115) [110]: Palatine process extends (0) or does not extend (1) significantly beyond anterior end of suborbital fenestra. (Adapted from Willis Reference Willis1993, character 2.)
(116) [118]: Palatine process generally broad anteriorly (0) or in form of thin wedge (1).
(117) [94]: Lateral edges of palatines smooth anteriorly (0) or with lateral process projecting from palatines into suborbital fenestrae (1).
(118) [85]: Palatine–pterygoid suture nearly at (0) or far from (1) posterior angle of suborbital fenestra.
(119) [88]: Pterygoid ramus of ectopterygoid straight, posterolateral margin of suborbital fenestra linear (0) or ramus bowed, posterolateral margin of fenestra concave (1).
(120) [90]: Lateral edges of palatines parallel posteriorly (0) or flare posteriorly, producing shelf (1). (Adapted from Norell Reference Norell1988, character 2.)
(121) [71]: Anterior border of the choana is comprised of the palatines (0) or choana entirely surrounded by pterygoids (1). (Benton & Clark Reference Benton, Clark and Benton1988; Clark Reference Clark, Fraser and Sues1994, character 43; Norell & Clark Reference Norell and Clark1990, character 1.)
(122) [72]: Choana projects posteroventrally (0) or anteroventrally (1) at maturity.
(123) [73]: Pterygoid surface lateral and anterior to internal choana flush with choanal margin (0) or pushed inward anterolateral to choanal aperture (1) or pushed inward around choana to form neck surrounding aperture (2) or everted from flat surface to form neck surrounding aperture (3).
(124) [107]: Posterior rim of internal choana not deeply notched (0) or deeply notched (1).
(125) [152]: Internal choana not septate (0) or with septum that remains recessed within choana (1) or with septum that projects out of choana (2).
(126) [116]: Ectopterygoid–pterygoid flexure disappears during ontogeny (0) or remains throughout ontogeny (1).
(127) [149]: Ectopterygoid extends (0) or does not extend (1) to posterior tip of lateral pterygoid flange at maturity (adapted from Norell Reference Norell1988, character 32.)
(128) [93]: Lacrimal makes broad contact with nasal; no posterior process of maxilla (0) or maxilla with posterior process within lacrimal (1) or maxilla with posterior process between lacrimal and prefrontal (2).
(129) [100]: Prefrontals separated by frontals and nasals (0) or prefrontals meet medially (1). (Norell Reference Norell1988, character 27.)
(130) [117]: Lacrimal longer than prefrontal (0), or prefrontal longer than lacrimal (1), or lacrimal and prefrontal both elongate and nearly the same length (2). (Modified from Norell Reference Norell1988, character 7.)
(131): Anterior tip of frontal (0) forms simple acute point or (1) forms broad, complex sutural contact with the nasals.
(132) [133]: Ectopterygoid extends along medial face of postorbital bar (0) or stops abruptly ventral to postorbital bar (1).
(133) [70]: Postorbital bar massive (0) or slender (1). (Norell Reference Norell1989, character 3.)
(134) [134]: Postorbital bar bears process that is prominent, dorsoventrally broad, and divisible into two spines (0) or bears process that is short and generally not prominent (1). (Adapted from Norell Reference Norell1989, character 2.)
(135) [146]: Ventral margin of postorbital bar flush with lateral jugal surface (0) or inset from lateral jugal surface (1). (Adapted from Benton & Clark Reference Benton, Clark and Benton1988; Norell & Clark Reference Norell and Clark1990, character 3.)
(136): Postorbital bar continuous with anterolateral edge of skull table (0) or inset (1). (Norell & Clark Reference Norell and Clark1990, character 3; Salisbury et al. Reference Salisbury, Molnar, Frey and Willis2006, character 175.)
(137) [103]: Margin of orbit flush with skull surface (0) or dorsal edges of orbits upturned (1) or orbital margin telescoped (2).
(138) [139]: Ventral margin of orbit circular (0) or with prominent notch (1).
(139) [96]: Palpebral forms from single ossification (0) or from multiple ossifications (1). (Adapted from Norell Reference Norell1988, character 8; Clark Reference Clark, Fraser and Sues1994, character 65.)
(140) [69]: Quadratojugal spine prominent at maturity (0) or greatly reduced or absent at maturity (1). (Adapted from Norell Reference Norell1989, character 1.)
(141) [114]: Quadratojugal spine low, near posterior angle of infratemporal fenestra (0) or high, between posterior and superior angles of infratemporal fenestra (1).
(142) [75]: Quadratojugal forms posterior angle of infratemporal fenestra (0) or jugal forms posterior angle of infratemporal fenestra (1) or quadratojugal-jugal suture lies at posterior angle of infratemporal fenestra (2). (Adapted from Norell Reference Norell1989, character 10.)
(143) [76]: Postorbital neither contacts quadrate nor quadratojugal medially (0) or contacts quadratojugal, but not quadrate, medially (1) or contacts quadrate and quadratojugal at dorsal angle of infratemporal fenestra (2) or contacts quadratojugal with significant descending process (3).
(144) [83]: Quadratojugal bears long anterior process along lower temporal bar (0) or bears modest process, or none at all, along lower temporal bar (1).
(145) [80]: Quadratojugal extends to superior angle of infratemporal fenestra (0) or does not extend to superior angle of infratemporal fenestra; quadrate participates in fenestra (1). (Adapted from Buscalioni et al. Reference Buscalioni, Sanz and Casanovas1992, character 6.)
(146) [163]: Postorbital–squamosal suture oriented ventrally (0) or passes medially (1) ventral to skull table.
(147) [84]: Dorsal and ventral rims of squamosal groove for external ear valve musculature parallel (0) or squamosal groove flares anteriorly (1).
(148) [132]: Squamosal–quadrate suture extends dorsally along posterior margin of external auditory meatus (0) or extends only to posteroventral corner of external auditory meatus (1).
(149) [102]: Posterior margin of otic aperture smooth (0) or bowed (1).
(150) [81]: Frontoparietal suture deeply within supratemporal fenestra; frontal prevents broad contact between postorbital and parietal (0) or suture makes modest entry into supratemporal fenestra at maturity; postorbital and parietal in broad contact (1) or suture on skull table entirely (2).
(151) [86]: Frontoparietal suture concavoconvex (0) or linear (1) between supratemporal fenestrae.
(152) [87]: Supratemporal fenestra with fossa; dermal bones of skull roof do not overhang rim at maturity (0) or dermal bones of skull roof overhang rim of supratemporal fenestra near maturity (1) or supratemporal fenestra closes during ontogeny (2). (Adapted from Norell Reference Norell1988, character 9.)
(153) [92]: Shallow fossa at anteromedial corner of supratemporal fenestra (0) or no such fossa; anteromedial corner of supratemporal fenestra smooth (1).
(154) [104]: Medial parietal wall of supratemporal fenestra imperforate (0) or bearing foramina (1). (Norell Reference Norell1988, character 51.)
(155) [131]: Parietal and squamosal widely separated by quadrate on posterior wall of supratemporal fenestra (0) or parietal and squamosal approach each other on posterior wall of supratemporal fenestra without actually making contact (1) or parietal and squamosal meet along posterior wall of supratemporal fenestra (2).
(156) [123]: Skull table surface slopes ventrally from sagittal axis (0) or planar (1) at maturity.
(157): Posterolateral margin of squamosal horizontal or nearly so (0) or upturned to form a discrete horn (1).
(158) [140]: Mature skull table with broad curvature; short posterolateral squamosal rami along paroccipital process (0) or with nearly horizontal sides; significant posterolateral squamosal rami along paroccipital process (1).
(159) [150]: Squamosal does not extend (0) or extends (1) ventrolaterally to lateral extent of paraoccipital process.
(160) [82]: Supraoccipital exposure on dorsal skull table small (0) absent (1) large (2) or large such that parietal is excluded from posterior edge of table (3). (Norell Reference Norell1988, character 11.)
(161) [164]: Anterior foramen for palatine ramus of cranial nerve VII ventrolateral (0) or ventral (1) to basisphenoid rostrum.
(162) [122]: Sulcus on anterior braincase wall lateral to basisphenoid rostrum (0) or braincase wall lateral to basisphenoid rostrum smooth; no sulcus (1).
(163) [129]: Basisphenoid not exposed extensively (0) or exposed extensively (1) on braincase wall anterior to trigeminal foramen. (Adapted from Norell Reference Norell1989, character 5.)
(164) [74]: Extensive exposure of prootic on external braincase wall (0) or prootic largely obscured by quadrate and laterosphenoid externally (1). (Adapted from Norell Reference Norell1989, character 5.)
(165) [115]: Laterosphenoid bridge comprised entirely of laterosphenoid (0) or with ascending process or palatine (1).
(166) [130]: Capitate process of laterosphenoid oriented laterally (0) or anteroposteriorly (1) toward midline.
(167) [154]: Parietal with recess communicating with pneumatic system (0) or solid, without recess (1).
(168) [127]: Significant ventral quadrate process on lateral braincase wall (0) or quadrate–pterygoid suture linear from basisphenoid exposure to trigeminal foramen (1).
(169) [128]: Lateral carotid foramen opens lateral (0) or dorsal (1) to basisphenoid at maturity.
(170): External surface of basioccipital ventral to occipital condyle oriented posteroventrally (0) or posteriorly (1) at maturity. (Modified from Hua & Jouve Reference Hua and Jouve2004, character 167; Salisbury et al. Reference Salisbury, Molnar, Frey and Willis2006, character 174.)
(171) [98]: Posterior pterygoid processes tall and prominent (0) or small and project posteroventrally (1) or small and project posteriorly (2).
(172) [113]: Basisphenoid thin (0) or anteroposteriorly wide (1) ventral to basioccipital.
(173) [119]: Basisphenoid not broadly exposed ventral to basioccipital at maturity; pterygoid short ventral to median eustachian opening (0) or basisphenoid exposed as broad sheet ventral to basioccipital at maturity; pterygoid tall ventral to median eustachian opening (1).
(174) [141]: Exoccipital with very prominent boss on paroccipital process; process lateral to cranioquadrate opening short (0) or exoccipital with small or no boss on paroccipital process; process lateral to cranioquadrate opening long (1).
(175) [147]: Lateral eustachian canals open dorsal (0) or lateral (1) to medial eustachian canal. (Adapted from Norell Reference Norell1988, character 46.)
(176) [151]: Exoccipitals terminate dorsal to basioccipital tubera (0) or send robust process ventrally and participate in basioccipital tubera (1) or send slender process ventrally to basioccipital tubera (2). (Adapted from Norell Reference Norell1988, character 20; Clark Reference Clark, Fraser and Sues1994, characters 57 and 60.)
(177) [121]: Quadrate foramen aerum on mediodorsal angle (0) or on dorsal surface (1) of quadrate.
(178) [165]: Quadrate foramen aereum is small (0), comparatively large (1), or absent (2) at maturity.
(179): Quadrate lacks (0) or bears (1) prominent, mediolaterally thin crest on dorsal surface of ramus.
(180): Attachment scar for posterior mandibular adductor muscle on ventral surface of quadrate ramus forms modest crests (0) or prominent knob (1). (Ösi et al. Reference Ösi, Clark and Weishampel2007, character 165.)
(181) [112]: Quadrate with small, ventrally-reflected medial hemicondyle (0) or with small medial hemicondyle; dorsal notch for foramen aerum (1) or with prominent dorsal projection between hemicondyles (2) or with expanded medial hemicondyle (3).
(182): Iris (0) greenish/yellowish or (1) brown.
(183): Two or more (0) or one (1) row of postoccipital osteoderms.
(184): Fewer than eight (0) or eight to 14 (1) or more than 14 (2) paired midline scale rows. (There is considerable variation in this character within species, and further work is needed to clarify the situation; data from Fuchs Reference Fuchs2006.)
(185): Ectopterygoid maxillary ramus forms less than (0) or more than (1) two-thirds of lateral margin of suborbital fenestra.
(186): Ectopterygoid maxillary ramus terminates at lateral margin of suborbital fenestra (0) or lateral to it, with maxilla separating the ectopterygoid from fenestra for short distance.
(187): Palatine–maxillary suture intersects suborbital fenestra at its anteromedial margin (0) or nearly at its anteriormost limit (1).
(188): Frontal lacks (0) or bears (1) prominent midsagittal crest between orbits.
(189): All cervical neural spines anteroposteriorly broad (0) or posterior neural spines thin and rod-like (1). (Modified from Clark Reference Clark, Fraser and Sues1994, character 90; Pol et al. Reference Pol, Turner and Norell2009, character 90.)
Bernissartia fagesii
??????0???0111102100000?0?000???0000?100010???0010?000???????10?0?00?001?1????000?0?0000?00030?001?????1000????1?0000?000??0100?0???000100?0?0??0?0?0010?0??00??0????????000?0000?000???00000
Allodaposuchus precedens
??????????????????????????????????????????????????????????????????????????????00010?0000001230000100?100000????000000010103000110000101100???0100000001100010000?????1??01?010001?000???0010?
Acynodon adriaticus
?????1?????????????01?100?1??????????010?10?????????01????????0???0??100??00??00010?000?0?10600001?0????110????00?00011010000?1?111?0?0100???0????0??010?0?0010??????????????1???01?0?????101
Acynodon iberoccitanus
??????????????????????????????????????????????10104101????????0???????0?0?????00010?0000001060000100??00100????0000000101?00???200100?0100?110?0000??010100000?1?????????1???0????010???1010?
Iharkutosuchus makadii
??????????????????????????????????????????????10124?????????110???00??10?1????0001??00000110610001?0???0110????00000010011000012111001?100?1?0?00?0???12???100?2?????1???1000000??110???1010?
Hylaeochampsa vectiana
????????????????????????????????????????????????????????????????????????0?????0??????????0?0?10001?00??0110?0?00000001001000?0?21110000120????1?0?000010100000000????1?001001000?0110?????10?
Borealosuchus threeensis
????0??????????????01???????0??????1?00??20???01002??1?????10?11?000001000??????????????????????????????????????????????????????????????????????????????????????????????????????????????????1
Borealosuchus formidabilis
000?000?0?11001001001000000101000001?000?20???0110200000?000?120000000100001??00020?0000?00231000100?00000010000000101001000101002001?11000000?00?000010100101000??????00100110000000???00001
Borealosuchus wilsoni
??????0??????????1001000000101??00?1?000?20???01002??100?0?001100000201000????00020?0?0???02310001?????00001?0?0?0010100100010100200101100?000100??001101001010000?00???010011?0?0000???00001
Borealosuchus acutidentatus
????????????????????????????0???????????????????002????????????000??????0?????00020?0?00??02310001?????0000????0?0??0???????1??0020?1?1100?0?0?0??0?011010?101000???????010??1?0?0000???0000?
Borealosuchus sternbergii
000000000?110010?1001000000101000001?00???0???011020000000?00020000000100000??00020?000000013100010001000001?0?000001110100010100000111100?000100?000000100101000000?1?00100110000000???00001
Eothoracosaurus mississippiensis
??????0????????????01??000????????00?00???0????122???3??????0?0???00?011?0????00120?000??1025000010??000000????0000101001000????000?000100???01???10000010?001?00????????100010000000???00001
Thoracosaurus neocesariensis
??????0??????111?1?010????0011??0??0?00???0???1122???3????????10?000?01?0?????00120?0000?10250000100??000000?0?00001010010000?000000000110?00010??1?000010000100000000?00100?10000000???00001
Thoracosaurus macrorhynchus
??????0???0?1111?1?01?????00????0????00???0???1?22???3??????0?100000001100????00120?00?0?10250000100??000000?0?000010100100001100000000110?000100?10001010?0010000???0??0100010000000???0000?
Eosuchus minor
??????0???0??111???01?00?0?01???0000?000??0???1122??0300?0000?20?000001100????00120?0000?1025?000100?000000?00000001010010000?100000000110???0100?10?100100001?0100??1???101010101003???00001
Eosuchus lerichei
??????0??????01????01???????1????????0????????1122???3??????????????????0?????00120?0000?10250000100?0000000???0000101001000001000000?0110?000?0??10?2001000010?1????????10?01?101003???00001
Eogavialis africanus
????????1?????11???010??????????0?????????0???1122??03?????10?200000??1101????00120?000?01025?000100?0000000000000010100100000100000000121?000100?1001001000010100?000??0101010100000???00?01
Gryposuchus colombianus
????0?0???001??????01??000????????????????0???11223?030100?000200000001100????0012010000?1025?000100?0?0000??0?000010100100000110000000121?000100?10011010000200000100?00121010100000???00101
Ikanogavialis gameroi
??????????????????????????????????????????????11223??3??????0?20?0?0?0???0????00020?0000?1025?0001?????0000?????0?01???010?????1000?000121?????0?????210?00002000???????012101010?000?????10?
Siwalik Gavialis
???????????????????010??????????????????????????22???3??????0?200000??1?0?????00?30?0??????2??0??1?0?0?0000??0?0000?0?00100?0?1???00000121?000100?100??0?000010000?????00121010100000???00101
Gavialis bengawanicus
?????????????111????1????????????????1????????11?2?003?0???000200000101100????0?13??0????10250000100?0000000???01?010???10?00??10000000121?000100010011010010100???0?0?001?101010?000???0010?
Gavialis gangeticus
020000000?001111011010000000111000000?0000000011223003000000002000000011000100001301000001025?00010000000000000000010100100000110000000121000010001001101000010000000000012101010000000100101
Boverisuchus vorax
????0?0???01001001?01?00000111??0100?10???1???1110?000?????0??2000001?1?0?????21010?0000?00030001100?000000??0?000000100100010100000111110???0100?010000100101000???01??1100110000002???00001
Boverisuchus geiseltalensis
?????????????0?0???01?000?0??1??0100?1???1????11102000??????0?2????0??1?1?????21?20?00000000300011???000000????0000001001010??100000111110?0?0?00?0?010010?10100?????????1???10000002???00001
Planocrania hengdongensis
????????????????????1?????????????????????????1110???1????????{12}????0?01?0?????20010?0?????01300??1????0?000????0????01001???????0?0?1?1100????????0?0?00100?0100?????1???1???10?10001???0000?
Planocrania datangensis
??????????????????????????????????????????????11?????0????????????????????????20010?000??10030000100??0?000???????000??01???????0?0?1?1110????????0?000010010??0?????????????10??0?0????0000?
Leidyosuchus canadensis
????0?0???????1????010000011?1??10?0?11??11???0110?00000?0?01?200000011101????00010?000000003000010000010001000100001110100010100010111100?010100100100010010100000001?00100110010001???00001
Diplocynodon ratelii
??????0??????000???010?00?1111001400?10??21???01002101?????0112000001?1101????00120?00000012300001000101000??0?000000111100010100010111100??10100100000010110100000001?10100110010001???00001
Diplocynodon hantoniensis
100???1?1?01000010001000011111??1400?101?21???011021010????011200000111101????00120?0?000?11300?0100?1010001?0?000000101100010100010111100?110100?001010101101000??0?1??010011?010001???00001
Diplocynodon darwini
100001001?010000?00010000?1111??1400?101121???011020010??0?01?20000011110100??00020?0000?01030000100?0??000??0?0000001111?001010001?111100?110100?011010101101000???????0100110010001???00001
Baryphracta deponiae
100?0?0???????0????01?0???1?????14?0?10??21????1?02??0?????01?20??0??1110?????001???000??010300001?0????000?????0?0001?11000?010001?111100?110100?0??011?0?101?00????????1???1??10001????0001
Stangerochampsa mccabei
????110???010010?0001000001111001000?01??11???111010100????0112110000?110?????00110?0002?11020000100?001000????000000011110010120011111100?110200?0?1100102101000??1?1??0100110010001???00001
Albertochampsa langstoni
??????????????????????????????????????????????????????????????????????????????001?0?0?0??110200001?0??01?00????00??00011110010?2001?111100???0?0??0?110010210100????????010011001?000?????00?
Brachychampsa montana
101011001?1100???0001??000111100?000?103111???11101101?????01120?00001110100??00110?0002?11010000100?0010001?0?010000011110010120011111100?110200?01110010110102000101?00100110010001???00001
Brachychampsa sealeyi
?????????????????????????????????????10???1???11101??0???????12??0000111?1????001?0?0002?1101000010???010?????????00????????10?200?1101?00???????????100???????2?????????????1??10??1??????0?
Alligator sinensis
101011001?01001000001011011111001100011210110011000112010010113000101111010011001000010000102000010000010001110000000011110010110111111110011020010112101021010100011110010011001000100100001
Alligator mississippiensis
101011001?01001000001011011111001100011210110011000112010010113000101111010011001000010000102000010000010001110000000011110010110111111110011020010112101021010100011110010011001000100100001
Alligator mefferdi
???????????????????????????1?????????11???????110000120100001130001011110100??00100?0100?01020000100?0?10001?1?0000000?1110010110111111110?110200?011210102101010??1???00100110010001???0000?
Alligator thomsoni
???????????????????01????????1?????0?1????????110000?2?????011300010011101????00100?010000102000010??0010001?1?000000????1?????10111111110?1?0200101?21010210101?001?11?0100110010001???0000?
Alligator olseni
??????0?1?????10???01?10011111???100?11???????11010010?????011300000011101????00100?01000?10200??100???1000??0?0?0001011111010110111111110?110200?011200102101010??????00100?1?010001???00001
Alligator mcgrewi
100010001?010010?00010000111?1101??0?11???1???11110010?100?011200000111101????00000?0100?01020000100?0010001?001000000111100101101111111000110200?011200102101010?01???00100110010001???00001
Alligator prenasalis
10001?0?1?????10?0?01000011111??1000?112111???11111010?????01120000011110100??00000?0100?01020000100?0010001100100000011110010110111111100?110200?011200102101000001?1?00100110010001???00001
Ceratosuchus burdoshi
???????????????????????????????????0?1????????11111??0?????01?20??00?11?01????00010?0?01?01020000100???1000??0????00???01???1?1??0??111100????????0????010?111???????????10??10010001??????0?
Hassiacosuchus haupti
001?1?0?1??????0???01?000?1111?????0??11?11???111110?0?????01?20??0??11101????00010?00???010?00001??????0001?0??????0?????????11001?111100?1?0200?01120010210101????????01???10010001???00001
Allognathosuchus polyodon
??????????????????????????????????????????????11111010?????01?21??00?111?1????00010?000??010200001?0?????00??0?00?00?0?111001?1?011?111100????????0??20010?101???????????1???1??1???1??????0?
Allognathosuchus wartheni
????1?0????????????0?0000?1111??1000?11???1???11111010?100?011210000011101????00010?0000?0102000010000010001?0?000000?0111001?110111111100?110200?0112001021010000?1?1?0010011001?001???00001
Navajosuchus mooki
??????0?1??????0???0?00???1111??1??0?111111???11111010?????01?20??00??110?????00010?0001?010200001???0?1000??0???00?00?1110010110011111100???020??0112?0102101000????1??010??10010001???00001
Wannaganosuchus brachymanus
????1?0???1?00?0???010000?1111001000?11???1???111110?0?????0??200?00?11?0?????00110?0000??10200??100???1000??0?100000???1??01?11011?111100????????0?12001??101000????????10011?010001???00001
Procaimanoidea kayi
????110?1??????0???010?00?1111??10?0?112121?????010?1??100?01?21000001110?????10?????0???010?00001???0?1000??0?0000000011100?01101111111000??0200?011200102101010???????0100110010001??????01
Procaimanoidea utahensis
??????????????????????????????????????????????110100?00??01011210??0011101????10110?0100?01020?00100??01000????0000000?111001011011?111100?1?0200?01120010?101000???????0100110010001???0000?
Arambourgia gaudryi
??????????????????????????????????????????????11010??0?????01?200?0??1110?????1001??010??010200001?0???10001000??000????11?0??11011?111100?1102?0?0?1210?0210100?????????10011?010?01??????0?
Necrosuchus ionensis
??????0????????????0???01?111???1300?11???1???1100???2????????2???0?????0?????00????????????????????????????????????????????????????????????????????????????????0??????????0????1?0?1???????1
Tsoabichi greenriverensis
???????????????????01????????????????10??20???1100???2????????2??????11???01??00010?10????10??0001????????0????????????????????0001?1111?0?1?0????0??211???101?2????????????????100????????0?
Purussaurus mirandae
????????????????????1?11001?????11?0?1????????1100?0?2??1?11?1301100?11?01????0?110?0001101020010100???1000????0000000111101?11111??111110?1102001011201???10102??????????0011?2?00?1??????0?
Purussaurus neivensis
101?100?1?000010?0??1????011???????0?11???1???1?00??1?1010?111301100011001????00110?0001?01020000100?0010001?0?0?00000111101211111?1111110?110201?0112011?210102000101??010??10210001???00?0?
Orthogenysuchus olseni
????????????????????????????????????????????????00??????????????????????0?????00121?0?01??10?0?0?1??????000????01?000????????????????11110????????0????11??101??0???????????????10001??????0?
Mourasuchus atopus
10??100?1?00?010?00?1?10?011????1300?11???1???1102?112?????01120?100011000????00121?0000?11050001100?0010001?0?01000001111012?1011??111110?110?00????2111??111?30????1???100110?10001???00?0?
Eocaiman cavernensis
??????????????????????????????????????????????1110???2?????11?2???????????????00????0??????0?????1?????100???0?1??0000???????0??????1??1?0?????0???????0?????1?3??????????0??1?2???????????0?
Caiman yacare
101111001?10001000001010111111001100011122111011002112101011102011020110010111001100000000112000010000010001100100000011110121111011111110011020110112011121010300010110010011021000100200001
Caiman crocodilus
101111001?1000100000101011111100110001112211101100211210101110201102011001011100110000000011200001000001000110010000001111012111(01)011111110011020110112011121010300010110010011021000100200001
Caiman latirostris
101110001?10001000001010?111110011000111221210110021121010111?20110201100???11001100000000102001010000010001100100000011110121110011111110011020110112111121010300010110010011021000100200001
Caiman lutescens
????????????????????????????????????????????????0?????????????????????????????00110?0000?01020010100?001000????1000000111101211200??1?1?10?????0???????1????0???????????????????100?1???0000?
Melanosuchus fisheri
??????0????????0?????????????????????????????????02????????1?12011????10?1????001?0?0000??1020010110???1000??0???0????11????2?11????111110????????0????11??1010?????????010??1?2?0001???0000?
Melanosuchus niger
101111001?1?00100000101011111100110001112212101100211210101111201102011001??11001100000000102001011000010001100100000011110121110011111110011020110112111121010300010110010011021000100200001
Paleosuchus trigonatus
100111111?01001010001000111111211300011132112011002122211111112011000110010111101100001000102000010000010001100010001011110111100011111110111020110112111?21010200010110010011021000110100001
Paleosuchus palpebrosus
100111111?010010101010001111112113000111321120110021222111?11?20110001100?0111101100001000102000010000010001100010001011110111100011111110111020110112111?21010200010110010011021000111100001
Mecistops cataphractus
10?001001?000010000011100111112012000111101101111041010100010020001110101?1001001200000010021000010001000001101000010110100010100000111110000101110012001001010001110101111001000000300?11101
Crocodylus niloticus
101000001?101010001011100111112012000111201101110021010100010120001110101110010011000000100210000100110000(01)110100(01)000110100010110000111110000101110012001001010001110101111001100000301100001
Crocodylus porosus
111000001?0010101010111000111120120001112011011100210101000101200011101011100100110000001002100011001100000110100(01)000110100010100000111110000101110012001001010001110101111001100000301100001
Crocodylus rhombifer
001000001?1010100010111001111120110001112011011100210101000101200011101011100100110000001002101001001100000110100(01)000110100010110000111110000101110012001001110001110101111001100000301200001
Euthecodon arambourgii
?????????????????????????????????????????????????????0??????????????????1?????00020?0000?10250001100?1000001101000000??0????0???0000111110??11011?001210100101100????1??11?0?1000?003???0000?
Osteolaemus tetraspis
??1?00001?00101010001110011111201110011111110111002101010001012000011010111001101000001010021000110001000001101100101110101010100010111110100101010012111001011001110101111011000000311000001
Osteolaemus osborni
??1?00001?00101010001110011111201110011111110111002101010000012000011010111001101100001010021000110001000001101100100100101010100000111110100101010012111001011001110101111011000000310000001
Voay robustus
??????0????????0???011?????111??1110??????1???1110210101000001200011111011????00110?0000?0021000110001000001101000101110101010100000111110?0010111001201100111110?1101?11110010000003???00001
Rimasuchus lloydi
?????????????????????????????????????????????????????1??????????????????1?????00110?0000?00210001100?100000110100000????10??1??1000?111110?001011?001200100101100111?1??1110?1000?003?????00?
Crocodylus pigotti
??????????00?010?00?11??01111????????10???0???11102??1????????2???111???1?????00010?0100?002100011000100000????0010101101010001?0?00111110???1?1??001210100101100??1?1??1100?10000003???0000?
Crocodylus megarhinus
??????0???????????001?????????????????????????11102101?????00120000??01011????00110?0000?0023000010001?0000??0?000000110100010120000111110?002?11?0012?01001010?01100???1110010000003???0000?
Australosuchus clarkae
??????0???????1??0??1?????11???????0?10???1???1110?101?????001200001101011????00110?000011021000010001000001?0?000000???????101000?0111110?002011?01120010010100011??1??11?001000?001???0000?
Kambara implexidens
??????0????????????01?????11????1100?10???1???11102101?????001200001101011????00110?000011021000010001000001?01000000100100010100000111110?002011?00120010010100011101?11110010000001???00001
Trilophosuchus rackhami
??????????????????????????????????????????????????????????????????????????????0?????0??????2?0??0??0?1000????0?1?000001010???01?0??0111110??12011101121010000102011101?111?0?1000?001??????0?
Tomistoma schlegelii
021000001?0010100010110001111110110001013011011122?104000001002000000010100001001200000011021000010101000001101000100100100010110000111110000011010012101001010001100101111001000000311100001
Tomistoma lusitanica
??????0????????????01????????????????10???1???1??2???4?????00?20000??0101?????00120?0000?1021?000101?100000110?000010100100010100000111110?0001?1?10121010010100011001??1110010000003???00001
Tomistoma petrolica
???????????????????????????1????????????????????????04??????0020?0?01?1?10????00???????????2?00?0??0???00001???0?00101001??????1?00?0?1110?0?2?0??0??200?0000100?????????1???1????003???0000?
Toyotamaphimaea machikanensis
00100100??11111100101100011111101100?00???1?????22??04?????10020?0000?101000??00120?000011004000010???00000????0000?010010000?100000011110??00?1??1??21010?1010??????????110010?00003???00001
Penghusuchus pani
00000011100010??10101??101111?1?0100?00???1???1?22?104?????00?20?000101?0?00??0?????0??????2500001?0??000001111000010000101000110200011100???????1???2?????1010??????0??011?010010003???0000?
Gavialosuchus eggenbergensis
????????????????????1?????????????????????????????????????????????????????????00120?0000?10240000100??00000????00?0101001000?012000?0?1110?0??????1?121010?10100????????11???10000003???0000?
Paratomistoma courtii
???????????????????????????????????????????????????????????0??20?000001010????00???????????2??0?0???????0001?0?????????????????0000?1??11?????1???1??200100001000?1000?101?0?100??00???????0?
Megadontosuchus arduini
???????????????????01????????????????1????????1120?????????0??20?????0???1????001?0?000?1102100001??????000???20???1????????????000?011110???0????1??2001??101???????????1???1????00????0000?
Tomistoma cairense
??????????????????????????????????????????????1??2???4????????2000001?1010????00?20?0?????025?0??1?0????000??0?000010100100010?00?0?011110???01???10?2001001010001??0???11?001?000003???0000?
Thecachampsa antiqua
02000000??001010?00011010111?1??11?0?00???1???1122???4?100????2000031?101?0???00120?0000110210000100?100000??0?0000101?0100010110000011110???011??101200100101010??00???11100100?2003???00001
Thecachampsa americana
??????????????1????01?000?1111??1100?00???1???1122???4?????00020?0031?1010????00120?0000110210000100?100000????0000100001000???1000?011110?000?10?10120010010101???0????11100100?2003???00001
Thecachampsa carolinense
????0000?????010?010110?01??????1?00?00???1???1122??04?????00020?0131?1010????00120?00001102100001????00000????0?00101001000?01?000?0?1110???0????1??20010?10101?????????1100100?2003???0000?
Dollosuchoides densmorei
001???0???111010?0001?0?001111??1?00??????????1120???0?????000200?0??0101?????00120?00?0?10210000100??00000????01?01010010001011000?011110????????1??20010?1010001???????10?01000?003???00001
Kentisuchus spenceri
??????0????????????????????????????????????????1?????0?????0??2000?1101?11????00110?0000?10210000100???0000??0?010010100100?1011000?111110??????0?0??20010?10100???????111?0?100?0?03???0000?
Brachyuranochampsa eversolei
???????????????????01????????????????1?????????11020100100000?20?000101011????00110?0000110210000100??00000??0?00010011010001010001?111100?????00?0?121010?101000???????1100010000003???0000?
Crocodylus acer
??????????????????????????????????????????????????????????????????????????????00110?0000?10210000100??000001???000100?10100010100010111100?002?01?00?2001001010001??0??11100010000003????000?
Asiatosuchus grangeri
??????????????????????????????????????????????1??0?101??????0?2000????101?????0???0????????1??0???????1?????????????????????????000???110????????????20?????0??????????????????????????????0?
Crocodylus depressifrons
???00?00??11000010011100001111??1100?10???1???11102101?????001200000001011????00110?0000?10110000100?1100001?0?0001001?01000101?0000111100?010200?00120010010100011001?11100010000003???00001
Crocodylus affinis
001001001?10001000011100001111001100?10???1???111021010100000120000000101100??00110?0000100110000100?010000??0?000100110100010100000111100?0?010??0012001001010001??0???1100010000003???00001
Asiatosuchus germanicus
001?0?0?1?001010?0101?000?1111??1??0??????1???11102000?????00120000??0101?00??00010?0000?0001000010???10000??0?00010???010001010000?111100???0100?001110100101000???????1100?10000003???00001
Prodiplocynodon langi
??????????????????????????????????????????????????????????????????????????????00110?0000??0030000100??100001??000010011?1000101???0?111100???0?0??0?11?010?1010001??01?01100110000003???0000?
5.2. Appendix 2. Planocraniid material used for coding characters in this study
Boverisuchus magnifrons – GM Ce IV 6044 (holotype), GM 10 128, GM VI 180, GM XVIII 3094, GM XVIII 3268, GM XVIII 486, GM XVIII 526, GM XVII 415, GM XXII 543, GM XXII 815, GM ce-II 5894, GM Leo X 8001 (holotype, Weigeltisuchus geiseltalensis), GM XXXV 127, GM XXXV 216, GM XXXV 94, GM XXXVI 245, GM XXXVI 274, GM XXXVI 499, GM XXXVII 111, GM XIV 616, HLMD Me 5346, HLMD Me 5609.
Boverisuchus vorax – YPM 249 (holotype), YPM 5890, UCMP 170767, USNM 12957, FMNH PR399, FMNH PR479, AMNH 2090, AMNH 29993.
Planocrania datangensis – IVPP V5016 (holotype).
Planocrania hengdongensis – IVPP V6079 (holotype).
6. Note added in proof
I recently had an opportunity to examine the holotype of Crocodilus bolcensis at the Musei Regionale di Scienze Naturali in Turin. Its preservation precludes identifying most cranial sutures, and so whether C. bolcensis is a distinct species or conspecific with Boverisuchus geiseltalensis remains unclear. However, the holotype does preserve serrated teeth, and several structures in the trunk region could be ventral osteoderms. I thank Massimo Delfino and Daniele Ormezzano for their hospitality and assistance during my visit.