

We were very pleased, and honoured, when asked to add ourselves to this tribute to Jenny Clack. A quick examination of the references for our contribution makes clear the fact that we could not have written this paper without leaning on Jenny's prolific work. Indeed, about 40% of our references boast Jenny as an author. Her efforts have made it possible for our own work to be better and we are immensely grateful for that. Vertebrate fossils can be difficult, as all of us know. Those tetrapods of the Upper Devonian and Lower Carboniferous that have been insulted by ca.380–330 million years of sedimentary deposition and jerked about by tectonic movements can be particularly difficult. Jenny continues to be a leading interpreter of these challenging relics and this is a testament to her depth of knowledge, technical skill, and bravery. We join our colleagues in offering our warmest congratulations, Jenny.
This paper is our fourth contribution toward a full description of Whatcheeria deltae Lombard & Bolt (Reference Lombard and Bolt1995), a Middle Mississippian (Viséan) tetrapod known from a single locality in SE Iowa. Lombard & Bolt (Reference Lombard and Bolt1995) summarised the skeletal anatomy as known at that time, Bolt & Lombard (Reference Bolt, Lombard and Heatwole2000) discussed functional anatomy and ecology, and Lombard & Bolt (Reference Lombard, Bolt, Carrano, Gaudin, Blob and Wi2006) described the very primitive lower jaw, which, inter alia, retains an extensive Meckelian ossification. The description of the skull in our 1995 paper was mostly of the exterior view. In that and subsequent papers, less space was devoted to the palate, braincase, and occiput, despite their considerable importance and the fact that Whatcheeria is ‘represented by numerous and often excellently preserved specimens' (Lombard & Bolt Reference Lombard and Bolt1995, p. 472). This is indeed the case; but these less fully described elements are quite difficult to characterise in most specimens. The reason is twofold: first, there is the crushing, which has affected many specimens; and second, the skull is both narrow and tall – a fact that we will document further. The skulls of early tetrapods are generally preserved as dorso-ventrally compressed. This result is taken to reflect the assumption of a state of least energy at burial for a skull that is wider than tall. Whatcheeria shares the reverse configuration with Crassigyrinus. Both preserve as fossils with the skull laterally compressed, the palatal bones sandwiched between the right and left skull halves, and the skull table flared out flat. In Whatcheeria, this mode of preservation produced a number of specimens in which the palate can be partially glimpsed through an orbit, for example, and is inaccessible without destructive preparation. With time, however, we have been able to prepare out partial palates from individual skulls as well as some isolated, sometimes three-dimensional, palatal elements, and can now present a description of individual palatal bones and a reasonable composite reconstruction as restored to three-dimensionality.
1. Materials and methods
All Whatcheeria specimens come from two adjacent collapse structures of Viséan 3b (Asbian) age, in the disused Hiemstra Quarry in SE Iowa (Witzke et al. Reference Witzke, McKay, Bunker and Woodson1990; Lombard & Bolt Reference Lombard and Bolt1995). Snyder (Reference Snyder2006) provides an analysis of the sedimentology and palaeoenvironment of the Hiemstra Quarry that indicates a primarily lacustrine environment at the time of deposition and an age of about 330mybp. Most of the specimens studied are in the collections of the Field Museum of Natural History (institutional identification in this paper, PR); others are University of Iowa specimens (institutional identification, SUI).
Specimens occurred in alternating bands of shale and lime mudstone. Those in mudstone generally required little special treatment initially, other than use of a rock saw, hammers, and chisels to reduce the size of the enclosing block. Specimens in shale usually had to be strengthened by cyanoacrylate adhesive and further supported by a plaster jacket in the case of larger specimens. In the laboratory, the adhesive was removed as needed with acetone. Matrix removal was by miniature pneumatic tools and handheld pin vises, under a binocular microscope. Repair, and further consolidation as needed, was usually done with cyanoacrylate adhesive; larger gaps were filled with an epoxy.
In one case (PR 1888), a lime-mudstone block was mechanically prepared from both sides while leaving just enough matrix for support. This produced valuable information, although in most cases the ‘hidden' surface proved too damaged, or too fragile, to permit extensive further preparation. Blocks could usually not be completely prepared from each side; in these cases, a ‘window' was developed on one side in order to maintain the necessary supporting matrix. One specimen (PR 1636) was transfer-prepared, with one side mechanically prepared, then completely embedded in transparent epoxy resin; the other was prepared mechanically, and left open.
1.1. Reconstructions
The palatal reconstruction (Fig. 1) is primarily based on three specimens: PR 1814, an anterior palate (Fig. 2a); PR 1792, the palatal bones in approximate anterior–posterior register (Fig. 5); and PR 1701, a braincase in ventral view (Fig. 6). PR 1814 was used to reconstruct the general outline of the snout, the articulation of the pterygoids with the lateral palatal bones, and the articulation of those bones with the marginal premaxilla and maxilla. This specimen also provided for the reconstruction of the choana and its placement, as well as the general disposition of some palatal teeth. PR 1792 provided the general shape of the pterygoid, the location of the basipterygoid joint relative to the marginal palatal series, the relations of the marginal bones to the vomer, palatine, ectopterygoid series, and their teeth, and the placement of the adductor fossa. PR 1701 provided the positioning of the basipterygoid joints relative to the midline and the foramen magnum. Many specimens listed in the following section were informative about details or provided confirmation of features present on the most informative specimens used for reconstruction.
The occlusal reconstruction (Fig. 10) is primarily based on the palatal specimens above plus PR 1809 (jaw), a three-dimensional jaw tip (Lombard & Bolt Reference Lombard, Bolt, Carrano, Gaudin, Blob and Wi2006, fig. 2.2), as well as PR 1644, which preserves the natural curvature of the left jaw in the horizontal plane. The numerous jaw specimens listed in Lombard & Bolt (Reference Lombard, Bolt, Carrano, Gaudin, Blob and Wi2006) also provided information on the disposition of the teeth.
1.2. Whatcheeria specimens studied
PR 1634. Articulated skull, jaws, and some postcranial elements on lime mudstone. Skull laterally compressed exposing left side and skull roof. Dorsal surface of left pterygoid including basipterygoid joint, as well as cultriform process of braincase, visible through orbit (Lombard & Bolt Reference Lombard and Bolt1995, text-fig. 1).
PR 1644. Three-dimensional jaws, disarticulated skull pieces, and some postcranial elements in lime mudstone.
PR 1651. Skull roof prepared free. Visible in dorsal view are the tabulars, postparietals, parietals with large parietal opening, frontals, supratemporals, intertemporals, and postfrontals. In ventral view, the basiparasphenoid with both stapes in articulation is crushed up against the underside of the dermal elements.
PR 1652. Partial skull roof prepared free. Posterior bones poorly demarcated, frontals broken off, and area around parietal foramen broken out. Internally, incomplete otic capsules are crushed and sheared to the left and an eroded-looking band connects them across the midline.
PR 1654. Basiparasphenoid, central portion only prepared free. Includes basipterygoid processes and basal plate of fused parasphenoid. Reveals where anterior extension of the basioccipital would penetrate anterior to the posterior edge of the parasphenoid. Missing cultriform process and posterior portions of cristae ventrolaterales.
PR 1655. Basiparasphenoid prepared free. Dorsoventrally compressed but retaining some three-dimensionality.
PR 1700. Holotype skull and postcranium on mudstone. Skull laterally compressed with skull roof and snout in dorsal view and right cheek rotated into same plane. Right and left maxillae partially exposed. Aspects of palatal bones exposed through orbit (Lombard & Bolt Reference Lombard and Bolt1995, pl. 1).
PR 1701. Basioccipital and basiparasphenoid in ventral view with three associated anterior vertebral elements and four nearby articulated vertebral elements of uncertain placement (Fig. 6).
PR 1747. Broken palate and jaw separately prepared. The palatal piece preserves the anterior parts of the maxillae, between which are the broken right premaxilla, choana, vomer, palatine, as well as eroded fragments of a pterygoid with denticles.
PR 1792. Laterally compressed and disarticulated skull and jaws on mudstone. Right jaw in medial view, left in lateral view. Broken cheek and snout with ventral exposure of skull roof. Both palatal and quadrate ramus of right pterygoid in articulation with right jaw and right ectopterygoid and palatine exposed in ventral view. The dentition on the jaw and palatal bones is in approximate register with some disruption anteriorly. Left pterygoid in dorsal view partially obscured by broken cheek bones.
PR 1809. Two separated specimens. 1. Skull: crushed skull prepared free of matrix. Right side of skull exposed on one side of specimen with parts of palate visible through the orbit. The obverse presents the skull table, occiput, and parts of the palate. 2. Jaw: anterior tip of the lower jaw in three dimensions, apparently with little distortion (Lombard & Bolt Reference Lombard, Bolt, Carrano, Gaudin, Blob and Wi2006, fig. 2.2).
PR 1813. Reasonably complete but laterally crushed skull that has been prepared free. One side preserves much of the right side of the skull and mandible. The other side presents the internal surface of the right mandible, the skull roof, and some occipital elements, some dermal elements of the left side of the snout including the left premaxilla, and unidentified fragments of the palate and occiput.
PR 1814. Anterior palate in ventral view in mudstone (Fig. 2). The marginal elements are sheared to the left but maintain their relative positions. Because of the shearing, the left premaxilla, choana, vomer, palatine, and pterygoid, all in articulation, are best exposed. Those same elements on the right are obscured by the folded-over premaxilla and maxilla. The internal surfaces of skull roofing bones are visible through the right choana.
PR 1816. Incomplete skeleton with cranium and postcranium, showing basiparasphenoid in ventral view.
PR 1817. Basiparasphenoid in ventral view with basipterygoid processes, cultriform process. Hand specimen in limestone.
PR 1818. Left maxilla prepared free of matrix except for that supporting the teeth (Fig 4a, b). The anterior end preserves the choanal surface; the posterior end is missing.
PR 1888. Partial disarticulated skull, plus disarticulated digital elements on a mudstone block prepared from both sides. One side preserves the right premaxilla and maxilla, dermal elements of the snout and skull roof. The other side preserves both jaws and a palatal piece that includes the right pterygoid with the basipterygoid joint, as well as a partial ectopterygoid.
PR 1954. Three-dimensional complex of pterygoid, ossified palatoquadrate, and quadrate in dorsal view on mudstone (Fig. 5). The pterygoid is damaged anteriorly and is partially covered by the right premaxilla and partial maxilla.
PR 2010. Right maxilla, lateral view, two halves broken in middle and posteriorly. In limestone.
PR 2891. Freestanding dorsoventrally crushed left palatine. Articular surfaces for maxilla and pterygoid are evident.
PR 1636. Anterior end of (mostly disarticulated) skull, transfer-prepared, in ventral view. Shows, inter alia, anterior terminus of denticulated left pterygoid.
PR 3061. Partial basiparasphenoid, partly in limestone. Missing area posterior to basipterygoid processes. Shows denticles on parasphenoid.
SUI 147644. Freestanding left premaxilla preserving a fang and fang socket, palatal shelf, and facial process, but missing the choanal margin and lateral marginal dentition (Fig. 3a, b).
SUI 52055. Small braincase prepared free (Fig. 7).
2. Description
We present first an overview of the palate as reconstructed in Figure 1. Next, the individual bones are considered in anterior to posterior sets, beginning with those at the margin and ending with those at the midline. This is followed by a description of the braincase and parasphenoid, the occiput, otic capsule, and stapes.

Fig. 1 Whatcheeria deltae. Schematic interpretive reconstruction of the palate in ventral view. The reconstruction is an estimate of maximum width and is primarily based on the anterior palate of PR 1814, pterygoids and maxillary teeth of PR 1792, and the braincase of PR 1701. The palatal ramus of the pterygoid is reconstructed as flat in the horizontal plane, the vertical arrow thus indicating that maximum palatal width controls skull width. In addition, the ectopterygoid is reconstructed as excluding contact by the pterygoid with the maxilla, though evidence is from a hint in one specimen. Thus, this is a conservative maximum width. Any vaulting of the palatal ramus dorsally from lateral to medial and/or direct contact of the pterygoid with the maxilla would narrow the skull. Abbreviations: ARF = adductor fossa; BIP = basiparasphenoid; BIO = basioccipital; BPJ = basipterygoid joint; CHO = choana; ECP ectopterygoid; MAX = maxilla; PAL = palatine; PMX = premaxilla and the label is on the premaxillary shelf; PTEr = palatal ramus of pterygoid covered in denticles; PTEq = quadrate ramus of pterygoid; QOJ = quadratojugal (and jugal?); QUA = quadrate = no quadrate at the jaw joint is preserved; VOM = vomer.
2.1. Overview of the palate
The palate is reconstructed as flat in the horizontal plane with the lateral palatal bones and the pterygoids coplanar, which places a constraint on the maximum width of the skull. The skull would be narrower than reconstructed if the lateral palatal bones and pterygoids formed a vaulted roof to the oral space. The exterior boundary of the palate is formed anterior to posterior by the premaxilla, maxilla, and quadratojugal. Whether the jugal contributes a small segment is unclear because the boundary between jugal and quadratojugal is not determinable. The premaxilla contributes a prominent shelf to the anterior palate, to which the vomers are joined. There is no anterior palatal fenestra. A large choana lies between the vomer and maxilla, and is bounded anteriorly by the premaxilla and posteriorly by the palatine. Through it, the external naris and interior surfaces of the nasal, lacrimal, and jugal are visible. The palate of Pederpes, the sister taxon to Whatcheeria in all analyses published to date, has been tentatively reconstructed (Clack & Finney Reference Clack and Finney2005, fig. 17), but for none of the preceding features is the anterior region well enough preserved for informative comparison. Among early large-bodied tetrapods, the absence of an anterior palatal fenestra coupled with a palatal shelf on the premaxilla has been described only in Proterogyrinus scheelei (Holmes Reference Holmes1984). The premaxilla and maxilla are toothed, and both carry large fang teeth. The vomer, palatine, and ectopterygoid are toothed, each having fangs that are slightly larger than the marginal dentition plus several teeth that are smaller than the marginal dentition. None of the bones in this series bears denticles, unlike those of Pederpes (Clack Reference Clack2002a; Clack & Finney Reference Clack and Finney2005). Medially, this series articulates with the pterygoid.
Each pterygoid has a palatal ramus that extends at least as far as the palatal shelf of the premaxilla and which, together, obscure the (presumed) intervomerine joint in palatal view. The palatal rami of the pterygoids are covered by denticles, extending posteriorly to the level of the basipterygoid articulation. The quadrate ramus lies in a roughly vertical plane and extends posteriorly to articulate with the quadrate, forming the ventromedial border of the opening of the adductor fossa. The lateral border of the adductor fossa is formed by the maxilla and quadratojugal (with possibly a small contribution from the jugal). The exact shape of anterior and posterior borders of the opening of the adductor fossa is uncertain and the exact arrangement of the elements forming the anterior border is unknown. We have reconstructed the anterior border as formed by the maxilla, ectopterygoid, and pterygoid. The pterygoid vacuities are small and the pterygoids join the midline basiparasphenoid at the basipterygoid joints.
2.2. Bones of the palate
2.2.1. Premaxilla
The external surface of the premaxilla is unsculptured but bears numerous pits, more densely spaced than elsewhere on the skull surface. External shape is shown in Lombard & Bolt (Reference Lombard and Bolt1995, fig. 1), and we can add little to that. The facial process is of fairly even height, with no sharp peak, and the nasal–premaxillary suture is strongly interdigitated. A foramen for the supraorbital lateral line enters the facial process approximately at its mid-width and dorsal to the palatal shelf. Its further course is covered. On the external surface dorsal to the teeth, numerous small foramina, some in a horizontal row, indicate sensory openings for the lateral line as preserved in PR 1814 and SUI 147644. The premaxillae meet at a butt joint. The premaxilla forms the anterior border of the external narial opening, and the anterior half of its ventral border; both surfaces are smooth and, with the exception of a foramen for the infraorbital lateral line canal, are featureless (Fig. 2a, b). The ventral narial border slants downward toward the joint with the maxilla. At their junction, the premaxillary portion is low and rounded in lateral view, and the joint area is small and smooth.

Fig. 2 (a) Whatcheeria deltae, PR 1814. Interpretive drawing of the anterior palate in ventral view. (b) Whatcheeria deltae, PR 1814. Key to (a). The choana is outlined in bold and the internal surfaces of dermal roofing bones and the external narial opening are visible through it. Labelled outlines: 1 = vomerine fang; 2 = palatine fang. Abbreviations: ECP = ectopterygoid; EN = external naris; IOL opening for infraorbital lateral line; JUG = jugal = LAC = lacrimal; MAX = maxilla; NAS = nasal; PAL = palatine; PMX = premaxilla; PTE = pterygoid; VOM = vomer.
In palatal view, the premaxilla is drawn out into an extensive, robust palatal shelf, the lateral portion forming the anterior border of the choana. The shelf is best seen in partial skull PR 1814 (Fig. 2a) and in a three-dimensional premaxilla (SUI 147644) in which some of the medial and posterior edge is missing (Fig. 3a, b). Its presence is not apparent on other separated (and heavily damaged) premaxillae available, but it is partially visible on, for example, the broken right premaxilla of PR 1747 and the left premaxilla of PR 1813. The palatal shelves articulate with the vomers and at the midline cover the anterior projections of the pterygoid (Figs 1, 2a). In SUI 147644 (Fig. 3a, b), the shelf is seen to tilt dorsally from lateral to medial, suggesting that the palate is vaulted, at least anteriorly. The dorsal interior junction of the shelf with the ascending process has a foramen of indeterminate function (Fig. 3b). No anterior palatal fenestrae are visible in the palatal shelf. In PR 1814 (Fig. 2a), the palatal shelves are visible and roughly in place, but no midline suture is visible. Details of the premaxilla–vomer suture are unclear, but evidence mainly from PR 1814 and PR 1792 indicates that it was a bevelled joint with the anterior end of the vomer lying dorsal to the palatal ledge of the premaxilla.
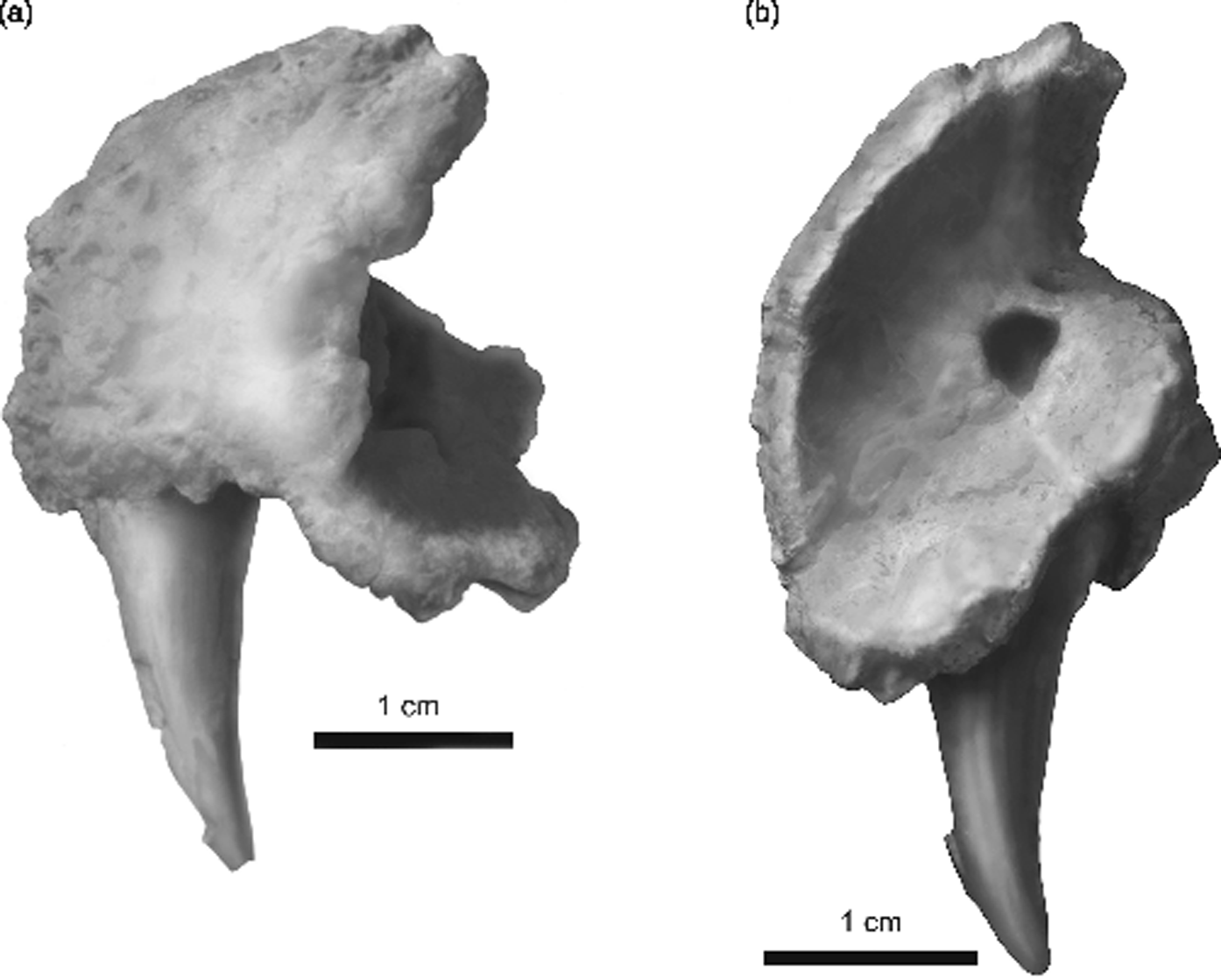
Fig. 3 Whatcheeria deltae, SUI 147644. Interpretive drawing of an isolated left premaxilla. (a) Lateral view. Anterior to the left, dorsal to the top. The premaxillary fang projects ventrally, the facial process dorsally, and the premaxillary shelf posteriorly. (b) Internal view from posterodorsal rotated counter clockwise to best display the premaxillary shelf. Medial to the right, dorsal to the top. The fang projects ventrally, the facial process dorsally.
A premaxillary tooth count is difficult to determine, but appears to be in the range of four to six tooth positions. A premaxillary fang occurs at or near the midline suture. Tooth morphology is similar to that of the maxilla, with bluntly conical tips that are curved inward.
2.2.2. Septomaxilla
One possible example of the septomaxilla is preserved in the left external naris of PR 1634. It was originally tubular in overall shape, but has been crushed and broken longitudinally into two halves. Beyond this, it shows no interpretable structure. It is partly buried in matrix, which was not removed because of the fragility of surrounding structures.
2.2.3. Maxilla
Good examples of the maxilla are preserved in several skulls, as well as isolated bones in a range of sizes. Medial and dorsal views are uncommon, however, and for these we depend on PR 1818 (Fig. 4), which has been prepared mostly free of matrix. The maxilla is long and low in lateral view, extending from its joint with the premaxilla ventral to the external naris to somewhere near the posterior margin of the jugal. It thus forms about 67% of the external ventral margin of the skull. There is a low facial process dorsal to the fangs. At this point, the facial process bulges medially as a massive buttress enclosing the roots of the maxillary fangs. The maxilla is joined to the lacrimal, and jugal dorsally and internally to the palatine and ectopterygoid. The lateral surface of the maxilla lacks sculpture and bears numerous fine pits. Absence of sculpture is likely not a simple function of size, as even large examples lack it. There is no indication of the lateral line, which runs in the jugal and lacrimal dorsal to the maxilla. At the external naris the lateral line canal appears to exit an anteriorly opening foramen in the lacrimal and presumably crosses the naris (perhaps in a missing lateral rostral) and enters the premaxilla, at a posteriorly facing foramen (IOL, Fig. 2b) and best seen in PR 1634. A branch may exit to the surface of the maxilla, posterior to the naris (see description of sulci in third paragrah following).
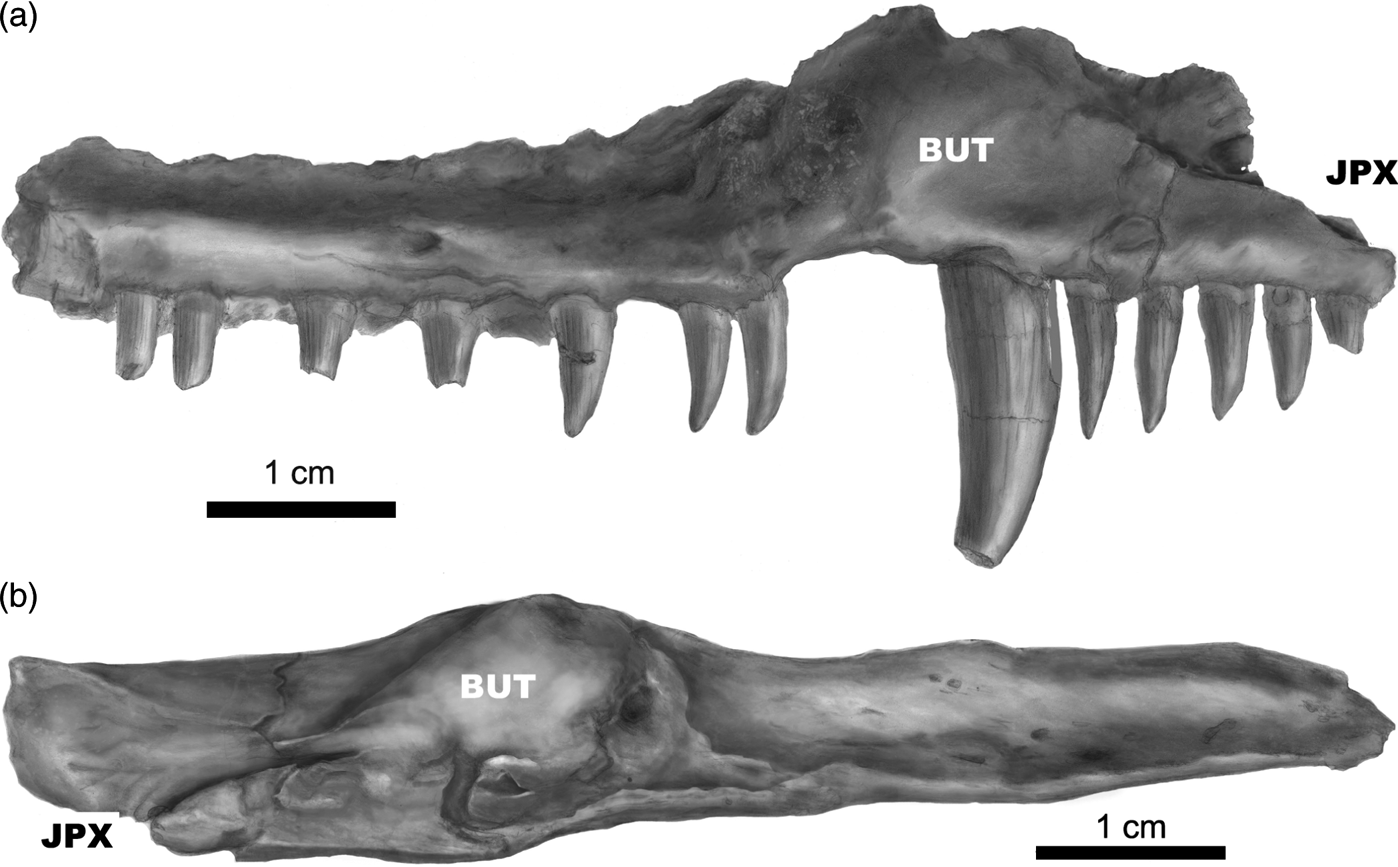
Fig. 4 (a) Whatcheeria deltae, PR 1818. Interpretive drawing of internal view of an isolated left maxilla. Anterior is to the right, dorsal to the top. The butt joint with the premaxilla (JPX) is at the anterior end. The extended posterior portion is missing. The smooth surface from the anterior tip to the fang base forms the lateral margin of the choana. BUT = thickened eminence for the maxillary fang(s). (b) Whatcheeria deltae, PR 1818. Interpretive drawing of an isolated left maxilla in dorsal view. Anterior is to the left, lateral to the top. The butt joint with the premaxilla (JPX) is at the anterior end. The extended posterior portion is missing. The large bulge (BUT) is the buttress at the base of the maxillary fang(s).
The maxilla forms the posterior one-half of the ventral border of the external naris, and a very small part of its posterior border. The ventral border is smooth finished bone, and is the width of the adjacent premaxillary process which meets it (Fig. 2a). It runs gradually downward toward the suture with the premaxilla, producing a somewhat wedge-shaped profile in lateral or medial view. The contact area for the premaxilla is small and rounded, with a slightly pitted surface but no indication of interdigitating or bevelling, mirroring the premaxillary side of its contact. The suture is, thus, best characterised as a butt joint.
Internally, the maxilla forms the lateral border of the choana, where in dorsal or ventral view it is neither noticeably curved nor excavated, but the large maxillary buttress for the fangs bulges into the posterior part of the lateral choanal outline to some degree (Fig. 4a). Most of the medial surface of the buttress is smooth, with some fine pitting. Posteriorly, its medial and posterior surface is incised by a rough surface that marks the contact with the anterolateral end of the palatine. This contact surface extends posteriorly on the medial and dorsomedial surface of the buttress, for a distance approximately equal to the length of the internal naris. Viewed from the medial side, it is slightly concave. The contact surface for the palatine becomes less rugose posteriorly, and ends in a small bony eminence. The same structure is visible in ventral view on the left side of skull PR 1814 (Fig. 2a), where it marked the junction between palatine and ectopterygoid. Posterior to this point, the lower medial surface of the maxilla is smooth finished bone with some fine pits. The maxilla and palatine appear to be in apposition along the entire length of the palatine and are so indicated in the reconstruction (Fig. 1).
The dorsolateral surface of the buttress, and of the pars facialis anterior to it, is thickened and coarsely irregular and clearly sutural with the lacrimal (+jugal?) (Fig. 4). It bears two sulci of unknown function. One sulcus is short and directed mediolaterally. The other runs ventrally and anteriorly from near the summit of the buttress, passing to the inner side of the posterior border of the external naris. In PR 1818 (Fig. 4), it opens to the exterior as two foramina whose borders are incomplete anteriorly. The ventral one of these is larger, and aligned with the sulcus; the dorsal one is much smaller, and is not so aligned. A notch in this position may be a general feature of Whatcheeria; we can confirm the presence of a notch in a right maxilla of PR 1888, right maxilla of PR 1814, and left maxilla of PR 1747. The absence of a notch or foramen in some other specimens does not necessarily imply that the sulcus is absent; apparent absence may be due to the availability of exterior views only.
Laterally on the dorsal surface of the maxillary buttress, and extending anteriorly and posteriorly to it, there is a clearly marked contact area for the lacrimal+jugal? (separate contact areas are not distinguishable), which at this point is a butt joint. Posterior to the buttress, this becomes a bevelled joint in which the ventrolateral surface of the jugal is overlapped by a low, thinner lamina of the maxillary pars facialis. In palatal view, the maxilla is excluded from contact with the vomer by the choana. Posteriorly, the maxilla appears not to have a joint with the pterygoid at the anterior margin of the adductor fossa, but this is uncertain. The degree to which the maxilla borders the adductor fossa is indeterminable from the available specimens. In our reconstruction (Fig. 1), we indicate a contribution but also emphasise uncertainty here.
Measurement of the maxilla is hampered by the fact that most specimens are more or less incomplete, and/or are preserved in a single view. Measurements in Table 1 are, thus, somewhat approximate allowing an impression of the size range of maxillae that are available as nearly complete specimens; this could certainly be extended to smaller sizes in particular, if less-complete maxillae were included. Large specimens such as PR 2010 give an impression of massiveness, and the measurements in Table 1 suggest that growth in height was more rapid than growth in length.
Table 1 Measurements (in mm) and tooth counts in the maxilla of Whatcheeria.

Maxillary teeth are similar to those of the premaxilla, with the obvious exception of fang placement, the maxillary fang being a synapomorphy of whatcheeriids (Clack & Finney Reference Clack and Finney2005). As seen in Table 1, maxillary fangs are preceded by four to six ‘normal-sized' marginal teeth. The fangs were alternately replacing, as indicated by a large empty space immediately behind the fang in many specimens including PR 1818, and the presence of two fangs in PR 2010. Empty tooth positions are common in maxillary specimens, presumably in most cases as a result of normal replacement; in PR 1818, the second tooth from the front shows a resorption pit at the base.
2.2.4. Vomer
There are no separated specimens of the vomer. Incomplete examples are preserved in PR 1792, PR 1747, and PR 1814 (Fig. 2a). In palatal view, the vomer is roughly rectangular and about twice as long as wide; it forms most of the medial margin of the choana (Fig. 1). The vomer is joined to the palatal shelf of the premaxilla anteriorly, which it overlies dorsally. There are no lateral vomerine extensions that form the anterior or posterior borders of the choana.
The intervomerine suture is more or less obscured in palatal view. In specimens where it is partially visible it appears to be a butt joint. Details of the potential joints between the vomers and with the anterior tips of the pterygoids are unclear from the specimens available, but may be, in part, a butt joint, or bevelled.
Several lines of evidence suggest that the vomer, palatine, and ectopterygoid contact the lateral border of the pterygoid in a flat, bevelled suture on its dorsal surface, and apparently with small overlap as, for example, is seen in cross sections in Acanthostega (Clack Reference Clack1994). This conclusion derives mostly from the following observations. In PR 1792, the right side of the palate is exposed in ventral view. The right pterygoid has been displaced laterally to cover the medial parts of the three paired lateral bones, and in the case of the palatine and ectopterygoid it completely covers the bones in ventral view, up to their tooth roots. Its anterior–posterior position relative to the paired bones seems approximately natural: the posterior end of the ectopterygoid either forms part of the border of the adductor fossa or is excluded from it by a contact of pterygoid with quadratojugal. The left pterygoid in this specimen has been folded over the right, whose medial portion it partly covers, while exposing the lateral portion of its own dorsal surface. The lateral margins of both left and right pterygoids are intact, and describe the same smooth curve. The right pterygoid is heavily denticulate up to its lateral margin, which precludes any overlap of the lateral bones onto its ventral surface. The lateral margins of both pterygoids are thin, and it seems unlikely that the contact with the lateral bones could have been a butt joint. The lateral dorsal surface of the left pterygoid shows no obvious sign of sutural contact. Although we have focused here on evidence from one specimen, no observations on other specimens contradict those observed in PR 1792.
In PR 1814 (Fig. 2a), vomerine teeth are borne on a somewhat raised crest that is concave laterally with the teeth lying along the border of the choana. A small tooth and two alternately replacing fang teeth lie in the anterior half of the vomer and two or three smaller teeth in the posterior half. The crown morphology of the single intact tooth is identical to that of the marginal teeth. The vomerine fangs in PR 1814 are the smallest of Whatcheeria's palatal fangs, in fact barely reaching the size standard advocated by Lombard & Bolt (Reference Lombard, Bolt, Carrano, Gaudin, Blob and Wi2006, p. 26): they must be ‘at least twenty-five percent greater in maximum basal diameter and/or height than the average of adjacent marginal teeth'. A replacement crown is visible anterior to the functioning fang on the left vomer; it appears substantially larger than that of the functioning fang. There are no denticles.
2.2.5. Palatine
In the form of an irregular rectangle with a laterally convex border, the palatine's exposure in palatal view is slightly over two times as long as it is wide, and is similar in size to the vomer. The palatine joins the maxilla laterally, the pterygoid medially, the ectopterygoid posteriorly, and the vomer anteriorly. Part of the anterior edge lateral to the joint with the vomer is excavated dorsally, forming the sloping posterior wall of the choanal opening in a manner identical to that illustrated for Proterogyrinus by Holmes (Reference Holmes1984, fig. 3c).
The palatine–vomer suture is apparently bevelled, and likely flat; but the superpositional relationships of the two bones in the suture are uncertain. The maxilla–palatine suture is partially visible in PR 1814 (Fig. 2a), where the two bones have pulled slightly apart, and the contact between the fang buttresses on the palatine and maxilla is partly visible. The bones are further separated posterior to the maxillary buttress, and at least the ventral portion of the suture on the maxilla is visible. This is a smooth surface. Palatine PR 2891, the only marginal palatal bone for which we have a freestanding example, is large, being some 4.7cm long in its greatest anteroposterior dimension, as opposed to an estimated maximum 3.0cm for the palatine of PR 1814. On the lateral sutural surface of PR 2891, a coarsely rugose and presumably sutural area continues nearly to the posterior extremity of the bone. This suggests that the largest maxillae would have a matching rugose sutural area on their medial surface.
The centre of the palatine is occupied by a pair of large fangs preceded and followed by two or three labyrinthine teeth that are smaller than the marginal dentition on the maxilla. The teeth are not on a markedly raised ridge. PR 2891 shows that the fang pair is supported by a massive buttress that extends along the lateral side of the palatine, with its greatest thickness at the location of the fangs. At this point, the palatine is approximately 1.3mm thick. There are no denticles.
2.2.6. Ectopterygoid
The ectopterygoid is partly visible in PR 1792 and PR 1814 (Fig. 2a). Based on these and other specimens, a reasonably accurate composite reconstruction of the ectopterygoid in palatal view is possible (Fig. 1). The bone is roughly rectangular with a length-to-width (L:W) ratio of about 3.3:1 as preserved; the ratio would undoubtedly be greater in a complete example. Due to damage/cover, the posterior relationship of the ectopterygoid to the adductor fossa is not certain; in the reconstruction it is shown as forming part of the anterior border of that opening. This is as generally illustrated for stem tetrapods and early temnospondyls; we have no evidence of the pterygoid joining the maxilla or jugal to exclude the ectopterygoid from the fossa as reconstructed in, for example, Proterogyrinus (Holmes Reference Holmes1984, fig. 3c) and Archeria (Holmes Reference Holmes1989, text-fig. 1). Anteriorly, the ectopterygoid sutures to the palatine, laterally the maxilla, and medially the pterygoid. The ectopterygoid–palatine suture is not interdigitated, but no other information about it is available.
In PR 1792, the palatine and ectopterygoid are subequal in length. The palatine and ectopterygoid dentitions are exposed on the right side, but the pterygoid has drifted laterally to cover some of the palatine and all of the ectopterygoid except the teeth, although the lateralmost part of the suture between them can be identified. From anterior to posterior, the ectopterygoid dental pattern seen on this specimen is: tp–t–t–Fp–F–tp–t–t–tp–t, where t=functioning small tooth, tp=tooth position that once held a small tooth, F=fang tooth, and Fp=fang-tooth position. The comparable series on the left side of PR 1814 (Fig. 2a) is t–tp–t–Fp–F, and the posterior part of the ectopterygoid is not exposed. In this specimen, one of the tooth positions is occupied by a replacement crown, as is the first fang position, which indicates that the fangs were alternately replacing, as expected. In both specimens, the row of small teeth begins at the palatine–ectopterygoid suture. All teeth are labyrinthine and denticles are not evident.
2.2.7. Pterygoid and epipterygoid (primary palatoquadrate)
Though the pterygoid is at least partly exposed in a number of specimens, in none could it be prepared free. Crushing has usually reduced this eminently three-dimensional bone to little more than two dimensions: the quadrate ramus, the palatal ramus, and the area of the basipterygoid articulation are preserved nearly in the same plane. An exception is PR 1954 (Fig. 5), which includes a pterygoid in dorsal view and a three-dimensional complex of pterygoid, ossified palatoquadrate, and quadrate. Though somewhat compressed dorsoventrally, the latter two components stand at approximately 90° to the plane of the palatal ramus of pterygoid. The vertical portion of palatoquadrate+quadrate ramus extends from approximately the basal process to the quadrate. The dorsal edge of the combined bones is damaged and incomplete. This posterior vertical portion of the palatoquadrate + quadrate ramus is continuous with the palatal ramus, which is also damaged and incomplete. Unfortunately, the basal process is hidden by a displaced nasal + premaxilla, which cannot safely be removed, and part of the transition from horizontal to vertical in the region of the basal process and columella cranii? (CAC? Fig. 5) has been mostly flattened from vertical to horizontal.

Fig. 5 Whatcheeria deltae, PR 1954. Interpretive drawing in dorsal view of a somewhat three-dimensional complex of pterygoid, ossified palatoquadrate and quadrate. Abbreviations: CAC? = columella cranii; EPT = epipterygoid; NAS = Nasal; PMX; premaxilla; PTE = (left) pterygoid; QUA = quadrate; REPT = right pterygoid.
Several aspects of the pterygoid are immediately apparent in this specimen. The interpterygoid vacuities are narrow. The profile of the lateral margin in both dorsal and ventral view is straight to smoothly curved, with no indication of suture with the lateral palatal bone series. The palatal ramus is slightly longer than the quadrate ramus. There is a prominent anterior digitiform process, shaped such that a ventrally facing trough is produced when the pterygoids are in articulation. This is very similar to Greererpeton (Smithson Reference Smithson1982, fig. 11). The quadrate ramus proper (exclusive of the epipterygoid) is strut-like, suggesting at first sight that the skull in posterior view might be open dorsal to the quadrate ramus. This idea is corrected in large skulls, where an extensive epipterygoid ossification meeting the squamosal is present. Even in a crushed specimen the socket of the basal joint is preserved standing somewhat proud of the plane of the rest of the pterygoid, and the tympanic excavation is identifiable immediately posterior to it (PR 1792). In crushed, two-dimensional specimens, it is apparent that for the basipterygoid joint to have functioned, the region of the pterygoid adjacent and posterior to the basal socket must be restored to a more vertical position relative to the palatal ramus. From this general consideration, plus the three-dimensional PR 1954, it is clear that the basipterygoid joint is roughly in the plane of the palatal ramus. A medially projecting basal process plus basal socket thus carried the articulating facets for the basipterygoid, such as has been described in many other early tetrapods.
The interior of the basal socket is not well preserved in any of the specimens, so it is unclear how closely it mirrored the facet(s) on the basipterygoid process. PR 1888 and PR 1809 (skull) show at least part of the articulating surface within the basal socket, but the composition of the articulating surfaces is uncertain. It is very likely, based on comparative anatomy, that the epipterygoid forms much of the socket and the articulating surfaces. We know that there is an ossified epipterygoid in Whatcheeria – there is an example in PR 1809 (skull). But as is often the case in early tetrapods, there is no clear suture within the socket between epipterygoid and (ento)pterygoid, and the extent of the epipterygoid outside of the basipterygoid region is difficult to determine on all but PR 1809 (skull). This specimen, massively distorted though it is, provides vital information. A columnar structure, flattened medio-laterally, is visible through the broken right orbit. This appears to be the right ascending process of the epipterygoid, visible in lateral view. It is continuous with and dorsal to a large plate of bone, visible through the orbit, which can only be the epipterygoid. The ascending process lies lateral and ventral to, and partly obscures, an area of thick and badly fractured bone, which includes a columnar projection very similar to the right ascending process. We interpret this as the left ascending process and epipterygoid in medial view. No basal socket is visible on the left ascending process, possibly because it is covered by the right process. Both ascending processes bear a terminal process that juts anteriorly at about 30°, whose function is unclear. It is uncertain whether the ossified ascending processes reached the underside of the skull roof, although it seems unlikely. If they did so, the basipterygoid joint would have to be very dorsally located, which appears not to be the case. Anterior to the basipterygoid joint, the extent of ossification in the palatoquadrate cartilage is unknown.
Clearly visible about a centimetre anterior to the ascending processes is the distal end of the right basipterygoid process, which, in this large specimen, shows clear dorsal and ventral facets. The epipterygoids are, thus, posteriorly displaced from their original position relative to the basipterygoid process. Some of the bone visible through the right orbit is likely part of the right otic capsule, but it is not interpretable in more detail.
The medial surface of the quadrate ramus of the right epipterygoid of PR 1809 (skull) is extensively visible in what might now be called the posterior view, relative to the orientation of the posterior skull table. In this view, it is in articulation with the quadrate ramus of the pterygoid and with the quadrate, although sutures between these bones and the epipterygoid cannot be reliably distinguished. It is clear from this specimen that most of the posteromedial wall of the adductor chamber was formed by the epipterygoid. In other words, the epipterygoid filled in the large gap between the slender quadrate ramus of the pterygoid and the occipital surface of the cheek bones, mostly the squamosal. The contact between the epipterygoid and cheek bones is not perfectly preserved in such a distorted specimen. In some places, though, the margin of the epipterygoid is apparently intact, and is a simple rounded edge that suggests a butt joint; certainly, there is no evidence for any complex suture.
2.3. Braincase and parasphenoid
As usual in early tetrapods, the basisphenoid and parasphenoid in Whatcheeria are fused, although the boundaries of each are often apparent. We refer to the composite ossification as the basiparasphenoid. The ossified braincase of Whatcheeria is limited to the basiparasphenoid and basioccipital; there is no sign of a sphenethmoid in any specimen available. The basiparasphenoid is seen in dorsal or ventral views in four separated specimens: PR 1654, PR 1655, PR 3061, and SUI 52055. It is visible only in ventral view in PR 1817 and PR 1701, and preserved in dorsal view in association with skull material in PR 1809 (skull) and PR 1888. No basioccipital has been recovered (or recognised) as a separated element, but it is visible in situ in ventral view on PR 1701, PR 1651, and PR 1816. Most braincase specimens are badly crushed. Therefore, we base the following description primarily on PR 1701 (Fig. 6) in ventral view and SUI 52055 (Fig. 7) in dorsal view. SUI 52055 is the best preserved, though smallest and least ossified, braincase available. Details are added from other specimens as appropriate.
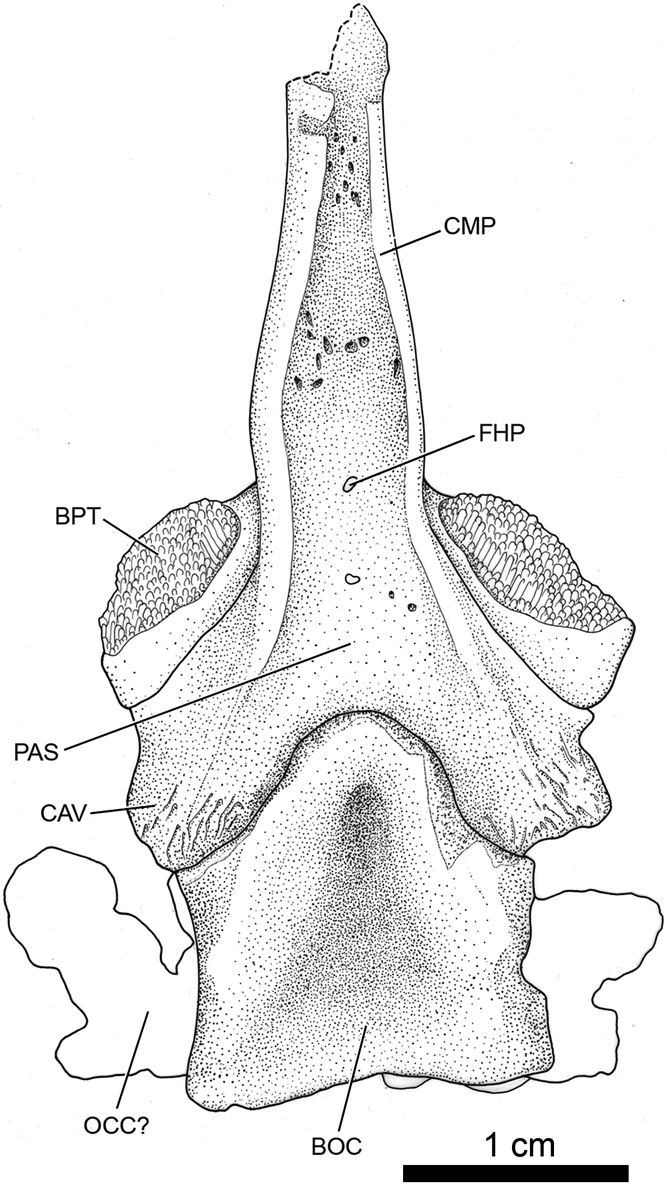
Fig. 6 Whatcheeria deltae, PR 1701. Interpretive drawing of the basioccipital and basiparasphenoid in ventral view. Anterior to the top. Abbreviations: BOC = basioccipital; BPT = basipterygoid joint surface; CAV = crista ventrolateralis; CMP = cultriform process; FHP = foramen hypophyseos; OCC? = otic capsule; PAS = parasphenoid.
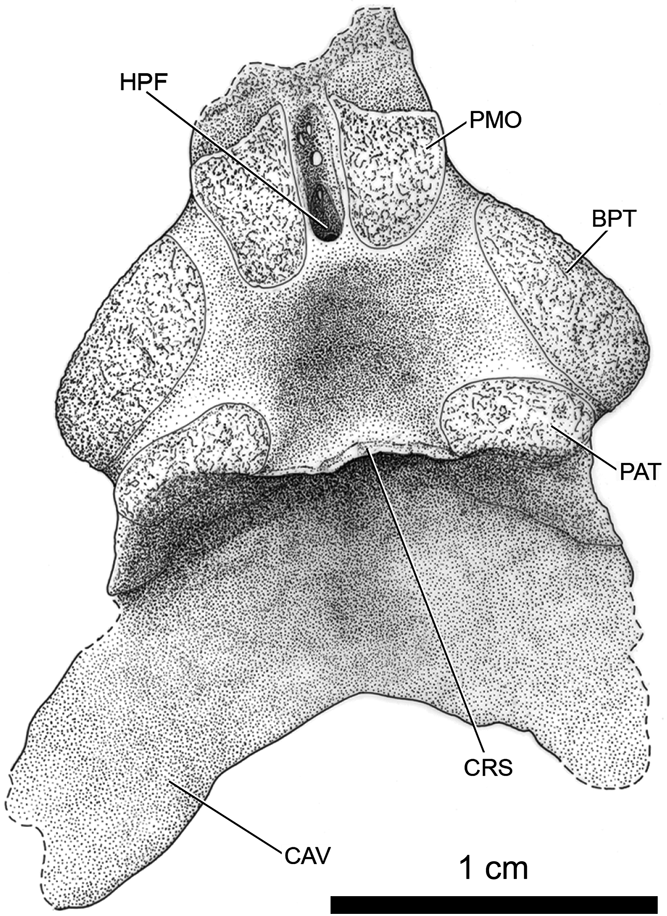
Fig. 7 Whatcheeria deltae, SUI 52055. Interpretive drawing of basiparasphenoid in dorsal view. The basioccipital is missing. Anterior is to the top. Abbreviations: BPT = basipterygoid joint surface; CAV = crista ventrolateralis; CRS = crista sellaris; HPF = hypophyseal fossa; PAT = pila antotica. PMO = pila metoptica.
The basipterygoid processes are very short. The joint surface is unfinished, and is directed anteriorly at about 45° from the centre line of the skull. In smaller specimens, the joint surface is in one plane; in larger specimens, each process shows two more or less distinct surfaces, directed respectively anterodorsally and anteroventrally, similar to those described in Proterogyrinus (Holmes Reference Holmes1984, p. 459). The centre of the basal plate of the parasphenoid is U-shaped and open posteriorly; it is also very short, extending posteriorly little beyond the basipterygoid processes. The medial area (base of the U) is thickened relative to the area anterior to it, with smooth periosteal bone forming its posterior border (also in PR 1655, the only other example in which the area is clearly seen). The arms of the U extend posteriorly alongside the basioccipital, are striated on dorsal, ventral, and medial sides near their termini, and appear to have been in sutural contact with the basioccipital. The contact appears to extend along the lateral side of the anterior half of the basioccipital, as is well shown in PR 1701. These posterior arms are likely homologous to the ‘cristae ventrolaterales' described by Smithson (Reference Smithson1982) in Greererpeton and identified in the primitive temnospondyl Dendrerpeton by Robinson et al. (Reference Robinson, Ahlberg and Koentges2005) and the ‘basal tubera' described in Pholiderpeton (Clack Reference Clack1987). As in Greererpeton, the cristae project laterally beyond the basisphenoid. Smithson suggested that in that genus they formed the lower rim of the fenestra ovalis and supported the ventral rim of the stapedial head. In Whatcheeria, an unknown amount of the lateral projection is likely due to crushing and the fenestra ovalis is not preserved. However, in PR 1651, both stapes, though crushed, are apparently approximately in place just posterior to the basisphenoid–basioccipital junction. The cristae ventrolaterales are in contact with the anterior one-half of the stapedial heads, as well as with the lateral edge of the basioccipital. Although no fenestra vestibuli has been observed in any specimen, we conclude that the stapes mark its position. Although there has been crushing, and likely some displacement of the stapes, the relationships described tend to confirm Smithson's suggestion.
Smithson described ‘tubera basisphenoidales' in Greererpeton, developed in the parasphenoid near the midline, and suggested these as attachment sites for subvertebral muscles. No similar structures are discernible in Whatcheeria, either in the parasphenoid or the basioccipital. PR 1701 (Fig. 6) shows a marked depression in the anterior part of the basioccipital, beginning anteriorly between the ‘cristae ventrolaterales' of the parasphenoid. This may be due to crushing, as the area dorsal to this depression was likely poorly ossified in PR 1701. The same border is visible in PR 1651 and PR 1816, although it does not stand much above the parasphenoid.
The base of the U-shaped curved contact between the parasphenoid basal plate and basioccipital is conventionally identified as marking the ventral cranial suture between the basioccipital and basisphenoid, as in Acanthostega (Clack Reference Clack1998), Crassigyrinus (Panchen Reference Panchen1985; Clack Reference Clack1998), and Pederpes (Clack & Finney Reference Clack and Finney2005). In Whatcheeria, this actually marks the posterior edge of the parasphenoid covering of the basioccipital. The basioccipital extends anteriorly dorsal to the parasphenoid to the level of the pilae antoticae and crista sellaris, where a posterior-facing joint surface at the base of each pila antotica and a joint (?) on the posteroventral surface of the crista sellaris form the basisphenoid–basioccipital joint. The ventral manifestation of this compound joint would be the ventral cranial suture but it is under cover of the parasphenoid and, thus, not visible. This morphology is revealed in specimens where the basioccipital has fallen out of contact with the basisphenoid, such as illustrated in dorsal view in the juvenile specimen SUI 52055 (Fig. 7) where the posterior edge of the parasphenoid and the cristae ventrolaterales are partially preserved and are lying ventral to the missing basioccipital.
In ventral view, much of the parasphenoid area of the basiparasphenoid is smooth-surfaced, finished bone. However, the centre of the parasphenoid basal plate up to some distance on the cultriform anterior to the basipterygoid processes consists of irregular-surfaced finished bone marked by numerous small punctae/foraminae. This area generally does not show denticles, but where the surface has not been prepared, it carries numerous very small denticles, borne on denticulate plates separate from the underlying parasphenoid (not depicted in specimen drawings). This is strongly suggested by PR 3061, in which the separation is marked by a line of white matrix. Further preparation has not been undertaken, in view of the fragility of the specimen. The number of plates and their ontogeny is unknown. On PR 1701, the cultriform process shows two small centrally located foramina. The largest of these is just anterior to the basipterygoid processes (Fig. 6, FHP). We identify this as likely the foramen hypophyseos (Goodrich Reference Goodrich1958, p. 240), identical in placement to the largest foramen in the floor of the hypophyseal fossa in dorsal view (Fig. 7, HYP). A foramen in the parasphenoid in this location has a long history in gnathostomes, being present in early arthrodire placoderms such as Kujdanowiaspis (Jarvik Reference Jarvik1954, ‘hypophysial canal'). It is also present in crossopterygian fish preceding the origin of tetrapods such as Porolepis and Eusthenopteron (Jarvik Reference Jarvik1954, 1972 as ‘hypophysial canal'). In early tetrapods, other than Whatcheeria, it has only been identified in Ichthyostega (Jarvik Reference Jarvik1996, as ‘buccohypophysial foramen') and most recently in Lethiscus stocki (Pardo et al. Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017, as ‘buccohypophyseal canal' and placed by their analysis stem-ward to the whatcheeriids). As in Whatcheeria, the foramen is likely to be small and easy to miss if the surface of the parasphenoid is not well preserved or is not finely prepared, and we would expect it to be more widespread than the current literature suggests.
The basal plate of the parasphenoid bears prominent paired ridges, which begin at the level of the basipterygoid processes and extend forward, converging gradually to become the borders of the cultriform process. It might be supposed that this morphology is the result of crushing, for example of a cylinder. This seems not to be the case, as the appearance, symmetry, and thickness variation are consistent between specimens. Immediately anterior to the basipterygoid processes, the cross section of the cultriform process is roughly that of an I-beam laid on its side, with a broad, horizontal web. Going forward, the process narrows and flattens. The process was possibly in contact with the pterygoids anteriorly, but the nature and location of that contact are not directly observable. The limited evidence available indicates that it lay dorsal to the pterygoids anteriorly and contacted them in a simple, flat, bevelled suture. The possibility of vomer–parasphenoid contact cannot be determined.
In dorsal view, the basiparasphenoid in SUI 52055 (Fig. 7) appears nearly uncrushed, and is almost schematically simple. A short and thin crista sellaris (as defined in Goodrich Reference Goodrich1958, p. 240), tilted slightly posteriorly, connects a pair of low and massive protuberances with unfinished dorsal ends. We consider these to be the pilae antoticae (also in Goodrich Reference Goodrich1958, p. 240). They continue posteroventrally for a short distance, which we are identifying as joint surfaces with a missing basioccipital. The crista shows a finished anterior surface and unfinished posterior surface, which was presumably continued in cartilage and may also have been part of the joint with the basioccipital. Immediately anterior to the crista sellaris, the surface of the basisphenoid is a shallow depression floored by finished bone bearing a number of tiny foramina, but otherwise featureless. Neither the depression, the anterior wall of the crista sellaris, nor any other part of the basisphenoid's dorsal surface bears markings that could be interpreted as muscle scars (e.g., for extrinsic eye muscles). Immediately anterior to the depression and to the basipterygoid processes, and dorsal to the base of the cultriform process, is a pair of closely spaced, very low protuberances with an unfinished dorsal surface. We interpret these as the ossified bases of the pilae metopticae. Between them is a narrow fossa that deepens posteriorly and is roofed over opposite to the posterior borders of the pilae metopticae. The fossa and a foramen in its base penetrate for an unknown distance into the basisphenoid. Several small foramina open into its anterior floor. We identify this as the hypophyseal fossa, consistent with placement of the ‘sella turcica' in Eryops (Sawin Reference Sawin1941), Edops (Romer & Witter Reference Romer and Witter1942), Eogyrinus, (Panchen Reference Panchen1972, brought into synonymy with Pholiderpeton by Clack Reference Clack1987), and Greererpeton? (Smithson Reference Smithson1982, as considered in an extended discussion in Clack & Homes Reference Clack and Holmes1988). In Whatcheeria, the placement of this fossa lines up with the putative foramen hypophyseos in ventral view (Fig. 6, FHP). No other significant foramina are visible in SUI 52055, which might provide for passage of the internal carotid artery or cranial nerves.
PR 1654, PR 1655, and PR 1888 show a large dorsum sellae borne on the crista sellaris, shaped as an equilateral triangle with the apex pointing forward and unfinished dorsal surface. It is especially marked in PR 1655, which appears to show a midline process directed anteriorly (cf. Robinson et al. Reference Robinson, Ahlberg and Koentges2005, in Dendrerpeton). Due to damage, the extent of this process in Whatcheeria is unknown. What is visible of the crista sellaris in these specimens is apparently somewhat sinuous, rather than straight.
2.4. Occiput and otic capsules
As noted in Section 2.3, we have not recovered a basioccipital in recognisable form as a separate element. In addition, we can only tentatively identify exoccipitals, even in articulated specimens where the basioccipital is present. This may be due to small sample size, or it may be due to absent sutural contact. In view of the generally primitive status of Whatcheeria, it is possible that there was still a metotic fissure anterior to the exoccipital – but this is speculative.
In ventral view, the best example of a basioccipital is PR 1701 (Fig. 6), but even here the basioccipital is heavily crushed and incomplete, being partially preserved only on the left side. Due to crushing, the shape of the condyle is uncertain. Similarly, neither can it be determined whether the condyle was notochordal, nor the extent (if any) of exoccipital participation. This specimen shows poorly ossified bones on either side of the basioccipital, which are likely otic capsules, but this identity cannot be confirmed. The best preserved but partial occiput, PR 1809 (skull) (Lombard & Bolt Reference Lombard and Bolt1995, text-fig. 2), is a considerably larger specimen, about 30% wider across the tabulars. It preserves the foramen magnum except for its ventral border. The dorsal and lateral borders of the foramen magnum appear to be unfinished or perhaps damaged, everywhere except at their lower lateral portions. These regions are separated from the otic capsule by symmetrical sutures (or cracks). We tentatively suggest that these are the exoccipitals, or at least part of them. These bones are certainly somewhat damaged, especially ventrally, but if correctly identified they suggest that the exoccipital played little role in the occipital condyle.
The otic capsule is partially represented in two specimens. PR 1652 preserves the dorsal portion of both otic capsules, although distorted and displaced to the left. Bone of this specimen has an eroded appearance, especially the inner surface of the otic capsules. In this specimen, the capsules are joined by a band of eroded bone/calcified cartilage apparently attached to the internal surface of the skull roof. This may either be the adult derivative of the tectum synoticum or the fusion of the otic capsules dorsal to the cranial cavity. PR 1809 (skull) preserves the otic capsules more completely, although they are severely cracked and are obscured by displaced cranial bones. These capsules appear to be continuous across the top of the foramen magnum, but there is no evidence for a separate supraoccipital. As noted previously (Lombard & Bolt Reference Lombard and Bolt1995), and in a composite restoration for Whatcheeria and Pederpes by Clack (Reference Clack2003a), the capsules would appear to exclude the exoccipitals from contact with the skull roof. Whether the exoccipitals were co-ossified with the basioccipital is uncertain.
Neither PR 1652 nor PR 1809 (skull) shows any sign of separate pro- and opisthotic, which is especially convincing in PR 1652 because, there, the preserved dorsal portion of the entire left capsule is visible. PR 1809 (skull) shows strong paroccipital processes, although broken (or unossified) distally and somewhat distorted by crushing; neither paroccipital process is in contact with the tabular. The occipital surface of both paroccipital processes is flat and high (about 4mm on the left, which is better preserved). Details of the contact between otic capsule and skull roof are uncertain. In PR 1809 (skull) (Lombard & Bolt Reference Lombard and Bolt1995, text-fig. 2), the distal end of the paroccipital process on both sides is separated from a strong ridge that originates at the posterolateral ventral corner of the tabular. On the right side the ridge appears to extend onto the ventral surface of the postparietal. In the modest-sized skull table PR 1651 (width 5.5cm), most of the underside of the tabular is occupied by a raised facet with unfinished surface, which is directed toward the paroccipital process. In the absence of direct evidence of contact, we conclude that the facet was the contact point for the paroccipital process. In any case, the posttemporal fossae must have been quite small.
On both sides of PR 1809 (skull), much of the occipital surface of the otic capsule dorsal to the posttemporal fenestra is occupied by a shallow, roughly quadrangular depression. There is a large foramen at the top centre of this depression, with that on the right side being the larger. We originally (Lombard & Bolt Reference Lombard and Bolt1995) identified these foramina as the posttemporal fenestrae; as pointed out by Clack (Reference Clack2003a), this was incorrect. Clack suggested that these foramina were actually for the occipital arteries, with which we agree. There are several much smaller foramina within the depressed areas; we count three on the left, two on the right. Their function is unclear, although some might be nutrient foramina.
2.5. Stapes
Two skull specimens have stapes more or less in place; no other stapes have been found. None of these are well-preserved, but the two stapes associated with PR 1651 yield some intriguing information. PR 1651 preserves a braincase in ventral view, crushed up against the underside of an isolated skull table. This braincase is slightly displaced to the left from its original position and preserves at least the proximal portion of both stapes approximately in normal contact with the braincase, although strongly crushed. The left stapes ends in contact with the facet for paroccipital process on the underside of the tabular. The left stapedial head is covered by the ‘crista ventrolateralis' almost up to the level of the large stapedial foramen, which is on the current ventral side. The right stapes falls short of the tabular facet, in part because of movement of the braincase. Rotatory position of these stapes can be interpreted by a simple convention: if a stapedial canal is present, it runs approximately antero-posteriorly (cf. Bolt & Lombard Reference Bolt and Lombard1985). The left stapes shows a large stapedial foramen, so, according to this convention, it appears to have been rotated by about 90° around its shaft. The right stapes is evidently not so rotated, as it does not show a stapedial foramen, despite preservation that should show it if present. Both stapes appear to show a small extension of the unfinished bone of the terminus, onto the stapedial shaft. Otherwise, both stapes apparently comprise finished bone. The right stapes shows slight constriction of the shaft, just distal to the footplate; but as preserved, neither shows a ‘plate-like' expansion of the shaft. This may well have been present distally; but whether they did or not, the point of interest here is that the stapes of Whatcheeria shows more of a distinct shaft than is generally present in other tetrapods of this age or older, including Pederpes finneyae (Clack Reference Clack, Webster, Fay and Popper1992; Clack & Finney Reference Clack and Finney2005). The Viséan aïstopod Lethiscus also has a short-shafted stapes (Pardo et al. Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017).
PR 1816 contains a braincase in ventral view, preserved as part of an incomplete skeleton. A crushed and poorly preserved right stapes is in articulation with the braincase immediately posterior to the basipterygoid process. This position aligns it with the likely placement of the (not visible) fenestra ovalis/fenestra vestibuli. The shaft extends laterally to contact the raised facet for the right paroccipital process on the underside of the tabular. The braincase + stapes is displaced about 2cm to the left relative to the skull table and the rotational orientation of the stapes is uncertain: a stapedial foramen appears to be present in ventral view, but this is uncertain due to the crushed condition of the stapedial footplate. The shaft is finished bone, on the (current) anterior and posterior sides, except that there is a strip of unfinished bone along the distal half of the (current) ventral edge (but well short of the foramen). The finished edges of this strip appear to be broken.
3. Discussion
The late Tournaisian Pederpes is morphologically similar to Whatcheeria and, together, they comprise the Whatcheeriidae (Clack Reference Clack2002a; Clack & Finney Reference Clack and Finney2005). The mid-Viséan Ossinodus pueri may/may not represent a third member of the family (Warren Reference Warren2007). The palate of Pederpes is more primitive than that of Whatcheeria in that the vomer, palatine, and ectopterygoid are covered with denticles. Otherwise, the details that are preserved in Pederpes do not present substantive compositional differences to those of Whatcheeria. The numerous specimens of Whatcheeria, however, allow a more complete reconstruction of the whatcheeriid palate than was previously possible. Four aspects in particular are brought into better focus: the general shape of the skull; the structure of the anteriormost palate; the choanae; and the occlusal pattern of the dentition on the lower jaw and palate.
3.1. General shape of the skull
With the palatal ramus of the pterygoid in the horizontal plane, the maximum estimate of skull width is controlled mainly by the breadth of the pterygoid at its greatest width from the midline. In our reconstruction with palatal ramus horizontal, the skull of Whatcheeria is very narrow; a rectangle encompassing this outline has a length about 2.2 times its width comparable to that of Crassigyrinus (1.9) as reconstructed by Clack (Reference Clack1998) and Proterogyrinus (2.0) as reconstructed by Holmes (Reference Holmes1984) (Fig. 8). The ratio in Whatcheeria can be seen to be very high compared to the tentative reconstruction based on the less complete whatcheeriid Pederpes, at 1.3 (our measurements; Clack & Finney Reference Clack and Finney2005, fig. 17), though Clack's earlier representation of a composite Pederpes/Whatcheeria in occipital view hints at a narrower skull (Clack Reference Clack2003a, fig. 10b); that is, Pederpes has been reconstructed as broad-headed with horizontal palatal bones partially controlling skull width, though the controlling factors are not specified. The ambiguity about palatal ramus orientation and its role in controlling skull width is universal in early tetrapod palate reconstructions. To further highlight the very narrow skull in Whatcheeria, all Palaeozoic tetrapods with a ratio equal to or greater than 1.4 gathered in preparation for Kimmel et al. (Reference Kimmel, Sidlauskas and Clack2009) are listed in Table 2 (Clack, personal communication). Ratios of less than 1.4 are characteristic of broad-headed, relatively flatter skulls, such as reconstructed for the very early tetrapods Tiktaalik (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006), Ichthyostega (Jarvik Reference Jarvik1996), and Acanthostega (Clack Reference Clack2002b).
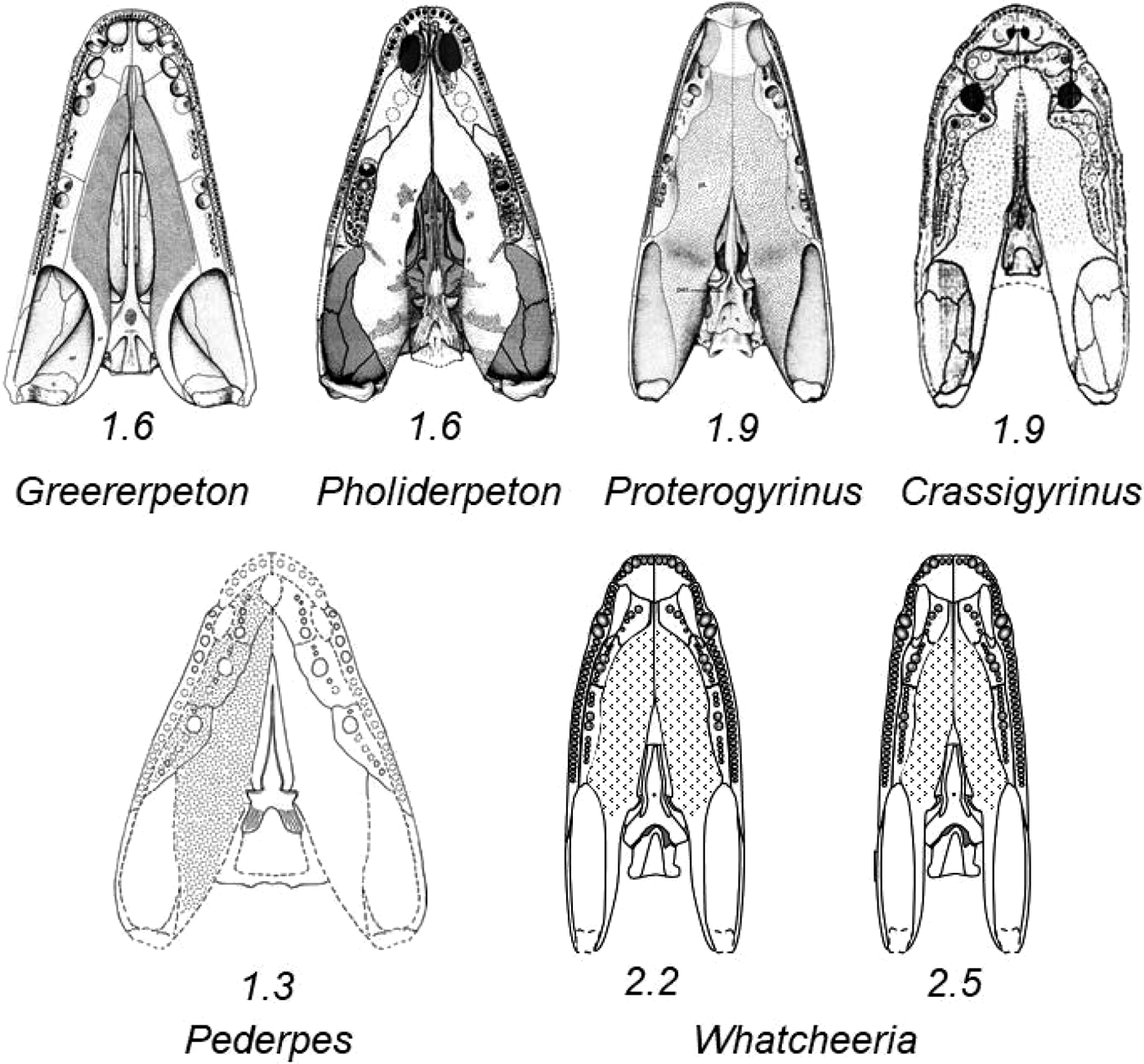
Fig. 8 Palatal reconstructions of selected early tetrapods. All reproduced to the same longitudinal dimension of a best-fit encompassing rectangle. Length-to-width ratios calculated from rectangle: Whatcheeria at 2.2, with all palatal bones in horizontal plane; Whatcheeria at 2.5, with ectopterygoid sloping ventrally away from midline to produce slightly vaulted palate. Illustrations of other taxa lightly edited from originals: Greererpeton, Smithson (Reference Smithson1982); Pholiderpeton, Clack (Reference Clack1987); Proterogyrinus, Holmes (Reference Holmes1984); Crassigyrinus, Clack (Reference Clack1998); Pederpes, Clack & Finney (Reference Clack and Finney2005).
Table 2 Length-to-width (L:W) ratio of 1.4 or greater for Palaeozoic tetrapod skulls. Data shared by J. Clack, gathered in the preparation of, but not published in, Kimmel et al. (Reference Kimmel, Sidlauskas and Clack2009). Some ratios differ by a tenth from those determined by us for this paper (Fig. 8).

It is possible that the skull of Whatcheeria is actually narrower than the 2.2 (Figs 1, 8). A narrower skull would result from a vaulted palate in which the pterygoid is not horizontal but, instead, still flat, but slightly angled ventrally away from the midline, or ventrally concave away from the midline. The resulting skull could have an aspect ratio of more like 2.5 (Fig. 8). The orientation of the palatal shelf in SUI 147644 suggests this possibility (Fig. 3a, b). Such a reorientation or shaping could also result from either shape or positioning changes to the braincase. In addition, a narrower skull would result from direct contact between the pterygoid and maxilla at the anterior end of the adductor fossa. In the reconstruction (Fig. 1), the posterior end of the ectopterygoid has been placed between the two, for which there is a hint in skull PR 1792. Braincase PR 1701 (Fig. 6), incorporated in our reconstruction, is dorso-ventrally compressed so that the basipterygoid joints are further from the midline than they would have been in life. Bringing the joints closer to the midline would result in a narrower skull. In addition, restoration of the original three-dimensionality of the braincase would raise the basipterygoid joints dorsally above the plane of the base of the basiparasphenoid with a similar result to the orientation of the pterygoids. Finally, the exact dorsoventral location of the braincase within the skull is unknown. No specimen preserves sufficient morphology to provide a control for its position. Our restoration with ratio 2.2 positions the braincase such that the ventral surface is coplanar with the rest of the palate. The 2.5 ratio restoration positions the ventral surface of the braincase slightly dorsally to the marginal palatal bones.
More or less complete skulls of Whatcheeria are preserved as laterally compressed, with the palatal elements and braincase sandwiched between the lateral surfaces and the skull roof splayed out to one side (Lombard & Bolt Reference Lombard and Bolt1995, pl. 1, text-fig. 1). This configuration suggests another feature of its skull shape. Not only is it narrow in the frontal plane, but also tall in the sagittal plane – a combination that favours the preservation typically found. Skull reconstructions of many primitive Palaeozoic tetrapods present a height-to-length (H:L) ratio of less than 0.30. Typical examples are: Acanthostega, 0.27 (Clack Reference Clack2003b); Proterogyrinus, 0.25 (Holmes Reference Holmes1984); and Greererpeton, 0.21 (Smithson Reference Smithson1982). Lethiscus (Pardo et al. Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017) has a long and narrow skull, but its H:L ratio is clearly quite small. In contrast, Pholiderpeton (Clack Reference Clack1987) and Crassigyrinus (Clack Reference Clack1998), both at 0.37, have considerably higher skulls. We estimate Whatcheeria at 0.38. We realise, of course, any such estimates are generally degraded by the imprecise measurements that must be made on distorted specimens. We think it worthwhile though to consider other early tetrapods that also present a form of preservation similar to Whatcheeria as candidates for skulls that are both narrow and tall. These taxa would include, in addition to Whatcheeria and Crassigyrinus, Pederpes, Proterogyrinus, and Archeria (Fig. 9). Aïstopods are also consistent with this narrow and elongate skull form (Pardo et al. Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017). Among the continuum of possibilities, Whatcheeria (and Crassigyrinus) are either exceptional in the proportions of their skulls and those with a similar preservation pattern convey no meaning about skull shape, or that pattern of preservation conveys important information about skull shape in any taxon in which it is found. Such taxa might benefit from renewed consideration of overall proportional shape, and a general contemplation of skull shape in early tetrapods is also suggested.

Fig. 9 Specimen illustrations of early tetrapods with a particular pattern of preservation. All are modified from the original. Images are reproduced to approximately the same posterior skull roof width with anterior to the left, though two are reversed to conform to this orientation. Original labelling has been removed. Elements of the skull roof are dark grey and parts of the lateral cheek and snout are light grey. Whatcheeria, Field Museum PR 1634 from Lombard & Bolt (Reference Lombard and Bolt1995, text-fig. 1); Crassigyrinus, British Museum R10000 ‘Cowdenbeath skull' from Panchen (Reference Panchen1985, fig. 3), British Museum BMNH 30532 in Clack (Reference Clack1998) has the preservation pattern common to broad and low skulls; Pederpes, Hunterian Museum GLAHMS 100815 from Clack & Finney (Reference Clack and Finney2005, fig. 4); Archeria, Museum of Comparative Zoology MCZ 2049 (reversed) from Holmes (Reference Holmes1989, text-fig. 7); Proterogyrinus, Museum of Comparative Zoology MCZ 4537 (reversed) from Holmes (Reference Holmes1984, fig. 8).
3.2. The anterior palate and choana
In early tetrapod fossils, the bones forming the tip of the snout – the premaxillae and vomers – are very often crushed and distorted at best, or undecipherable in detail, or missing at worst. It follows that the morphology of the internal choana and external naris, the details of the dentition, and presence or shape of any anterior palatal fenestra(e) or fossa(e) are also difficult or impossible to reconstruct with certainty, if at all. In consequence, many skull reconstructions, especially those of the palate, are graced with dotted outlines as unsupported by interpretable specimens, such as in Pederpes (Fig. 8). The premaxillae, anterior ends of the vomers, and the anterior processes of the pterygoids are missing in the material of Pederpes presently available (Clack & Finney Reference Clack and Finney2005, fig. 18b). Though still not permitting a reconstruction absent any doubt, Whatcheeria provides for a reasonable presentation because several premaxillae, much of the anterior end of the vomers, the anterior processes of the pterygoids, and the articulated anterior end of the palate in PR 1814 (Fig. 2a) are preserved. This material indicates that Whatcheeria has an extensive premaxillary shelf with no evidence of an anterior palatal fenestra or fossa, and probably no room for either. Among early tetrapods, a well-developed palatal shelf is present only in Proterogyrinus, which also lacks an anterior palatal fenestra (Holmes Reference Holmes1984).
Together, PR 1814 (Fig. 2a) and the several freestanding maxillae allow reconstruction of the choana as both elongate and possibly positioned so as to receive the dentary fang teeth (Fig. 10).
3.3. Areas of uncertainty in the palate
There are four areas of uncertainty in the palate: the possible degree of palatal vaulting; the details of the intersection of the pterygoid palatal processes with the vomers and premaxillary shelf, as well as with the cultriform process in that region; the details of the bones involved in the anterior border of the adductor fossa; and the assembly of the quadrate with ventral borders of the quadratojugal laterally and quadrate process of the pterygoid medially.
3.4. Occlusion of jaw–palate dentition
Here we mean simply the spatial relationships of the upper and lower teeth to each other and to non-dental structures. The general occlusal pattern of teeth (excluding denticles) in the majority of early tetrapods is clear enough: from lateral to medial, are premaxillary/maxillary, dentary, lateral palatal, and coronoid teeth (e.g., Jarvik Reference Jarvik1980, fig. 94). In the course of determining the shape of the Whatcheeria snout in palatal view, we reconstructed the opposing sides of the lower jaw to the appropriate size for a given skull size. Dental occlusion in Whatcheeria, as we reconstruct it, includes a close relationship between the dentary fang and choana (Fig. 10). This reconstruction resulted from detailed tracings of maxillary, dentary, and adsymphyseal tooth positions at the front of the skull, the latter two from PR 1809 (jaw) (Lombard & Bolt Reference Lombard, Bolt, Carrano, Gaudin, Blob and Wi2006, fig. 2.2). This suggests the possibility that the large dentary fang on each side might have passed into or at the edge of the choana with the mouth closed. This is not as far-fetched as it seems: some osteolepiform fish, and even some tetrapods, accommodate palatal fangs in intercoronoid fossae in the lower jaw, or dentary fangs in anterior palatal fenestrae or fossae, for example. This is simply analogous, and indicates that the choana might not have been exclusively devoted to water sampling/respiration. That the fit is not perfect is likely due to the imprecise nature of reconstruction, but it is a fact that the very large dentary fangs would need some accommodation for jaw closure.

Fig. 10 Whatcheeria deltae. Schematic reconstruction of occlusal pattern. Palatal tooth positions, which would be projecting out of the plane of the illustration, are black circles on outlines of palatal bones; pterygoid shagreen not shown. Jaw tooth positions are open circles that would be projecting down into the plane of the illustration. The bones of the jaw and the prearticular shagreen are not shown. The marginal dentition of the premaxilla and maxilla are labial to the marginal dentition of the dentary. The large dentary fangs are related to the choana. The dentition of the vomer, palatine, and ectopterygoid are lingual to the marginal dentitions. Lingual to all palatal dentition are the teeth of the adsymphyseal and coronoids 1–3 from anterior to posterior. Abbreviations: ADF = adductor fossa; CHO = choana; ECP = ectopterygoid; MAX = maxilla; PAL = palatine; PMX = premaxilla; PTY = pterygoid; VOM = vomer. Tooth position spacing on the palatal bones based on PR 1792 and PR 1814 (Fig. 2) with contributions from other specimens. Tooth position spacing on the jaw from Lombard & Bolt (Reference Lombard, Bolt, Carrano, Gaudin, Blob and Wi2006).
3.5. Evolution of the stapes
A number of primitive-tetrapod stapes have now been described, including those of Greererpeton burkemorani (Smithson Reference Smithson1982), Pholiderpeton scutigerum (Clack Reference Clack1983), Acanthostega gunnari (Clack Reference Clack1989), Kyrinion martilli (Clack Reference Clack2003a), Ichthyostega sp. (Clack et al. Reference Clack, Ahlberg, Finney, Dominguez Alonso, Robinson and Ketcham2003), and Pederpes finneyae (Clack & Finney Reference Clack and Finney2005). Expectations for the morphology of the stapes in early tetrapods were at one time set by the first such stapes to be described, namely that of Greererpeton (Carroll Reference Carroll and Panchen1980; Smithson Reference Smithson1982). In this model, the shaft was expected to be expanded (‘plate-like'), beginning slightly distal to the stapedial foramen, and with an unfinished distal margin around most/all of the plate. The plate was applied to the posterior surface of the pterygoid/epipterygoid quadrate ramus. Such a stapes was presumed to have little or no role in perceiving airborne sound. Most of the early tetrapod stapes described so far fit this model quite well. This definitely included Pederpes finneyae (Clack & Finney Reference Clack and Finney2005). The stapes of Ichthyostega is the first known exception: it is sui generis and is associated with a uniquely specialised middle ear.
The stapes of Whatcheeria may be another exception to expectations. Not only is there no sign of a plate-like expansion, the short strips of unossified bone along the distal part of the shaft begin well distal to the stapedial foramen. The shaft itself appears to consist of finished bone, and we believe that the three specimens available allow inspection of the entire circumference of the shaft. Of course, we cannot say what the morphology of an intact stapes may have been. But the present picture is that the stapes either lacks the expanded plate, or the expansion began considerably distal to the expected proximal point. The Viséan aïstopod Lethiscus appears to fit this model quite well (Pardo et al. Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017).
Given the damaged condition of all available stapes, our suggestion is plausible but not certain. If the Whatcheeria stapes did have the suggested morphology, it may be an early example of evolution of a more ‘rod-like' stapes. As has been suggested for the fenestra ovalis (e.g., Clack Reference Clack1994), the ‘rod-like' stapes probably arose more than once.
3.6. Position of Whatcheeria in the tetrapod phylogeny
It has been obvious since its original description (Lombard & Bolt Reference Lombard and Bolt1995) that Whatcheeria is a very primitive tetrapod. Clack (Reference Clack2002a) described Pederpes finneyae from the Tournaisian of Scotland and included a phylogeny that placed Pederpes and Whatcheeria as sister taxa in a new family, Whatcheeriidae, near the base of the tetrapod tree. In 2005, Clack & Finney published a full description of Pederpes, with a strict-consensus tree that also showed Pederpes and Whatcheeria as sister taxa, still very near the base of the tetrapod tree. That phylogeny was based on more data than the initial one, and (unsurprisingly) differed from Clack's original one in some respects. The position of Whatcheeria and Pederpes was, however, nearly identical to Clack's original finding. The contribution to the whatcheeriid data matrix was based primarily on Pederpes, but included Whatcheeria cranial and postcranial data, from the literature and Clack's own observations. The most recent phylogenies of Clack et al. (Reference Clack, Bennett, Carpenter, Davies, Fraser, Kearsey, Marshall, Millward, Otoo, Reeves, Ross, Ruta, Smithson, Smithson and Walsh2016) and Pardo et al. (Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017), which include citations to all preceding relevant phylogenies, confirm Whatcheeria and Pederpes as sister taxa. In addition, their common stem is among the earliest and generally falls in sequence by geological age among the earliest tetrapod taxa. The constant linkage of Whatcheeria and Pederpes and their placement with respect to other early taxa is not surprising given that the characters available, and our understanding of them, have changed very little since the initial descriptions of Lombard & Bolt (Reference Lombard and Bolt1995) and Clack & Finney (Reference Clack, Ahlberg, Finney, Dominguez Alonso, Robinson and Ketcham2005). The present study adds new morphological details and in the Appendix we have inserted the states for Whatcheeria palatal characters in the most recent character list available (Pardo et al. Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017). Previously known states for the cranial and postcranial skeleton have been added to the compilation as well. We do not present a phylogenetic hypothesis as the new information is not likely to make a difference to the general understanding now available. Additional cranial characters or character states for Whatcheeria are likely to be relatively few in the future, but the postcranium currently under study by ourselves and B. Otoo has the potential to add important new information.
4. Acknowledgements
W. Simpson, H. Rittenhouse, A. Leman, R. Masek, S. McCarroll, and A. Shinya prepared and repaired the numerous specimens of Whatcheeria. We are very grateful to them for their cheerful and exceptionally skilful work. The preparation and conservation work was supported by the Field Museum of Natural History and the National Science Foundation through grants DEB 9207475 and DEB 9306294 to the authors. The National Geographic Society supported the initial field work through a grant to J. Bolt. We thank the University of Iowa for the loan of specimens, and the many members of field crews for their assistance, without which such an extensive collection could not have been amassed. We appreciate the assistance and interest of the late Jasper Hiemstra, owner of the Delta quarry. Lori Grove and Marlene Donnelly skilfully created the included illustrations and we very much appreciated their willingness to accommodate our change requests as our knowledge increased. We are grateful for the comments of two reviewers and especially appreciate the insight offered on the tilt of the premaxillary shelf.
5. Appendix
This list of 370 characters is that compiled for the phylogenetic analysis of Pardo et al. (Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017). Their original character numbers are retained, with text edited for consistent punctuation and capitalisation. The original source of individual characters within the list is noted via an acronym as per Pardo et al.:
CABF: Clack, Ahlberg, Blom, and Finney (2012)
HPSA: Huttenlocker, Pardo, Small, and Anderson (Reference Huttenlocker, Pardo, Small and Anderson2013)
MA: Maddin & Anderson (Reference Maddin and Anderson2012)
PSAA: Pardo, Szostakiwskyi, Ahlberg, and Anderson (Reference Pardo, Szostakiwskyj, Ahlberg and Anderson2017)
Modifications of a few character states to accommodate Whatcheeria are noted as BL: Bolt & Lombard.
States determined for Whatcheeria are in bold. Characters where a state for Whatcheeria might be determined with better specimens, or have yet to be determined where work is in progress, are indicated as Not determinable. Characters where no state listed fits Whatcheeria are indicated as Not applicable.
1. Basal Skull Length (HPSA 001): (0) >70mm; (1) 50–70mm; (2) 30–50mm; (3) <30mm.
2. Skull:trunk ratio (HPSA 002): (0) 0.45; (1) 0.30–0.45; (2) 0.20–0.29; (3) 0.20. Not determinable.
3. Skull proportions (HPSA 003): (0) longer than wide; (1) wider than long.
4. Intertemporal (PSAA 4): (0) present; (1) replaced by anterior extension of supratemporal or tabular; (2) replaced by lateral extension of parietal. (PSAA note: this character serves as a replacement for HPSA 004, 007, 008, 050, 051, and 052.)
5. Supratemporal (HPSA 005): (0) present; (1) absent.
6. ST exposure on occiput (HPSA 006): (0) absent; (1) present.
7. Postfrontal shape (HPSA 009): (0) broadly quadrangular; (1) falciform.
8. Squamosal–tabular contact (HPSA 010): (0) absent; (1) present; (2) fused.
9. Lacrimal–prefrontal suture (HPSA 012): (0) simple butt joint; (1) interdigitating; (2) prefrontal broadly underplates lacrimal. Not determinable.
10. Lacrimal (HPSA 013): (0) present; (1) absent.
11. Lacrimal extends to naris (HPSA 014): (0) present; (1) absent.
12. Lacrimal extends to orbit (HPSA 015): (0) absent; (1) present.
13. Lacrimal orbital processes (HPSA 016): (0) only ventral present; (1) dorsal and ventral present; (2) neither present. Not applicable.
14. Lacrimal–jugal contact (HPSA 017): (0) present; (1) absent.
15. Quadratojugal (HPSA 018): (0) present; (1) absent.
16. Quadratojugal–jugal contact (HPSA 019); (0) present; (1) absent.
17. Quadratojugal–maxillary contact (HPSA 020): (0) present; (1) absent. Not determinable.
18. Frontals (HPSA 021): (0) paired; (1) fused.
19. Frontal into orbital margin (HPSA 022): (0) no; (1) yes.
20. Anterior laterally flaring frontals (HPSA 023): (0) absent; (1) present.
21. Nasals (HPSA 024): (0) present; (1) absent.
22. Narial flange (HPSA 025): (0) absent; (1) present.
23. Alary processes of premaxilla (HPSA 026): (0) absent; (1) present.
24. Septomaxilla (HPSA 028): (0) ossified; (1) unossified.
25. Prefrontal into external narial margin (HPSA 029): (0) distant from; (1) near; (2) present.
26. External naris in dorsal view (HPSA 030): (0) exposed; (1) not exposed. Not determinable.
27. External naris shape (HPSA 031): (0) circular; (1) posteriorly extended, along lacrimal–prefrontal suture; (2) posteriorly extended excavation of lacrimal only. Not determinable.
28. Dorsal exposure of premaxilla (HPSA 032): (0) broad pars dorsalis anteromedial to external naris; (1) pars dorsalis limited, but naso–premaxillary suture exposed dorsally; (2) none.
29. Dorsal shape of skull (HPSA 033): (0) triangular; (1) diamond; (2) rounded.
30. Posterior skull margin (HPSA 034): (0) concave; (1) straight; (2) convex; (3) undulating.
31. Snout shape (HPSA 035): (0) blunt; (1) pointed.
32. Snout length (HPSA 036): (0) less than ¼ total skull length; (1) greater than ¼ total skull length.
33. Quadrate articulates along internal flange of squamosal (HPSA 037): (0) absent; (1) present. Not determinable.
34. Otic/spiracular notch (HPSA 038): (0) present; (1) absent.
35. Large otic notch approaching orbit (HPSA 039): (0) more than ½ postorbital length of skull; (1) between ¼ and ½ postorbital length of skull; (2) less than ¼ postorbital length of skull.
36. Otic notch structure (HPSA 040): (0) open posteriorly; (1) closed posteriorly.
37. Semilunar flange of supratemporal (HPSA 041): (0) absent; (1) present.
38. Supratympanic flange (HPSA 042): (0) absent; (1) present.
39. Supratympanic shelf (HPSA 043): (0) absent; (1) present.
40. Raised orbital rim (HPSA 044): (0) absent; (1) present.
41. Postorbital (HPSA 045): (0) present; (1) absent.
42. Jugal–postorbital interfingered processes (HPSA 046): (0) absent; (1) present.
43. Postorbital participates in orbital margin (HPSA 047): (0) present; (1) absent.
44. Shape of postorbital (HPSA 048): (0) irregular trapezoid; (1) triangular, apex caudal.
45. Palpebral ossifications (HPSA 049): (0) absent; (1) mosaic of bony plates in orbit (PSAA new state); (2) single large plate above orbit (new state).
46. Parietal–tabular contact (HPSA 052): (0) absent; (1) present.
47. Postparietals (HPSA 053): (0) paired; (1) fused; (2) absent.
48. Parietal foramen (HPSA 054): (0) present; (1) absent.
49. Postparietal size (HPSA 055): (0) much smaller than parietals; (1) approximately as large or larger than parietals.
50. Postparietal squamosal contact (HPSA 056): (0) absent; (1) present.
51. Postparietal length (HPSA 057): (0) large, quadrangular; (1) abbreviated anteroposteriorly, elongate lateral rectangle.
52. Squamosal–jugal contact (HPSA 058): (0) present; (1) absent.
53. Tabular (HPSA 059): (0) present; (1) absent.
54. Posterolateral projection from lateral margin of tabular above squamosal embayment (HPSA060): (0) absent; (1) present.
55. Tabular horns (HPSA 061): (0) absent; (1) present.
56. Tabular horns shape (HPSA 062): (0) parallel or slightly divergent; (1) widely divergent.
57. Squamosal forms base of tabular horn (HPSA 063): (0) absent; (1) present.
58. Lateral line canal grooves (HPSA 064): (0) present; (1) absent.
59. Dermal sculpturing (HPSA 065): (0) circular pits; (1) shallow ridges and grooves; (2) little to none; (3) tuberculate.
60. Premaxilla anterior margin (HPSA 066): (0) vertical; (1) overturned.
61. Maxilla into external naris (HPSA 068): (0) present; (1) absent.
62. Maxilla entire ventral naris (HPSA 069): (0) absent; (1) present.
63. Maxilla (HPSA 070): (0) longer than palatine; (1) shorter than palatine.
64. Marginal teeth orientation (HPSA 071): (0) vertical; (1) turned medially.
65. Marginal teeth largest anterior (HPSA 072): (0) absent; (1) present.
66. Marginal teeth shape (HPSA 073): (0) pointed pegs; (1) blunt pegs; (2) large cones.
67. Number of premaxillary teeth (HPSA 074): (0) 10–20; (1) 5–9; (2) <5; (3) 20 or more.
68. Number of maxillary teeth (HPSA 075): (0) 30–40; (1) 20–29; (2) 15–19; (3) <15; (4) >40.
69. Teeth laterally compressed (HPSA 076): (0) no; (1) yes.
70. Enlarged teeth mid toothrow (maxillary) (HPSA 077): (0) absent; (1) present.
71. Teeth (HPSA 078): (0) simple points; (1) multiple cusps.
72. Multiple cusp orientation (HPSA 079): (0) labio-lingual; (1) antero-posterior. Not applicable.
73. Enamel fluting (HPSA 080): (0) absent; (1) present.
74. Labyrinthine infolding (HPSA 081): (0) present; (1) absent.
75. Jaw articulation (HPSA 087): (0) posterior to occiput; (1) even with occiput; (2) anterior to occiput; (3) far anterior (>20% basal skull length).
76. Internal nares (HPSA 088): (0) widely separated; (1) narrowly separated.
77. Lateral exposure of the palatine (LEP) (HPSA 091): (0) absent; (1) present.
78. Anterior palatine (HPSA 092): (0) short anteromedial process articulating with vomer at choana; (1) long anteromedial process more medial than lateral; (2) palatine absent.
79. Teeth on pterygoid (HPSA 098): (0) absent; (1) present.
80. Tooth pedicely (HPSA 099): (0) absent; (1) present.
81. Palatal teeth size (HPSA 103): (0) larger than marginal teeth; (1) equal to marginal; (2) smaller than marginal.
82. Parasphenoid (HPSA 104): (0) medial of stapes; (1) under footplate of stapes. Not determinable.
83. Interpterygoid vacuities (HPSA 115): (0) narrow or closed; (1) wide; (2) fused at midline.
84. Anterior extent of pterygoids (PSAA 84): (0) pterygoids contact anteriorly; (1) pterygoids exclude vomer from interpterygoid vacuity but excluded from median contact by parasphenoid; (2) pterygoids contact vomer but do not exclude vomer from interpterygoid vacuity; (3) pterygoids reach anteriorly only as far as palatine; (4) pterygoids reach ectopterygoid only; (5) pterygoids do not contact lateral palatal bones at all. (PSAA note: this character serves as a replacement for HPSA 116, 118, and 119.)
85. Lateral process of pterygoid into posttemporal fossa (HPSA 120): (0) absent; (1) present.
86. Ectopterygoid palatine width (HPSA 122): (0) wider than maxilla; (1) narrower than maxilla.
87. Pharyngeobranchial pouches (HPSA 123): (0) absent; (1) present. Not determinable.
88. Dentary (HPSA 124): (0) tooth row greater than 50% of total jaw length; (1) tooth row less than 50% of total jaw length.
89. Dentary forms coronoid process (HPSA 125): (0) absent; (1) present.
90. Surangular (HPSA 126): (0) subequal in size to angular; (1) substantially smaller than angular; (2) absent.
91. Angular (HPSA 127): (0) approximately as deep as the dentary mid-tooth-row, or less; (1) deeper than dentary mid-toothrow.
92. Number of splenials (HPSA 128): (0) 2; (1) 1; (2) 0.
93. Splenial exposed laterally (HPSA 129): (0) present; (1) absent.
94. Meckelian fossae (HPSA 130): (0) 2 or more; (1) 1; (2) 0.
95. Ventral border of Meckel's fossa (HPSA 131): (0) splenial; (1) angular; (2) splenial, postsplenial, angular, surangular (BL new state).
96. Retroarticular process presence (HPSA 132): (0) absent; (1) present.
97. Retroarticular process shape (HPSA 133): (0) straight; (1) hooked. Not applicable.
98. Vertical position of jaw articulation relative to tooth row (HPSA 134): (0) above; (1) equal; (2) below.
99. Extent of angular in lateral view (HPSA 135): (0) posterior tooth row; (1) middle of tooth row.
100. Number of coronoids (HPSA 136): (0) 3; (1) 2; (2) 1; (3) 0.
101. Splenial participates in symphysis (HPSA 139): (0) yes; (1) no.
102. Jaw sculpture (HPSA 140): (0) present; (1) absent.
103. Ossified hyoids (HPSA 141): (0) present; (1) absent.
104. Gill osteoderms (HPSA 142): (0) absent; (1) present non-interdigitating; (2) toothed interdigitating rakers.
105. Parahyoid (HPSA 143): (0) absent; (1) present.
106. Number of accessory articulations (HPSA 144): (0) 0; (1) 1; (2) 2 or more.
107. Number of presacrals (HPSA 145): (0) 25–35; (1) 20–24; (2) >35; (3) <20.
108. Vertebral development (HPSA 146): (0) arches then centra; (1) centra and arches simultaneously. Not determinable.
109. Caudal processes between depressions (HPSA 147): (0) absent; (1) present.
110. Trunk intercentra (HPSA 148): (0) present; (1) absent.
111. Trunk neural arch to centrum in adults (HPSA 149): (0) loosely articulated; (1) sutured; (2) fused.
112. Base of neural spine (HPSA 150): (0) equal to or wider than haemal; (1) smaller than haemal spine. Not determinable.
113. Height of neural spines (HPSA 151): (0) even; (1) alternating.
114. Dermal armour associated with neural arches (HPSA 152): (0) absent; (1) present. (PSAA note: ‘This character was established to recognise the expanded, sculptured structures seen at the top of neural arches in some nectrideans. Upon further examination, this character represents fusion of osteoderms to the neural arch. As this is present in a number of temnospondyls as well as chroniosuchians, we have amended this diagnosis to allow for comparisons outside nectrideans.')
115. Neural spine shape in lateral view (HPSA 153): (0) anterior and posterior sides parallel, forming a rectangular surface; (1) non-parallel, triangular.
116. Neural spine lateral surface (HPSA 154): (0) smooth; (1) crenulated.
117. Pleurocentra (HPSA 155): (0) paired rhachitomous; (1) closely approaching ventrally; (2) fused dominant weight-bearing element; (3) embolomerous; (4) dorsally fused rhachitomous (BL state added).
118. Haemal arch presence (HPSA 156): (0) present; (1) absent.
119. Haemal arch fusion (HPSA 157): (0) loosely articulated to intercentra; (1) fused to mid length of centrum.
120. Haemal arch length (HPSA 158): (0) longer than or equal to neurals; (1) shorter than neurals. Not determinable.
121. Haemal accessory articulations (HPSA 159): (0) none; (1) one; (2) two. Not determinable.
122. Haemal arch shape (HPSA 160): (0) non parallel triangular; (1) parallel rectangular. Not determinable.
123. Tail termination in skeleton (HPSA 161): (0) tapers; (1) deep with sudden end. Not determinable.
124. Tail length (HPSA 162): (0) elongate equal to or exceeding trunk and skull length; (1) foreshortened markedly shorter than trunk. Not determinable.
125. Trunk arches (HPSA 163): (0) paired; (1) fused.
126. Spinal nerve foramina (HPSA 164): (0) absent; (1) present.
127. Extended transverse processes (HPSA 165): (0) absent; (1) present.
128. Transverse process (HPSA 166): (0) on arch pedicle; (1) on centrum. Not applicable.
129. Atlas axis intercentra (HPSA 167): (0) present; (1) absent.
130. Atlas anterior centrum (HPSA 168): (0) same size as posterior; (1) laterally expanded. Not determinable.
131. Atlas centrum (HPSA 169): (0) multipartite; (1) single notochordal; (2) single odontoid. Not determinable.
132. Atlas neural arch centrum fusion (HPSA 170): (0) loosely articulated; (1) sutured to centrum; (2) fused to centrum. Not determinable.
133. Atlas parapophyses (HPSA 171): (0) on centrum; (1) on transverse process; (2) absent. Not determinable.
134. Atlas neural arch midline fusion (HPSA 172): (0) paired; (1) sutured at midline; (2) fused at midline. Not determinable.
135. Atlas accessory articulation (HPSA 173): (0) absent; (1) zygosphene; (2) zygantra. Not determinable.
136. Proatlantes (HPSA 174): (0) present; (1) absent. Not determinable.
137. Second cervical arch (HPSA 175): (0) more expanded to subsequent; (1) equal to subsequent; (2) shorter than subsequent. Not determinable.
138. Atlas ribs (HPSA 176): (0) one pair; (1) two pairs; (2) absent. Not determinable.
139. Cervical rib distal shape (HPSA 177): (0) spatulate; (1) pointed.
140. Ribs anterior to sacrum (HPSA 178): (0) short; (1) long.
141. Ribs (HPSA 179): (0) elongated and sometimes curved; (1) straight; (2) short simple rod.
142. Costal process at rib head (HPSA 180): (0) absent; (1) present. Not determinable.
143. Number of sacrals (HPSA 181): (0) 1; (1) 2; (2) 3.
144. Sacral parapophysis (HPSA 182): (0) on centrum; (1) on transverse process. Not determinable.
145. Number pairs of caudal ribs (HPSA 183): (0) 5 or more; (1) 4; (2) 3; (3) 2 or fewer. Not determinable.
146. Interclavicle posterior stem length (HPSA 184): (0) no or short; (1) long.
147. Interclavicle posterior stem breadth (HPSA 185): (0) wide; (1) narrow.
148. Interclavicle shape (HPSA 186): (0) diamond-shaped; (1) T-shaped.
149. Interclavicle anterior plate (HPSA 187): (0) broad; (1) narrow.
150. Interclavicle shape (if diamond present) (HPSA 188): (0) broad diamond; (1) narrow diamond.
151. Interclavicle anterior fimbriation (HPSA 189): (0) present; (1) absent.
152. Interclavicle sculpture (HPSA 190): (0) present; (1) absent.
153. Cleithrum head dorsal extent (HPSA 191): (0) aligned along anterior rim of scapula; (1) posterodorsally enlarged head wrapping around dorsal scapula.
154. Cleithrum head size and shape (HPSA 192): (0) dorsally greatly expanded much wider than shaft; (1) simple rod without or slight dorsal expansion.
155. Cleithrum ossification (HPSA 193): (0) ossified; (1) unossified.
156. Cleithrum overall shape (HPSA 194): (0) rounded or pointed dorsally; (1) T- or Y-shaped.
157. Proximal clavicle blades (HPSA 195): (0) widely separate; (1) articulate medially; (2) interdigitate.
158. Supraglenoid foramen (HPSA 196): (0) present; (1) absent. Not determinable.
159. Number coracoid foramina (HPSA 197): (0) none; (1) 1; (2) 2. Not determinable.
160. Scapulocoracoid ossification (HPSA 198): (0) both; (1) scapula only; (2) absent.
161. Torsion in humerus (HPSA 200): (0) absent; (1) less than 80°; (2) more than 80°. Not determinable.
162. Deltapectoral crest (HPSA 201): (0) weak or absent; (1) intermediate; (2) prominent. Not determinable.
163. Supinator process (HPSA 202): (0) absent; (1) present.
164. Humerus length (HPSA 203): (0) long (∼4 trunk centra); (1) short.
165. Radius–humerus ratio (HPSA 204): (0) >0.7; (1) 0.5–0.7; (2) <0.5. Not determinable.
166. Olecranon process (HPSA 205): (0) unossified; (1) ossified.
167. Carpals (HPSA 206): (0) fully or partially ossified; (1) unossified.
168. Basale commune (HPSA 207): (0) absent; (1) present. Not determinable.
169. Number digits manus (HPSA 208): (0) 5; (1) 4; (2) 3; (3) >5. Not determinable.
170. Pelvis (HPSA 209): (0) fused; (1) sutured; (2) poorly ossified.
171. Anteriorly inclined ilium (HPSA 210): (0) absent; (1) present.
172. Iliac blade (HPSA 211): (0) 2 dorsal processes; (1) narrowly bifurcate; (2) single blade.
173. Internal trochanter articulation (HPSA 212): (0) distinct; (1) continuous. Not determinable.
174. Femoral shaft (HPSA 213): (0) robust; (1) slender.
175. Femur (HPSA 214): (0) long; (1) short.
176. Tarsals (HPSA 215): (0) ossified; (1) unossified. Not determinable.
177. Elongate tibiale and fibulare (HPSA 216): (0) absent; (1) present. Not determinable.
178. Number of distal tarsals (HPSA 217): (0) 6; (1) 5 or fewer. Not determinable.
179. Astragalus (HPSA 218): (0) absent; (1) present. Not determinable.
180. Number of digits pes (HPSA 219): (0) 5; (1) 4 or less; (2) >5. Not determinable.
181. Dorsal margin of splenial only contacts first coronoid (HPSA 220): (0) absent; (1) present.
182. Postparietal lappet (HPSA 224): (0) mostly exposed posteriorly; (1) equal posteriorly and dorsally; (2) mostly exposed dorsally. Not determinable.
183. Cheek emargination (HPSA 225): (0) absent; (1) present.
184. Parietal anterior waisting (HPSA 226): (0) absent; (1) present.
185. Parietal width relative to frontal (HPSA 227): (0) greater; (1) equal or less.
186. Trabecula cranii (PSAA 186): (0) without significant median fusion posterior to solum nasi (platytrabic); (1) fused medially posterior to solum nasi to form elongate trabecula communis (tropitrabic). Not determinable.
187. Dorsal trabeculae (PSAA 187): (0) dorsal trabeculae provide dorsolateral bridge between sphenoid region and nasal capsule; (1) dorsal trabeculae absent or incomplete, no dorsolateral bridge between sphenoid region and nasal capsule. Not determinable.
188. Ossification between optic foramen and pila antotica (PSAA 188): (0) ossification complete between optic foramen and pila antotica; (1) pila metoptica and associated cartilaginous taenia unossified. Not determinable.
189. Ossification within columella ethmoidalis (PSAA 189): (0) absent; (1) present. Not determinable.
190. Path of profundus branch of trigeminal nerve (PSAA 190): (0) enclosed in lateral wall of sphenoid region of braincase and exits separately from maxillomandibular branch via series of small foramina; (1) extramural, exits antotic fissure with other branches of trigeminal. Not determinable.
191. Foramina for optic nerve and trigeminal nerve (PSAA 191): (0) confluent; (1) widely separate. Not determinable.
192. Lateral head vein (PSAA 192): (0) no distinct foramen for lateral head vein; (1) distinct foramen within the antotic fissure serving the lateral head vein.
193. Anterior extent of cultriform process of parasphenoid (PSAA 193): (0) cultriform process extends to anterior margin of sphenethmoid; (1) cultriform process extends far anterior to sphenethmoid; (2) cultriform process does not reach anterior margin of sphenethmoid. Not determinable.
194. Olfactory bulbs (PSAA 194): (0) narrow; (1) endocasts swollen, leaving considerable impressions in lateral and ventral wall of sphenoid region and in ventral surface of frontal. Not determinable.
195. Flange from skull roof articulating with sphenethmoid (modified from HPSA 223): (0) absent; (1) present on frontal and parietal; (2) present on frontal only.
196. Descending lamina of parietal invades medial orbital wall between ‘pleurosphenoid' and ‘sphenethmoid' elements (PSAA 196): (0) no; (1) yes. Not determinable.
197. Foramen for oculomotor nerve (PSAA 197): (0) exits braincase far dorsal to foramen for optic nerve; (1) exits braincase at or below optic nerve. Not determinable.
198. Intermaxillary fossa (modified from HPSA 198): (0) present; (1) absent. Not determinable.
199. Intermaxillary fossa (PSAA 199): (0) paired; (1) unpaired. Not determinable.
200. Sphenethmoid forms interorbital septum (PSAA 200): (0) no; (1) yes. Not determinable.
201. Anterior extent of cultriform process along palate (PSAA 201): (0) cultriform process extends anteriorly to level of posterior margin of choana; (1) cultriform process dramatically shortened, barely reaching the level of the posterior margin of the orbit. Not determinable.
202. Sutural contact between cultriform process of parasphenoid and vomer (PSAA 202): (0) no; (1) yes. Not determinable.
203. Lateral wall of the nasal capsule underplated by lateral processes of the vomer and palatine (PSAA 203): (0) no; (1) yes. Not determinable.
204. Cultriform process vaulted high above palatal surface (PSAA 204): (0) no; (1) yes. Not determinable.
205. Posterior extent of parasphenoid beneath braincase (PSAA 205): (0) floors sphenoid region only; (1) floors sphenoid and otic region; (2) floors sphenoid, otic, and occipital regions.
206. Basal tubera (PSAA 206): (0) present, with significant endochondral contribution; (1) present, with contribution of parasphenoid only; (2) absent.
207. Path of common internal carotid artery (PSAA 207): (0) does not run alongside braincase, enters braincase directly in region of sella turcica; (1) follows vidian sulcus along posterior surface of basal plate of parasphenoid, enters parasphenoid via vidian canal in basal plate of parasphenoid, divides into cerebral and palatal branches after entering parasphenoid; (2) follows vidian sulcus along posterior surface of basal plate of parasphenoid or lateral wall of braincase, divides into cerebral and palatal branches prior to entering the skull. Not determinable.
208. Buccohypophyseal foramen in parasphenoid (PSAA 208): (0) open; (1) absent.
209. Morphology of pila antotica (PSAA 209): (0) pila antotica is a thin, broad sheet; (1) pila antotica is a robust dorsoventral pillar bracing the skull roof against the palate.
210. Basicranial fissure (PSAA 210): (0) present; (1) absent. Not determinable.
211. Location of vidian sulcus (PSAA 211): (0) along ventral surface of braincase; (1) along lateral surface of braincase. Not determinable.
212. Basipterygoid joint (PSAA 212): (0) epipterygoid comprises entire conus recessus; (1) substantial contribution to conus recessus by pterygoid; (2) conus recessus comprises entirely pterygoid without epipterygoid participation; (3) pterygoid and parasphenoid broadly sutured without development of a conus recessus. Not determinable.
213. Hypophyseal fossa (PSAA 213): (0) single unpaired sulcus; (1) paired sulci divided medially by ridge originating on dorsum sellae.
214. Bone flanking the dorsum sellae (PSAA 214): (0) concurrent with fully ossified lateral braincase; (1) subparallel with sagittal plane (‘pleurosphenoid'); (2) strongly oblique to or perpendicular to sagittal plane (‘laterosphenoid'); (3) restricted to dorsum sellae only. Not determinable.
215. Basal plate of parasphenoid (HPSA 105): (0) roughly quadrangular, basipterygoid articulations narrowly spaced; (1) rectangular laterally, anteroposteriorly narrow, basipterygoid articulations distant. Not determinable.
216. Sphenethmoid (HPSA 114): (0) ossified; (1) unossified.
217. Ossification within the synotic tectum (PSAA 217): (0) synotic tectum massively co-ossified with otic capsules; (1) supraoccipital paired at some point in ontogeny; (2) supraoccipital unpaired throughout ontogeny; (3) no supraoccipital bone, synotic tectum invaded by dorsal processes of exoccipitals.
218. Median ascending process of supraoccipital (PSAA 218): (0) absent; (1) present. Not applicable.
219. Lateral ascending processes of the supraoccipital (PSAA 219): (0) absent; (1) present. Not applicable.
220. Margin of fenestra vestibuli (PSAA 220): (0) parasphenoid excluded by neurocranial elements (basisphenoid and basioccipital); (1) parasphenoid contributes to anteroventral margin of fenestra vestibuli; (2) parasphenoid floors entire fenestra vestibuli; (3) ossification of otic capsule surrounds entire fenestra vestibuli. Not determinable.
221. Crista intervestibularis (PSAA 221): (0) crista intervestibularis absent; (1) crista intervestibularis present. Not determinable.
222. Morphology of crista parotica (PSAA 222): (0) crista parotica meets exoccipitals only, forming lateral wall of posttemporal fossa but not bracing against dermal skull; (1) crista parotica drawn out dorsolaterally into paroccipital process that contacts the tabular; (2) crista parotica drawn out laterally into paroccipital process that contacts the cheek and/or suspensorium. Not determinable.
223. Dorsal process of stapes (HPSA 111): (0) absent; (1) present; Not determinable.
224. Facets on dorsal surface of supraoccipital (PSAA 224): (0) absent; (1) present. Not applicable.
225. Otoccipital fissure (PSAA 225): (0) present; (1) absent. Not determinable.
226. Crista parotica (PSAA 226): (0) descends posteriorly; (1) horizontal along the extent of its length. Not determinable.
227. Position of quadrate with respect to otic capsules (PSAA modified from HPSA 82): (0) quadrates ventral and lateral to otic capsules; (1) quadrates mostly lateral to and greater or equal to twice the width of the otic capsules; (2) quadrates mostly ventral to otic capsules; (3) quadrates approaching or abutting lateral wall of otic capsules.
228. Size of otic capsules (PSAA 228): (0) otic capsules comprise less than 2/3 the width of otoccipital region; (1) otic capsules comprise greater than 2/3 total width of otoccipital region. Not determinable.
229. Otic trough (PSAA 229): (0) absent; (1) present. Not determinable.
230. Articulation between the epipterygoid and prootic (PSAA 230): (0) none; (1) elongate facet on anterior surface of prootic for articulation of epipterygoid. Not determinable.
231. Opisthotic obscures occipital in lateral view (PSAA 231): (0) no; (1) yes. Not determinable.
232. Fenestra vestibularis at end of broad, wing-like lateral extension of the otic capsule (PSAA 232): (0) no; (1) yes. Not determinable.
233. Cristae in otoccipital region (PSAA 233): (0) primarily comprises ascending flanges from braincase; (1) primarily comprises descending flanges from skull roof. Not determinable.
234. Opisthotic excluded from the occipital surface by tabular process of the exoccipital (PSAA 234): (0) no; (1) yes. Not determinable.
235. Insertion of epaxial musculature on occiput (PSAA 235): (0) deep within post-temporal fossae; (1) in broad, shallow fossae along occipital surface of postparietals. Not determinable.
236. Foramen for internal jugular vein (PSAA 236): (0) between supraoccipital and exoccipital; (1) between opisthotic and exoccipital; (2) through exoccipital; (3) posterior notch of fenestra vestibule. Not determinable.
237. Foramina for hypoglossal nerve (PSAA 237: (0) multiple; (1) single; (2) none. Not determinable.
238. Occipital condyle shape (PSAA 238 modified from HPSA 085): (0) round; (1) U-shaped; (2) paired. Not determinable.
239. Ventral process of exoccipital reaches basipterygoid joint along palatal surface (PSAA 239): (0) absent; (1) present. Not determinable.
240. Occipital condyle shape (HPSA 84): (0) concave; (1) convex. Not determinable.
241. Columella of stapes (PSAA modified from HPSA 108): (0) perforate; (1) imperforate.
242. Orientation of stapes (PSAA modified from HPSA 109): (0) dorsal, towards tabular or otic notch; (1) anteroventral, towards quadrate.
243. Stapedial footplate shape (HPSA 110): (0) oval; (1) round; (2) palmate. Not determinable.
244. Dorsal sinus between synotic tectum and skull roof (PSAA modified from HPSA 222): (0) absent; (1) present. Not determinable.
245. Ossification within septum internasale (PSAA 48): (0) absent; (1) present, sphenethmoid; (2) present, mesethmoid. Not applicable.
246. Ossification of septum internasale invades nasal tectum (PSAA 49): (0) absent; (1) present. Not applicable.
247. Insertion of hypaxial musculature (PSAA 50): (0) sphenoid; (1) otic; (2) basioccipital. Not determinable.
248. Prootic supported by pedicel extending from basisphenoid lateral to prootic foramen (PSAA 51): (0) no; (1) yes. Not determinable.
249. Position of pineal foramen with respect to hypophyseal foramen (PSAA 52): (0) anterior to; (1) approximately the same; (2) far posterior to.
250. Pineal foramen or fossa (PSAA 53): (0) well anterior to posterior margin of cerebral fossa; (1) at or behind posterior margin of cerebral fossa. Not determinable.
251. Median wall of otic capsule (PSAA 54): (0) completely unossified; (1) ossification of opisthotic; (2) ossification of supraoccipital and basioccipital. Not determinable.
252. Conical recess in basioccipital (PSAA 55): (0) absent; (1) present. Not determinable.
253. Semicircular canals (PSAA 56): (0) separated from utricular region by bone; (1) not separated from utricular region by bone. Not determinable.
254. Basioccipital (PSAA 57): (0) robust ossification ventral to foramen magnum present; (1) absent.
255. Accessory articulation processes with proatlantal facet on exoccipitals (PSAA 58): (0) absent; (1) prominent; Not determinable.
256. Exoccipital proatlantal facets incorporated into occipital joint (PSAA 59): (0) no; (1) yes. Not determinable.
257. Anterior tectal (modified from CABF 001): (0) anterior tectal present; (1) absent.
258. Ectopterygoid/palatine exposure (CABF 002): (0) more or less confined to tooth row; (1) broad medial exposure additional to tooth row; (2) reduced to thin sliver.
259. Ectopterygoid: (0) present; (1) absent. BL note: no attribution given in PSAA.
260. Ectopterygoid as long or longer than palatines (CABF 003): (0) yes; (1) no.
261. Ectopterygoid reaches subtemporal fossa (CABF 004): (0) no; (1) yes. Not determinable.
262. Frontal (CABF 005): (0) absent; (1) present.
263. Jugal (CABF 007): (0) does not extend anterior to orbit; (1) extends anterior to orbit. Not determinable.
264. Lateral rostral present (CABF 009): (0) yes; (1) no.
265. Maxilla makes interdigitating suture with vomer (CABF 010): (0) no; (1) yes.
266. Maxilla external contact with premaxilla (CABF 011): (0) narrow contact point, not interdigitated; (1) interdigitating suture.
267. Maxilla extends behind level of posterior margin of orbit (CABF 012): (0) yes; (1) no.
273. Pterygoid quadrate ramus margin in subtemporal vacuity (CABF 018): (0) concave; (1) with some convex component. Not determinable.
274. Vomers separated by parasphenoid>half length (CABF 019): (0) yes; (1) no. Not determinable.
275. Vomers (CABF 022): (0) as broad as long or broader; (1) about twice as long as broad or longer.
276. Basipterygoid process (CABF 023): (0) not strongly projecting with concave anterior face; (1) strongly projecting with flat anterior face.
277. Ethmoid (CABF 024): (0) fully ossified; (1) partly or wholly unossified. Not applicable.
278. Hypophyseal region (CABF 025): (0) solid side wall pierced by small foramina for pituitary vein and other vessels; (1) single large foramen. Not determinable.
279. Lateral commissure of otic capsule bearing hyomandibular facets (CABF 026): (0) present; (1) absent. Not determinable.
280. Parasphenoid (CABF 027): (0) does not overlap basioccipital; (1) overlaps basioccipital.
281. Denticulate field of parasphenoid (CABF 028): (0) present; (1) absent.
282. Sphenoid (CABF 029): (0) fully ossified, terminating posteriorly in intracranial joint or fused to otoccipital; (1) separated from otoccipital by unossified gap. Not applicable.
283. Ectopterygoid fang pairs (CABF 030): (0) present; (1) absent.
284. Ectopterygoid row (3+) of smaller teeth (CABF 031): (0) present; (1) absent.
285. Ectopterygoid/palatine shagreen field (CABF 032): (0) absent; (1) present.
286. Palatine row of smaller teeth (CABF 034): (0) present; (1) absent.
287. Pterygoid shagreen (palatine ramus) (PSAA 287 modified from CABF 035): (0) dense; (1) organised radiating rows of denticles; (2) disorganised patches or absent.
288. Dentition of transverse flange of pterygoid (PSAA 288): (0) denticle field indistinct from palatine denticle field; (1) distinct raised denticle field; (2) organised tooth row; (3) absent. Not applicable.
289. Premaxillary tooth proportions (CABF 036): (0) all approximately same size; (1) posteriormost teeth at least twice height of anteriomost teeth; (2) anteriormost teeth largest.
290. Vomerine fang pairs (CABF 037): (0) present; (1) absent.
291. Vomerine fang pairs noticeably smaller than other palatal fang pairs (CABF 038): (0) no; (1) yes.
292. Vomer anterior wall forming posterior margin of palatal fossa bears tooth row meeting at midline (CABF 039): (0) yes; (1) no. Not applicable.
293. Vomerine row of teeth (CABF 040): (0) present; (1) absent.
294. Vomerine shagreen field (CABF 041): (0) absent; (1) present.
295. Adductor fossa (CABF 042): (0) faces dorsally; (1) faces medially.
296. Adductor crest (CABF 043): (0) absent; (1) peak anterior to adductor fossa, dorsal margin of fossa concave; (2) peak above anterior part of adductor fossa, dorsal margin of fossa convex.
297. Angular-prearticular contact (CABF 044): (0) prearticular contacts angular edge to edge; (1) absent; (2) medial lamina of angular sutures with prearticular.
298. Coronoid (anterior) contacts splenial (CABF 045): (0) no; (1) yes.
299. Prearticular extends between infradentaries and middle coronoid (modified from CABF 046): (0) yes; (1) no.
300. Coronoid (middle) contacts postsplenial (CABF 047): (0) no; (1) yes.
301. Coronoid (posterior) posterodorsal process (CABF 048): (0) absent; (1) present.
302. Coronoid (posterior) posterodorsal process visible in lateral view (CABF 049): (0) no; (1) yes. Not applicable.
303. Dentary external to angular and surangular, with chamfered ventral edge and no interdigitations (CABF 050): (0) no; (1) yes.
304. Dentary ventral edge (CABF 051): (0) smooth continuous line; (1) abruptly tapering or ‘stepped' margin.
305. Dentary suture with splenial and postsplenial marked by deep furrow (CABF 052): (0) no; (1) yes.
306. Mandibular sensory canal (CABF 053): (0) present; (1) absent.
307. Mandibular canal exposure (CABF 054): (0) entirely enclosed, opens through lines of pores; (1) mostly enclosed, short sections of open grooves; (2) mostly open grooves, short sections opening through pores; (3) entirely open.
308. Oral sulcus/surangular pit line of mandible (CABF 055): (0) present; (1) absent.
309. Meckelian bone floors precoronoid sulcus (CABF 056): (0) yes; (1) no. Not applicable.
310. Meckelian bone ossified in middle part of jaw (CABF 057): (0) yes; (1) little or no ossification.
311. Meckelian bone exposure in middle part of jaw (CABF 058): (0) depth much less than prearticular; (1) depth similar to prearticular.
312. Meckelian foramina/fenestrae, dorsal margins (CABF 059): (0) Meckelian bone; (1) prearticular; (2) infradentary.
313. Meckelian foramina/fenestrae height (CABF 060): (0) much lower than adjacent prearticular; (1) equal to or greater than depth of adjacent prearticular.
314. Parasymphyseal lateral foramen present (CABF 061): (0) no; (1) yes.
315. Parasymphyseal mesial foramen present (CABF 62): (0) no; (1) yes.
316. Postsplenial with medial lamina (CABF 063): (0) no; (1) small; (2) expanded, contacting precoronoid.
317. Postsplenial pit line present (CABF 064): (0) yes; (1) no.
318. Postsplenial suture with prearticular present (PSAA modified from CABF 065): (0) no; (1) yes.
319. Prearticular sutures with surangular (CABF 066): (0) no; (1) yes.
320. Medial lamina of splenial (PSAA modified from CABF 067): (0) absent; (1) present.
321. Prearticular with longitudinal ridge below coronoids (CABF 068): (0) no; (1) yes.
322. Prearticular with mesially projecting flange on dorsal edge along posterior border of adductor fossa (CABF 069): (0) no; (1) yes.
323. Prearticular centre of radiation of striations (CABF 070): (0) level with posterior end of posterior coronoid; (1) level with middle of adductor fossa; (2) level with posterior end of adductor fossa. Not Applicable.
324. Splenial has free ventral flange (CABF 071): (0) yes; (1) no.
325. Splenial, rearmost extension of medial lamina (CABF 072): (0) closer to anterior end of jaw than to adductor fossa; (1) equidistant; (2) closer to anterior margin of adductor fossa than to the anterior end of the jaw.
326. Coronoids; at least one has fang pair recognisable because at least twice the height of coronoid or marginal teeth (CABF 073): (0) yes; (1) no.
327. Coronoids: at least one has fangs recognisable because noticeable lingual to vertical lamina of bone and to all other teeth (CABF 074): (0) yes; (1) no.
328. Coronoids: at least one has organised tooth row (CABF 075): (0) yes; (1) no.
329. Coronoids: at least one carries shagreen (CABF 076): (0) no; (1) yes.
330. Coronoids: size of teeth (excluding fangs) on anterior and middle coronoids relative to dentary tooth size (CABF 077): (0) about the same; (1) half the height or less.
331. Dentary tooth row (CABF 078): (0) homodont; (1) markedly heterodont.
332. Dentary with parasymphyseal fangs internal to marginal tooth row (CABF 079): (0) yes; (1) no.
333. Dentary teeth (CABF 080): (0) same size as maxillary teeth; (1) larger than maxillary teeth; (2) smaller than maxillary teeth.
334. Dentary with a row of very small teeth or denticles lateral to tooth row (CABF 081): (0) yes; (1) no.
335. Parasymphyseal tooth plate (CABF 082): (0) present; (1) absent.
336. Parasymphyseal plate dentition (CABF 083): (0) shagreen or irregular tooth field; (1) organised dentition aligned parallel to jaw margin; (2) no dentition.
337. Parasymphyseal plate has fang pair (CABF 084): (0) no; (1) yes.
338. Parasymphyseal plate has tooth row (CABF 085): (0) no; (1) short tooth row, separated from coronoid row by diastema; (2) long tooth row reaching coronoid.
339. Prearticular shagreen field distribution (CABF 086): (0) gradually decreasing from dorsal to ventral; (1) well-defined dorsal longitudinal band; (2) scattered patches or absent.
340. Anterior palatal fenestra (CABF 087): (0) single; (1) double; (2) absent
342. Interpterygoid vacuities (CABF 089): (0) absent; (1) at least 2× wider than long; (2) 2× longer than wide.
343. Intracranial joint (CABF 090): (0) present in dermal skull roof; (1) absent in dermal skull roof.
344. Nature of dermal ornament (CABF 091): (0) tuberculate; (1) fairly regular pit and ridge; (2) irregular; (3) absent or almost absent.
345. Nature of ornament: starbursts of radiating ornament on at least some bones (CABF 092): (0) no; (1) yes. Not applicable.
346. Anocleithrum (CABF 093): (0) oblong with distinct anterior overlap area; (1) drop-shaped with no anterior overlap area; (2) absent.
347. Cleithrum (CABF 094): (0) ornamented; (1) not ornamented.
348. Cleithrum, postbranchial lamina (CABF 095): (0) present; (1) absent.
349. Autopod (PSAA modified from CABF 096): (0) radials; (1) digits.
350. Humerus (CABF 097): (0) narrow tapering entepicondyle; (1) square or parallelogram-shaped entepicondyle.
351. Ilium, iliac canal (CABF 098): (0) absent; (1) present.
352. Ilium, posterior process (CABF 099): (0) oriented posterodorsally; (1) oriented approximately horizontally posteriorly.
353. Interclavicle (CABF 100): (0) small and concealed or absent; (1) large and exposed.
354. Interclavicle shape (CABF 101): (0) ovoid; (1) kite-shaped; (2) with posterior stem.
355. Lepidotrichia in paired appendages (CABF 102): (0) present; (1) absent.
356. Posttemporal and supracleithrum (CABF 103): (0) present; (1) absent.
357. Radius and ulna (CABF 104): (0) radius much longer than ulna; (1) approximately equal in length.
358. Ribs, trunk (CABF 107): (0) all cylindrical; (1) some or all bear flanges from posterior margin which narrow distally; (2) some or all flare distally.
359. Scapular blade (CABF 108): (0) absent; (1) small with narrow top; (2) large with broad top.
360. Scapulocoracoid (modified from CABF 109): (0) small and tripodal; (1) large plate. Not applicable.
361. Subscapular fossa (CABF 110): (0) broad and shallow; (1) deeply impressed posteriorly. Not determinable.
362. Squamation (CABF 111): (0) complete body covering of scales, all similar; (1) ventral armour of gastralia. Not applicable.
363. Pectoral process (CABF112): (0) absent; (1) present; Not determinable.
364. Proximal limb of oblique ridge (CABF 113): (0) present, separated from anterior margin of humerus by prepectoral space; (1) absent, replaced by deltopectoral crest. Not determinable.
365. Latissimus dorsi attachment (CABF 114): (0) diffuse ridged area; (1) distinct process.
366. Foramina piercing oblique ventral ridge (CABF 115): (0) many; (1) one moderately large foramen in addition to entepicondylar foramen; (2) entepicondylar foramen is the only large opening, other foramina are pinpricks or absent; (3) none.
367. Humerus with well-developed anterior plate (PSAA 367): (0) yes; (1) no. Not determinable.
368. Subclavicular ossifications (PSAA 368): (0) absent; (1) sometimes present.
369. Radial capitulum (PSAA 369 modified from Sigurdsen & Green Reference Smithson2011): (0) approximately same size as ulnar facet; (1) greatly enlarged and rounded, >2× size of ulnar facet.
370. Lissamphibian inner ear sensu Maddin and Anderson (Reference Maddin and Anderson2012) (MA 220): (0) no; (1) yes. Not determinable.














