
The development of limbs from fins was a defining stage in the evolution of tetrapods (Clack Reference Clack2012). The earliest limbs with digits are thought to have been used to facilitate underwater bottom walking (Coates & Clack Reference Coates and Clack1995), but the critical next step was their use in terrestrial locomotion. Although improved understanding of limb development in both extant and many fossil tetrapods (Shubin & Alberch Reference Shubin and Alberch1986; Wagner & Larsson Reference Wagner, Larsson and Hall2007; Callier et al. Reference Callier, Clack and Ahlberg2009), and a greater knowledge of limb structure in Devonian tetrapods (Lebedev & Coates Reference Lebedev and Coates1995; Coates Reference Coates1996; Jarvik Reference Jarvik1996; Ahlberg Reference Ahlberg2011; Pierce et al. Reference Pierce, Clack and Hutchinson2012) have provided new insight into limb development, the evolution of limbs enabling tetrapods to use quadrupedal gaits on land is still poorly understood (Nyakatura et al. Reference Nyakatura, Andrada, Curth and Fischer2013; Pierce et al. Reference Pierce, Hutchinson and Clack2013).
Here, we name and describe new material from the Early Carboniferous of the Scottish Borders collected by Stanley Wood that reveals an early adaptation for walking. It forms part of a rich assemblage of early tetrapods discovered recently in rocks dating from Romer's Gap (Coates & Clack Reference Coates and Clack1995), an apparent hiatus in the fossil record, which lasted for up to 20 my following the end-Devonian extinction. It was previously characterised by a paucity of tetrapod fossils, which has now been shown by Wood and others to be the result of collection failure (Smithson et al. Reference Smithson, Wood, Marshall and Clack2012; Clack et al. Reference Clack and Kilman2016). The new material includes a humerus structurally intermediate between the plesiomorphic tetrapod condition found in Devonian tetrapodomorph fishes and the tetrapod Acanthostega (Coates Reference Coates1996) and that seen in the humeri of more derived tetrapods from the later Carboniferous, which was carried forward into modern forms. Our new data document one of the earliest modifications of the forelimb in its adaptation for terrestrial walking. Combined with data from fossil tetrapodomorph fishes, later Palaeozoic tetrapods, and extant taxa, we show how increasing the angle of humeral torsion not only transformed the course of the brachial artery and median nerve through the bone, but also potentially improved walking in early tetrapods by contributing to an increase in stride length.
1. Material and methods
The material comprises a right clavicle, humerus and ulna, with a neural arch, centrum and rib, preserved on a single block of matrix, National Museums Scotland (NMS) G. 2012.39.13 (Fig. 1). It was collected from Bed 3 at Willie's Hole, near Chirnside, Scottish Borders, in the Ballagan Formation, dated as Tournaisian, CM palynozone (Smithson et al. Reference Smithson, Wood, Marshall and Clack2012; Clack et al. Reference Clack and Kilman2016). Bed 3 is a laminated micaceous grey siltstone with abundant plant remains, exposed near the base of c.1 m thick sequence containing three distinct fossiliferous horizons (see Ross et al. (this volume), fig. 4 for a detailed stratigraphic log). The associated fauna includes lungfish, actinopterygians, crustaceans and myriapods. An isolated left ilium, NMS G. 2012.39.138, the only other tetrapod element recovered from this horizon, is tentatively referred to this new species.

Figure 1 Mesanerpeton woodi gen. et sp. nov. Type specimen NMS G. 2012.39.13, pectoral girdle, forelimb and axial skeleton bones from Willie's Hole, Scottish Borders, Scotland.
The humerus was removed from the block using a dental mallet and mounted needles. A small portion of the ectepicondyle remains in the matrix, and the proximal posterior edge of the bone and the latissimus dorsi process were damaged slightly during the original collection, but otherwise the bone is intact. The fibula and right side of a neural arch had originally been prepared from the matrix by Mr Wood and then glued back in place. This glue was softened with acetone and the bones removed from the block. The humerus has been compressed slightly, constricting the foramen for the brachial artery and median nerve, but otherwise there is no other evidence of significant post mortem crushing on any of the bones.
Institutional abbreviations. ANSP, Academy of Natural Sciences of Philadelphia; NHMD, Natural History Museum of Denmark; NMS, National Museum Scotland; UMZC, University Museum of Zoology, Cambridge.
2. Systematic palaeontology
Zoobank reference: LSID urn:lsid:zoobank.org:pnb:8BF2CA45–
B378–433A–97A2–4403ICB76503.
Mesanerpeton woodi gen. et sp. nov.
Derivation of name. From μεσαίος (mesaios), meaning ‘intermediary' and ερπετο (erpeto), meaning ‘crawler'; referring to the intermediate condition of the humerus.
woodi sp. nov.
Derivation of the name. In honour of Stan Wood who discovered the tetrapod fauna at Willie's Hole and collected the material.
Type specimen. NMS G. 2012.39.13, right clavicle, humerus and ulna, with a neural arch, centrum and rib, preserved on a single block of matrix.
Type horizon and locality. Laminated micaceous grey siltstone (Bed 3) at Willie's Hole, near Chirnside, Scottish Borders, in the Ballagan Formation, dated as Tournaisian, CM palynozone (Smithson et al. Reference Smithson, Wood, Marshall and Clack2012).
Diagnosis. Neural arch consists of separate bilateral halves lacking clear zygapophyses; humerus with low angle of torsion; a prepectoral space between the proximal head and deltopectoral crest and a foramen for the brachial artery and median nerve which pierces the posterior edge at the base of the entepicondyle; ulna with well-developed olecranon and pronounced lateral keel.
Referred specimen. Left ilium, NMS G. 2012.39.138.
3. Description
3.1. Axial skeleton
The axial skeleton is represented by the two bilateral halves of a neural arch, a centrum and a rib (Fig. 1). The left side of the neural arch has been freed from the matrix (Fig. 2 A–C) but the other elements remain attached to the block.

Figure 2 Mesanerpeton woodi gen. et sp. nov. (A–C) left neural arch NMS G. 2012.39.13: (A) posterior view; (B) anterior view; (C) lateral view. (D–F) reconstruction of vertebra: (D) anterior view; (E) posterior view; (F) lateral view. Scale bar = 10 mm.
3.1.1. Centrum
The centrum is preserved in dorsal view. It is a gently curved strap of unfinished bone, 6 mm long in the midline, tapering to 3 mm long at the lateral ends and 10 mm in diameter. One face is straight, while the other bulges outwards to create the extra length in the middle. The centrum presumably formed a thin bony sheath around the ventral half of the notochord. None of the external surface is visible and the areas of articulation with the neural arch are ill-defined.
3.1.2. Neural arch
The description is based on the left side of the neural arch (Fig. 2). This is half of a bilateral pair which are unfused along the midline. The neural arch most closely resembles those of Crassigyrinus (Panchen Reference Panchen1985; Panchen & Smithson Reference Panchen and Smithson1990). The zygapophyses are poorly developed and, as in Crassigyrinus, it is difficult to determine which are the prezygapophyses and which are the postzygapophyses (see discussion in Panchen & Smithson Reference Panchen and Smithson1990, p.39). Here, the kidney-shaped process, in anterior view, that projects slightly from the edge of the arch is interpreted as the prezygapophysis, and the recess on the inner surface of the arch, which receives it, is interpreted as the postzygapophysis (Fig. 2). This is consistent with Panchen's interpretation of Crassigyrinus (Panchen Reference Panchen1985; Panchen & Smithson Reference Panchen and Smithson1990) where the projecting process was also considered to be the prezygapophysis.
The left half is almost complete, missing only the dorsal part of the neural spine. It is lightly built and lacks the buttressing of the zygapophyses. The transverse process is more clearly defined than those figured from Crassigyrinus and projects laterally from the anteroventral edge of the arch. The articulation for the rib is tall and narrow, extending from a position level with the ventral edge of the postzygapophysis down to the base of the arch. The postzygapophysis is an unfinished area on the internal surface of the arch's posterior edge and is only visible in posterior view. It is bounded laterally by a fine, gently curved ridge, which has the same contour as the lateral edge of the prezygapophysis. This process projects slightly from the arch's anterior edge. The unfinished articulating surface is kidney-shaped, with a convex lateral edge and a concave medial edge.
Inspection of the neural arches of Crassigyrinus described by Panchen (Reference Panchen1985) revealed that undamaged specimens had a similar morphology. The prezygapophysis of neural arch 17 has a convex lateral edge and a concave medial edge. It has an unfinished surface that is raised slightly above the smooth lateral surface of the arch. The postzygapophysis is a gently curved unfinished area on the internal surface of the posterior edge. In neural arch 14 (Panchen Reference Panchen1985, figs 17g, 18b), the prezygapophysis on the left is damaged and has been slightly over-emphasised in the reconstruction. The postzygapophysis is similar to that of neural arch 17. In the vertebra associated with the hind limb material of Crassigyrinus described by Panchen & Smithson (Reference Panchen and Smithson1990, fig. 8d–f)), the morphology of the arch is the same, but the zygapophyses are mislabelled. The posterior view of the neural arch is mislabelled as anterior view and the unfinished area on its internal surface of the arch is mislabelled as prezygapophysis.
3.1.3. Rib
The double-headed rib is 53 mm long. Overall, it is nearly straight, but with a slight dorsal curvature (Fig. 1). The proximal end is flat and the two rib heads are adjacent.
3.2. Appendicular skeleton
The appendicular skeleton is represented by a right clavicle, humerus and ulna, preserved together on the same block with the vertebral elements and rib, and a left ilium. These two specimens are the only tetrapod postcranial remains collected from Bed 3 at Willie's Hole. They are most likely to be from the same animal, but it is possible that the ilium is from a different individual and taxon. However, given that the ilium is of a size expected in an animal with fore limb bones of the size found on NMS G. 2012.35.13, it is described alongside the other material of Mesanerpeton.
3.2.1. Clavicle
The clavicle is preserved in internal (dorsal) view (Fig. 1). It has a flat, triangular-shaped clavicular blade and thickened clavicular stem. The blade is approximately 50 mm long with a maximum width of 25 mm and has similar proportions to the clavicles attributed to Doragnathus (Smithson & Clack Reference Smithson and Clack2013). Although the blade lacks the dorsal ridge seen in Doragnathus, which was thought to have strengthened the union with the interclavicle, it does bear a number of ridges and grooves along the long axis of the posterior half of the blade, which probably served the same purpose. The clavicular stem projects dorsally at an angle of about 40 degrees. On its posterior edge is a thin curved lamina of bone that probably received the ventral half of the cleithrum.
3.2.2. Humerus
The humerus of Mesanerpeton (Fig. 3) has the characteristic L-shape of early tetrapods, with a broad proximal articulation and large rectangular entepicondyle. It is 55 mm long. The bone is twisted midway along its shaft and the angle of torsion between the proximal and distal articulations is c.25 degrees. The insertions of the principal locomotory muscles from the pectoral girdle to the proximal end of the humerus are well developed.

Figure 3 Mesanerpeton woodi gen. et sp. nov. (A–D) right humerus, NMS G. 2012.39.13: (A) posterior view; (B) dorsal view, plane of the proximal dorsal surface; (C) ventral view, plane of the proximal dorsal surface; (D) anterior view. (E–J) reconstruction of right humerus: (E) posterior view; (F) dorsal view, plane of the proximal dorsal surface; (G) ventral view, plane of the proximal dorsal surface; (H) anterior view; (I) proximal view; (J) distal view. Abbreviations: br for = brachial foramen; d = deltoid; ect = ectepicondyle; ent = entepicondyle; lat d = latissimus dorsi; p = pectoralis; pre = prepectoral space; rad = radial condyle; s = supinator; ul = ulnar condyle. Scale bars = 10 mm.
The proximal posterior edge is slightly concave. Despite the slight damage in this area, there is no evidence of the pre-entepicondylar ridge found in Acanthostega. The foramen for the brachial artery and median nerve pierces the edge of the bone at the junction where the proximal posterior edge of the humerus meets the medial edge of the entepicondyle. It is clearly visible in posterior view but, unlike in the humeri of other Carboniferous tetrapods, where the foramen pierces the body of the entepicondyle, the entrance is not visible in dorsal view. The exit is conspicuous in the centre of the ventral surface of the entepicondyle. In the area immediately distal to the ventral edge of the humeral head is a shallow depression marking the insertion of the coracobrachialis muscle.
On the dorsal surface is a prominent ectepicondyle. This starts as a low swelling in front of the entrance of the entepicondylar foramen and extends anterodistally as a dorsally curved, gently rounded ridge. It terminates immediately before the distal edge of the humerus, between the radial and ulna condyles. In the centre of the humerus, midway between the proximal articulation and the start of the ectepicondyle, is a low ridge which formed the insertion of the latissimus dorsi. It may have terminated proximally in a distinct process, but the bone is slightly damaged in this area and the height of the ridge is uncertain.
The well preserved anterior edge on the humerus shows the insertions of the deltoideus and pectoralis muscles very clearly. They form a swelling on the proximal third of the anterior edge, the deltopectoral crest. Each insertion is a slightly excavated area of unfinished bone almost triangular in outline. The pectoralis insertion lies proximal to that for the deltoideus and is visible in both anterior and ventral views. In contrast, the deltoideus insertion is only visible in anterior view.
Lying between the pectoral insertion and the proximal head of the humerus is a deep recess, the prepectoral space. Its ventral margin forms the ventral edge of the humerus, but its dorsal edge curves towards the latissimus dorsi process so that the space is visible in both anterior and dorsal views. In tetrapods, the prepectoral space has only previously been described in Ichthyostega (Jarvik Reference Jarvik1996; Callier et al. Reference Callier, Clack and Ahlberg2009). In tetrapodomorph fishes, this space is a large area on the proximal part of the anterior surface of the humerus, between the dorsal and ventral edges. It appears to have been lost as the dorsal and ventral edges came togeher, uniting the widely separated deltoid process on the dorsal edge and the pectoral process on the ventral edge to form the tetrapod apomorphy of the deltopectoral crest.
On the ventral surface, extending from the pectoralis insertion to the anteroproximal edge of the entepicondyle, is a low, poorly differentiated ventral ridge. The distal slope of the pectoralis articulation is scarred by shallow grooves, which mark the origin of the brachialis muscle. An oval-shaped swelling on the anterior edge of the humerus, distal to the deltoid articulation, marks the origin of the supinator muscle. This area of unfinished bone lies proximal to the exit of the entepicondylar foramen, as it does in Eoherpeton (Smithson Reference Smithson1985); in contrast to Acanthostega (see below) where the supinator process lies distal to the foramen. The anterior edge terminates with the radial condyle. It is an oval-shaped, unfinished area that is raised in the centre and is partially visible in ventral view. The ulnar condyle is an oval-shaped recess with a pronounced dorsal lip, lying partially below the distal end of the ectepicondyle, and is only visible in distal view.
3.2.3. Ulna
The ulna of Mesanerpeton (Fig. 4) is a short rod 30 mm long, expanded proximally to form a relatively large area of articulation with the humerus, and tapered distally to form a small area of articulation with the carpus. When compared with the length of the humerus, it is relatively longer than the ulnae of the Devonian taxa Acanthostega (Coates Reference Coates1996), Ichthyostega (Jarvik Reference Jarvik1996) and Tulerpeton (Lebedev & Coates Reference Lebedev and Coates1995) and, instead, is similar to the ulna of Archeria (Romer Reference Romer1957).
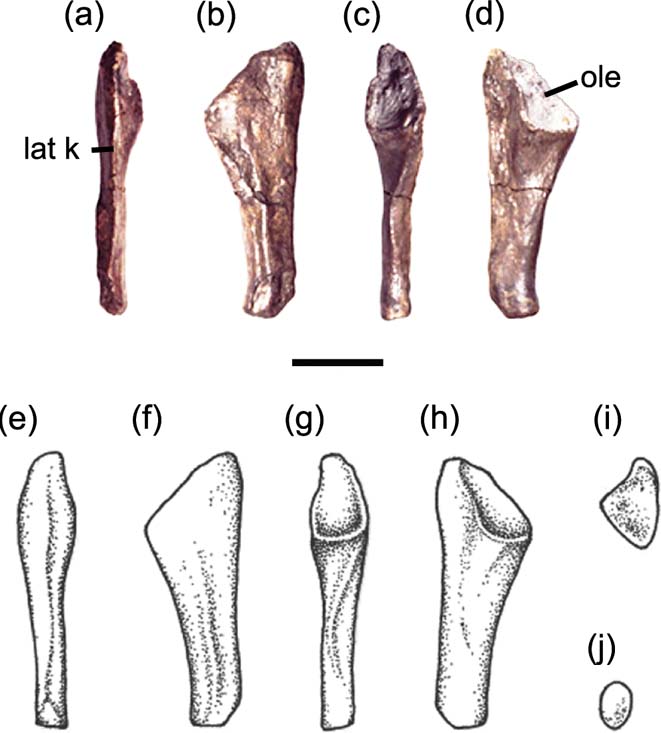
Figure 4 Mesanerpeton woodi gen. et sp. nov. (A–D) right ulna, NMS G. 2012.39.13: (A) lateral view; (B) dorsal (flexor) view; (C) medial view; (D) ventral (extensor) view. (E–J) restoration of right ulna: (E) lateral view; (F) dorsal (flexor) view; (G) medial view; H ventral (extensor) view; (I) proximal view; (J) distal view. Abbreviations: lat k = lateral keel; ole = olecranon. Scale bar = 10 mm.
Unlike in many early tetrapods, the olecranon is well ossified and makes up approximately one-third of the length of the ulna. It fully surrounds the articulation with the humerus. It is tear drop-shaped in medial view (Fig. 4c), with a narrow proximal rim and wide distal lip. Its dorsal (flexor) surface is embayed laterally to expose the articulating surface, but it is concealed ventrally (cf Fig. 4b, d). The ulnae of Archeria (Romer Reference Romer1957, fig. 5) and Proterogyrinus (Holmes Reference Holmes and Panchen1980, fig. 7) are similar in this regard, in contrast to Eryops (Pawley & Warren Reference Pawley and Warren2006, fig. 7) where the area of articulation is essentially only visible in medial view. In amniotes, both the dorsal and ventral surfaces are embayed to produce the sigmoid notch that articulates with the trochlea on the humerus (Romer Reference Romer1956).
The lateral edge of the ulna is essentially straight and bears a strong ridge (the lateral keel) along its length, which separated the extensor and flexor musculature. The medial edge below the distal lip of the humeral articulation is also mainly straight before curving medially immediately above the distal articulating surface. It also bears a sinuous ridge along much of its length.
The distal end is very narrow. It is approximately half as wide as the proximal end in both lateromedial and dorsovental views. It bears two articulating surfaces of similar size, set at an angle of 120 degrees to each other, for the intermedium and ulnare.
3.2.4. Ilium
The left ilium attributed to Mesanerpeton (Fig. 5) is a robust, well ossified, biramus bone, with an iliac blade and post iliac process dorsally and a long suture with the puboischiadic plate ventrally. In between, the waist is relatively thick.

Figure 5 Mesanerpeton woodi gen. et sp. nov. (A–B) left ilium, NMS G. 2012.39.138: (A) lateral view; (B) medial view. (C–E) restoration of left ilium: (C) lateral view; (D) medial view; (E) dorsal view. Abbreviations: supra b = supra-acetabula buttress; supra n = supra-acetabular notch. Scale bar = 10 mm.
The iliac blade and post iliac process are slightly off set (Fig. 5C). The iliac blade occupies a more medial position and the dorsal edge of the post iliac process forms a continuation of the anterior edge of the ilium, as it extends upwards and backwards above the acetabulum. When the ventral edge of the ilium is orientated horizontally, as in Figure 5, the post iliac process projects gently posterodorsally and the iliac blade extends slightly above it. The post iliac process tapers to a thin blade posteriorly, with a sharp edge ventrally. Its posterior tip is unfinished and it probably continued in cartilage in life. The dorsal portion of the medial surface of the iliac blade is roughly textured probably marking its connection with the sacral rib. A faint transverse line extends across the lateral surface of the ilium, between the top of the acetabulum and the dorsal edge of the post iliac process. It is less prominent than in Acanthostega (Coates Reference Coates1996) and Eoherpeton (Smithson Reference Smithson1985).
The ventral portion of the ilium is well ossified and formed the dorsal half of the acetabulum. The anterior tip of the ventral edge is broken and a small area is missing. We estimate the suture between the ilium and the pubo-ischiadic plate was 34 mm long. The acetabulum occupies approximately 80 % of its length. There is a well-developed supra-acetabular buttress in the anterior half of the acetabulum and a relatively deep supra-acetabular notch above the posterior half. As in Acanthostega (Coates Reference Coates1996, p. 389), the buttress lies far forward, in front of the iliac blade, and thus in front of the sacrum. On the mesial surface below the waist, a faint ridge extends ventrally down the centre the ilium, dividing it into anterior and posterior halves. A faint ridge occupies a similar position on the Blue Beach pelvis described by Anderson et al. (Reference Anderson, Smithson, Mansky, Meyer and Clack2015, fig. 13A). It is more prominent in Archeria (Romer Reference Romer1957, fig. 3), where it continues on to the pubis.
A small portion of the pubis is preserved in NMS G. 2012.39.138 below the ilium (Fig. 5). The anterior edge was broken during collection, but the other incomplete edges appear to have been eroded prior to preservation. The ventral part of the acetabulum is preserved in lateral view and the pubo–iliac suture is marked on the medial surface by a slight depression along its length. This surface is rough and appears to lack the complete periosteal covering found on the ilium.
4. Discussion
Our description of the humerus of Mesanerpeton prompted a re-examination and reinterpretation of the humerus of Acanthostega, based on Natural History Museum of Denmark specimen NHMD 29020 (Fig. 6) and further preparation of the left humerus, specimen NHMD 29019. In Coates (Reference Coates1996), the entry point for the brachial foramen was shown in both dorsal and ventral views of the humerus (Coates Reference Coates1996, fig. 16C, D). However, examination of NHMD 29020 shows that it lies on the ventral surface, close to, but not on, the edge of the entepicondyle. It is not visible in dorsal view, but penetrates the ventral aspect of the thickened edge of the entepicondyle, described by Callier et al. (Reference Callier, Clack and Ahlberg2009) as part of the distal portion of the ventral ridge. The posterior edge of the humeral head is essentially straight, here labelled the pre-entepicondylar ridge, and lacks the sigmoid curvature of the humeri of most stem tetrapods which is also seen in Mesanerpeton (cf Figs 5, 6). The indentation at the inception of the entepicondyle shown by Coates (Reference Coates1996, fig. 16C, D), results from damage to NHMD 29019 in that area. It is not present in NHMD 29020.
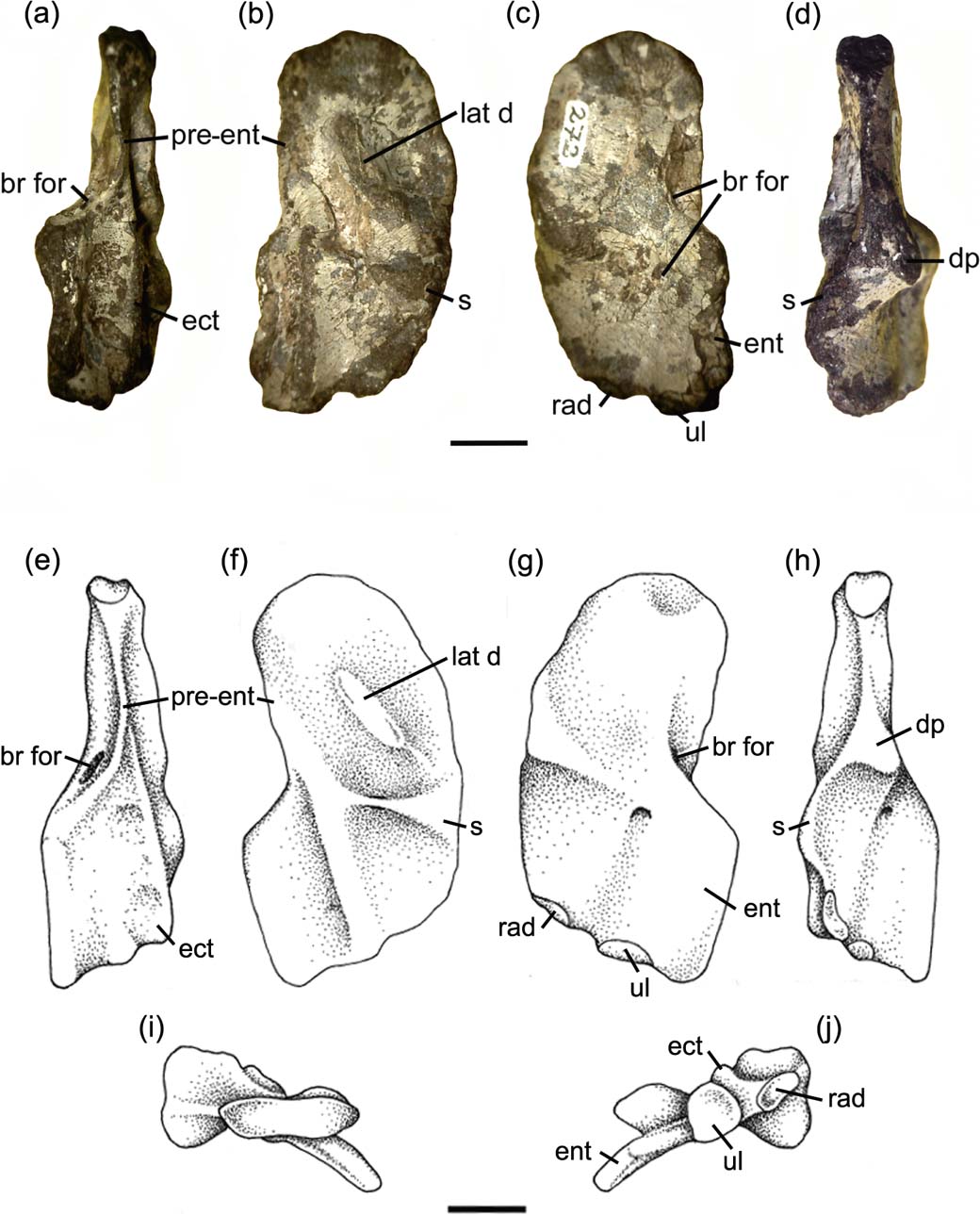
Figure 6 Acanthostega gunnari. (A–D) right humerus, NHMD 29020: (A) posterior view; (B) dorsal view, plane of the proximal dorsal surface; (C) ventral view, plane of the proximal dorsal surface; (D) anterior view. (E–J) reconstruction of the right humerus: (E) posterior view; (F) dorsal view, plane of the proximal dorsal surface; (G) ventral view; plane of the proximal dorsal surface; (H) anterior view; (I) proximal view; (J) distal view. Abbreviations: br for = brachial foramen; d = deltoid; dp = deltopectoral crest; ect = ectepicondyle; ent = entepicondyle; lat d = latissimus dorsi; p = pectoralis; pre = prepectoral space; pre-ent = pre-entepicondylar ridge; rad = radial condyle; s = supinator; ul = ulnar condyle. Scale bars = 10 mm.
4.1. Humerus evolution
New information on the morphology of the Acanthostega humerus and the discovery of Mesanerpeton allows us to reassess the early evolution of the tetrapod humerus and forelimb.
In Devonian tetrapodomorph fishes such as Tiktaalik (Shubin et al. Reference Shubin, Daeschler and Jenkins2006) and Gogonasus (Holland Reference Holland2013), the course of the brachial artery and median nerve was entirely ventral and they passed through the ventral ridge. The vessels follow the same course in Acanthostega, and this probably represents the pleisiomorphic tetrapod condition. Most living amniotes and most stem tetrapods show the condition in which the brachial artery and median nerve passed from the dorsal (or anterior) to the ventral (or posterior) surface of the entepicondyle (e.g., Gregory Reference Gregory1949; Landry Reference Landry1958; Smithson Reference Smithson1985; Godfrey Reference Godfrey1989; Lombard & Bolt Reference Lombard and Bolt1995; Clack & Finney Reference Clack and Finney2005; Saber Reference Saber2013; Bishop Reference Bishop2014). Mesanerpeton represents an intermediate condition in which the vessels passed into the humerus through the posterior edge of the entepicondyle, between the ventral and dorsal faces of the bone. The same intermediate condition is present in the humerus of Ossirarus (Clack et al. Reference Clack, Bennett, Carpenter, Davies, Fraser, Kearsey, Marshall, Millward, Otoo, Reeves, Ross, Ruta, Smithson, Smithson and Walsh2016). This is a small, unrelated tetrapod with gastrocentrous vertebrae and well-ossified neural arches. It was collected from a horizon in the Ballagan Formation at Burnmouth, which is slightly younger than the beds exposed at Willie's Hole (Clack et al. Reference Clack, Bennett, Carpenter, Davies, Fraser, Kearsey, Marshall, Millward, Otoo, Reeves, Ross, Ruta, Smithson, Smithson and Walsh2016), and will be described by the present authors in due course.
This list of tetrapodomorph fishes, stem tetrapods and amniotes should not be regarded as a temporal or phylogenetic sequence, as some Devonian tetrapods also show the typical tetrapod condition with a dorsal point of entry. These include material attributed by Callier et al. (Reference Callier, Clack and Ahlberg2009) to Acanthostega (UMZC T.1295) and the scanned and rendered material of Ichthyostega (NHMD 29017a) (Callier et al. Reference Callier, Clack and Ahlberg2009). Tulerpeton (Lebedev & Coates Reference Lebedev and Coates1995) may also have the typical tetrapod condition, although the humerus is damaged in the critical area. These observations do not help resolve the identity of the Red Hill humerus, ANSP 21350 (Shubin et al. Reference Shubin, Daeschler and Coates2004; Ruta & Wills Reference Ruta and Wills2016). This humerus, thought to be that of a tetrapod (Shubin et al. Reference Shubin, Daeschler and Coates2004; Ahlberg Reference Ahlberg2011), shows little torsion and is pierced by a number of different foramina, but the course of the brachial artery and median nerve through the bone is uncertain.
The change of course of the brachial artery and median nerve through the humerus is brought about by humeral torsion. During evolution, twisting of the distal half of the humerus relative to the proximal half appears to have brought the bone across the path of the vessels and changed their positional relationship. Primitively, in a humerus with little or no torsion, the vessels ran under the bone and passed through the ventral ridge. Humeral torsion depressed the entepicondyle, initially causing the vessels to pass through the edge of the entepicondyle and eventually through the condyle itself (Fig. 7).

Figure 7 Humeri of tetrapodomorphs in dorsal (left) and ventral (right) views showing the course of the brachial artery and median nerve (in red): (A) Gogonasus (after Holland Reference Holland2013); (B) Tiktaalik (after Shubin et al. Reference Shubin, Daeschler and Jenkins2006); (C) Acanthostega; (D) Mesanerpeton; (E) Eoherpeton (after Smithson Reference Smithson1985); (F) Captorhinus (after Holmes Reference Holmes1977); (G) Dimetrodon (after Romer & Price Reference Romer and Price1940); (H) Ornithorhynchus, based on UMZC A2.2/2. Scale bars = 10 mm.
The angle between the long axis of the head of the humerus and the proximal edge of the entepicondyle increases along a morphocline, starting with Acanthostega, and reaches its greatest angle in large Permian stem amniotes such as Diadectes and Limnoscelis (Gregory Reference Gregory1949) and early amniotes such as Captorhinus (Holmes Reference Holmes1977) and some sphenacodonts (Romer & Price Reference Romer and Price1940). Twisted humeri which retain a brachial foramen are found in many extant amniotes with a sprawling gait. These include the reptiles Sphenodon (Smithson pers. obs. UMZC R2586) and Varanus (Smithson pers. obs. UMZC R9410) and the monotremes Ornithorynchus (Smithson pers. obs. UMZC A2-2/2) and Tachyglossus (Smithson pers. obs. UMZC A1.3\1).
Increasing humeral torsion appears to be one way in which early tetrapods improved walking. Plesiomorphically, the elbow joint had limited excursion, so that elbow flexion and extension was minimal (Jenkins Reference Jenkins1973; Hopson Reference Hopson, Dial, Shubin and Brainerd2015). As a consequence, the whole forelimb moved as a unit at the glenoid and locomotion was achieved by a combination of movements of the humerus (Romer Reference Romer1922). Increasing humeral torsion has its largest impact on walking at the beginning of the power stroke, when the head of the humerus is horizontal. With a greater angle of twist, the epipodials can achieve a greater elevation and step further in front of the anterior edge of the glenoid. This leads to an increase in stride length, the distance travelled between the end of one power stroke, when the epipodials are essentially vertical (Jenkins Reference Jenkins1973), and the start of the next. If all other components of the locomotory cycle remain constant, increasing the angle of torsion increases stride length by the simple relationship:
 $$s = e \sin a$$
$$s = e \sin a$$
where s is increase in stride length, e is epipodial length and sin a is sine angle of torsion (Fig. 8A). At low angles of torsion, the effect is relatively small, but an angle of 45 degrees increases stride length by more than 70 % (Fig. 8B). Further twisting increases stride length even more. The increase in angle is usually accompanied by an increase in epipodial and digit length, the earliest examples of which are observed in the Viséan stem amniotes Casineria (Paton et al. Reference Paton, Smithson and Clack1999) and Westlothiana (Smithson et al. Reference Smithson, Carroll, Panchen and Andrews1994), with angles of torsion of c.60 and 70 degrees respectively (Smithson pers. obs.).

Figure 8 Changes in manus placement with increasing angles of humeral torsion: (A) the geometry of humeral torsion and its relation to manus placement; (B) representation of changes in manus placement with increasing angle of humeral torsion, lateral view. The taxa represented here have been studied by the authors. The angles of torsion of other early tetrapods are listed in Appendix 1 (see section 7).
Humeral torsion in early tetrapods has been discussed by numerous authors. It was noted by Watson (Reference Watson1917), Romer (Reference Romer1922, Reference Romer1957; Romer & Price Reference Romer and Price1941) and Miner (Reference Miner1925), and more recently by Holmes (Reference Holmes and Panchen1980), Smithson (Reference Smithson1985) and Coates (Reference Coates1996), but was considered in most detail first by Evans & Krahl (Reference Evans and Krahl1945) and later by Andrews & Westoll (Reference Andrews and Westoll1970). In these detailed studies, a number of axes were identified along the proximal and distal ends of the humerus and the angle between them measured. The angle we have measured above is the same as Andrews & Westoll's axis of the ‘flattened extremities' (Andrews & Westoll Reference Andrews and Westoll1970, p. 249). However, in none of these studies was the changing position of the brachial foramen discussed, or the possible impact of humeral torsion on stride length considered.
Humeri with a typical entepicondylar (brachial) foramen and angle of twist greater than 45 degrees have recently been described from the Blue Beach Formation at Blue Beach in Nova Scotia (Anderson et al. Reference Anderson, Smithson, Mansky, Meyer and Clack2015). These strata are coeval with the Ballagan Formation of the Scottish Borders, in which we have also found the earliest five-digited autopod (Smithson et al. Reference Smithson, Wood, Marshall and Clack2012). This suggests that tetrapods in the early Carboniferous were experimenting with limb morphology and developing new ways of walking. Compelling evidence of this comes from the large number of trackways that have been found at Blue Beach, where at least five different trackway morphotypes have been identified, in varying sizes and demonstrating different gaits (Mansky & Lucas Reference Mansky and Lucas2013; Clack Reference Clack and Kilman2016).
5. Conclusions
During the evolution of tetrapods, the course of the brachial artery and median nerve through the humerus changes. This change is due to humeral torsion, the twisting of the distal end of the bone relative to its proximal end. Torsion transformed the course of the vessels through the bone, from an entirely ventral path, found in Devonian tetrapodomorph fishes and the tetrapod Acanthostega, to one in which the vessels pass through the entepicondyle from the dorsal to the ventral surface. This arrangement is seen in most fossil and extant tetrapods which retain the entepicondylar (brachial) foramen. The humerus of Mesanerpeton from the early Carboniferous of Scotland shows an intermediary condition. Humeral torsion is suggested to improve walking in early tetrapods by contributing to an increase in stride length.
6. Acknowledgements
The new material described here was collected by the late Stan Wood. His discoveries of early Carboniferous vertebrates in Scotland made an unprecedented contribution to our understanding of the structure, relationships and evolution of early tetrapods. We thank Stig Walsh and Andy Ross, National Museums Scotland, for the loan of the material and permission to prepare it; the Natural History Museum of Denmark, for the loan of the Acanthostega specimens; Sandra Chapman, Natural History Museum, London, for access to Crassigyrinus; Daniel Brinkman, Yale Peabody Museum, Anthony Howell, Redpath Museum and Deborah Skilliter, Nova Scotia Museum, for access to specimens from Blue Beach; Chris Mansky and Sonia Wood, Blue Beach Fossil Museum, for showing us the Blue Beach trackways; and Matthew Lowe, University Museum of Zoology, Cambridge, for access to skeletons of extant amniotes. We thank Per Ahlberg, Mike Coates and Stephanie Pierce for discussion following our preparation and reinterpretation of the humerus of Acanthostega, and Nick Fraser for discussion on the relationship between humeral torsion and stride length. We are grateful to our reviewers, John Hutchinson and Marcello Ruta, for their helpful comments and suggestions that greatly improved the manuscript. This is a contribution to the TW:eed Project, Tetrapod World: early evolution and diversity, and we thank members of the project team for discussion and support. This work was carried out with the aid of NERC research grant NE/J022713/1.
7. Appendix 1. Angle of torsion and position of entrance of the brachial foramen in early tetrapods









