Phytosauria is recognised as a monophyletic group of archosauriforms (Sereno Reference Sereno1991; Brusatte et al. Reference Brusatte, Benton, Desojo and Langer2010; Nesbitt Reference Nesbitt2011), and most recently the clade was hypothesised to be the sister taxon to Archosauria (Nesbitt Reference Nesbitt2011). Recent phylogenetic analyses recovered increasingly resolved relationships within Phytosauria (Hungerbühler Reference Hungerbühler2002; Parker & Irmis Reference Parker, Irmis, Parker, Ash and Irmis2006; Stocker Reference Stocker2010, Reference Stocker2012). However, those phylogenies were focused on resolving relationships among derived phytosaurid phytosaurs; relationships among non-phytosaurid phytosaurs and their taxonomy remain contentious and unresolved.
Paleorhinus was named based on a fragmentary specimen (FMNH UC 632; holotype specimen of Paleorhinus bransoni Williston, Reference Williston1904; Lees Reference Lees1907) from the Popo Agie Formation of Wyoming. Other species of Paleorhinus were since named based on additional specimens from the Popo Agie Formation (Paleorhinus parvus Mehl, Reference Mehl1928), from the Dockum Group of Texas (Paleorhinus scurriensis Langston, Reference Langston1949; Paleorhinus sawini Long & Murry, Reference Long and Murry1995), and from the Late Triassic of Morocco (Paleorhinus magnoculus Dutuit, Reference Dutuit1977). Additionally, the name Paleorhinus was applied to other specimens of non-phytosaurid phytosaurs from the Late Triassic of India (=Parasuchus hislopi), Poland (multiple specimens from the Krasiejów clay pit; Dzik Reference Dzik2001; Dzik & Sulej Reference Dzik and Sulej2007), Austria (=‘Francosuchus’ trauthi), Germany (=Mesorhinosuchus fraasi, Paleorhinus angustifrons (sensu Butler et al., Reference Butler, Stocker, Rauhut, Lautenschlager and Bronowicz2012), Francosuchus latus, Ebrachosuchus neukami, Francosuchus broilii) and the United States, specifically Arizona (MNA V3728 from the Downs Quarry) and Texas (Promystriosuchus ehlersi, specimens from the Otis Chalk localities). Most of those taxa were synonymised within Paleorhinus/Parasuchus (Gregory Reference Gregory1962; Ballew Reference Ballew, Lucas and Hunt1989; Hunt & Lucas Reference Hunt and Lucas1991; Long & Murry Reference Long and Murry1995), based on shared plesiomorphic morphological characters and low stratigraphic positions within their respective lithologic units.
Several researchers proposed the possibility that ‘Paleorhinus’ is a paraphyletic group of non-phytosaurid phytosaurs (Hungerbühler Reference Hungerbühler1998, Reference Hungerbühler2000; Wroblewski Reference Wroblewski2003; Irmis Reference Irmis2005; Parker & Irmis Reference Parker, Irmis, Parker, Ash and Irmis2006). The hypothesis of a paraphyletic Paleorhinus was supported by a recently published cladistic analysis (Stocker Reference Stocker2010). In that study, ‘Paleorhinus’ scurriensis was recovered as the basal-most phytosaur taxon included in the analysis (Fig. 1). Additionally, characters that had been considered diagnostic of Paleorhinus (e.g. Ballew Reference Ballew, Lucas and Hunt1989) were reinterpreted as plesiomorphic for Phytosauria (Stocker Reference Stocker2010).

Figure 1 Strict consensus based on Stocker (Reference Stocker2010) showing position of Wannia scurriensis gen. nov. and paraphyly of ‘Paleorhinus’. Bootstrap values are listed to the lower left of the node, and Bremer support values are listed to the upper left of the node.
A challenge to understanding the phylogenetic relationships and taxonomy of non-phytosaurid phytosaurs is the limited inclusion of those species in published phylogenetic analyses (Ballew Reference Ballew, Lucas and Hunt1989; Hungerbühler Reference Hungerbühler2002; Parker & Irmis Reference Parker, Irmis, Parker, Ash and Irmis2006; Stocker Reference Stocker2010, Reference Stocker2012). Most phylogenetic analyses that examined phytosaur in-group relationships were focused on more derived leptosuchomorph phytosaurs, and included a limited sample of non-phytosaurid phytosaurs or a single non-phytosaurid phytosaur as an out-group taxon only. Those analyses were not developed to test which characters – if any – are synapomorphies that unite various species in a monophyletic Paleorhinus. Additionally, the majority of specimens referred to ‘Palerohinus’ require a modern comparative description of their anatomy in order to better understand morphological evolution at the base of Phytosauria.
The type and only known specimen of ‘Paleorhinus’ scurriensis (TTU P-00539) is a partial skull collected from the base of the Dockum Group in Scurry County, Texas, (Langston Reference Langston1949) and is an important specimen with respect to the application of the Otischalkian land vertebrate faunachron (lvf) (Hunt & Lucas Reference Hunt and Lucas1991). TTU P-00539 was first described over sixty years ago (Langston Reference Langston1949). Using an explicitly cladistic framework, I re-examined the holotype of ‘Paleorhinus’ scurriensis and provide a full redescription of that specimen in a comparative context. I focus on where my interpretation differs from the original description, or where the new morphology is visible because of further preparation, and I provide comparisons with other North American specimens previously referred to ‘Paleorhinus,’ as well as to a number of other taxa of phytosaurs.
Institutional abbreviations. AMNH FARB, American Museum of Natural History, Fossil Amphibian, Reptile, and Bird collections, New York; BSPG, Bayerische Staatssammlung für Paläontologie und Geologie, Munich; FMNH UC, Field Museum of Natural History, University of Chicago collections, Chicago; MNA, Museum of Northern Arizona, Flagstaff; MNHN, Muséum National d'Histoire Naturelle, Paris; MU, University of Missouri, Columbia; NHMW, Naturhistorisches Museum Wien, Vienna; PEFO, Petrified Forest National Park, Arizona; TMM, Vertebrate Paleontology Laboratory, Texas Natural Science Center, Austin; TTU P, Texas Tech University, Lubbock; UCMP, University of California Museum of Paleontology, Berkeley; UMMP, University of Michigan Museum of Paleontology, Ann Arbor; ZPAL, Institute of Palaeobiology, Polish Academy of Sciences, Warsaw, Poland.
1. Geographic and geologic setting
TTU P-00539 was collected by Dr. John Clark in 1937, from a “white, fine-grained, calcareous sandstone” on the H. G. Bryan farm, 2·5 miles north-east of the town of Camp Springs, Scurry County, Texas (Langston Reference Langston1949, p. 324; B. Mueller, pers. com. 2011; Fig. 2) within what is now known as the Camp Springs Formation of the Dockum Group. That unit originally was called the Camp Springs Conglomerate (Beede & Christner Reference Beede and Christner1926) and referred to the lower part of the Dockum Group in this area of Texas. The precise stratigraphic location of the locality is unclear because of the complex sedimentology and stratigraphy of the basal conglomeratic units of the Dockum Group (e.g. Lehman Reference Lehman1994; Lucas et al. Reference Lucas, Anderson, Hunt, Ahlen, Peterson and Bowsher1994; Riggs et al. Reference Riggs, Lehman, Gehrels and Dickinson1996; Martz Reference Martz2008). The Camp Springs Member was correlated tentatively with the Santa Rosa Sandstone, the basalmost unit of the Dockum Group in both Texas and New Mexico (Martz Reference Martz2008). However, the Santa Rosa Sandstone potentially comprises two units, with the Camp Springs Formation in Texas only correlative to the lower, Tecololito Member of the Santa Rosa Formation in New Mexico (Lehman Reference Lehman1994; Lucas et al. Reference Lucas, Anderson, Hunt, Ahlen, Peterson and Bowsher1994; Martz Reference Martz2008).

Figure 2 Map of Texas, USA, showing surface exposure of the Upper Triassic Dockum Group. Arrow indicates the general location of the town of Camp Springs within those exposures. Larger map indicating the approximate location of the type locality (within the dashed box) of Wannia scurriensis gen. nov., with respect to the cities of Snyder, Sweetwater, Colorado City and Big Spring, based on locality information reported by Langston (Reference Langston1949).
The age of the base of the Dockum Group is not well constrained. The Camp Springs Formation was correlated with the Colorado City Formation (=the Iatan Member of Lucas & Hunt [Reference Lucas, Hunt, Lucas and Morales1993], encompassing the Otis Chalk localities) within the Otischalkian lvf because of the shared presence of Paleorhinus (Hunt & Lucas Reference Hunt and Lucas1991; Lucas & Hunt Reference Lucas, Hunt, Lucas and Morales1993). The age of the Otischalkian was hypothesised based on correlations with the Opponitzer Limestone (late Carnian) of Austria, using the presence of NHMW 1905/0007/0052, a fragmentary rostrum that was identified as Paleorhinus trauthi (e.g. Huene Reference Huene1939; Trauth Reference Trauth1948; Hunt & Lucas Reference Hunt and Lucas1991). However, re-evaluation of Paleorhinus trauthi revealed that it differs from other non-phytosaurid phytosaurs in multiple features, and instead that specimen more likely represents a non-phytosaurian saurian (Butler et al. Reference Butler, Stocker, Rauhut, Lautenschlager and Bronowicz2012). Based on radioisotopic and magnetostratigraphic revisions to the Late Triassic timescale (Furin et al. Reference Furin, Preto, Rigo, Roghi, Gianolla, Crowley and Bowring2006; Muttoni et al. Reference Muttoni, Kent, Olsen, Di Stefano, Lowrie, Bernasconi and Hernández2004), the Carnian–Norian boundary was re-dated at approximately 228 Ma. That change to the placement of the Carnian–Norian boundary, along with recent radioisotopic dates for portions of the Chinle Formation (e.g. Irmis et al. Reference Irmis, Mundil, Martz and Parker2011; Ramezani et al. Reference Ramezani, Hoke, Fastovsky, Bowring, Therrien, Dworkin, Atchley and Nordt2011), indicate that exposures of Chinle Formation in Arizona and New Mexico are entirely post-Carnian (Irmis et al. Reference Irmis, Martz, Parker and Nesbitt2010, Reference Irmis, Mundil, Martz and Parker2011; Olsen et al. Reference Olsen, Kent and Whiteside2011; Martz et al. Reference Martz, Mueller, Nesbitt, Stocker, Parker, Atanassov, Fraser, Weinbaum and Lehane2013, this volume). If the Santa Rosa Sandstone is equivalent to the Norian-aged Shinarump Member as demonstrated by detrital zircon signatures (Riggs et al. Reference Riggs, Lehman, Gehrels and Dickinson1996), then that is additional supporting data for a Norian age of the base of the Dockum Group. Based on the Adamanian (Norian) age of the Post Quarry, which is within the stratigraphically higher Cooper Canyon Formation of the Dockum Group, the Santa Rosa Sandstone can be constrained to be older than the hypothesised age range of 220–215 Ma for the Post Quarry (Martz et al. Reference Martz, Mueller, Nesbitt, Stocker, Parker, Atanassov, Fraser, Weinbaum and Lehane2013, this volume).
2. Material and methods
The specimen described here (TTU P-00539) is preserved in two parts: one part includes the posterior portion of the skull roof including the frontals, parietals, squamosals, the dorsal portions of the quadrates and quadratojugals, and the braincase; and the second part includes the region around the nares and antorbital fenestrae including the anterior portions of the jugals and lacrimals, the right ectopterygoid, parts of the pterygoids, the palatines, maxillae, nasals, ‘septomaxillae,’ and posterior portions of the premaxillae. No recognisable portions of the prefrontals, postfrontals, or postorbitals are present. No mandibles or postcrania were collected with this specimen.
Original preparation methods for TTU P-00539 are not fully known. However, the specimen already was prepared and coated with what was interpreted as an orange-coloured shellac prior to its original description (W. Langston, pers. comm. 2011). In order to remove that material, Langston scrubbed the specimen with rags dipped in acetic acid. A coating of Butvar B-78 was applied over the outer surface of the entire skull in the late 1990s, and both segments were moulded in March of 2010 (B. Mueller, pers. comm. 2011).
3. Systematic palaeontology
Archosauriformes Gauthier et al., Reference Gauthier, Kluge and Rowe1988
Phytosauria Meyer, Reference Meyer1861, sensu Doyle & Sues, Reference Doyle and Sues1995
Wannia gen. nov.
Type species.Wannia scurriensis (Langston Reference Langston1949).
Etymology. For Dr. Wann Langston, Jr., Professor Emeritus at The University of Texas at Austin in honour of his extensive work on archosaur palaeontology. This species was the first that Dr. Langston described and named in a peer-reviewed scientific publication (Langston Reference Langston1949).
Diagnosis. Same as for only known species.
Occurrence. Camp Springs Formation, Dockum Group, Scurry County, Texas.
Wannia scurriensis comb. nov.
Figure 3 Type specimen of Wannia scurriensis gen. nov. (TTU P-00539): (A) antorbital region of skull in right lateral view with line drawing interpretation. Anterior is to the right; (B) antorbital region of skull in left lateral view with line drawing interpretation. Anterior is to the left. Abbreviations: AOF=antorbital fenestra; ect=ectopterygoid; j=jugal; la=lacrimal; mx=maxilla; n=nasal; pal=palatine; pm=premaxilla; ‘sm’ = ‘septomaxilla’; t.=tooth. Scale bar=1 cm.
Figure 4 Type specimen of Wannia scurriensis gen. nov. (TTU P-00539): (A) antorbital region of skull in dorsal view with line drawing interpretation; (B) antorbital region of skull in ventral view with line drawing interpretation. Anterior is to the top of the page. Abbreviations: AOF=antorbital fenestra; CH=choana; ect=ectopterygoid; j=jugal; la=lacrimal; m.=matrix; mx=maxilla; n=nasal; NA=nares; pal=palatine; pm=premaxilla; ‘sm’=‘septomaxilla’; SOF=suborbital fenestra; v=vomer. Scale bar=1 cm.
Figure 5 Type specimen of Wannia scurriensis gen. nov. (TTU P-00539). Braincase region of skull in posterodorsal view with line drawing interpretation. Abbreviations: bo=basioccipital; bs=basisphenoid; FM=foramen magnum; m.=matrix; oto=otooccipital; p.f.=possible parietal foramen; pa=parietal; par=paroccipital process of the opisthotic; pt=pterygoid; PTF=posttemporal fenestra; q=quadrate; so=supraoccipital; sq=squamosal; STF=supratemporal fenestra. Scale bar=1 cm.
Figure 6 Type specimen of Wannia scurriensis gen. nov. (TTU P-00539). Braincase region of skull in posterior view with line drawing interpretation. Abbreviations: bo=basioccipital; bt=basisphenoid; fo=foramen ovale; FM=foramen magnum; m.=matrix; mf=metotic foramen; oto=otooccipital; pa=parietal; par=paroccipital process of the opisthotic; pt=pterygoid; PTF=post-temporal fenestra; q=quadrate; so=supraoccipital. Scale bar=1 cm.
Figure 7 Type specimen of Wannia scurriensis gen. nov. (TTU P-00539). Braincase region of skull in ventral view with line drawing interpretation. Abbreviations: bo=basioccipital; bs=basisphenoid; CN I=cranial nerve I; CN II=cranial nerve II; epi=epipterygoid; ls=laterosphenoid; m.=matrix; pa=parietal; pbs=parabasisphenoid rostrum; pr=prootic; pt=pterygoid; PTF=posttemporal fenestra; q=quadrate; sq=squamosal. Scale bar=1 cm.

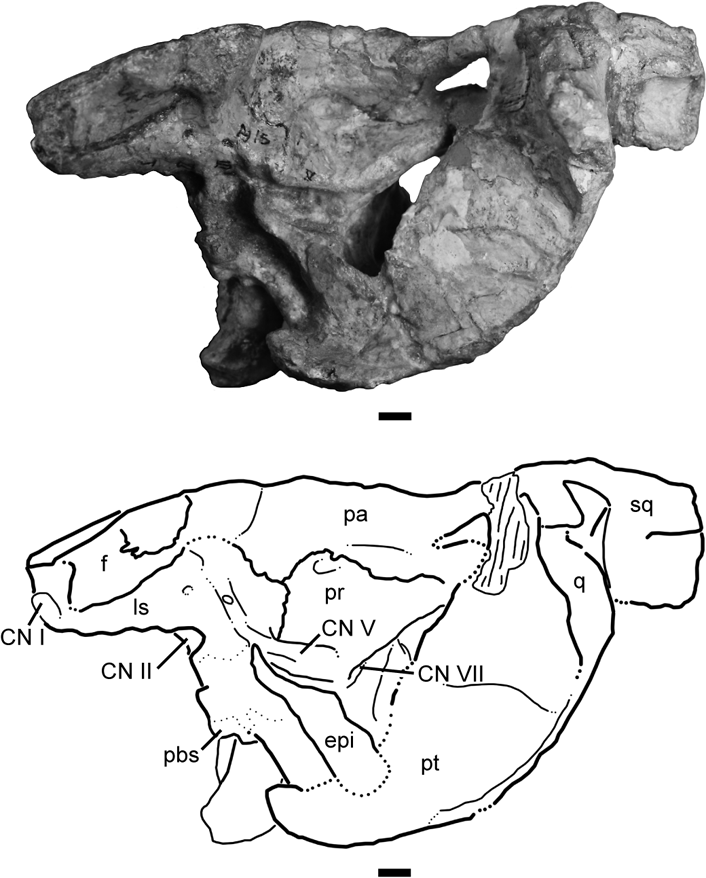
Figure 8 Type specimen of Wannia scurriensis gen. nov. (TTU P-00539). Braincase region of skull in left lateral view with line drawing interpretation. Abbreviations: bs=basisphenoid; CN I=cranial nerve I; CN II=cranial nerve II; CN V=cranial nerve V; CN VII=cranial nerve VII; epi=epipterygoid; f=frontal; ls=laterosphenoid; pa=parietal; pbs=parabasisphenoid rostrum; pr=prootic; pt=pterygoid; q=quadrate; sq=squamosal. Scale bars=1 cm.
Holotype. TTU P-00539, partial skull preserved in two parts.
Revised diagnosis.Wannia differs from all other phytosaurs in the following characters: basitubera that are widely separated mediolaterally; presence of a ridge on the lateral surface of the jugal; presence of a thickened shelf along the posteroventral edge of an expanded pterygoid–quadrate wing; ‘septomaxillae’ that do not contact one another and do not form part of the internarial septum; and presence of a nasal swelling posterior to the posterior borders of the nares. Wannia shares the following characters with all other phytosaurs (based on cladistic analyses by Stocker (Reference Stocker2010), Nesbitt (Reference Nesbitt2011) and Stocker (Reference Stocker2012)): nares directed dorsally (Stocker Reference Stocker2010: 1[1]); nares nonterminal (Stocker Reference Stocker2010: 2[1+2]; Nesbitt Reference Nesbitt2011: 139[1+2]); interpremaxillary fossa present (Stocker Reference Stocker2010: 8[1+2]); facial portion of the maxilla anterior to anterior edge of antorbital fenestra equal in length or longer than portion posterior to anterior edge of fenestra (Nesbitt Reference Nesbitt2011: 14[1]); anterior extent of the maxilla anterior to the nasals (Nesbitt Reference Nesbitt2011: 19[1]); posterior portion of the maxilla ventral to the antorbital fenestra expands dorsoventrally at the posterior margin of the maxilla (Nesbitt Reference Nesbitt2011: 27[2]); dorsal head of the quadrate has a sutural contact with the paroccipital process of the opisthotic (Nesbitt Reference Nesbitt2011: 77[1]); presences of separate ossification (=‘septomaxilla’) anterior to the nasals surrounded by the premaxilla (Nesbitt Reference Nesbitt2011: 150[1]). Wannia scurriensis differs from leptosuchomorph phytosaurs in possessing supratemporal fenestrae that are in the same horizontal plane as the skull roof (Stocker Reference Stocker2010: 32[0]) and are exposed dorsally (Stocker Reference Stocker2010: 33[0]); shared with Paleorhinus bransoni, Paleorhinus angustifrons, Ebrachosuchus neukami, ‘Paleorhinus’ sawini, Angistorhinus, and Brachysuchus megalodon). Wannia differs from Angistorhinus and Brachysuchus megalodon in possessing nares with their posterior borders anterior to the anterior borders of the antorbital fenestrae (Stocker Reference Stocker2010: 2[1]); shared with Paleorhinus bransoni, Paleorhinus angustifrons, and Ebrachosuchus neukami). Wannia scurriensis differs from Angistorhinus, Brachysuchus megalodon, and ‘Paleorhinus’ sawini in having parietal–squamosal bars that trend straight posterior to their attachments on the squamosals (Stocker Reference Stocker2010: 34[0]); shared with Paleorhinus bransoni, Paleorhinus angustifrons, and Ebrachosuchus neukami). Wannia scurriensis differs from Paleorhinus bransoni in lacking a premaxilla–palatine contact (Stocker Reference Stocker2010: 42[0]); shared with Paleorhinus angustifrons and Ebrachosuchus neukami). Wannia scurriensis differs from Paleorhinus bransoni and Paleorhinus angustifrons in the presence of a ridge, rather than a row of nodes, on the lateral surface of the jugal.
4. Morphological description
4.1. Premaxilla
The premaxillae are incomplete; they are broken transversely at the lateral exposure of the premaxilla-maxilla suture (Figs 3, 4). I infer from the preserved portions that the premaxillae are narrow mediolaterally and possibly tube-like. The presence or absence of a premaxillary crest cannot be confirmed. The contact between the premaxilla and maxilla is a zig-zag suture on the lateral surface of the rostrum, with an anterior process of the maxilla inserted between a smaller, lower process and a larger, posteriorly-directed ascending process of the premaxilla. That posterior process does not appear to be as extensive on the right side as on the left (Fig. 3A); on the left side, the laterally-directed ascending process of the premaxilla extends posteriorly almost to the posterior margin of the nares (Fig. 3B). Narrow processes of the premaxillae comprise approximately three-fourths of the anteroposterior length of the internarial septum; the nasals contribute the remaining one-fourth of the septum. The premaxillae are separated from the medial margins of the nares by thin extensions of both the ‘septomaxillae’ and nasals.
On the palatal surface, alveolar ridges extend anteriorly as sharp crests on either side of a wide, rounded interpremaxillary groove. Palatal processes of the premaxillae posteriorly extend to the choanae and contribute to their anteromedial margins; the choanae are located slightly more posterior than the nares. The premaxilla does not contact the palatine in Wannia scurriensis, whereas the premaxilla and palatine contact in all other North American phytosaurs I examined. A premaxilla–palatine contact previously was cited as a synapomorphy of Phytosauria (Sereno Reference Sereno1991). However, Paleorhinus angustifrons, Ebrachosuchus neukami, and the Krasiejów phytosaur also lack a premaxilla-palatine contact (Stocker pers. obs. 2011). The anteriormost alveolus present on the right side, and possibly the anteriormost on the left, are preserved in small, posterior fragments of the premaxillae that remain articulated with the maxillae. Those alveoli are small and rounded. No teeth appear to be preserved in any of the premaxillary alveoli.
4.2. Maxilla
The maxilla is bordered dorsally by the lateral ascending process of the premaxilla and by the nasal posterior to the posterior tip of the premaxilla. The maxilla surrounds the anterior border of the antorbital fenestra; an antorbital fossa is present on the maxilla anterodorsally (Fig. 4A) but not on the maxilla at the ventral border of the fenestra (Fig. 3). The lateral surfaces of the maxillae are oriented dorsoventrally and are marked by multiple small foramina randomly distributed anterior to the antorbital fenestra. The maxillae gently widen posterior to the anterior borders of the antorbital fenestrae. There they become much wider mediolaterally, though the posterior processes do not increase in dorsoventral thickness. The suture with the jugal is directed posteroventrally from the posterior edge of the antorbital fenestra and is clearly visible on the right side.
Ventrally, the maxillae have narrow palatal processes. The alveolar ridges continue posteriorly from the premaxillae, but the ridges remain low and are not visible in lateral view. The maxillae gently widen posteriorly until approximately the eighth alveolus from the posterior end of the tooth row. There, the maxillae widen much more dramatically and have laterally convex lateral margins around the last five alveoli. The posterior tip of the maxilla contacts the ectopterygoid and the jugal in ventral view. The maxilla forms the anterolateral margin of the suborbital fenestra. Just anterior to the suborbital fenestra, at the lateral apex of the palatine, is another, smaller fenestra. This smaller fenestra also is bordered laterally by the maxilla but is cut medially into the palatine. Lateral to the posterior tips of the palatal processes of the premaxillae, the palatal processes of the maxillae form rounded margins of the choanae.
All preserved alveoli that were mechanically prepared are separated by bony septa. The lateral edges of the alveoli are not more ventrally expanded than their medial edges. There are 16 alveoli preserved in the right maxilla, and at least five of those alveoli preserve broken teeth. All alveoli are circular in outline, though their diameters vary. TTU P-00539 exhibits anterior maxillary alveoli that are smaller in diameter than more posterior maxillary alveoli and smaller than the few preserved premaxillary alveoli; Nicrosaurus kapffi also features a reduction in size of the anteriormost maxillary alveoli posterior to the maxilla-premaxilla suture (Hungerbühler Reference Hungerbühler2000). The lack of a mediolateral constriction of the rostrum at the premaxilla-maxilla suture indicates the presence of a bipartite dentition (Hungerbühler Reference Hungerbühler2000). The teeth preserved in the right maxilla are too fragmentary to make detailed observations; however, the two that are visible in the posterior portion of the maxilla appear to have long axes oriented mesiodistally with labial surfaces that are more laterally convex than their lingual surfaces.
4.3. ‘Septomaxilla’
The triangular ‘septomaxillae’ (see Stocker (Reference Stocker2010) for discussion of the homology of this element in phytosaurs) form the slightly tapered anterior borders of the nares. The anterior tips of the ‘septomaxillae’ do not extend as far anteriorly as the anterior processes of the maxillae. The dorsal surfaces of the ‘septomaxillae’ are faintly roughened, unlike the anteroposteriorly-grooved dorsal surfaces of the ‘septomaxillae’ of Paleorhinus angustifrons (BSPG 1931 X 502), Ebrachosuchus neukami (BSPG 1931 X 501), Brachysuchus megalodon (UMMP 10336), Angistorhinus grandis (FMNH UC 631), some Angistorhinus-like specimens from the Otis Chalk localities (e.g. TMM 31098-1, TMM 31100-1332), and Angistorhinus talainti (MNHN TAL 1). Medially, the ‘septomaxillae’ in TTU P-00539 are separated from each other completely by the ascending processes of the premaxillae (Fig. 4A), rather than sutured to each other along the midline and forming most or all of the internarial septum as Sereno stated was characteristic for phytosaurs (Sereno Reference Sereno1991, p. 16). Thin laminae continue posteriorly to form the medial margins of the nares. Laterally, the ‘septomaxillae’ contact the nasals to form part of the lateral borders of the nares.
4.4. Nasal
The anterior process of the nasal articulates with the ‘septomaxilla’ dorsally and the premaxilla ventrally. Medially, thin laminae project anteriorly lateral to the premaxillae to form the medial margins of the nares, and those laminae contact the ‘septomaxillae’, approximately 2 cm anteriorly along the anteroposterior length of the internarial septum. The nasals form the majority of the lateral borders of the nares, which are inclined anterodorsally. The lateral edges of the nares project more dorsally than the internarial septum, and this projection is more prominent posteriorly. At the squared-off posterior borders of the nares, the nasals form a dorsal prominence. However, it can be inferred that the nares would not have been dorsal to the level of the skull roof. A slight constriction in mediolateral width is present just ventral to the narial margins, and this exaggerates the posterior widening of the nares in dorsal view. Laterally, the nasals are sculptured slightly by faint grooves and ridges that are oriented dorsoventrally. The posterolateral portions of the nasals are broken away and, as currently preserved, the nasals do not form any part of the inset antorbital fossae. The dorsal margins of the antorbital fenestrae are too damaged to determine whether the nasals contributed to those margins or whether the maxillae excluded the nasals. Posterior to the nares, the nasals possess a concave morphology that is ornamented more heavily. The articulations with the frontals are not preserved.
4.5. Frontal
The frontals are broken across their midsections, and the anterior portions are missing. The posterior portions of the frontals are preserved in articulation with the parietals. The anteroposteriorly oriented interdigitating suture between the left and right frontal is raised on a low ridge along the preserved length. At the posterior termination of that suture, an irregular depression is present at the junction of the frontals and the parietals. The distance between the orbits was hypothesised to have been narrower in TTU P-00539 than in Paleorhinus bransoni (Langston Reference Langston1949). The amount of breakage across the frontals requires a cautious assessment of interorbital width in TTU P-00539, and the incomplete rostrum makes estimation of skull length difficult. The frontals were at least 25 mm wide versus a length of approximately 340 mm from the premaxilla-maxilla suture to the occipital condyle in Wannia scurriensis, whereas the measured widths in two specimens of Paleorhinus bransoni are approximately 38 mm wide versus an approximate length of 342 mm in TMM 31100-101, and approximately 27 mm wide versus an approximate length of 345 mm in TMM 31025-172.
4.6. Lacrimal
The lacrimals form a large antorbital fossa around the dorsal and posterior borders of the antorbital fenestra (Fig. 4). Dorsally, the lacrimal portion of the antorbital fossa contacts a contribution from the maxilla, thus appearing to exclude the nasal from the margin of the antorbital fenestra. Though both right and left lacrimals are incomplete posterior to their portions of the antorbital fossae, the left lacrimal is less distorted than the right. The smooth fossa surrounding the posterior corner of the antorbital fenestra is widest at the posteroventral portion. Along the ventral margin of the antorbital fenestra, the lacrimal is reduced to a thin, anteriorly-directed process that excludes the jugal from participation in the antorbital fenestra.
4.7. Jugal
The anterior rami of both left and right jugals articulate along an anterodorsally–posteroventrally oriented suture with the maxillae. The anterior ramus of the jugal extends anterior to the posterior corner of the antorbital fenestra, but it does not appear to participate in any part of the margin of the fenestra or the fossa. At the ventral expression of the maxilla–jugal suture the mediolateral width of the skull decreases, enhancing the swollen appearance of the posterior portions of the maxillae in ventral view. Posteriorly, the ventral margin of the jugal curves posterodorsally so that the posterior ramus of the right jugal is raised relative to the ventral margin of the maxilla and the ectopterygoid is visible in lateral view (Fig. 3). The most posterior portion of the jugal is preserved as a rod-like process, and the contact with the quadratojugal is not preserved. On the lateral surface of the jugal, a continuous ridge slopes posteriorly from the posterior corner of the antorbital fossa and along the posterior ramus of the jugal. This continuous ridge differs from the row of raised nodes present on the lateral surface of the jugal in Paleorhinus bransoni (e.g. TMM 31100-101), Paleorhinus angustifrons (BSPG 1931 X 502), and the Krasiejów phytosaurs (Dzik & Sulej Reference Dzik and Sulej2007).
4.8. Parietal
The parietals articulate along the midline of the skull roof with an interdigitating suture. At approximately midlength along that suture, an elongate pit is present (Fig. 5). This pit was hypothesised to be separated from the endocranial cavity (Langston Reference Langston1949), but infilled matrix prevents further definitive assessment of Langston's interpretation. The parietals do not greatly overhang the dorsal portion of the supraoccipital, but they form a slight lip at the posterior margin of the skull roof. The parietals have long, mediolaterally narrow squamosal processes that are dorsally exposed and in the plane of the skull roof. Small depressions are present in the dorsal surface of the parietals at the base of those processes. Along the posterolateral extension of the squamosal processes, the parietals clearly form mediolaterally thin extensions. It is unclear how far dorsoventrally the parietals expand. In posterior view, it appears that the parietals could be only the thin dorsal expressions of the squamosal processes (Fig. 6) and separate ossifications may be in place between the parietals and supraoccipital. However, sutures in this area often are difficult to delimit, and the dorsal edges of the parietal–squamosal bars are slightly damaged.
4.9. Squamosal
Only the posterodorsal corners of the squamosals are preserved. What remains of the squamosals appears to be ornamented dorsally with rugose ridges and grooves. On the dorsal surface of the right squamosal, a trace of an anteroposteriorly trending groove is preserved. The squamosals posteriorly enclose large supratemporal fenestrae that have rounded posterior borders in dorsal view (Fig. 5). The squamosals lack posterior processes; there is no posterior expansion of the squamosals beyond the paroccipital processes of the otoocciptials (see section 4.19 for note on nomenclature). There appears to be a long descending process of the squamosal anterior to the quadratojugal, but preservation in that area of the skull is poor.
4.10. Quadratojugal and quadrate
Only the dorsal-most portion of the right quadratojugal is preserved, and no new observations can be made. The dorsal portion of the right quadrate also is preserved. A tab-like flange is visible laterally and forms part of the dorsal and lateral edges of a large, posteriorly-oriented quadrate foramen. On both the left and right sides, the large, flat quadrate flanges of the pterygoid–quadrate wings are preserved. Those wings are oriented with their flat surfaces directed anterolaterally (Fig. 6). A faint suture is distinguishable on the anterior surface of each pterygoid–quadrate wing indicating the overlapping contact of the quadrate on the pterygoid. Portions of the sutures are visible posteriorly, but poor preservation prevents observation of the entire suture.
4.11. Vomer
Portions of the vomers may be present along the midline of the palate, but that area is poorly preserved. The articulated vomers appear to comprise the choanal septum, though a fracture separates the vomers from the premaxillae.
4.12. Palatine
The well-preserved palatines are triangular in ventral view. Their medial edges are slightly convex and diverge from each other posteriorly. They are approximately 3·5 cm apart anteriorly and approximately 7·5 cm apart at their posterior corners. Anteriorly, small points of the palatines insert into the maxillae at the widest points of the choanae. The interdigitating sutures between the anterior portions of the palatines and maxillae are well defined by matrix (Fig. 4B). Posteriorly, the connection between the palatines and maxillae becomes irregular with asymmetrical fenestrae present near the lateral apices of the palatines. The posterolateral edges of the palatines form the medial edges of the elongate suborbital fenestrae. Those fenestrae have long axes oriented posteromedially. On the left side the posterior-most portion of the palatine appears to be broken away, though the right is complete and articulates with the ectopyterygoid along a short straight suture. Portions of the dorsomedial flanges of the palatines are preserved and visible though it is difficult to distinguish palatine from pterygoid on the right side. A slight curved ridge is present along the length of the palatine on each side at the juncture between the palatal flange and the dorsomedial flange.
4.13. Pterygoid
The pterygoids are preserved on both parts of the skull as it is preserved currently. The right anterior process is articulated with the right palatine on the facial portion of the skull, and the posterior portions of both the left and right pterygoids are preserved in articulation with the quadrates and basipterygoid processes of the parabasisphenoid on the braincase portion of the skull. The anterior portion of the right pterygoid is difficult to observe because that area of the palate was not fully prepared; however, it does not appear that the pterygoids form the midline of the palate or the choanal septum.
Posteriorly, the pterygoids articulate with the quadrates by overlapping flanges on the dorsoventrally deep pterygoid-quadrate wings (Fig. 6). A sharp shelf is formed by the ventral edge of the pterygoids on their posterior surfaces. Those shelves become more pronounced toward the midline as they approach the basipterygoid processes, terminating by wrapping anteriorly around the basipterygoid processes with broken anterior edges.
4.14. Ectopterygoid
The base of the right ectopterygoid is preserved, articulated with the maxilla and jugal laterally, and the palatine and pterygoid medially and posteriorly (Figs 3, 4). The suborbital fenestra extends between the ectopterygoid and the palatine for approximately two-thirds of the length of the fenestra, and the posteriormost corner of the fenestra is incised into the ventral surface of the ectopterygoid. Along the edge of that fenestra, the ectopterygoid has a rough and undulating edge. In lateral view, the body of the ectopterygoid has a strong ventral curvature posterior to the articulation with the jugal and maxilla, so that the entirety of the element is visible in lateral view. The posterior process of the ectopterygoid is poorly preserved, and both the ectopterygoid foramen and the articulation with the pterygoid are broken away. Only the lateral portion of the left ectopterygoid is preserved, still articulated with the maxilla and a small section of the left jugal, but this side does not contribute additional information about the element.
4.15. Epipterygoid
Both left and right epipterygoids are preserved in articulation with the lateral surfaces of the endocranial region (Figs 7, 8). Each long, slender epipterygoid is angled approximately 45° anterodorsally from its ventral articulation with the pterygoid and basipterygoid process to its tapered dorsal point near the exit for the trigeminal nerve in the prootic. The epipterygoids do not appear to be rounded in cross-section but rather they have flat lateral surfaces.
4.16. Basioccipital and parabasisphenoid
The rounded occipital condyle is visible in dorsal view, protruding posteriorly beyond the edge of the supraoccipital, although the exoccipital portions of the otooccipitals meet to cover nearly the entire dorsal surface of the condyle and exclude the basioccipital from the floor of the foramen magnum. In posterior view, the basioccipital is a tall element (approximately 4·5 cm in dorsoventral height) with the basitubera completely visible ventral to the condyle. Ventrally, the basioccipital has a sharp edge separating the condyle from the anteroposteriorly short neck (Fig. 7). The basioccipital forms the posterior portions of the basitubera, and the suture with the parabasisphenoid appears to be exposed more posteriorly than ventrally. A deep anteroposterior cleft widely separates the tubera along the midline, and at the posteriormost expression of this cleft there appears to be a basioccipital recess (sensu Gower, Reference Gower2002). The basitubera extend further laterally than do the basipterygoid processes, unlike the morphology in Paleorhinus bransoni (TMM 31100-101) where the basipterygoid processes are wider than the basitubera.
The parabasisphenoid forms the anterior portions of the basitubera and has a much smoother surface than the basioccipital portions of the basitubera. The basitubera and basipterygoid processes in TTU P-00539 are anteroposteriorly separated from each other by no more than 1 cm; most of the separation between these structures is in a dorsoventral direction because the basipterygoid processes extend farther ventrally than do the basitubera. At the midline between the basitubera and the basipterygoid processes is a rounded, ventrally concave depression that may be homologous with the median pharyngeal recess of other archosaurs (Witmer Reference Witmer, Currie and Padian1997; Gower Reference Gower2002). The basipterygoid processes are surrounded laterally and anteriorly by portions of the pterygoids. The exits for the internal carotid arteries are not visible ventrally; the foramina should exit the parabasisphenoid laterally in phytosaurs (Gower Reference Gower2002), and in this specimen they are most likely blocked from view by the epipterygoids. Anteriorly, the cultriform process of the parabasisphenoid is in the same transverse plane as the basioccipital and a small raised tubercle is present on its anteroventral surface. The preserved portion of the cultriform process is broken just anterior to its base, although a portion of it may continue into the broken posterior portion of the palate in the other part of the specimen (Fig. 4B).
4.17. Laterosphenoid
The left and right laterosphenoids are articulated with the ventral surfaces of the frontals and parietals, and with the prootics posteriorly (Fig. 8). These complex elements enclose two anteriorly directed foramina for the passage of cranial nerves I and II (Fig. 8). Two additional small foramina are visible on the lateral surface of the left laterosphenoid; these may be for other cranial nerves (possibly CN III and IV), but it is unclear which nerves would pass through each foramen or if they are actually blind pits that do not communicate with the endocranium. A faint groove is present that crosses the posteriormost foramen on the lateral surface of the laterosphenoid, and this may have held a branch of cranial nerve V that exited dorsally from the prootic.
4.18. Prootic
The prootic is a large element that is exposed on the lateral surface of the braincase (Fig. 8). An extensive articulation with the parietal is present at the dorsal margin of the prootic, and a foramen (possibly for the dorsal head vein) sits in a deep pocket at the posterodorsal contact between the prootic and the parietal. Posteroventrally, the prootic forms the crista prootica, which dorsally covers cranial nerve VII and forms the anteromedial border of the pteroccipital fenestra in this specimen. The trigeminal foramen is oriented anterodorsally near the anteroventral edge of the prootic, but it is difficult to discern whether the prootic forms the entire foramen in this specimen.
4.19. Otooccipital
The opisthotic and exoccipital appear to be indistinguishably fused to one another; therefore, the two elements are described together here as the otooccipital (sensu Evans Reference Evans, Gans, Gaunt and Adler2008). The exoccipital pillars are angled posteroventrally to meet posteriorly on the occipital condyle and nearly cover the basioccipital in dorsal view. Posteriorly, the otooccipitals form nearly the entire border of the circular foramen magnum (Fig. 6). Small depressions are present on the ventrolateral surfaces of the pillars, and these are interpreted as the exits for cranial nerve XII. The anterior edges of the pillars form the posterior borders of the metotic foramina, and those foramina are each separated from the fenestra ovale on each side by the crista interfenestralis (=ventral ramus of the opisthotic; Gower Reference Gower2002; Nesbitt Reference Nesbitt2011).
The nearly unexpanded paroccipital processes extend posterolaterally from their bases and form the ventral borders of small, almond-shaped posttemporal fenestrae. Proximally, the otooccipitals contact the supraoccipital in approximately horizontal sutures. Distally, the paddle-like paroccipital processes contact the squamosals and quadrates. That area is too damaged on the left side to discern sutures among these elements, though on the right side the paroccipital process appears to articulate against the posteriormost portion of the squamosal dorsomedially without covering the dorsal surface of the squamosal. The distal edges of the paroccipital processes may have been expanded more posteriorly at their ventral corners, but erosion of the tips of both processes prevents direct confirmation.
4.20. Supraoccipital
The wide supraoccipital is dorsally exposed and barely contacts the dorsal edge of the foramen magnum, which instead is bordered almost completely by the exoccipital processes of the otooccipitals. The suture with the otooccipitals is nearly horizontal, and the supraoccipital tapers to a point at the median corners of the posttemporal fenestrae. On the dorsally-exposed surface, an anteroposterior ridge trends along the midline of the element and terminates just slightly under the posterior edge of the parietals (Fig. 5). There may be two small fenestrae on either side of the midline at the anterior end of the supraoccipital ridge, but this is difficult to interpret without further preparation.
5. Discussion
Many diagnoses for phytosaur taxa, including that of Wannia scurriensis, were published at a time when paraphyly was accepted and embraced as evidence of evolution of multiple lineages from an ‘ancestral stock’ (Simpson Reference Simpson1944). Specimens were assigned to taxa based on the presence of shared characters, irrespective of character polarity. With the introduction of cladistic methodology, there was a modification of taxonomic methods; shared characters were examined with respect to a single, common ancestry and went from defining a group or specimen as part of a particular ‘kind’ to diagnosing a monophyletic group (Rowe Reference Rowe1987). Because of this change in perception, characters that were thought previously to be synapomorphies of a group often were recognised as plesiomorphies for a clade. It is only from examinations that are grounded in an evolutionary framework of monophyly that it is possible to recognise those characters and analyse sequences of evolutionary character transformations and speciation events. Reassessment of those character state distributions in the framework of monophyly will limit arbitrary and unnatural taxonomic identifications and will form the basis for testable diagnoses.
The original species-level diagnosis of Wannia scurriensis was based on: its small size, the anterior position of the nares; a laterally-visible, though posteriorly-positioned, quadrate foramen; dorsoventral flattening of the skull; an internarial septum composed of premaxillae and nasals; ‘septomaxillae’ not meeting on the midline; exceptionally large palatine foramina (=suborbital fenestrae); elongate posttemporal fenestrae; and presence of a persistent parietal foramen in immature individuals (Langston Reference Langston1949). However, only a single specimen (TTU P-00539) was known at the time of publication. Of those characters, some have potential phylogenetic utility but were not examined in the context of all known phytosaur specimens or non-phytosaurian archosauriform outgroups.
Characters traditionally utilised to diagnose Paleorhinus must be re-examined now that most are demonstrated to be distributed across a grade of taxa at the base of Phytosauria (Hungerbühler Reference Hungerbühler1998; Stocker Reference Stocker2010, Reference Stocker2012). An anterior position of the nares with respect to the antorbital fenestrae was listed as a diagnostic character of ‘Paleorhinus’ scurriensis, suggesting “a close relationship to the genus Paleorhinus” (Langston Reference Langston1949, p. 325). However, nares completely anterior to the antorbital fenestrae characterise not just all non-phytosaurid phytosaurs, but archosauriforms generally (e.g. Stocker Reference Stocker2010; Nesbitt Reference Nesbitt2011). Phytosaurid phytosaurs such as Angistorhinus and Machaeroprosopus have nares with posterior borders that are either parallel with, or posterior to, the anterior borders of the antorbital fenestrae (Stocker Reference Stocker2010), and it is the plesiomorphic lack of this character that characterises non-phytosaurid phytosaurs. Using plesiomorphic characters to identify all basal phytosaurs as ‘Paleorhinus’ masks what is actually a much greater diversity of non-phytosaurid phytosaurs than was recognised previously.
The “apparent presence of a persistent parietal foramen” and the “uncrowded arrangement” of the maxillary dentition were identified as morphological features that could be indicative of immaturity in TTU P-00539 (Langston Reference Langston1949, p. 325). Currently, morphological transformations of phytosaur external anatomy that are associated with ontogeny are limited to features of the vertebrae (Irmis Reference Irmis2007). The presence of a parietal foramen in phytosaurs is a character that remains unresolved. In addition to the holotype specimen of Wannia scurriensis, a parietal foramen was reported as present for a small, potentially immature specimen referred to Smilosuchus lithodendrorum (UCMP 27181; Camp Reference Camp1930), whereas the holotype of ‘Paleorhinus’ magnoculus (MNHN ALM 1) possesses multiple morphological features that indicate its juvenile status (Fara & Hungerbühler Reference Fara and Hungerbühler2000) and does not preserve a parietal foramen. In the holotype specimen of Pravusuchus hortus (AMNH FARB 30646), there is a small dorsal convexity of the parietals in this location, but it is not open dorsally (Stocker Reference Stocker2010). The absence of a parietal foramen is a synapomorphy of Archosauriformes (Nesbitt Reference Nesbitt2011), and other than the potential presence in UCMP 27181 and TTU P-00539, no archosauriforms show a reversal of this character. Rigorous evaluation of this feature in phytosaurs requires evaluation of external and internal cranial morphology to determine whether the openings in the parietals of TTU P-00539 and UCMP 27181 are natural structures associated with immaturity, teratologies, or the result of preparation methods.
Both Camp (Reference Camp1930) and Colbert (Reference Colbert1947) noted that smaller phytosaur specimens tended to have more widely spaced alveoli, and in larger specimens the dentition became ‘crowded’ and small alveoli often were obliterated. Where the maxillary alveoli are free from matrix in TTU P-00539, bony divisions separate the alveoli, and no preserved alveoli in TTU P-00539 appear to be confluent. However, maxillary alveoli are also separated by septa in Smilosuchus adamanensis (UCMP 26699) and Machaeroprosopus pristinus (sensu Parker et al., Reference Parker, Hungerbühler and Martz2013, this volume; MU 525), though the maxillary alveoli in Smilosuchus gregorii (UCMP 27200) are separated by thin septa or are confluent. Among other archosauriforms, the maxillary alveoli in Euparkeria capensis also are separated by bony septa (Senter Reference Senter2003). The presence of septa, and their thicknesses, appears to be a complex relationship between ontogeny, overall skull size, and robustness of the rostrum. Collection and careful preparation of new, small specimens can provide the basis for a badly needed examination of skeletal transformations associated with ontogeny in phytosaurs.
5.1. Comparisons with other non-phytosaurid phytosaurs
Wannia scurriensis, Paleorhinus angustifrons (BSPG 1931 X 502), Ebrachosuchus neukami (BSPG 1931 X 501), and the Krasiejów phytosaur (Dzik & Sulej Reference Dzik and Sulej2007) lack a premaxilla–palatine contact, though the premaxilla and palatine contact each other in Paleorhinus bransoni (TMM 31100-101). All other known phytosaurs also have a premaxilla–palatine contact.
In Wannia scurriensis, the internarial septum is formed mainly by the premaxillae with a small contribution supplied by the nasals, whereas in Paleorhinus bransoni (TMM 31100-101) and Paleorhinus angustifrons (BSPG 1931 X 502), the ‘septomaxillae’ contact the nasals and form the posterior portions of the internarial septum. The ‘septomaxillae’ of Wannia scurriensis do not contact each other, are separated completely by the premaxillae and are limited to the anterior borders of the nares, whereas in Paleorhinus bransoni (TMM 31100-101, TMM 31025-172) and possibly ‘Paleorhinus’ sawini (TMM 31213-16), the ‘septomaxillae’ contact each other posteriorly, and in leptosuchomorphs (e.g. UCMP 27200, PEFO 34852), the ‘septomaxillae’ contact along nearly their entire length.
The large swelling of the nasals surrounding the posterior borders of the nares is unique in Wannia scurriensis. A slightly similar swelling is present in Paleorhinus angustifrons (BSPG 1931 X 502), but posterior to the swelling in P. angustifrons is a depression that is not present in Wannia (TTU P-00539). Paleorhinus bransoni (TMM 31100-101), ‘Paleorhinus’ sawini (TMM 31213-16), and Ebrachosuchus neukami (BSPG 1931 X 501) lack a swelling of the nasals posterior to the nares.
The antorbital fenestrae in Wannia scurriensis nearly are surrounded by wide antorbital fossae that are also seen in Paleorhinus angustifrons (BSPG 1931 X 502). The long axis of the antorbital fenestra in W. scurriensis is angled posterodorsally, whereas the fenestrae in P. angustifrons and Ebrachosuchus neukami are approximately parallel with the horizontal ventral margin of the skull. The antorbital fossa in Wannia scurriensis nearly surrounds the antorbital fenestra except for a small area on the dorsal edge of the ventral margin as formed by the maxilla; a similar, though not quite as wide exposure is observed in Paleorhinus bransoni and P. angustifrons, and is much more extensive than that in Ebrachosuchus neukami (BSPG 1931 X 501), where it is not expressed on the dorsal border of the antorbital fenestra.
A ridge present on the lateral surface of the posterior ramus of the jugal in Wannia scurriensis is similar to the ridge present on the lateral surface of the jugal in the proterochampsid Chañaresuchus and the early theropod dinosaurs Herrerasaurus ischigualastensis and Coelophysis bauri (Nesbitt Reference Nesbitt2011: 75[1]). In Paleorhinus bransoni (TMM 31100-101, TMM 31025-172), Paleorhinus angustifrons (BSPG 1931 X 502), and the Krasiejów phytosaur (Dzik & Sulej Reference Dzik and Sulej2007) the lateral surface of the jugal is ornamented with a row of low nodules rather than a ridge, in Ebrachosuchus neukami (BSPG 1931 X 501) there is a raised rugose surface in this area of the jugal, and in ‘Paleorhinus’ sawini (TMM 31213-16) and some leptosuchomorph phytosaurs (e.g. Leptosuchus studeri: UMMP 14267; Smilosuchus adamanensis: UCMP 26699) the surface is unadorned.
The suborbital fenestra in Wannia scurriensis is long and straight as in Ebrachosuchus neukami (BSPG 1931 X 501), though it is wider in Wannia scurriensis; in Paleorhinus angustifrons (BSPG 1931 X 502), the suborbital fenestra has two main axes that form an approximately 150° angle, and in the Krasiejów phytosaur (ZPAL Ab III 200, 1943, Dzik & Sulej Reference Dzik and Sulej2007) the suborbital fenestra is curved. The basitubera of Wannia scurriensis are widely separated as in Paleorhinus angustifrons (BSPG 1931 X 502), but they are rounded in Wannia scurriensis. The basitubera of Paleorhinus bransoni (TMM 31025-172, TMM 31100-101) and Ebrachosuchus neukami (BSPG 1931 X 501) are wider, not as separated from each other, and possess a weakly-developed mediolateral ridge seen in leptosuchomorph phytosaurs (e.g. UMMP 14267, AMNH FARB 30646, UCMP 26699).
Wannia scurriensis bears a thickened, sharp shelf along the posteroventral edge of the deeply expanded pterygoid–quadrate wing; in Paleorhinus bransoni (TMM 31100-101), Paleorhinus angustifrons (BSPG 1931 X 502) and Ebrachosuchus neukami (BSPG 1931 X 501), the posteroventral edge of the unexpanded pterygoid–quadrate wing does not have a shelf, although there is a slight posterior curvature to the quadrate portion of the wing near the quadrate body. The pterygoid–quadrate wings of Paleorhinus angustifrons (BSPG 1931 X 502) and Ebrachosuchus neukami (BSPG 1931 X 501) likely were affected by dorsoventral compression.
A small partial skull previously catalogued as ‘Paleorhinus’ scurriensis (TTU P-11422) does not share any diagnostic characters with the holotype specimens of Wannia scurriensis. TTU P-11422 does not preserve the braincase or palate, and the only portion of this specimen that includes elements also found in TTU P-00539 is the area surrounding the right antorbital fenestra. TTU P-11422 does have a large antorbital fossa with a slightly posterodorsally inclined antorbital fenestra, but the nasals just posterior to the nares are not swollen. The skull is incomplete and embedded in a hard matrix, but further preparation of TTU P-11422 may reveal additional morphological details that would facilitate comparisons with other non-phytosaurid phytosaurs.
Wannia scurriensis clearly differs from other North American phytosaurs (Stocker Reference Stocker2010, Reference Stocker2012). In previously published analyses of mainly North American phytosaurs (Stocker Reference Stocker2010, Reference Stocker2012), Wannia scurriensis was shown to lack a contact between the premaxilla and palatine but does possess: an internarial septum formed mainly by the premaxillae; ‘septomaxillae’ that do not contact one another and; much more separated basitubera. All other phytosaurs included in those analyses have: a premaxilla–palatine contact; have ‘septomaxillae’ that contact and form at least some part of the internarial septum; and have basitubera that are closer together and have a mediolaterally-oriented ridge on their ventral surfaces. However, additional comparisons with the non-phytosaurid phytosaurs Paleorhinus angustifrons, the Krasiejów phytosaur, and Ebrachosuchus neukami reveal a more complex distribution of morphological characters at the base of Phytosauria.
5.2. Distribution and correlations
Non-phytosaurid phytosaurs are not confined to a single depositional basin or regional area but are found in both the Dockum Group of Texas and the Popo Agie Formation of Wyoming in North America (Williston Reference Williston1904; Langston Reference Langston1949; Hunt & Lucas Reference Hunt and Lucas1991; Long & Murry Reference Long and Murry1995). Specimens from Poland (Dzik & Sulej Reference Dzik and Sulej2007), Germany (Paleorhinus angustifrons and Ebrachosuchus neukami), India (Parasuchus hislopi; Chatterjee Reference Chatterjee1978), and Morocco (Fara & Hungerbühler Reference Fara and Hungerbühler2000) reveal a nearly worldwide distribution of early-branching phytosaur taxa (Hunt & Lucas Reference Hunt and Lucas1991). This indicates that the clade was already widespread by the time the earliest recognised phytosaur fossils from the Late Triassic. However, this study demonstrates that the early radiation was not a diversification of a single basal clade of phytosaurs; rather it represents multiple cladogenetic events along the spine of Phytosauria. The results of recent cladistic analysis of Archosauriformes (Nesbitt Reference Nesbitt2011) incorporating Xilosuchus (Nesbitt et al. Reference Nesbitt, Liu and Li2011) and Ctenosauriscus (Butler et al. Reference Butler, Brusatte, Reich, Nesbitt, Schoch and Hornung2011) support an Early Triassic time of divergence for Phytosauria. The lack of known phytosaur specimens from the Early and Middle Triassic is a limiting factor with respect to the understanding of character transformations in the early history of the clade.
The occurrences of phytosaur specimens are integral in the biostratigraphic and biochronologic correlation of Late Triassic deposits because of the paucity of numerical dates for that time. Although more radioisotopic dates were inferred recently (Irmis & Mundil Reference Irmis and Mundil2008; Mundil et al. Reference Mundil, Pálfy, Renne, Brack and Lucas2010; Irmis et al. Reference Irmis, Mundil, Martz and Parker2011; Ramezani et al. Reference Ramezani, Hoke, Fastovsky, Bowring, Therrien, Dworkin, Atchley and Nordt2011), there are still few localities where both phytosaur specimens and numerical dates are known or can at least be correlated unambiguously using lithostratigraphic data. For the early part of the Late Triassic, several localities previously were correlated with the Otischalkian land-vertebrate faunachrons (LVFs) based on the presence of Paleorhinus (Hunt & Lucas Reference Hunt and Lucas1991; Lucas & Hunt Reference Lucas, Hunt, Lucas and Morales1993; Lucas Reference Lucas1998). The Placerias Quarry in the Chinle Formation of Arizona was correlated with the Adamanian based on the presence of “Rutiodon” (Lucas et al. Reference Lucas, Heckert and Hunt1997). However, an isolated narial region of a phytosaur rostrum (MNA V3728; Long & Murry Reference Long and Murry1995) identified as Paleorhinus based on an anterior position of the nares (Hunt & Lucas Reference Hunt and Lucas1991), created a potential overlap of Paleorhinus and “Rutiodon” (Lucas et al. Reference Lucas, Heckert and Hunt1997). That identification was criticised (Padian Reference Padian, Fraser and Sues1994; Irmis Reference Irmis2005) because it was based on morphology that is plesiomorphic for Archosauriformes, and Irmis (Reference Irmis2005) suggested that MNA V3728 could be constrained only as a non-phytosaurid phytosaur. The posterior borders of the nares of MNA V3728 are not unambiguously anterior to the anterior border of the antorbital fenestra as they are in Wannia, Paleorhinus, and Ebrachosuchus. Instead, the posterior borders of the nares in MNA V3728 are just posterior to the anterior borders of the antorbital fenestrae (Stocker Reference Stocker2010: 2[2]), though not as far posterior as is observed for Brachysuchus (UMMP 10336), Angistorhinus (FMNH UC 631), or leptosuchomorph phytosaurs. The position of the nares in MNA V3728 most closely resembles that of the holotype of ‘Paleorhinus’ sawini (TMM 31213-16). Therefore, MNA V3728 is not diagnosable as Paleorhinus, but clearly it does not share the morphology of other non-phytosaurid phytosaurs either. Furthermore, this modified identification removes any possible correlation between the Placerias Quarry in the Chinle Formation and the Otis Chalk localities in the Dockum Group based on Paleorhinus. This may mean that much of our understanding of the early part of the Late Triassic was compressed artificially to a shorter time interval because of the previous practice of identifying all non-phytosaurid phytosaurs as Paleorhinus. More accurate specimen identifications based on unique combinations of characters and autapomorphies in a phylogenetic systematic context will yield more refined biostratigraphic and biochronologic signals for the early Late Triassic and will lead to a more robust understanding of the early history of the Phytosauria.
Resolution of the taxonomic and phylogenetic issues that have plagued Paleorhinus for nearly a century has important consequences for archosauriform relationships. Recognising the species previously referred to Paleorhinus as a paraphyletic grade documents extensive, previously hidden morphological diversity at the base of Phytosauria. Moreover, fragmentary specimens cannot be diagnosed as Paleorhinus based on characters demonstrated to be plesiomorphic for Phytosauria. Recovery of previously referred species of Paleorhinus as a paraphyletic assemblage of taxa consequently impacts the use of Paleorhinus for early Late Triassic biostratigraphy. Because Paleorhinus is the current index taxon for the Otischalkian IVF (Hunt & Lucas Reference Hunt and Lucas1991; Lucas & Hunt Reference Lucas, Hunt, Lucas and Morales1993; Lucas Reference Lucas1998), a reassessment of all specimens previously identified as Paleorhinus is required before their continued use for Late Triassic biostratigraphy and biochronology can be supported or sustained.
6. Acknowledgments
This project would not have proceeded were it not for the scientific generosity of Dr. Wann Langston, Jr., whose library was always open and who shared much of his palaeontological knowledge with me. Dr. Langston worked on an extensive diversity of topics and taxa, but his first published description was on phytosaurs. I thank W. Parker for the invitation to contribute to this volume. This project is part of dissertation research completed in partial fulfillment of a PhD at The University of Texas at Austin under the guidance of and discussion with C. Bell, J. Clarke, T. Rowe, E. C. Kirk and G. Gunnell. R. Butler, K. Claeson, A. Kennedy, B. Mueller and S. Nesbitt provided additional helpful discussion that greatly improved this manuscript. S. Chatterjee, R. Ethington, M. Goodwin, G. Gunnell, C. Holliday, P. Holroyd, C. Mehling, B. Mueller, L. Murray, M. Norell, K. Padian, W. Parker, O. Rauhut, T. Rowe, J. C. Sagebiel, W. Sanders, B. Simpson, M. Smith, H.-D. Sues, T. S. Williams and J. Wilson provided access to specimens in their care. Randall Irmis and Richard Butler provided helpful reviews that greatly improved the quality of this manuscript. Funding from the University of California Museum of Paleontology Doris and Samuel P. Welles Fund, the Jackson School of Geosciences, The University of Texas at Austin Lundelius Fund and The University of Texas at Austin William Powers Jr. Presidential Graduate Fellowship supported this research.