
The Nectridea is a clade of small, mostly aquatic tetrapods occurring in the Lower Pennsylvanian to Permian of North America, Europe and Morocco comprising three families: Urocordylidae, Diplocaulidae and Scincosauridae. Most nectrideans were less than 500mm long, with relatively short trunks and long tails, and possessed characteristic holospondylous vertebrae with fused neural and haemal arches and spines. Expanded fan-shaped haemal spines conferred a long, laterally flattened tail that was adapted primitively for subcarangiform swimming. Nectrideans were traditionally grouped together with microsaurs and aïstopods as Lepospondyli on the basis of the shared holospondylous vertebra (Carroll et al. Reference Carroll, Bossy, Milner, Andrews and Wellstead1998). However, the phyletic unity of lepospondyls was questioned by Thomson & Bossy (Reference Thomson and Bossy1970) and Bossy (Reference Bossy1976), who suggested that holospondylous vertebrae are the product of functional convergence and the lepospondyl concept might no longer be considered valid. However, a monophyletic lepospondyl clade was found by Ruta et al. (Reference Ruta, Coates and Quicke2003) and Ruta & Coates (Reference Ruta and Coates2007), although lepospondyls were regarded as polyphyletic by Pardo et al. (Reference Pardo, Szostakwiskji, Ahlberg and Anderson2017, Reference Pardo, Carter, Lennie, Sallan, Anderson , Murray and Holmes2018).
The diplocaulids, or horned nectrideans, are distinguished by long posteriorly or posterolaterally directed tabular horns. The Lower Pennsylvanian diplocaulids – together, representatives of the urocordylids and aïstopods – were members of equatorial coal-swamp assemblages in Europe and North America. The earliest occurrence is the Langsettian fauna from Jarrow Colliery, Kilkenny, Ireland. The Jarrow locality is, thus, of great historical importance to the understanding and interpretation of coal-swamp assemblages (Milner Reference Milner1980b). The Huxley & Wright (Reference Huxley and Wright1867) description of the Jarrow non-amniote tetrapods was the first detailed and illustrated account of a coal-swamp assemblage and provided the basis for interpreting the later Middle Pennsylvanian coal-swamp fauna from Linton, Ohio, first discovered in the 1850s (Newberry Reference Newberry1867).
Little has been published on the Jarrow nectrideans since Huxley & Wright's (Reference Huxley and Wright1867) monograph, apart from Steen (Reference Steen1938), a brief review by Milner (Reference Milner and Panchen1980a) and a comparative review of the Nectridea by Bossy & Milner (Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998). The diplocaulid Keraterpeton galvani is the commonest of the 11 taxa identified in the Jarrow assemblage (Sequeira Reference Sequeira1996). A detailed morphological account is given here, drawing on unpublished data from Milner (Reference Milner1978) and additional specimens subsequently made available for study. A second species, Keraterpeton longtoni from the Bolsovian (Westphalian C) of the North Staffordshire Coalfield, is described and newly figured.
Specimens are held in the following institutions: British Geological Survey (BGS), Sedgwick Museum of Earth Sciences, University of Cambridge (CAMSM), Geological Survey of Ireland (GSI), Museum of Comparative Zoology, Harvard University (MCZ), The Natural History Museum, London (NHMUK), National Museum of Ireland New Geology Collection (NMING), National Museums of Scotland (NMS), Trinity College Dublin (TCD) and Warwickshire Museums Service (WARMS).
1. The Jarrow Carboniferous coal-swamp fauna
1.1. History of discovery and collection
Amphibian fossils were first discovered and collected in 1864 by W. B. Brownrigg from Jarrow Colliery in Kilkenny, Republic of Ireland. He identified six or seven forms that he presumed were reptiles (Brownrigg Reference Brownrigg1865). The fossils derived from shales roofing a deep oxbow in the Jarrow Coal seam, the so-called ‘Jarrow Channel', lying 64m below ground. The Jarrow Coal was worked from 1853, but until 1864 had produced only occasional plant fossils, although anecdotal reports of ‘large lizards and snakes with feet' date back to 1858 (Bailey Reference Bailey and Hardman1881). Brownrigg communicated his findings to E. P. Wright of TCD, anticipating joint collaboration on the material. Wright then procured a grant from the British Association to fund the further collecting of plants, fishes, and additional non-amniote tetrapod specimens. Nonetheless, Wright simultaneously invited T. H. Huxley to describe the material (Etheridge Reference Etheridge1866; Wright & Huxley Reference Wright and Huxley1866; Huxley & Wright Reference Huxley and Wright1867). In doing so, Wright side-lined Brownrigg, who had expected to be involved in the scientific description of his finds and chose to collaborate with Huxley. Wright's behaviour caused resentment, conflict of interests, and overrode potential priority claims of local geologists (Wyse Jackson et al. Reference Wyse Jackson, DeArce and Monaghan2011). Brownrigg was sufficiently disaffected and aggrieved by the whole affair that he abandoned geology (DeArce et al. Reference DeArce, Monaghan and Wyse Jackson2011). He sold his personal collection to the British Museum (Natural History) in January 1870. The original register entry NHMUK PV OR 41851 notes that ‘92 blocks from the Jarrow coal containing fishes and amphibians and that many were subsequently destroyed because of pyrite decay'.
A total of c.120 identifiable tetrapods specimens are recorded in the collections of the institutions listed in the previous section.
1.2. Geology and preservation
The Jarrow Coal Formation is dated as Langsettian (Westphalian A) in the lower part of the communis zone (Eagar Reference Eagar and Nevill1961, Reference Eagar1964; Nevill Reference Nevill1961). Higgs & O'Conner (Reference Higgs and O'Connor2005) confirmed on palynological grounds that the Jarrow seam lies above the SS–RA miospore biozonal boundary that correlates closely with the boundary of the lenisulcata and communis non-marine bivalve chronozones. The Jarrow seam, 35cm deep, extends over 51.7km2 of the Leinster Coalfield, expanding into an oxbow 167–251m wide and 1.2m deep (Hallissy Reference Hallissy1939). The whole lake basin is U-shaped in cross section, 14m deep and represents an abandoned channel (Commission of Enquiry 1921; Hook & Ferm Reference Hook and Ferm1988). Unusually, the Jarrow channel is infilled with anthracite and cannel coal (Kendall Reference Kendall1923). The non-amniote tetrapods and fish fossils derive from bright coal and carbonaceous shales in millimetre laminae overlying the anthracite coal. The matrix is pyrite-rich and Jarrow specimens are poorly and variably preserved although usually as articulated individuals. The bone is subject to pyrite decay, mobilisation and replacement of calcium phosphate Ca3(PO4)2 by calcite and/or prismatic apatite crystals and sphalerite (Zn,Fe)S that collapses the bone structure (Rayner Reference Rayner1971). Extensive dissolution results in bituminous residues with indistinct outlines, as noted by Huxley & Wright (Reference Huxley and Wright1867, p. 354), and, ultimately, low-resolution ‘ghost' moulds. Rayner (Reference Rayner1971) concluded that the interstitial waters of the lake-bottom sludge were intermittently sufficiently acid to cause the diagenetic changes that degrade anatomical detail and hinder interpretation of the material. The usual technique for the preparation of coal fossils – acid-etching and casting the resulting high-fidelity moulds – is, thus, not applicable to the Jarrow material. Fine-surface films of cannel were removed where possible under a binocular microscope with an S. S. White industrial abrasive using sodium bicarbonate powder (Stucker Reference Stucker1961).
2. Systematic palaeontology
Order Nectridea Miall, Reference Miall1875
Emended diagnosis. Small Palaeozoic tetrapods with a relatively short trunk, between 14 and 26 presacral vertebrae, and a long, usually laterally compressed tail. No otic notch and no stapes known. Prefrontal contacts external naris in majority of taxa, palatal fangs absent. Intertemporal absent. Postorbital–parietal contact (except Scincosauridae) and parietal–tabular contact. Braincase largely cartilaginous but opisthotic long and well-ossified where known. Single coronoid and splenial in lower jaw. Cleithrum present primitively. Vertebrae holospondylous with single spool-shaped centra fused to neural and haemal arches. Atlas vertebra with reduced neural spine. Neural spines rectangular to fan-shaped, ornamented with longitudinal grooves and distal crenelations. Haemal spines expanded and bearing similar ornament. At least one pair of accessory articulations present above the zygapophyses between adjacent vertebrae in the trunk and proximal region of tail. Limbs relatively small, manus and pes with five digits where known.
Family Diplocaulidae Cope, Reference Cope1881
(=Ceraterpetontidae Jaekel, Reference Jaekel1903.
Keraterpetontidae Jaekel, emend. Romer, Reference Romer1945)
Emended diagnosis. Nectridean non-amniote tetrapods with broad moderately to strongly flattened akinetic skulls with cheek and skull table firmly sutured. Skull with short snout and long postorbital region. Nasal participates in the posteromedial border of the naris. Intertemporal and supratemporal absent. Frontals paired primitively. Large parietal contacting frontal, postfrontal, squamosal, tabular and postparietal. Tabular large with long posteriorly or posterolaterally directed horn. Squamosal with an internal flange contacting quadrate. Quadrate anterior to level of occiput. Sutural connection between parasphenoid, exoccipitals and basioccipital. Mandible short with short marginal tooth row, primitively with 10–12 teeth. Retroarticular process present on surangular. Single Meckelian fenestra in mandible. Dermal ornament a shallow pit-and-ridge system. Lateral-line canals present. Short trunk with a maximum of 17 presacral vertebrae. Neural arches of presacral vertebrae never waisted, haemal arch insertion always symmetrically waisted. Haemal spine articulation a simple overlap where present. Elongate rectangular to triangular clavicle plates. Cleithrum ‘T'-shaped primitively. Unossified carpals and tarsals.
Included taxa. Batrachiderpeton, Diceratosaurus, Diploceraspis, Diplocaulus, Ductilodon, Keraterpeton.
Genus Keraterpeton Huxley in Wright & Huxley, Reference Wright and Huxley1866
(=Urocordylus Miall, Reference Miall1875 non Wright & Huxley, Reference Wright and Huxley1866,
Ceraterpeton Cope, Reference Cope1875,
Keratespeton Oldham, Reference Oldham1876 in errore,
Ceraterpetum Lydekker, Reference Lydekker1890,
Keraterpetum Andrews, Reference Andrews1895)
Diagnosis. Primitive characters with respect to all other diplocaulids.
Maxilla excluded from orbit margin by lacrimal–jugal contact. Nasal contributes to posteromedial border of the naris. Squamosal margin concave, similar in area to parietal and extends onto the lateral tabular horn base. Postparietal border straight. Parasphenoid with long cultriform process contacting vomers. Ossified basibranchial present. Elongate dorsally directed retroarticular process on surangular. Seventeen presacral vertebrae. Dorsal neura1 spines twice as tall as wide. Dorsal edge of neural spines bear ridged, striate to slightly rugose ornament. Caudal vertebrae lack zygantrum–zygosphene articulations and neural and haemal spines slightly waisted. Haemal spines wider than centra and overlap along the tail. Haemal spines wider than centra. Long tapering tail with a whip-like end in which spine heights gradually reduce distally to small centra only. Simple crescentic humerus.
Primitive characters shared with Diceratosaurus.
Prefrontal reaches external naris margin. Cultriform process on parasphenoid and small interpterygoid vacuities present. Tabular horns straight and parallel. Cleithrum with short, rounded anterior process lying in close proximity to the posterior end of the tabular horn and long posterior process with straight dorsal margin. Paired accessory dermal ossifications lateral to clavicles. Distal caudal vertebrae with waisted haemal arches.
Primitive characters shared with Diceratosaurus and Batrachidepeton.
Postorbital contributes to posterior margin of orbit. Dermal ornament a pit-and-ridge system. Proximal caudal vertebrae with haemal arches inserted along full length of centrum and overlapping haemal spines.
Distribution. England, Republic of Ireland; Langsettian–Bolsovian, Pennsylvanian.
Included species. Type species Keraterpeton galvani, Keraterpeton longtoni.
2.1. Keraterpeton galvani Wright & Huxley, Reference Wright and Huxley1866
Diagnosis. As for genus plus postorbital with narrow insertion on orbit margin and elongate posterior contact inserting between parietal and squamosal. Parietal–squamosal suture convex. Transverse lateral-line sulcus follows squamosal parietal suture. Skull table slightly longer than pre-parietal region of skull. Three pairs of caudal ribs usually present.
Holotype. NMING F14735 (ex GSI B.2866). The largest known individual in dorsal aspect lacking the tail. Length 170mm as originally preserved (Fig. 1a). The proximal caudal vertebrae and hind limb has since been lost and the specimen is embedded in and covered by paraffin wax to combat pyrite decay (N. T. Monaghan, pers. comm.) (Fig. 1b). The species was named for the collector Charles Galvan of the GSI. Huxley proposed the name in November 1865, in reply to a letter from J. B. Jukes, Director of the GSI, returning a sketch sent to him by Galvan.
Locality. Jarrow Colliery, Castlecomer near Clough, County Kilkenny, Ireland.
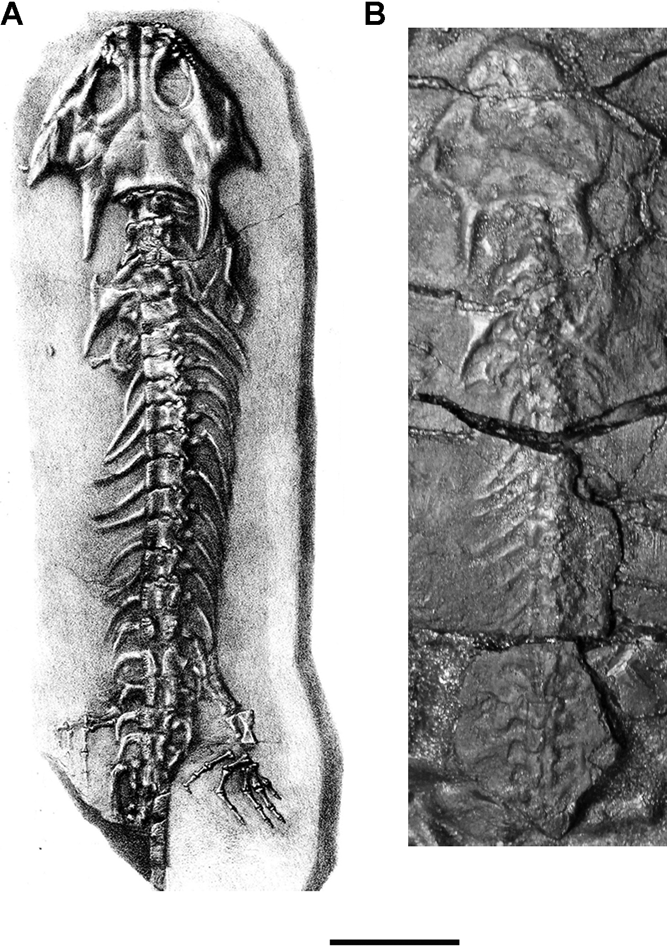
Figure 1 NMING F:14735 Keraterpeton galvani holotype skeleton in dorsal aspect. (A) Figured by Huxley & Wright (Reference Huxley and Wright1867, pl. 19, fig. 1). (B) Photograph as now preserved, showing missing hind limbs. Scale bar=30mm.
Horizon. Jarrow Coal, Coolbaun Coal Formation (Higgs & O'Conner Reference Higgs and O'Connor2005), Leinster Coalfield, Langsettian (=Westphalian A), Pennsylvanian.
Referred material. BGS 12107, presented by James George Robertson in 1889, originally to the Royal School of Mines. Ghost impression of skull, trunk and disarticulated hind limb elements in dorsal aspect.
BGS 26842 (ex a 6 XVII 2), presented by James George Robertson in 1889 (originally to the Royal School of Mines). Skull in dorsal aspect figured Steen (Reference Steen1938, figs 9c, 10) with associated but detached pectoral girdle.
CAMSM A.5969 (Walton Collection, 16w). Skull figured Steen (Reference Steen1938, pl. 2, fig. 3) trunk and proximal five caudal vertebrae in dorsal aspect. Length 115mm.
CAMSM E.14378 (Walton Collection, unnumbered). Ghost impression of skull and skeleton including proximal caudal vertebrae described Wilson (Reference Wilson1875), figured Oldham (Reference Oldham1876, pl. vii), cited Woods (Reference Woods1891). Originally in the Rugby School Natural History Collection. Length c.140mm.
CAMSM E.14379 presented by W. Clayton Browne in 1900. Skull in ventral aspect with complete vertebral column with complete tail. Length c.200mm.
CAMSM E.14380 (Walton Collection, 8w). Skull and trunk in dorsal and ventral aspect mounted in glass. Length c.70mm.
CAMSM E.19869 (Walton Collection, 17w). Skull and trunk in dorsal aspect plus five associated articulated posterior dorsal vertebrae. Length 74mm.
CAMSM E.19870 (Walton Collection, 2w). Ghost impression of skull and anterior postcranium in ventral aspect. Length c.70mm.
CAMSM E.19871 (Walton Collection, 3w). Fragmentary skull and vertebral column including c.23 caudal vertebrae. Length c.150mm.
CAMSM E.19872 (Walton Collection, 33w). Skull anterior trunk and pectoral girdle in ventral aspect. Length 61mm.
NHMUK PV R 8452, purchased from William Bookey Brownrigg, Reference Brownrigg1870. Skeleton in dorsal aspect complete to third caudal vertebra. Length 165mm.
NHMUK PV R 8457, purchased from William Bookey Brownrigg, Reference Brownrigg1870. Skeleton in ventral aspect including hind limb and proximal caudal vertebrae as impressions. Length 116mm.
NHMUK PV R 8460, purchased from William Bookey Brownrigg, Reference Brownrigg1870. Poorly preserved skull, trunk and proximal caudal vertebrae. Length 80mm.
NHMUK PV R 8472, purchased from William Bookey Brownrigg, Reference Brownrigg1870. Counterpart impression of NHMUK PV R 8460.
NMING F14711 (ex G1-1970), presented by Joseph Dobbs, 1890. Poorly preserved skull and trunk in ventral aspect. (No accurate measurements possible.)
NMING F14212 (ex G2-1970), presented by Joseph Dobbs, 1890. Poorly preserved skull and trunk in dorsal aspect. (No accurate measurements possible.)
NMING F14714 (ex G4-1970), presented by Joseph Dobbs, 1890. Poorly preserved ghost impression of skull fragments and trunk. (No accurate measurements possible.)
NMING F14715 (ex G5-1970), poorly preserved ghost impression. (No accurate measurements possible.)
NMING F14734 (ex GSI collection). (Not examined.)
NMING F14742 (ex GSI collection). (Not examined.)
NMING F16888. (Not examined.)
NMS 1950.56.6, well-preserved skeleton in ventral aspect originally with complete tail. Figured Woodward (Reference Woodward1897, pl. 12).
TCD F70 (ex NMING F14694, G47–1959). Skeleton in ventral aspect figured Huxley & Wright (Reference Huxley and Wright1867, pl. 19, fig. 2).
TCD R252 (ex NMING F14695, G48–1959, T252/426). Skeleton preserved in ventral aspect complete to 16th caudal vertebra. Length c.167mm.
TCD 38352 (ex NMING F14696, G49–1959). Skull in dorsal aspect, trunk and proximal five caudal vertebrae in left lateral aspect. Length 127mm.
WARMS Gz 2470. Fragmentary vertebral column and hind limb.
WARMS Gz 2780. Postcranial fragments on small blocks.
MCZ 2193, presented by E. P. Wright. Skeleton in ventral and lateral aspect, complete to 6th caudal vertebra. Length 155mm.
2.2. Keraterpeton longtoni Milner, Reference Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998
(=Urocordylus wandesfordii Wright & Huxley, Miall, Reference Miall1875 non Wright & Huxley, Reference Wright and Huxley1866;
Keraterpeton galvani Wright & Huxley, Ward, Reference Ward1890 non Wright & Huxley, Reference Wright and Huxley1866)
Diagnosis. As for genus plus postorbital squarish with a broad insertion on orbit margin and short parietal contact. Parietal–squamosal suture concave. Transverse lateral-line sulcus crosses squamosal. Cranial lateral-line sulci present as pronounced pits including transverse occipital and infraorbital pit-lines. Skull table slightly shorter than pre-parietal region of skull. Dorsal edges of neural spines with markedly coarsely rugose ornament.
Holotype. NHMUK PV R 2395 presented J. Ward (Reference Ward1890). The only certainly known specimen. Skull and partial postcranial skeleton preserved in dorsal aspect on three blocks (Fig. 2). Skull figured Ward (Reference Ward1890, pl. ix, fig. 2); Andrews (3, fig. 1); Steen (Reference Steen1938, text-fig. 9a, pl. 2, fig. 1); Milner (Reference Milner1978, figs 22–25 (unpublished thesis)).

Figure 2 NHMUK PV R 2395 Keraterpeton longtoni skeleton in dorsal aspect. (A) Photograph of two blocks comprising most of the skeleton. (B) Interpretative drawing of skull and anterior postcranium. (C) Composite outline of skull including anterior snout preserved on small counterslab partly as a mould. Abbreviations: ac = anterior commissure; AT = atlas; CLA = clavicle; CLE = cleithrum; DO = dorsal osteoderms; = ios = infraorbital sulcus; js = jugal sulcus; L = lacrimal; MAND = mandible; P = parietal; PP = postparietal; QJ = quadratojugal; SC = scapulocoracoid; sos = supraorbital sulcus; tos = transverse commissure; ts = temporal sulcus. Scale bars = 10mm.
Locality. Longton Hall Colliery, Longton, Staffordshire, England.
Horizon. Rowhurst Coal, known locally as the Ash Coal, Middle Coal Measures, Bolsovian (=Westphalian C), Moscovian, Pennsylvanian (Rees & Wilson Reference Rees and Wilson1998). Noted by Ward (in Gibson et al. Reference Gibson, Barrow, Wedd and Ward1905) as a bluish-black shale intercalated with a thin band of ironstone overlying the Ash Coal.
Possible related material.
Locality. Fenton Colliery, Fenton, Staffordshire, England.
Horizon. Winghay Coal, otherwise known as the Knowles Coal, from an ironstone band termed the ‘Knowles Ironstone', Middle Coal Measures, Bolsovian (=Westphalian C), Pennsylvanian (Rees & Wilson Reference Rees and Wilson1998).
Material. NHMUK PV R 8967 (acquisition history unknown). Four articulated dorsal vertebrae, two of which are shattered with an embedded impression of a large tooth indicating probable predation. Not diagnostic but may belong to Keraterpeton longtoni since the dorsal edges of neural spines bear identical coarsely rugose ornament.
3. Description
3.1. General proportions of the skeleton in Keraterpeton galvani
Almost complete skeletons are preserved in NMS 1950.56.6 and CAMSM E.14379. Woodward (Reference Woodward1897, pl. 12) figured the skeleton of NMS 1950.56.6 in ventral aspect with 49 or 50 caudal vertebrae, but only the proximal 25 now remain. The distal portion of the tail was lost during a subsequent remounting of the block in a wooden frame (pers. obs.). Based on Woodward's figure, the proportions of the skull:trunk:tail are 1:3.5:7, the entire skeleton being about 170mm long (Fig. 3). The articulated tail of an otherwise poorly preserved 200-mm-long individual, CAMSM E. 14379, includes c.53 caudal vertebrae. Accurate overall dimensions are not possible for most specimens because of the variable effects of preservation and crushing, but the ratio of skull length to trunk length falls within the range of 1:3.0–3.5. Midline skull lengths range from c.20 to 40mm in Keraterpeton galvani, giving an estimated total length ranging from c.l70mm to 400–420mm.

Figure 3 NMS 1950.56.6 Keraterpeton galvani skeleton in ventral aspect. Distal region of tail restored from Woodward (Reference Woodward1897, pl. 12). Scale bar=20mm.
3.2. Skull
3.2.1. Skull roof
The holotype NMING F14735 is the largest recorded individual with a skull length of c.40mm (Fig. 1). Skull lengths in other individuals range between 20 and 29mm. The major features of the skull roof are depicted in Huxley & Wright's original figure (Reference Huxley and Wright1867, pl. 19, fig. 1), but the specimen is unavailable for detailed study (see Section 2.1). The skull roof in dorsal aspect is broadly triangular in shape, with a rounded snout. The posterolateral borders of the skull table bear parallel straight-sided slender and posteriorly directed tabular horns. The skull roof description is based principally on BGS 26842 (Fig. 4) with a skull length of c.28.5mm unless otherwise indicated. The snout region is broad, shallow and squared off anteriorly with sub-terminal external nares. The orbits are large, slightly oval in shape and dorsolaterally oriented. The deep cheek region is triangular in shape and describes a shallow curve with the margin of the skull table. It describes an almost 90° concave curve to the lateral margin of the tabular, which it meets on a line slightly medial to the lateral snout margin. The cheek region is preserved flattened in the same plane as the skull table in the majority of specimens. In some individuals it is fractured along the mid-length of the orbit – the line of least resistance in the skull roof – and crushed flat against the palate.

Figure 4 BGS 26842 Keraterpeton galvani skull in dorsal aspect. (A) Photograph. (B) Interpretative drawing. (C) Reconstruction of skull in dorsal aspect. (D) Reconstruction of skull in left lateral aspect. Abbreviations: A = articular; AN = angular; D = dentary; FR = frontal; J = jugal; L = lacrimal; mg = Meckelian groove; MX = maxilla; N = nasal; P = parietal; PMX = premaxilla; POF = postfrontal; PP = postparietal; PRF = prefrontal; Q = quadrate; QJ = quadratojugal; RT = retroarticular process; SA = surangular; SQ = squamosal; T = tabular. Scale bars = 10mm.
The dermal ornament of the skull roof is a uniformly distributed pit-and-ridge system. The centre of ossification of each bone is marked by deep pitting that radiates out as elongated ridges along the lines of growth.
The right premaxilla preserves four conical marginal teeth (see Section 3.2.3), is rectangular in shape and the posterolateral margin borders the external naris. Steen (Reference Steen1938, fig. 9c) depicted the premaxilla as the same length as the nasal, but it is only half as long in the midline. The maxilla is slender and splint-like and the marginal teeth are sharply conical; five anterior ones plus a gap are present on the left side (see Section 3.2.5). The right maxilla is slightly displaced at its anterior end exposing the broad overlapping suture with the premaxilla on the lateral margin of the external naris. The maxilla is excluded from the orbit margin by the lacrimal and jugal.
The nasal is short and broad anteriorly where it borders the medial narial margin and narrow posteriorly. The frontal is long and narrow with parallel lateral and medial margins and a length–width ratio of 3:1.
The parietal is the largest element in skull roof with convex lateral margins so that the posterior width is 2.5 times the anterior width in contrast to Steen's (Reference Steen1938, figs 9b, 10) interpretation of the parietals as comparatively parallel-sided. The parietal contacts the frontal and postfrontal anteriorly and forms the posterolateral margin of the postorbital. The contact with the squamosal posterolaterally and the postparietal and tabular posteriorly effectively preclude any kinesis between the skull table and cheek. The postparietal is rectangular in shape with a straight posterior margin and a smooth convex ridge curving ventrally towards the occiput. The dorsal aspect of the exoccipitals are exposed beyond posterior margin of the postparietals and tabulars, although not well preserved.
The prefrontal is long and slender and contacts the posterior narial margin in CAMSM A.5969 (Fig. 5). The postfrontal borders the posteromedial margin of the orbit and contacts the prefrontal, frontal, parietal and postorbital (Figs 4, 5). The postorbital is elongate with a narrow neck inserting on the posterior orbit margin contra Steen's figure (Reference Steen1938, figs 9c, 10) of a wide insertion occupying most of the posterolateral orbit margin. A long posterior process carrying the temporal sulcus forms a v-shaped contact between the parietal and squamosal.
The slender anterior process of the jugal borders the lateral orbit margin and contacts the postorbital medially. The right jugal is displaced laterally in CAMSM A.5969 (Fig. 5), exposing a long overlapping contact with the lacrimal. It has a long lateral-sloping suture overlying the maxilla and forms the posterior margin of the mouth. It expands posterolaterally, with a concave margin at the junction of the snout and the flared-out cheek contacting the quadratojugal at the lateral extremity of the cheek and the squamosal posteriorly. The quadratojugal occupies the ventrolateral extremity of the cheek region. The posterior margins form a concave notch with the squamosal contact to accommodate the movement of the mandibular retroarticular process.

Figure 5 CAMSM A.5969 Keraterpeton galvani skull interpretive drawing in dorsal aspect with right mandible displacing anterior end of maxilla. Abbreviations: ext. n. = external naris; J = jugal; L = lacrimal; MAND = mandible; MX = maxilla; N = nasal; PMX = premaxilla. Scale bar = 10mm.
The squamosal is very broad anteriorly and comprises an extensive part of the cheek region contacting the jugal, quadratojugal, parietal, postorbital and jugal. The lateral margin is acutely concave from the quadratojugal notch, forming a splint-like contact with the distal margin of the tabular horn base. The tabular contacts the parietal anteriorly, the postparietal medially so that the posterior margin describes a smooth concave curve with the postparietal margin of the skull. The anterior part of the tabular extends posteriorly to form the straight parallel-sided horn. The lateral margin of the horn is straight and continues anteriorly to contact the squamosal process. Dermal ornament is restricted to the dorsal surface and the posteromedial surface is relatively smooth and lightly striated.
3.2.2. Lateral-line system
Traces of the lateral-line system are preserved as a combination of pit-lines and grooves. Traces of the left and right supraorbital sulci follow the course of the sutures between the frontals and pre- and postfrontals, and traverse laterally across the postorbitals and the jugals, intersecting with anterior end of the temporal sulcus at the mid-point of the postorbitals. The temporal sulcus runs posteriorly, following the line of the parietal squamosal suture. The anterior ends of the temporal sulci run posteriorly across the postorbitals from the points of intersection with the supraorbital sulci in BGS 2684 (Fig. 4).
3.2.3. Palate
Few specimens are preserved in ventral aspect and none show a complete undistorted palate, particularly the braincase and occipital regions. The palatal elements are extensively crushed, overlain by the folded right-cheek region and mandible in TCD R252 (Fig. 6) or the pectoral girdle plates in MCZ 2193 (Fig. 7). The features of the palate are described as viewed, rather than reversed with respect to the dorsal aspect of the skull.

Figure 6 TCD R252 Keraterpeton galvani skull in ventral aspect. The dorsal aspect of the right cheek and mandible, broken along the lateral orbit margin is crushed over, obscuring the midline region of the palate. (A) Photograph. (B) Interpretative drawing of identifiable structures. Abbreviations: A = articular; AN = angular; D = dentary; J = jugal; LPT = left pterygoid; MX = maxilla; PAL = palatine; PMX = premaxilla; PT = ptergoid; Q = quadrate; RSQ = right squamosal = SA = surangular; SPL = splenial; SQ = squamosal; T = tabular; V = vomer. Scale bar = 10mm.

Figure 7 MCZ 2193 Keraterpeton galvani photograph of skull in ventral aspect. Abbreviations: ADO = accessory dermal ossification; B = ossified basibranchial; BOC = basioccipital; b.pr = basipterygoid process; CL = clavicle; EO = exoccipital; EOC = exoccipital condyle; ICL = interclavicle; ipt.v = interpterygoid vacuity; OP = opisthotic; PAL = palatine; PMX = premaxilla; PSP = parasphenoid; PT = pterygoid; Q = quadrate; QJ = quadratojugal; R.SQ = right squamosal; SQ = squamosal; T = tabular; V = vomer. Scale bar = 10mm.
The left maxilla, premaxilla, internal nostril, palatine and vomer are preserved in TCD R252 (Fig. 6), although the sutures between the elements are unclear. The premaxillary tooth count is between four and eight, but cannot be determined accurately as only a small trace of the probable midline suture is visible in line with a trace of the midline vomer suture.
The anterior region of the left premaxilla bears a ring of seven or eight small teeth round the rim of the internal nostril margin and two larger ones placed anterolateral to it. The bone preservation of the posterior region is badly degraded and lacks the finished bone surface, although the posterior end of the suture with the right vomer is visible. The left palatine lies between the maxilla and jugal laterally and vomer medially. It carries a row of 16 conical teeth (allowing for two gaps) that diminish in size anteriorly towards the premaxillary contact. The palatine does not appear to contact the posterior narial margin but preservation is not sufficiently clear.
The palatal ramus of the right pterygoid borders the lateral margin of a small interpterygoid vacuity in MCZ 2193 (Fig. 7), establishing that the palate was not closed, contra Bossy & Milner (Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998). The anterior region is crushed and incomplete, revealing the ventral side of the skull roof and border of the orbit. A medial ridge is preserved, running anteriorly parallel to the cultriform process of the parasphenoid and contacts the vomer, although the suture is not clear. The posterior region of the palatal ramus bears fine diagonal parallel striations radiating towards the medial margin, marking scars for the insertion of the adductor musculature. No denticle-covered area is present, contra Milner (Reference Milner and Panchen1980a) and Bossy & Milner (Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998). The medial margin has a vertical contact with the parasphenoid at the posterior end of the interpterygoid vacuity (see Section 3.2.4). The quadrate ramus appears to have been sheared off distally and a large tongue-like process of smooth bone protrudes posterolaterally. This is interpreted as the distal part of the ramus that has been compressed and rotated to lie in the same plane as the palatal ramus. It overlies most of the quadrate except for the exposed condyles that are separated by a deep intercondylar groove. The underlying margin of the quadratojugal is visible, marked by a distinct crack that continues anterodistally onto the ventral exposure of the squamosal. The right quadrate ramus is partially exposed in TCD R252 (Fig. 6), oriented ventrolaterally and contacts the quadrate with a strongly interdigitating suture. The medial quadrate condyle is still in articulation with the articular glenoid (see Section 3.2.5). A small area of the internal shelf of the squamosal that braces the quadrate is exposed in ventral aspect. A reconstruction of the skull in palatal aspect with the pterygoid and quadrate restored in life position is based on TCD R252 and MCZ 2193 (Fig. 8).

Figure 8 Reconstruction of the palate of Keraterpeton galvani based on Figures 6 and 7. Abbreviations: BOC = basioccipital; b.pr = basipterygoid process; EO = exoccipital; EOC = exoccipital condyle; ipt.v = interpterygoid vacuity; J = jugal; MX = maxilla; OP = opisthotic; PAL = palatine; PMX = premaxilla; PSP = parasphenoid; PT = pterygoid; Q = quadrate; QJ = quadratojugal; SQ = squamosal; T = tabular; V = vomer. Scale bar = 10mm.
3.2.4. Braincase and occiput
The parasphenoid is preserved in MCZ 2193 (Fig. 7). It bears a long parallel-sided cultriform process with a square-ended anterior contact with the vomers in the midline. The left basipterygoid process is preserved in situ on the posterolateral region of the cultriform process. The right process is compressed against and articulated with the pterygoid. The parasphenoid baseplate is diamond-shaped and sutures with the posteromedial margin of the right pterygoid ramus. The posterior end of the plate is v-shaped and the posterolateral margin shows a long robust interdigitating suture with the anterior margin of the left exoccipital and a midline suture with a poorly preserved basioccipital.
The left occipital condyle is borne on a stout pedestal and the condyle is oriented slightly towards the midline. The contact with the tabular is disrupted by a broken suture and missing segment of bone. A fractured contact parallel and anterior to the tabular with a slightly detached bone fragment is interpreted as a probable paroccipital process of the left opisthotic (Fig. 8). A fragment of the right exoccipital plate is visible together with the oval condyle, although the condylar surface is partly eroded.
3.2.5. Mandible
The mandible is straight, sub-parallel, narrowing anteriorly and curving medially towards the symphysis. The lateral surface ornament is identical to that of the skull roof. The mandibles are preserved in articulation in several individuals and the long dorsally oriented surangular retroarticular process projects well behind the posterior margin of the cheek. The dentary bears a single row of sharply pointed, conical marginal teeth. A complete tooth row is not preserved in any individual, but, assuming equal spacing, there appears to be space for 10–12 teeth. The posterior end of the left dentary tooth row is exposed anterior to the edentulous dorsal margin in TCD R252 (Fig. 6). Two teeth occupy the same socket, indicating possible evidence of tooth replacement previously unreported in diplocaulids. The mesial surface of the dentary is visible in the partially disarticulated mandibles of BGS 26842 (Fig. 4), exposing a shallow widening Meckelian groove and an elongate surangular dorsal crest. The left mandible in TCD R252 (Fig. 6) is preserved in lateral aspect. The splenial has a wide exposure on the mandibular symphysial margin and a long sloping contact with the angular. The angular has a long straight dorsal contact with the dentary and a sloping contact posteriorly with the surangular. The anterodorsal process of the surangular is slightly disarticulated, showing the long overlapping contact with the posterior region of the dentary. The glenoid surface of the articular bears prominent rounded facets either side of a deep u-shaped asymmetric embayment accommodating the quadrate condyles. The course of the suture with the surangular is not determinable.
3.2.6. Branchial skeleton
A single ossified basibranchial element in ventral aspect is present in MCZ 2913, located ventral to the parasphenoid (Fig. 9). A basibranchial is known in some basal tetrapods (Witzmann Reference Witzmann2013) but is hitherto unrecorded in any nectridean contra Bossy & Milner (Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998). It is a robust rod-like element with a rounded expanded anterior head resembling a roller joint. The head bears deep laterally curving grooves, the left being somewhat compressed. The shaft widens gradually and then expands markedly with a roughened and lightly rugose surface to the point where it is overlain by the left clavicle plate. Two ridges run anteroposteriorly from the base of the head offset from the midline, between which runs a shallow groove terminating in faint radiating ridges. The morphology bears little resemblance to the basibranchials of basal tetrapods described and figured by Witzmann (Reference Witzmann2013) other than the overall basic shape of a rod-like shaft with expanded anterior and posterior ends and a ventral groove as described in dvinosauroid temnospondyls.

Figure 9 MCZ 2193 Keraterpeton galvani interpretative drawing of ossified basibranchial from Figure 6. Scale bar=5mm.
3.3. Postcranial skeleton
3.3.1. Vertebrae and ribs
The vertebral column comprises 17 presacral vertebrae (NMING F14735; Fig. 1). Regional variation in the column is negligible, the depth and length of the vertebrae are almost constant with a slight reduction in spine height in the pelvic region (CAMSM A.5969; Fig. 10). The neural arches and spines of the dorsal vertebrae are fused along the full length of the centrum so that adjacent elements are firmly in contact throughout their full height. The neural spines are ornamented with light striations and pitting with thickened ridged dorsal margins bearing prominent indented vertical striations. Transverse processes are positioned anterodorsally on the neural arches adjacent to the anterior zygapophyses. The postzygapophyses are distinctly larger and overlap the prezygapophyses on each succeeding vertebra and the articulating surfaces between the zygopophyses are horizontal or nearly so. Zygantrum–zygosphene articulations are present above the zygapophyses. The posterodorsal margin of each neural arch is developed into a distinct spine, which fits into a corresponding notch on the anterodorsal neural arch margin of the succeeding vertebra. The adjacent edges of the neural spines show additional small complementary indentations above the zygantrum–zygosphene (Fig. 10). The horizontal position of the zygapophyses and the extra articulations lock the vertebrae together, minimising dorsoventral flexing but allowing lateral movement between the vertebrae. The caudal neural arches and spines are slightly waisted on the proximal centra and the spines abut dorsally between adjacent vertebrae. Haemal arches fused ventrally to the centra are carried along the length of the tail. The haemal spines span the full length of the centra and are tightly appressed in the first four post-caudal rib-bearers. They have slightly waisted insertions on the centra from the fifth caudal vertebra distalwards. The spines are expanded and fan-shaped so that the posterior margins are longer than the centra so that they overlap the spines of the succeeding vertebrae. The overlapping generally follows a consistent sequence, with the anterior end of each spine placed to the same side of the posterior end of the preceding spine to facilitate lateral flexibility. The zygapophyses are dorsally placed and rather crescent-shaped. The prezygapophyses project anteriorly from a pronounced lateral ridge on the neural arch. The postzygapophyses curve dorsolaterally from the lateral ridge and dorsoventrally at its posterior extremity and almost completely overlap the prezygapophyses (Fig. 11). The haemal arches bear no articulations. Both neural and haemal spines gradually decrease in height distally and become more waisted so that the tail gradually tapers. Traces of low spines persist at least as far as the 22nd caudal vertebra and the distalmost elements are reduced to notochordal rings forming a whip-like end to the tail (Fig. 3).

Figure 10 CAMSM A.5969 Keraterpeton galvani skeleton. (A) Skull in dorsal aspect with vertebral column in left lateral aspect. (B) Interpretative drawings of proximal presacrals 5 and 6. (C) Distal presacrals 13 and 14. Abbreviations: CAP = capitulum; NA = neural arch; TP = transverse process; TUB – tuberculum; za = prezygapophysis; zp = postzygapophysis; zyga = zygantrum; zygp = zygosphene. Scale bars = 10mm (A); 5mm (B), (C).

Figure 11 CAMSM E.19871 Keraterpeton galvani interpretative drawing of mid-caudal vertebrae in left lateral aspect. Abbreviations: HA = haemal arch; HS = haemal spine; NA = neural arch; NS = neural spine; za = prezygapophysis; zp = postzygapophysis. Scale bar = 5mm.
Robust curved dorsal ribs are borne by all the post-axial presacral vertebrae gradually becoming shorter and more incurved (Fig. 12). The sacral rib is stout and straight with posteriorly reflexed distal end in NMING G1-1970 (Fig. 13). The sacral vertebra bears a short stout rib and the first three caudals carry sharply reflexed caudal ribs. Two caudal rib-bearers are apparent in NMS 1950.56.6 (Fig. 2), but other individuals, including the holotype NMING F:14735 (Fig. 1), possess three pairs. The ribs are double-headed with a short tuberculum articulating with the transverse process and a longer capitulum that articulates obliquely either intercentrally or possibly on the posterior region of the preceding centrum, although no articular facets are preserved (Fig. 10).

Figure 12 NHMUK PV R 8452 Keraterpeton galvani skeleton in dorsal aspect. (A) Photograph. (B) Interpretive drawing. Abbreviations: CLA = clavicle; CLEI = cleithrum; CR = caudal rib; HS = haemal arch; SR = sacral rib. Scale bar = 10mm.

Figure 13 NMING F14711 Keraterpeton galvani interpretative drawing of sacral region. Abbreviations: CR = caudal rib; IL = ilium; SR = sacral rib; SV = sacral vertebra. Scale bar = 10mm.
3.3.2. Pectoral girdle and forelimb
The dermal girdle comprises paired cleithra, clavicles and a single median interclavicle. The clavicles and interclavicle overlap in many individuals due to post-mortem distortion. The articulated dermal plates are preserved in ventral aspect in BGS 26842 (Fig. 14). The broad rectangular clavicle plate bears heavily ornamented pits and ridges radiating out from the centre of ossification adjacent to the base of the unornamented dorsal process. The dorsal process arises from the distal margin of the plate slightly more than halfway towards the posterior end. In articulated specimens it slopes posteriorly (Figs 15, 16) to contact the cleithrum. The similarly ornamented interclavicle bears symmetrical depressions for reception of the clavicles. The posterior margin, where completely preserved, is broadly convex. Paired accessory dermal ossifications lateral to the clavicle plates and bearing similar ornament are present in TCD R252 and figured in MCZ 2913 (Fig. 7), NMING F14696 (Fig. 15) and NHMUK PV R 8457 (see Section 3.5). They are rather oval to triangular in shape with a pointed dorsal apex and have not been observed previously in Keraterpeton, although they are present in Diceratosaurus brevirostris from the Late Pennsylvanian assemblage at Linton, Ohio (Jaekel Reference Jaekel1903, pl. 5 and pers. obs.).

Figure 14 BGS 26842 Keraterpeton galvani interpretative drawing of pectoral girdle and associated forelimb elements in ventral aspect. Abbreviations: CLA = clavicle; DO = dorsal scale; H = humerus; ICL = interclavicle; R = rib; RA = radius; SC = scapulocoracoid; U = ulna. Scale bar = 10mm.

Figure 15 NMING F14696 Keraterpeton galvani photograph of skull and anterior region of trunk in left lateral aspect. Abbreviations: ADO = accessory dermal ossification; CLA = clavicle; CLEI = cleithrum; ICL = interclavicle; SC = scapulocoracoid. Scale bar = 10mm.

Figure 16 TCD R252 Keraterpeton galvani interpretative drawing of right forelimb in medial and ventral aspects. Abbreviations: CL = clavicle; CLEI = cleithrum; COR = coracoid; end = endepicondyle; ent = entepicondyle; gl = glenoid; H = humerus; RA = radius; SC = scapula; supc.f = supracoracoid foramen; U = ulna. Scale bar = 5mm.
The cleithrum is ‘T'-shaped with a long shaft inclined anteriorly from the perpendicular. The anterior head is short and broadly rounded and is projected posteriorly into a long tapering posterior spur with a straight dorsal margin. The anterior margin of the shaft follows a smooth convex concave to a sharply rounded ventral point. The posterior margin follows a convex curve to intersect with the base of the posterior arm of the head. The lateral face of the cleithrum is lightly ornamented. The medial face is smooth and the anterior head bears a distinct shallow notch in TCD R252 (Fig. 16). The cleithrum shaft lies parallel to the clavicle shaft and the head overlaps its distal margin (NMING F14696; Fig. 13). The posterior tip of the tabular horn lies very close to the anterior process and may have abutted against or fitted into the medial face of the notch. The skull is skewed laterally post-mortem in several individuals including NMING F 14696 (Fig. 15), suggesting a lack of any fixed osteological connection, although presumably there was a connective tissue attachment in life.
The right scapulocoracoid is unusually well preserved in left lateral aspect in TCD R252, partially overlain by the cleithrum shaft, although vertical cracks across the shaft reveal the broadly l-shaped outline (Fig. 16). The margins of the dorsal scapular region are almost parallel. The medial margin turns through a 90° curve demarcating the coracoid region. The coracoid is quite robust with a thick but eroded medial edge. A distinctly raised ridge on the posterior margin demarcates the elongate oval glenoid fossa. The supracoracoid foramen is marked by a distinct pit on the dorsolateral margin of the ridge.
The right humerus is preserved in ventral aspect in TCD R252 (Fig. 16), is 7mm long and kidney-shaped with no discernible features or foramina. The articular surfaces are distally placed and the radius and ulna lie in articulation with the entepicondyle.
The radius is shorter than the ulna with a robust shaft and rounded distal condyles. The ulna has a slender shaft and an incompletely preserved flattened asymmetric distal region in TCD R252 (Fig. 16). The radius and ulna are associated with the pectoral girdle in BGS 24682 (Fig. 14) and the proximal end of the ulna bears a small olecranon process.
The carpals are unossified. The digit count is ambiguous in most specimens, but five metacarpals are preserved in articulation in MCZ 2193 (Fig. 17) together with some phalanges of digits IV and V. A typed note on the reverse of the accompanying label states, ‘right manus damaged in preparation; original ?-?-?-4–3', which refers to an excavated area between the ulna and radius and the metacarpals. A five-digit count is also preserved in NMS 1950.56.6 (Fig. 2), contra the four-digit manus given in the diagnosis of the Diplocaulidae by Milner (Reference Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998, p. 122). The phalanges are slender with short curved unguals. Digit four is the longest and the phalangeal formula is probably 2–3–3–4–3.

Figure 17 MCZ 2193 Keraterpeton galvani interpretative drawing right of left forelimb in ventral aspect. Abbreviations: H = humerus; RA = radius; T = tabular; U = ulna; I = digit one; II = digit two; III = digit three; IV = digit four; V = digit five. Scale bar = 5mm.
3.3.3. Pelvic girdle and hind limb
The dorsally directed iliac blade is rectangular and square-ended in ventral aspect, and the posterior margin describes a convex curve widening into the baseplate in NMING F14696 (Fig. 13), although it is partly obscured by the right sacral and proximal caudal rib. The most complete hind limb is preserved in ventral aspect in TCD R252 (Fig. 18). The femur is straight with a slender shaft, and the distal end is c.40% wider than the proximal end. The tibia is robust and slightly curved with a wide proximal end. The shaft narrows distally but is incomplete. The fibula is c.60% shorter than the tibia with a narrow proximal and distal ends and the shaft curves markedly towards the tibia. The tarsus is unossified. The metatarsals are slender shafted with broad proximal ends and narrower distal ends. The phalanges are relatively long and slender with short curved unguals. Although no complete articulated pes is preserved, the phalangeal formula is probably 2–3–4–3–3.

Figure 18 TCD R252 Keraterpeton galvani interpretative drawing of hind limb. Abbreviations: CR = caudal rib; F = femur; FB = fibula; SR = sacral rib; SV = sacral vertebra; TB = tibia. Scale bar = 10mm.
3.3.4. Scalation
Isolated dorsal osteoderms overly the right scapulocoracoid and a rib in Keraterpeton galvani BGS 26842 (Fig. 14). They exhibit a pit-and-ridge ornament identical to the skull roof and pectoral girdle plates and the shape varies from roughly oval to quadrangular. Overlapping rows of small ventral scales between the pectoral and pelvic girdles were figured in TCD F70 by Huxley & Wright (Reference Huxley and Wright1867, pl. 19, figs 2–4). The scales were illustrated, albeit rather indistinctly, with crenelated edges. Ventral gastral scales are preserved clearly in NHMUK PV R 8457, covering the right ribs and presacral centra (Fig. 19). The individual scales are rectangular and arranged in overlapping parallel diagonal rows. They are heavily ornamented mirroring the pits and ridges that extend to posterior margins of the scales so that the edges are coarsely denticulate. Paired triangular-shaped accessory dermal plates are also preserved in association with the lateral margins of the clavicles (see also Section 3.4.2).

Figure 19 NHMUK PV R 8457 Keraterpeton galvani skeleton in ventral aspect with gastral scales overlying right ribs and presacral centra. Inset: close up of scales overlying right ribs. Abbreviations: ADO = accessory dermal ossification; CL = clavicle; CLEI = cleithrum; GS = gastral scales. Scale bar = 10mm.
3.4. Keraterpeton longtoni
Keraterpeton longtoni is represented by a single small individual with a midline skull length of c.15mm (NHMUK PV R 2395; Fig. 2). The bone preservation from the Ash Coal shale is complete with no loss of detail, in contrast with acid degradation of bone in the Jarrow Coal (Section 1.2). The dermal pitting on the skull roof is uniformly deeper and more densely distributed, and runs in distinct furrows to the margins of the midline elements (Fig. 2b) in comparison with Keraterpeton galvani (Fig. 4). The parietal is narrower with a concave lateral margin (Fig. 2b) in comparison with the convex margin in K. galvani (Fig. 4b). The skull roof is split between counterpart slabs that, between them, reveal more of the skull roof and lateral-line system. The postorbital is short and square (Fig. 2c) with a small posteromedial parietal contact in contrast to the elongate ‘v-shaped' parietal–squamosal contact in K. galvani (Fig. 4b). The lacrimal borders the posterior margin of the external naris and the anterior margin of the orbit and sutures narrowly with the jugal on the lateral orbit margin. A more complete lateral-line complement is present as deep pit-lines (Fig. 2b, c). The right infraorbital sulcus and transverse commissure crosses the tabulars and squamosals and the temporal sulci traverse the squamosals lateral to the parietals, in contrast to K. galvani (Fig. 4b) and continue posteriorly to intersect with the transverse sulcus that crosses the postparietals and tabulars.
The atlas vertebra, exposed clearly in right lateral aspect (Fig. 2b), lacks a transverse process on the neural arch and has double zygantrum–zygosphene articulations with succeeding second dorsal vertebra. This detail is not visible in any of the less well-preserved Keraterpeton galvani specimens. Several ‘scales' are thinly scattered across the anterior trunk region in Keraterpeton longtoni (Fig. 2b). The shapes vary between rectangular and oval, with pitted ornament and crenelated borders. These are interpreted as probable dorsal osteoderms based on the presence of pitted dermal ornament.
4. Discussion
The diplocaulids are characterised by broad, short-snouted akinetic skulls with large tabular bones bearing long posteriorly or posterolaterally directed horns, short trunks and long tails. They possess broad elongate pectoral girdle plates and unossified carpals and tarsals, adaptations seen in other secondarily aquatic groups of Carboniferous tetrapods (temnospondyls and anthracosaurs) and analogously with modern newts and salamanders (Milner Reference Milner and Panchen1980a). The diagnostic characters of the group express the unique morphology of highly specialised predatory aquatic animals. Keraterpeton is the earliest diplocaulid represented in the fossil record and the revised generic diagnostic characters in Section 2 represent the primitive condition with respect to all other diplocaulids. The presence of an ossified basibranchial element, although preserved in a single individual of Keraterpeton galvani only, and the primitive crescentic humerus are previously undiscovered basal diplocaulid characters. Milner (Reference Milner and Panchen1980a) discussed the jaw mechanics and locomotor adaptations in relation to the skull–pectoral girdle association in Keraterpeton and argued that it functioned in a dual role as a damping mechanism. Minimising lateral head oscillation to support a straight-line approach to prey as a consequence of subcarangiform swimming and buffering stress and skull tilting on rapid jaw opening of the depressor musculature were both mediated by the tabular–cleithrum connection.
The primitive skull–girdle association is shared with Diceratosaurus from the Pennsylvanian swamp assemblage at Linton, Ohio. Both genera also share paired extra dermal ossifications lateral to the clavicle plates that conferred extra rigidity to the pectoral girdle and anterior trunk. The primary function in Diceratosaurus was hypothesised to buffer the anterior trunk against hind limb paddling action rather than the subcarangiform swimming mode in Keraterpeton (Milner Reference Milner and Panchen1980a; Bossy & Milner Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998). The skull–girdle connection was lost in all other diplocaulids in relation to posterolateral tabular horn divergence and adaptations to different locomotor patterns and feeding strategies (Bossy & Milner Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998).
The size range of Keraterpeton galvani at Jarrow is restricted to individuals with a midline skull length of c.20–40mm. The size range for the genus is increased to c.15–40mm with the addition of the single specimen of Keraterpeton longtoni from the later Longton locality. Very small diplocaulids are rare in the fossil record. Diplocaulus primus from the Artinskian of Oklahoma is the smallest known individual with a midline skull length of 7.1mm. Despite being a juvenile, it exhibits a high degree of precocial ossification, lacking only ossified carpals and tarsals associated with an aquatic lifestyle (Milner Reference Milner1996). A Diplocaulus sp. with a midline skull length of 8.3mm, known from the Texas Early Permian, is also a juvenile but is a well-ossified individual (Chaney et al. Reference Chaney, Sues, DiMichele, Lucas and Zeiger2005). In a growth and variation study of Diplocaulus, Olson (Reference Olson1970) established a ratio of midline skull length to antorbital length of <5 as diagnostic of Diplocaulus magnicornis. However, he observed values of >5 for some skulls less than 24mm long and Milner (Reference Milner1996) questioned the diagnostic value of this ratio in small individuals. Nonetheless, the wide size range of D. magnicornis skulls, 8.3–147mm, establishes the possible growth range in diplocaulids. Batrachiderpton reticulatum from the Duckmantian coal-swamp assemblage in the Northumberland coalfield ,and with a similar aquatic active predatory lifestyle, is represented by specimens with a midline skull length of between 16 and 98mm, including well-ossified juveniles (Milner Reference Milner1978). Precocial ossification of the postcranial skeleton at small body size is a developmental pattern that occurs in both fossil and modern groups with holospondylous vertebrae, including nectrideans (Carroll Reference Carroll1989), and is suggestive of either direct development or a tiny larval stage of which there is no fossil record. The size range of K. galvani from Jarrow might represent a component of a single faunal mass death event. However, the variable state of preservation of individual specimens, combined with the fact that there are no records to indicate if the material came from a single horizon within the Jarrow channel, offers no clues. All that can be surmised is that individuals of between c.l70 and 420mm long inhabited the shallow oxbow lake that gave rise to the Jarrow Channel deposit and died either in a single or multiple events along with the other aquatic members of the fauna and a few terrestrial erratics. Most of the aquatic individuals are represented by articulated skeletons, suggesting rapid burial in quiet, perhaps anoxic, conditions.
A character analysis of the Nectridea conducted by Milner (Reference Milner and Panchen1980a) recovered a monophyletic clade supported by characters of the membrane bone vertebrae but not by cranial characters, and a monophyletic urocordylid clade as outgroup to Scincosaurus as the sister taxon to the diplocaulids. The diplocaulid taxa formed a nested set with Keraterpeton as the most primitive taxon. The remaining taxa were structured as Batrachiderpeton, Diceratosaurus, Peronedon, Diploceraspis, Diplocaulus. Characters shared by Batrachiderpeton and Diceratosaurus, the maxilla bordering orbit margin separating the lacrimal and jugal and emarginated midline postparietal borders, were cited as derived with respect to Keraterpeton. However, the emargination of the postparietal borders in Batrachiderpeton is an apomorphic feature related to the presence of postparietal horns (Bossy & Milner Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998, figs 57b, 58b). It is a very variable feature in Diceratosaurus (Bossy & Milner Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998, fig. 57c and pers. obs.) and not a reliable character state for this taxon. Peronedon primus Olson, Reference Olson1970 was redescribed and reduced in synonymy to Diplocaulus primus (Olson) by Milner (Reference Milner1996) and falls within the generic operational taxonomic unit (OTU) for Diplocaulus.
Relatively few cladistic analyses of early tetrapods have incorporated data on nectrideans. A detailed morphological review of the Moscovian terrestrial scincosaurid nectridean Scincosaurus by Milner & Ruta (Reference Milner and Ruta2009) included a critical review of large-scale investigations of early tetrapods by Anderson (Reference Anderson2001, Reference Anderson, Anderson and Sues2007), Ruta et al. (Reference Ruta, Coates and Quicke2003) and Ruta & Coates (Reference Ruta and Coates2007). Their data matrix of 173 characters (109 cranial and 64 postcranial) recovered a monophyletic nectridean clade and a pattern of relationships within the diplocaulid clade in agreement with Milner (Reference Milner and Panchen1980a). A revaluation of the Moroccan Late Permian Diplocaulus minimus Dutuit, Reference Dutuit1988 by Germain (Reference Germain2010) employed 45 cranial and four postcranial characters to investigate internal nectridean relationships. He found a topology that grouped Diceratosaurus with Batrachiderpeton and the long-horned diplocaulids, supported by the participation of the maxilla in the lateral orbit margin. The node grouping Batrachiderpeton as the sister taxon of the long-horned genera included Peronedon as a separate OTU and was supported by the naris opening anterior to the nasal (but see preceding paragraph). However, the nasal reaches the posteromedial border of the naris in Keraterpeton and Diceratosaurus and is a primitive character for diplocaulids.
As a result of the revised diagnoses (Section 2), primitive characters shared by all diplocaulids are indicated in Figure 20a. Keraterpeton and Diceratosaurus share three cranial and three postcranial primitive characters (Fig. 20b). The exposure of the postorbital occupying the posterior margin of orbit, proximal caudal vertebrae with haemal arches inserted along full length of centrum and overlapping haemal spines are synapomorphies that place Batrachiderpeton crownwards (Fig. 20c), reversing the order of taxa thus: Keraterpeton, Diceratosaurus, Batrachiderpeton. This agrees with the topology in Germain (Reference Germain2010), although that was based erroneously on a primitive diplocaulid character (see previous paragraph).

Figure 20 Cladogram depicting the internal relationships of Keraterpeton, Diceratosaurus and Batrachiderpeton. (A) Primitive characters shared by diplocaulids. (B) Primitive characters shared by Keraterpeton and Diceratosaurus. (C) Primitive characters shared by Diceratosaurus and Batrachiderpeton and long-horned forms.
The outgroup to nectrideans was found to be a node comprising three representative aïstopod taxa in Milner & Ruta's (Reference Milner and Ruta2009) analysis. A detailed study of the neurocranial anatomy of the earliest (Viséan) aïstopod, Lethiscus stocki, by Pardo et al. (Reference Pardo, Szostakwiskji, Ahlberg and Anderson2017) demonstrated a very primitive cranial morphology and a braincase organisation consistent with Devonian stem tetrapods, rather than with other ‘lepospondyl' groups, and placed aïstopods as a stem tetrapod group. Those authors did not include nectrideans in their analysis, but on the basis of their results it appears unlikely that they have any close relationship to aïstopods as found by Milner & Ruta (Reference Milner and Ruta2009).
The derived and specialised nature of nectrideans at their first appearance in the fossil record implies an as yet undiscovered lineage extending back to the earliest Carboniferous. A new analysis, beyond the scope of this paper, is clearly needed, incorporating data from large-scale tetrapod phylogenies to test the interrelationships of nectrideans among early tetrapod groups (Milner & Ruta Reference Milner and Ruta2009). In addition, the newly defined primitive characters of diplocaulids represented by Keraterpeton provide data to test the monophyly of the nectridean clade. The skull morphology of urocordylids reflects adaptations to feeding mechanisms involving a mobile kinetic skull table and snout that is radically different to the akinetic skull organisation of diplocaulids (Milner Reference Milner and Panchen1980a; Bossy & Milner Reference Bossy, Milner, Carroll, Bossy, Milner, Andrews and Wellstead1998). A separate analysis restricted to skull characters alone would prove an interesting exercise in this regard.
5. Acknowledgements
I am indebted to the following colleagues for access to collections and loan of material under their care: Mahala Andrews†, NMS; Colin Forbes†, CAMSM; Mike Howe, BGS; Hugh Ivimey-Cook†, BGS; Nigel Monaghan, NMI; Daniel Pemberton, CAMSM; John Playle, WARMS; Barrie Rickards†, CAMSM; Matt Riley, CAMSM; Chuck Schaff†, MCZ; Patrick Wyse Jackson, TCD. I also thank especially Nigel Monaghan, Matt Riley and Patrick Wyse Jackson for providing catalogue details of their Jarrow material. Nigel Monaghan also kindly supplied images of the holotype of Keraterpeton galvani. Phil Crabb and Phil Hurst (Natural History Museum Image Resources) photographed NHMUK and selected NMI/TCD specimens. I am very grateful to my husband, Andrew Milner, for his help, advice and comments on the draft manuscript, and I thank Florian Witzmann and one other referee for their detailed and careful refereeing that has much improved this manuscript. It is a privilege and pleasure to contribute this work to my long-time friend and colleague Jenny Clack in honour of her dedicated and groundbreaking work that has revolutionised our understanding of early tetrapods.




















