

Hitherto, the earliest Carboniferous deposits of the Tournaisian stage have been considered depauperate, as a result of a poor fossil record of vertebrates and arthropods. A mass extinction event at the end of the Devonian saw the demise of many ancient fish groups such as placoderms, acanthodians and the majority of sarcopterygians that had flourished in the Devonian ‘Age of Fishes' (Sallan & Coates Reference Sallan and Coates2010). Tetrapods of the Devonian, notable for fish-like primitive characters and multidigited limbs, were apparently wiped out (but see Anderson et al. Reference Anderson, Smithson, Mansky, Meyer, Carroll and Clack2015). The poor fossil record following the end-Devonian was taken to be a long-lasting result of this extinction event. Tetrapods from the Tournaisian were virtually unknown, the record consisting only of a range of isolated limb and girdle bones from Nova Scotia (Clack & Carroll Reference Clack, Carroll, Heatwole and Carroll2000; Anderson et al. Reference Anderson, Smithson, Mansky, Meyer, Carroll and Clack2015), and a single articulated tetrapod from western Scotland (Clack Reference Clack2002; Clack & Finney Reference Clack and Finney2005).
Recent excavations in the Borders region of Scotland by members of the TW:eed project (Tetrapod World: early evolution and diversification) have shown that the hiatus is, to a large extent, an artefact of collection failure. A multitude of specimens, including new taxa of tetrapods as well as other vertebrate groups such as chondrichthyans and lungfishes, plus arthropods and plants, are now coming to light (Smithson et al. Reference Smithson, Wood, Marshall and Clack2012; Clack et al. Reference Clack, Ahlberg, Blom and Finney2012). Here, we describe a partial lower jaw similar to the mid-Carboniferous genus Crassigyrinus, from the Tournaisian of Scotland. The specimen was previously mentioned and figured in Smithson et al. (Reference Smithson, Wood, Marshall and Clack2012, fig. 2C, D).
Crassigyrinus scoticus is known from five main specimens, all from Scotland, dated, with some margin for error, as ‘Upper Viséan' or ‘Lower Namurian E1' and, in more recent terminology, uppermost Viséan to lowest Serpukovian, in the Brigantian to Pendleian European substages (Gradstein et al. Reference Gradstein, Ogg, Schmitz and Ogg2012). The holotype specimen consists of the right snout, cheek and suspensorial region of a skull (NMS G.1859.33.104) from the Gilmerton Ironstone near Edinburgh and first described by Watson (Reference Watson1929). Other specimens include NHMUK R310, a partial lower jaw exposed in external view; GSE 4722, an almost complete lower jaw exposed in internal view; and NHMUK 30532, a partial cranium (all from Gilmerton near Edinburgh, and probably all from the Gilmerton Ironstone); NHMUK R10000, an articulated individual including a skull and much of the postcranium from the Dora Bone Bed, in the Limestone Coal Formation near Cowdenbeath. Additionally, NMS G.1975.40.50/51, also from the Dora Bone Bed, consists of matching articular and quadrate. These were described and/or listed by Panchen (Reference Panchen1973, 1985), Clack (Reference Clack and Milner1996, Reference Clack1998) and Ahlberg & Clack (Reference Ahlberg and Clack1998).
The new specimen described here (Fig. 1) is about 20 million years older than any of these specimens, potentially extending the geological range of the genus much further down towards the base of the Carboniferous, and about 10 million years after the end of the Devonian. When first collected, its identity, obscured by the coarse matrix in which it was preserved, was eventually recognised to be part of a lower jaw. It was still largely in the matrix when Stanley Wood first identified it as belonging to Crassigyrinus, an intuitive observation that is probably correct.

Figure 1 Cf. Crassigyrinus UMZC 2011.9.1: (A) specimen photograph in lateral view. Asterisk indicates the break at which the photograph in (F) was taken; (B) interpretive drawing of specimen in external view; (C) interpretive drawing of specimen in medial view; (D) photograph of specimen in medial view. Double-ended arrow shows portion scanned in Figure 2; (E) enlargement of area showing main articulation point for prearticular; (F) Section at level of asterisk in (A), arrow shows the section through the canal in Figure 2. Grey fill indicates matrix, diagonal hatch indicates broken bone. Scale bars = 10 mm.
1. Material and methods
The new specimen, UMZC 2011.9.1, from the locality of Burnmouth, in the Borders Region of Scotland, comes from deposits dated as within the CM (claviger-macra) palynozone, although the palynozonation of the Tournaisian stage in Scotland is being reviewed as a result of work undertaken as part of the TW:eed project. The sequence at Burnmouth covers much of the Tournaisian stage, somewhat condensed, but extending from slightly below the Devonian–Carboniferous boundary, through to the base of the Viséan Fell Sandstone.
The specimen was collected from a 200 mm-thick localised conglomerate lag that occurs at the base of a 4 m-thick fluvial sandstone channel deposit, about 383 m above the base of the Ballagan Formation at Burnmouth. The conglomerate lies between sandstones 4 & 5 of Scrutton & Turner (Reference Scrutton, Turner and Scrutton1995) and is weakly bedded, poorly sorted, matrix-supported and cemented with dolomite, which makes preparation extremely difficult in places. Centimetre-sized clasts (average size 30 mm) are composed of red and grey siltstone and bioclasts, within a coarse sandstone matrix. The clasts and matrix are likely derived from lakes (grey silt) and palaeosols (red silt) on the floodplain, which were eroded and incorporated into the lag during river flow and subsequent sediment deposition. Bioclasts are more concentrated towards the base of the unit and include numerous isolated bones, including some very large examples, of lungfishes, rhizodonts, gyracanth spines and girdle elements, actinopterygians, a few teeth of the stem chondrichthyan Ageleodus, and occasional tetrapod elements. The bed also contains charcoalified plant stems, charcoal fragments and wood fragments. However, the more fragile ostracod and bivalve fossils that are a common element of floodplain sediments are absent. The jaw described here is one of the larger finds from this bed.
Collection from this bed presents difficulties as it is almost vertically orientated like all other beds at Burnmouth, and is set between the overlying massive sandstone and a cementstone, close to the cliff face. There is a high risk of losing fragments into the gap between these two harder beds left by erosion of the relatively soft conglomerate.
The jaw was prepared mechanically by JAC, mainly with a mounted needle under binocular microscope, and occasional judicious use of a dental mallet. The specimen was set into a series of silastomer jackets as each face was prepared in turn. The bone was extremely fragile and required careful consolidation with the resin Paraloid B72. Matrix has been left on parts of the lingual surface, where the bone is too delicate to support itself.
For comparison with existing Crassigyrinus specimens, NHMUK R10000, NHMUK R310 and GSE 4722 were scanned using micro-computed tomography (µCT) at the Imaging and Analysis Centre of the Natural History Museum (London, UK) on an X-Tek HMX-ST µCT 225 scanner (Nikon Metrology, Tring, UK). Scans of NHMUK R10000 produced two separate data sets consisting of 1958 DICOM slices of the anterior half of the specimen and 1953 DICOM slices of the posterior half of the specimen with a resolution of 0.1083 mm/voxel. GSE 4722 is preserved in three large pieces, each of which was scanned separately. The anterior portion of this specimen produced 1995 DICOM slices with a resolution of 0.1271 mm/voxel. The central-posterior portion was scanned to produce 1995 slices with a resolution of 0.1099 mm/voxel. The posterior portion of GSE 4722 was scanned to produce 1687 DICOM slices with a resolution of 0.1099 mm/voxel. NHMUK R310 was scanned in two parts: the anterior half produced 1956 DICOM slices at a resolution of 0.1034 mm/voxel; while the posterior half produced 1939 DICOM slices at a resolution of 0.1034 mm/voxel. Scans were segmented to remove matrix from bone and separate individual bones from each other, then rendered into three-dimensional models using Avizo 7.1.1 (FEI Visualization Sciences Group, Mérignac Cedex, France). An isolated articular, NMS 1975.48.50, was also studied and part of UMZC 2011.9.1 was µCT-scanned at the Cambridge Biotomography Centre (Zoology Department) at the University of Cambridge on an X-Tek H 225 µCT scanner (Nikon Metrology, Tring, UK). This produced a data set of 1866 16bit TIFF images with a resolution of 0.0465 mm/voxel. These 16-bit TIFFs were converted to 8-bit TIFFs using imageJ and the resulting dataset loaded into Mimics (Materialise NV – Technologielaan 15, 3001 Leuven, Belgium).
Institutional abbreviations. GSE, British Geological Survey, Edinburgh; NHMUK, Natural History Museum, London; NMS, National Museum Scotland; UMZC, University Museum of Zoology, Cambridge.
2. Systematic palaeontology
Osteichthyes Huxley, Reference Huxley1880
Sarcopterygii Romer, Reference Romer1955
Tetrapodomorpha Ahlberg, Reference Ahlberg1991
Family Crassigyrinidae Huene, Reference Huene1948
Genus cf. Crassigyrinus Watson, Reference Watson1929
Type species. Crassigyrinus scoticus.
Specimen. UMZC 2011.9.1. cf. Crassigyrinus.
3. Description
3.1. Comparative anatomy of the Burnmouth jaw
The preserved part of the specimen measures 250 mm and includes the angular, postsplenial and parts of the splenial, dentary and surangular of the lower jaw. It lacks the symphysial region, prearticular, articular and all the dentigerous bones, apart from fragments of the dentary (Fig. 1).
The lateral line is a deep open groove approximately following the ventral margin, although approaching it more closely near the mid-point of the angular. It continues to the rearmost point of the surangular. Ornamentation ventral to the lateral line takes the form of long, fairly regular grooves and ridges, less marked on the angular, which instead bears a few elongate, individual pits. Dorsal to the lateral line, the ornamentation consists of sparsely distributed individual pits, very shallow on the angular, deeper on the postsplenial and surangular, and shallow radiating grooves. The fragment of dentary, parts of the angular and the surangular also bear fine striations. There is no indication of the extent or orientation of the adductor fossa, nor whether or not there was a surangular crest, and there is no evidence of a postsplenial pit line.
By comparison with the isolated articular NMS 1975.48.50, it appears that the articular in UMZC 2011.9.1 is missing. The smooth excavated surface at the rear of the jaw is more likely to have been contributed by the surangular, as seen in the NMS articular. A series of grooves and ridges at the extreme posterior end of the jaw could indicate a suture to the articular. From that point, the ventral margin of the jaw is smooth until approximately the midpoint of the angular, suggesting no sutural contact between either the angular or the surangular and the prearticular until that point. Sutures between the angular and surangular are difficult to determine on the internal surface in part because of breakages and adhering matrix.
From the section of jaw exposed in a break, a circular feature indicated the existence of a canal running through the lower part of the jaw (Figs 1F, 2). Micro-CT scanning has revealed the shape and course of the canal and confirmed that it does not appear connected to the lateral line, except possibly at the extreme posterior end, where matrix in the lateral line may obscure a foramen (Figs 1B, 2). It takes a medial course through the angular (Fig. 2B, arrows) and is distinct from the groove of the lateral line canal. At least some of the branches off the canal appear to be genuine features, although some may be artefacts as a result of breakages. The canal could be the course of a blood vessel or nerve, but it appears to end blindly rather than exiting via a foramen. No such canal has been identified in any of the other specimens, but most of them are too fragmented to be certain. On the lingual surface near the articulation, two foramina appear to be linked by what may be a branch of this canal (Fig. 2A arrows). The status of this feature as an apomorphy is unknown; it could be at individual or specific level rather than generic.
A series of robust ridges and crests marks the ventral edge and inner face of what is probably the angular, suggesting a sutural contact between it and the prearticular along a length of about 50 mm (Fig. 1). This may constitute a very limited mesial lamina of the angular as noted by Ahlberg & Clack (Reference Ahlberg and Clack1998) in specimen GSE 4722 (Fig. 3) and NHMUK R10000 of Crassigyrinus scoticus. Thereafter, the lower margin of the Burnmouth jaw is smooth, except for a small protuberance near the posterior end of the postsplenial. This may represent a point of attachment for a pillar defining a Meckelian fenestra. There is no other contact surface for the prearticular along the length of the jaw. Only the posteriormost part of the splenial is preserved, including its contact with the postsplenial, but the region where the mesial lamina would normally be situated is missing. Figure 4 shows the three C. scoticus jaw specimens compared with the Burnmouth jaw and to the same scale.
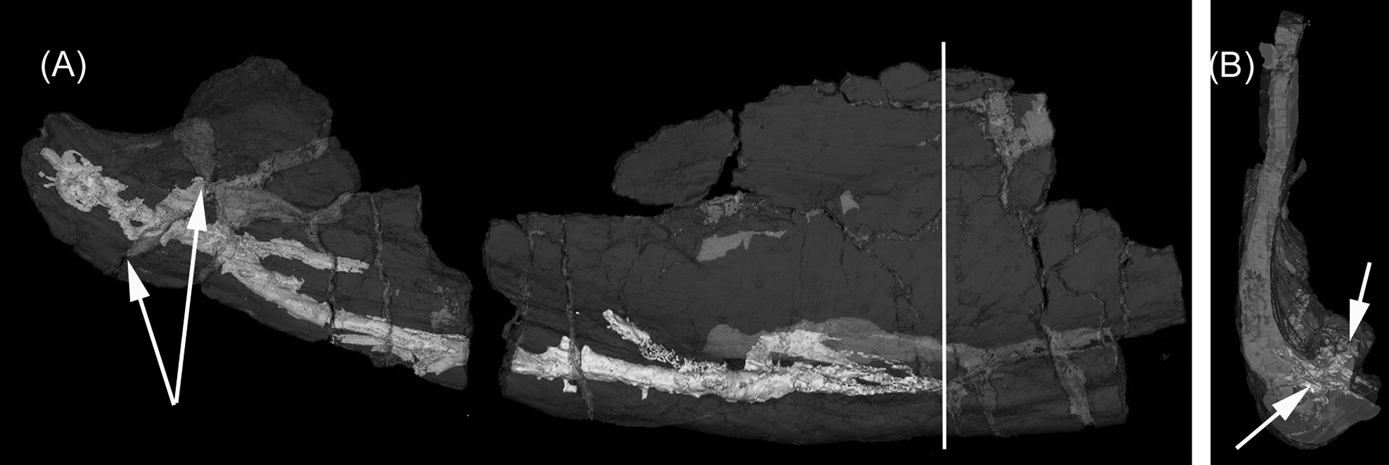
Figure 2 Cf. Crassigyrinus UMZC 2011.9.1: (A) micro-CT of the posterior region showing the course of the canal (see double arrow in Figure 1(D). Vertical line shows the position of the view in (B). Paired arrows show the possible canal that crosses the surangular to emerge on the internal surface; (B) view from the posterior side of the section shown in (A), looking anteriorly along the jaw. At this point, only the tip of the canal can be seen (left arrow): other traces (right arrow) are breaks.
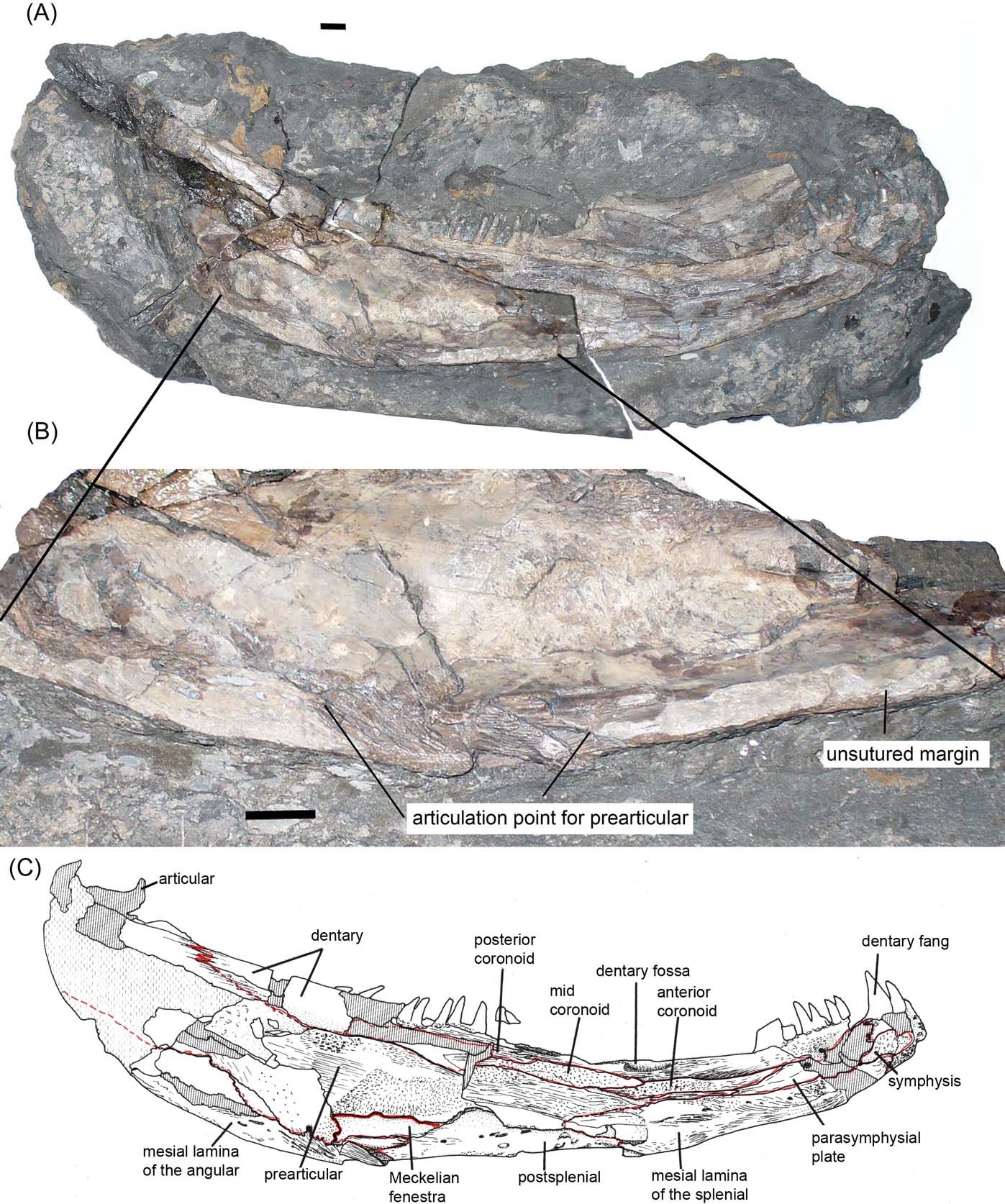
Figure 3 Crassigyrinus scoticus, GSE 4722: (A) complete specimen, exposed in medial view; (B) enlargement of middle section showing the lower margin and articulation points for the prearticular. Note that areas to the right and left of the main articulation point have been abraded away; (C) interpretive drawing of specimen from Ahlberg & Clack (Reference Ahlberg and Clack1998), with sutures shown in red. Scale bars = 10 mm.

Figure 4 Crassigyrinus lower jaws: (A) C. scoticus, Part of NHMUK R10000, placed on the reconstruction of the jaw by Panchen (Reference Panchen1985), with part of the posterior end represented by a line drawing. Reproduced from Panchen (Reference Panchen1985) with permission; (B) Cf. Crassigyrinus, UMZC 2011.9.1, lateral view; (C) C. scoticus, NHMUK R310; (D) C. scoticus, GSE 4722. Photographs all to the same scale. Scale bars=10 mm.
3.2. Micro-CT data from C. scoticus jaws
The inner face of the jaw of C. scoticus is most clearly exposed in specimen GSE 4722 (Fig. 3), most recently described and reinterpreted by Ahlberg & Clack (Reference Ahlberg and Clack1998), but the external surface of this specimen is inaccessible by conventional means. The specimen is broken into three parts, all of which are exposed in medial view and which we scanned. The middle portion (Fig. 5E) preserves parts of the prearticular, angular and the posterior portion of the postsplenial. The mesial lamina of the angular is present, but incomplete, on the central portion of GSE 4722. Strong deformation and breakage may have reduced the lamina in this specimen to a pronounced ridge. Although damaged, visual inspection and µCT-scanning demonstrate that this ridge medially overlaps the thickened ventral margin of the prearticular in this specimen. Nonetheless, there are significant regions along the ventral edge where it is clear that no sutural contact existed between the angular and prearticular. The extent of the sutural area on the angular for the prearticular in GSE 4722 can be estimated as about 100 mm long, and is thus more extensive than that in the Burnmouth jaw.
The lateral surface of the Burnmouth jaw shows the ornament to be consistent with that of other specimens of C. scoticus, although the finer striations are beyond the resolution of the scan data (Fig. 5E). The lateral line expression is similarly deep and continuous, but more dorsally placed than in C. scoticus.
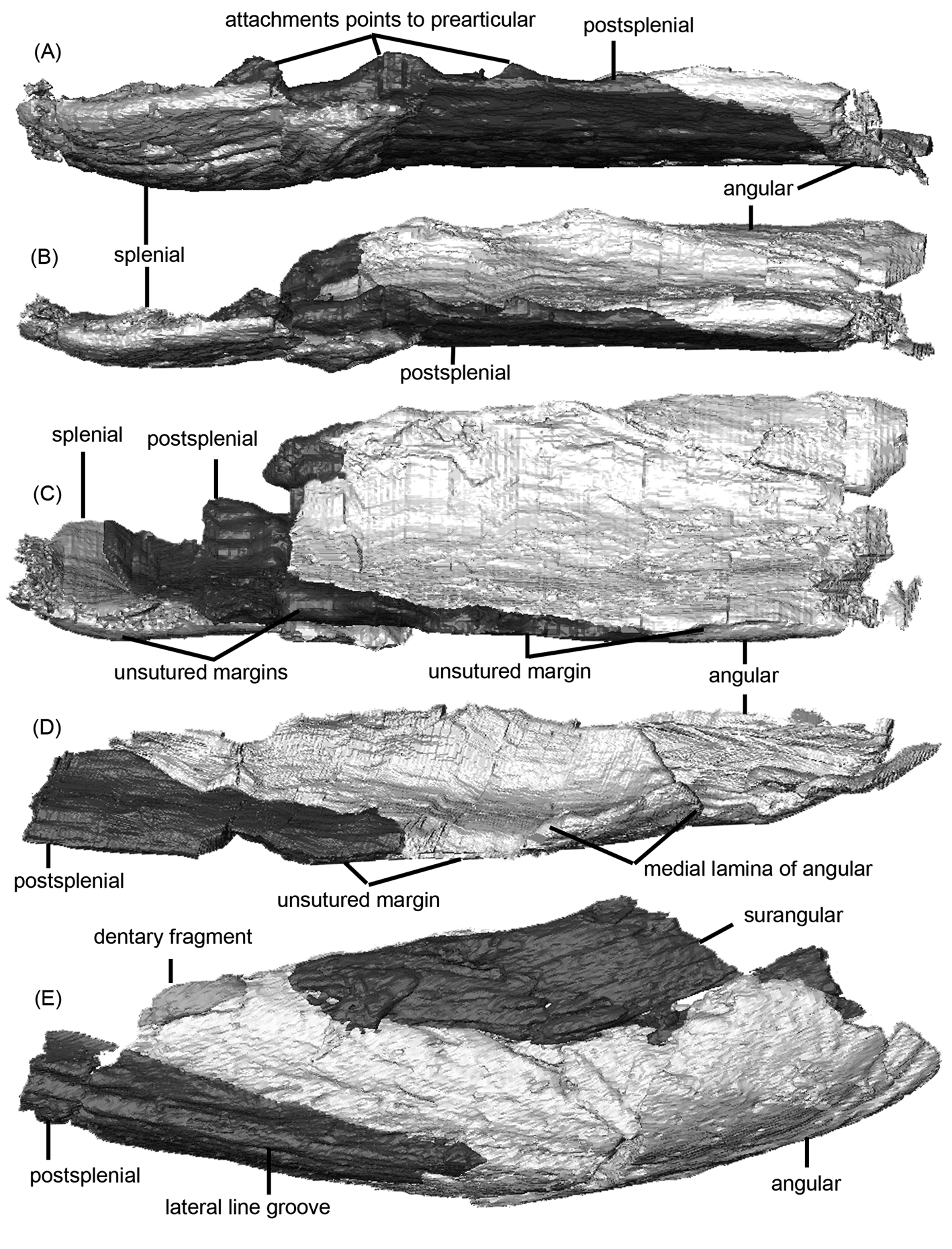
Figure 5 Crassigyrinus scoticus. Micro-CT renders of lower jaws: (A–C) posterior portion (isolated angular, postsplenial and splenial) of NHMUK R310 in (A) ventral view, (B) ventro-medial view and (C) medial view – i.e., the specimen is rotated about its long axis in (A–C); (D) posterior portion (isolated angular and postsplenial) of right lower jaw ramus of NHMUK R10000; (E) middle portion (isolated surangular, angular and postsplenial) of GSE 4722 in lateral view, showing lateral line and dermal ornament.
Specimen NHMUK R310 (Fig. 5A–C) preserves the portion of the jaw from the symphysis to the posterior end of the postsplenial. The ornament pattern on the postsplenial is in many ways similar to that of the Burnmouth jaw, with elongate ridges and grooves ventral to the lateral line, but more numerous and shorter pits and grooves dorsal to it. The dentary and a preserved fragment of the surangular are marked by fine almost parallel striations. The lateral line itself is similarly expressed as a deep groove and positioned parallel to the ventral edge. Segmentation of µCT-scan data reveals the medial surface of the specimen for the first time (Fig. 5A–C). The mesial lamina of the splenial can be visualised and internally overlaps the adsymphysial and the anterior process of the prearticular. Posterior to the mesial lamina, the splenial and prearticular separate and the ventral margin of the prearticular approaches but does not firmly contact the postsplenial. The angular weakly contacts the prearticular, although the mesial lamina of the angular is not preserved in this specimen.
Specimen NHMUK R10000 is the most complete skull known for C. scoticus; unlike GSE 4722 and NHMUK R310, NHMUK R10000 experienced dorsoventral (as opposed to mediolateral) compression. The anterior portion of the right jaw is illustrated in Figure 4A. The labial surfaces of both antimeres have similar ridge and groove ornament to that of the Burnmouth jaw, if more marked. The lateral line is likewise an open groove parallel to the ventral margin. Much of the internal face including the prearticular is damaged and broken, but µCT scans have clarified the morphology and sutural contacts of the medial aspect of the lower jaw. Both sides preserve the prearticular, articular and infradentaries, as well as all the dentigerous bones. Posterior to the mesial lamina of the splenial, the prearticular intermittently makes point and lapping contacts with the splenial and postsplenial, but these are almost certainly due to the deformation undergone by the specimen and are artefacts. The ventral margin of the prearticular thickens at the level of the angular (as in GSE 4722) and passes lateral (internal) to the mesial lamina of the angular on both sides of NHMUK R10000. The prearticular does not contact the surangular on either side of this specimen and terminates by ventrally and medially lapping the articular. The surangular wraps around the articular, broadly overlapping it both laterally and posteriorly. No fine interdigitations are visible in µCT scans between the surangular and articular; however, larger-scale ridging is present between these two bones. This arrangement leaves only a small area of the articular externally exposed posteromedially, in addition to the jaw joint surface. Scans and the resulting three-dimensional models (Fig. 5D) of NHMUK R10000 confirm the presence of a substantial mesial lamina of the angular. This is broken and folded dorsally in the right ramus, but is better visualised than on the left ramus. It overlaps the ventral margin of the prearticular medially (Fig. 5D). These observations all accord with the condition seen in the Burnmouth jaw. Altogether, the morphology of the medial aspect of the lower jaw in NHMUK R10000 is very similar to that of the Burnmouth jaw.
4. Discussion
The new Burnmouth lower jaw specimen UMZC 2011.9.1 requires comparison with other known Early Carboniferous tetrapod jaws, as well as those of Late Devonian ones, to establish its identity and to examine its combination of features both primitive and derived. Among Early Carboniferous forms, those of Pederpes (Clack & Finney Reference Clack and Finney2005) Sigournea (Bolt & Lombard Reference Bolt and Lombard2006), Whatcheeria (Lombard & Bolt Reference Lombard, Bolt, Carrano, Gaudin, Blob and Wible2006), Greererpeton (Smithson Reference Smithson1982; Bolt & Lombard Reference Bolt and Lombard2001) and a lower jaw from Parrsboro, Nova Scotia (Sookias et al. Reference Sookias, Böhmer and Clack2014) are the most appropriate comparators. Of these, only Pederpes is Tournaisian in age, with all the others being late Viséan or early Serpukhovian. The Parrsboro jaw preserves mainly the dentigerous bones, but has a narrow denticulated prearticular dorsally bounding a large Meckelian space. There are few points of comparison between this and the Burnmouth jaw. Sookias et al. (Reference Sookias, Böhmer and Clack2014) found the Parrsborough jaw to nest within a polytomy of mainly later Carboniferous forms. In that analysis Sigournea, Whatcheeria and Greererpeton were all found to be basal to Crassigyrinus. Clack et al. (Reference Clack, Ahlberg, Blom and Finney2012) did not include Sigournea, but their analysis also placed Whatcheeria and Greererpeton basal to Crassigyrinus. This is in contrast to the analysis of Ahlberg & Clack (Reference Ahlberg and Clack1998), in which Greererpeton sat crownward of Crassigyrinus. Pederpes and Whatcheeria sometimes emerge as basal to Crassigyrinus (Warren Reference Warren2007) and sometimes immediately crownward (Ruta & Clack Reference Ruta and Clack2006). Crassigyrinus has also appeared in a more basal position amongst embolomeres and Eoherpeton (Klembara et al. Reference Klembara, Clack, Milner and Ruta2014), or with baphetids (Laurin Reference Laurin2004).
Sigournea, which has so far been included in only one phylogenetic analysis (Sookias et al. Reference Sookias, Böhmer and Clack2014), resembles the Burnmouth jaw in the position and expression of the lateral line canal, but differs in having a mesial lamina of the angular that appears to suture with the prearticular to a more posterior extent. The postsplenial shows evidence of several separate Meckelian fenestrae along its ventromedial margin. Larger specimens of the jaw of Whatcheeria preserve an ossified Meckelian bone between the angular, postsplenial and the prearticular, though in smaller specimens it is absent or unossified, and there is no angular–prearticular or postsplenial–prearticular contact. There is very little ornamentation on the external face of the jaw, and the lateral line is expressed as a series of pores, some of which are elongated. The lower jaw of Pederpes is not well enough preserved for comparison. Greererpeton possesses well-marked radiating pit and ridge ornament on the external face of the lower jaw, and the lateral line, though mainly open, is interrupted in places and posteriorly is seen as pores. There is a strong sutural contact between the surangular and prearticular and the angular and prearticular, although there is a large single Meckelian fenestra in which the prearticular does not contact the postsplenial. These combinations of characters appear to rule out the Burnmouth specimen from all three of these genera.
Ahlberg & Clack (Reference Ahlberg and Clack1998) showed a progressively increasing degree of sutural contact between the prearticular and the infradentary bones through the evolution of early tetrapods, namely the postsplenial and angular. Although there is a contact between the angular and prearticular in some Devonian taxa such as Ventastega and Ichthyostega, it does not appear to be strongly sutured, and there is no such contact in Acanthostega, Metaxygnathus or Ymeria (Clack et al. Reference Clack, Ahlberg, Blom and Finney2012). In Whatcheeria, the condition is variable depending on size, with either no contact or an intervening Meckelian ossification, but no mesial lamina of the angular. Such ontogenetic effects are interesting and could be more widespread than we currently appreciate. Both Greererpeton and Crassigyrinus may represent early stages of the development of contacts between the prearticular and the infradentary series. In Greererpeton, the prearticular has a smooth and excavated margin defining a large single Meckelian fenestra, an apparently more specialised condition compared to the ragged and poorly defined ventral margin in Crassigyrinus, as seen in GSE 4722.
When compared with the known specimens of C. scoticus the new jaw shows similarities such as the overall shallow and curved shape of the jaw and the condition of the dermal ornament. More general early tetrapod characters include the open lateral line sulcus and the degree of contact between the prearticular with the angular and postsplenial. The position of the lateral line is further dorsally placed for most of its length than in C. scoticus. There are also differences in proportion between the infradentary bones between the Burnmouth jaw and C. scoticus, as seen in Figure 6. However, such differences could be expected in taxa 20 million years apart, and could simply indicate specific rather than generic distinctions. In summary, the Burnmouth jaw shows most resemblance to that of C. scoticus amongst Early Carboniferous tetrapod lower jaws, and in fact also later ones such as baphetids.
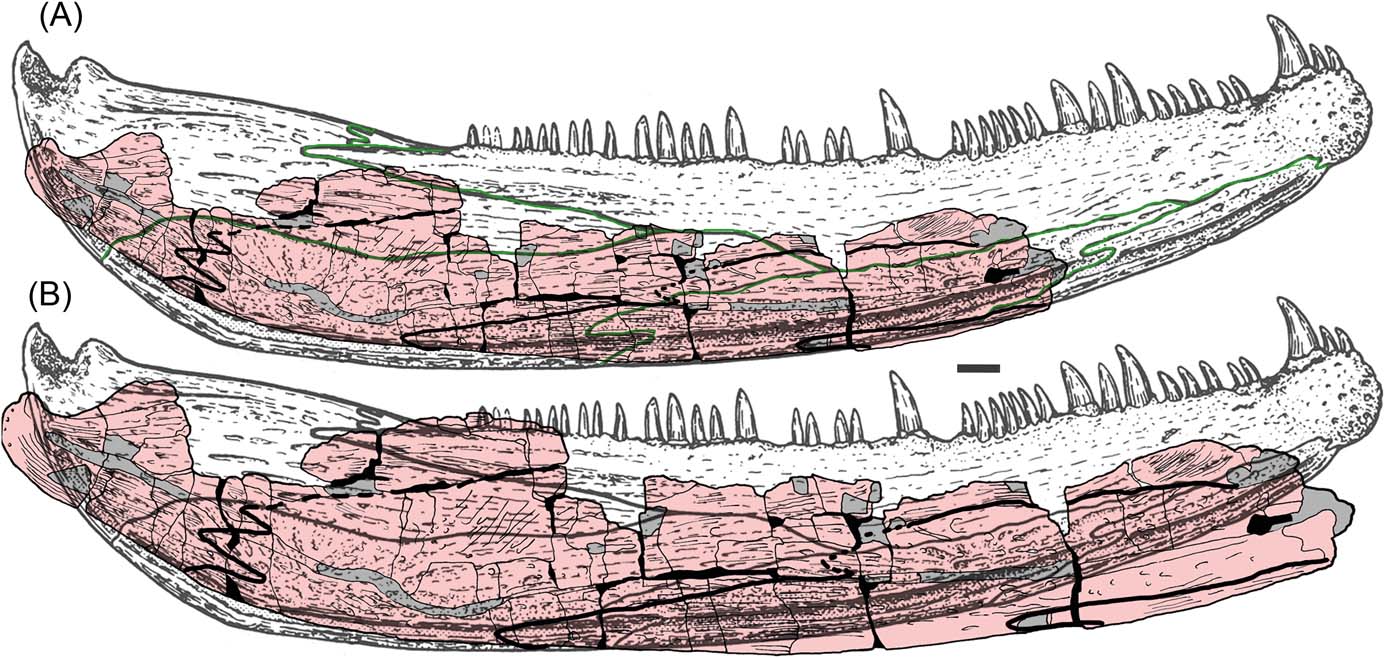
Figure 6 C. scoticus lower jaw reconstruction from Panchen (Reference Panchen1985), with UMZC 2011.9.1 superimposed. Panchen's sutures shown in green: (A) with UMZC 2011.9.1 to the same scale; (B) with UMZC 2011.9.1 to approximately the same size.
From the articular to the point of inception of the splenial, all C. scoticus jaws are similar in size. In GSE 4722, the total length is about 320 mm, and the specimen shows a greater degree of curvature than the Burnmouth jaw. Differential crushing and distortion could account for that, in that the bone of GSE 4722 is thicker and much more robust than that of the Burnmouth jaw, and so is less liable to distortion.
The holotype specimen of C. scoticus, NMS G.1859.33.104, consists of the right side of a skull from the tip of the snout to the quadrate. It is c.336–340 mm long. Assuming that the lower jaw would be slightly shorter, it is possible to estimate its length at c.325–330 mm. The ‘Dora Bone Bed' specimen, NHMUK R.10000, has a lower jaw length of 328 mm, and a restoration of NHMUK R310 (Fig 4), based on the reconstruction by Panchen (Reference Panchen1985) measures 320 mm. A feature of C. scoticus is that all specimens are of almost identical size. This could be considered a co-incidence, but it could also imply that the genus reached, if not a determinate size, at least one in which growth slowed considerably after a certain ontogenetic age.
Attribution of the Burnmouth jaw to a genus only known from much later in time might be problematic. However, the genus Neoceratodus, the modern Australian lungfish, is known from the Early Cretaceous (Kemp & Molnar Reference Kemp and Molnar1981); although Cavin et al. (Reference Cavin, Suteethorn, Buffetaut and Tong2007) suggested that it might extend back to the Early Triassic. Such longevity for a genus is thus not unprecedented.
On the other hand, there is a precedent for animals with similar skull morphology to have very different postcranial characteristics and be placed in different genera. The Viséan tetrapod Whatcheeria, whilst having a similar skull shape and dermal bone anatomy to that of the Tournaisian Pedepes, has a humerus that differs radically from that of Pederpes. There has been doubt expressed as to the validity of the family Whatcheeriidae (Warren Reference Warren2007); in addition to the differences between Pederpes and Whatcheeria, Ossinodus lacks many of the features characteristic of the other members. Such a wide disparity between constituent members of a family might suggest caution in placing them together. Without further evidence, a similarly cautious argument could be used against attributing such a small fragment of a lower jaw to a genus with a highly specialised postcranium.
The Burnmouth jaw is less robust than the later specimens of Crassigyrinus, and the degree of prearticular–angular contact less extensive. However, given the similarities in size, ornamentation, lateral line expression and overall construction, attribution to the genus Crassigyrinus may be justified, but only further evidence from Tournaisian specimens could corroborate this attribution.
5. Acknowledgements
With this publication, we acknowledge the perspicacity and intuition, as well as the life-time achievements, that Stan Wood brought to the pursuit of Carboniferous vertebrate palaeontology in the UK. We wish to thank staff of the Natural History Museum in London, Zerina Johanson, Emma Bernard and Sandra Chapman, for help with access to collections and to the µCT facility. Micro-CT-scanning of C. scoticus specimens was carried out by Dan Sykes (Imaging and Analysis Centre, Natural History Museum, London, UK). Keturah Smithson of the UMZC undertook µCT scanning and segmenting of UMZC 2011/.9.1 for Figure 2. Alejandra Sánchez-Eróstegui and Jean-Luc Garnier (FEI Visualization Sciences Group, Mérignac Cédex, France) provided technical support for Avizo. Rob Clack collected the specimen in the field; and the TW:eed team members contributed to our understanding of the geology of Burnmouth. LBP was funded by a Marie Curie International Incoming Research Fellowship (“Tetrapods Rising”, 300161); and JAC and CEB by NERC consortium grants NE/J022713/1 and NE/J020729/1, known as the TW:eed project. This is a contribution to IGCP project 596.






