

Phytosaur skulls are abundant in the Upper Triassic Chinle Formation in the southwestern United States, especially those of pseudopalatine phytosaurs, the most derived clade of phytosaurs that includes various species that have been referred to Pseudopalatus Mehl, 1928 and Redondasaurus Hunt & Lucas, Reference Hunt, Lucas, Lucas and Morales1993. As a rough estimate, more than 75 skulls of these two genera have been recovered over the last hundred years in Arizona, Colorado, New Mexico, Texas and Utah. This number represents approximately one-third of all phytosaur skulls known from North America. Given the abundance, it is striking how little information is available about the detailed cranial anatomy of this group. The seminal paper on phytosaur skull morphology is still Camp's (Reference Camp1930) study based on observations on the types of Smilosuchus gregorii (Camp Reference Camp1930) and Smilosuchus adamanensis (Camp Reference Camp1930). Because the configuration of the skull elements in many specimens of pseudopalatine phytosaurs, among them almost all type specimens, is rather imperfectly preserved, the majority of accounts deal more with the general appearance of the skull rather than with osteological details (Cope Reference Cope1881; Huene Reference Huene1915; Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993; Long & Murry Reference Long and Murry1995; Lucas et al. Reference Lucas, Heckert, Zeigler, Hunt, Heckert and Lucas2002). More recent accounts of older collections from Arizona (Heckert & Lucas Reference Heckert, Lucas, McCord and Boaz2000) and New Mexico (Zeigler et al. Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b) and recently discovered, well-preserved material of pseudopalatine phytosaurs from the Petrified Forest Member of the Chinle Formation and the Redonda Formation of New Mexico (Heckert et al. Reference Heckert, Harris, Lucas, McCord and Boaz2000; Zeigler et al. Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002b, Reference Zeigler, Heckert, Lucas, Zeigler, Heckert and Lucas2003a) similarly provide few detailed osteological data. Parker & Irmis (Reference Parker and Irmis2006) have provided, to date, the most detailed cranial description of a North American pseudopalatine phytosaur, with their description of the partial skull of M. jablonskiae from the Petrified Forest. Other than that, the most informative overall studies of a pseudopalatine phytosaur from North America to date remain Camp's (Reference Camp1930) account on M. tenuis and Mehl's (Reference Mehl1922) description of M. andersoni. Parker & Irmis (Reference Parker and Irmis2006) defined a node-based clade for the Pseudopalatinae following Hungerbühler (Reference Hungerbühler2002). They defined the clade as Nicrosaurus, Mystriosuchus, Pseudopalatus, Redondasaurus, and all the descendants of their last common ancestor.
Here we present a detailed description of the cranial anatomy of the genus Machaeroprosopus on the basis of three skulls. The excellent preservation of the specimens provides a wealth of new morphological information on previously debated anatomical structures, in particular relating to the braincase and the palate, both regions of the skull that are either frequently inaccessible or rarely preserved because of their fragile nature. Moreover, two of the specimens represent a new species of Machaeroprosopus. They provide additional evidence for sexual dimorphism in phytosaurs, and their unusual combination of characters has important implications for the taxonomy of North American pseudopalatine phytosaurs.
Taxonomic note. Parker et al. (Reference Parker, Hungerbühler and Martz2013 (this volume)) demonstrate that Pseudopalatus Mehl, Reference Mehl1928 is a junior synonym of Machaeroprosopus Mehl, Reference Mehl1916. We argue below that there are good reasons to consider the genus Redondasaurus Hunt & Lucas, Reference Hunt, Lucas, Lucas and Morales1993 a junior synonym of Machaeroprosopus as well. Consequently, in the text we refer to all nominal species that are currently included in Pseudopalatus and Redondasaurus as species of Machaeroprosopus. We provisionally follow Stocker (Reference Stocker2010) in using the genus-group names Leptosuchus Case, Reference Case1922sensu strictu, Smilosuchus Long & Murry, Reference Long and Murry1995 and Pravusuchus Stocker, Reference Stocker2010 for taxa that were traditionally part of a more inclusive concept of Leptosuchus.
Institutional abbreviations. AMNH, American Museum of Natural History, New York, NY; CM, Carnegie Museum of Natural History, Pittsburgh, PA; GPIT, Paläontologische Sammlung, University of Tübingen, Germany; NHMUK, The Natural History Museum, London, UK; NMMNH, New Mexico Museum of Natural History and Science, Albuquerque, NM; PEFO, Petrified Forest National Park, AZ; SMNS, Staatliches Museum für Naturkunde Stuttgart, Germany; TTU, Museum of Texas Tech University, Paleontology Division, Lubbock, TX; UCMP, University of California, Museum of Paleontology, Berkeley, CA; UMMP, Museum of Paleontology, University of Michigan, Ann Arbor, MI; UMo, University of Missouri, Department of Geological Sciences, Vertebrate Paleontology Collection, Columbia, MO; YPM Yale Peabody Museum, New Haven, CT.
1. Geological setting of the Patricia Site (locality number TTU VPL 3870)
Upper Triassic strata of the Dockum Basin are exposed along the eastern escarpment of the Southern High Plains in the Texas Panhandle, and continue along the western escarpment and the Pecos River Valley in eastern New Mexico. Researchers from New Mexico (e.g., Lucas & Hunt Reference Lucas, Hunt, Lucas and Hunt1989; Lucas et al. Reference Lucas, Anderson, Hunt, Ahlen, Peterson and Bowsher1994, Reference Lucas, Heckert, Hunt, Lucas and Ulmer-Scholle2001) and Texas (Lehman Reference Lehman, Ahlen, Peterson and Bowsher1994a, Reference Lehmanb; Lehman & Chatterjee Reference Lehman and Chatterjee2005; Martz Reference Martz2008) advocate two competing stratigraphic schemes. We note that this debate is largely about priority and nomenclature, and both stratigraphic schemes agree that the units relevant here, the upper portion of the Cooper Canyon Formation in Texas and the Bull Canyon Formation in New Mexico, are equivalents in time. We follow the stratigraphic nomenclature as proposed by Lehman (Reference Lehman, Ahlen, Peterson and Bowsher1994a). Accordingly, the Dockum Group of Texas includes, in ascending order, four mappable units: the Santa Rosa Sandstone, the Tecovas Formation, the Trujillo Sandstone and the Cooper Canyon Formation (upper Cooper Canyon of Martz Reference Martz2008). A fifth and uppermost unit of the Dockum Group, the Redonda Formation, is restricted to eastern New Mexico and seems to grade eastward and southward into the upper part of the Cooper Canyon Formation.
The strata forming the Trujillo Sandstone and Cooper Canyon Formation represent a thick (maximum more than 150 m) alluvial depositional sequence, separated from the underlying Tecovas Formation by a locally angular unconformity. The basal part of the Trujillo Sandstone consists of thick, multistoried and laterally-extensive fluvial channel-sand bodies that reflect multiple phases of channel incision, lateral migration and aggradation. The Trujillo Sandstone grades and intertongues upward with an increasing proportion of fluvial flood-plain mudstone, and thus the overlying Cooper Canyon Formation is largely mudstone-dominated. Interbedded fluvial channel sandstone units in the Cooper Canyon Formation mostly reflect single phases of channel migration and aggradation, and so tend to be single-storied.
The Patricia Site TTU VPL 3870 served as an example for vertebrate bone accumulation in channel facies (Lehman & Chatterjee Reference Lehman and Chatterjee2005). The site is located about 13 km southwest of Post, in Garza County. Vertebrate fossils, along with carbonaceous plant remains, occur here within the upper part of the Cooper Canyon Formation, in channel deposits of fine-grained sandstone and green mudstone (Fig. 1).

Figure 1 Patricia Site TTU Vertebrate Paleontology Locality 3870, Garza Co., Texas: (A) view of Site 1C, the locality of TTU-P10074; (B) composite section of Locality 3870, with depositional interpretation and the stratigraphic position of phytosaur specimens. Arrows indicate the position of the two characteristic conglomeratic beds that bracket the main fossiliferous strata.
The most abundant fossils collected from this site are phytosaurs, representing more than 90% of the identifiable remains. In addition TTU VPL 3870 yielded isolated remains of a palaeonisciform actinopterygian, a temnospondyl amphibian, the aetosaur Typothorax, the rauisuchid Postosuchus, the poposauroid Shuvosaurus and a theropod dinosaur (Cunningham et al. Reference Cunningham, Hungerbühler, Chatterjee and McQuilkin2002).
2. Systematic paleontology
Archosauriformes Gauthier, Kluge & Rowe, Reference Gauthier, Kluge and Rowe1988
Phytosauria Jaeger, Reference Jaeger1828sensu Reference NesbittNesbitt 2011
Phytosauridae Jaeger, Reference Jaeger1828sensu Doyle & Sues Reference Doyle and Sues1995
Pseudopalatinae Long & Murry, Reference Long and Murry1995sensu Parker & Irmis Reference Parker and Irmis2006
Machaeroprosopus Mehl, Reference Mehl1916
Synonyms.Arribasuchus Long & Murry, Reference Long and Murry1995; Pseudopalatus Mehl, Reference Mehl1928; Redondasaurus Hunt & Lucas, Reference Hunt, Lucas, Lucas and Morales1993.
Type species.Belodon buceros Cope, Reference Cope1881.
Diagnosis. Pseudopalatinae that are tentatively distinguished from the other pseudopalatine phytosaur genera Nicrosaurus and Mystriosuchus by several derived character states: ventrally convex suture of the maxilla with the premaxilla and nasal (also in Leptosuchus and Smilosuchus ssp.); exposure of supratemporal fenestra on skull roof reduced to narrow slit or entirely closed (also in Angistorhinopsis ruetimeyeri (Huene Reference Huene1911): Huene Reference Huene1922); distinct subsidiary opisthotic process of squamosal (also in Pravusuchus Stocker, Reference Stocker2010); lateral corner of posttemporal fenestra formed by squamosal, rather than by union of squamosal and paroccipital process; base of paroccipital process of opisthotic expands posteriorly in a sinuous curve, resulting in the exoccipital pillar being in extension of midline, rather than the posterior rim of the paroccipital process, and the posterior outline of the posttemporal fenestra being oblique rather than parallel to the axis of paroccipital process (also in Leptosuchus, Smilosuchus and Pravusuchus ssp.); basioccipital condyle receded under supraoccipital shelf and obscured in dorsal view (also in Leptosuchus, Smilosuchus and Pravusuchus ssp.); vomers on anterior half of interchoanal septum flat and broad, rather than narrow and sharp. The validity of these apomorphic characters as synapomorphies of Machaeroprosopus needs to be tested by a more comprehensive phylogenetic analysis.
Distribution. Restricted to the uppermost units of the Triassic of the southwestern USA (the stratigraphic nomenclature follows Parker & Martz (Reference Parker and Martz2011) for Arizona; Lucas (Reference Lucas1993, Reference Lucas, Dickins, Yang, Yin, Lucas and Acharyya1997) and Heckert & Lucas (Reference Heckert, Lucas, McCord and Boaz2000) for New Mexico; and Lehman (Reference Lehman, Ahlen, Peterson and Bowsher1994a) for Texas): Upper part of the Sonsela Member (Parker & Irmis Reference Parker and Irmis2006; Parker & Martz Reference Parker and Martz2011), Petrified Forest Member (Heckert & Lucas Reference Heckert, Lucas, Heckert and Lucas2002; Parker & Martz Reference Parker and Martz2011), and Owl Rock Member (Kirby Reference Kirby, Lucas and Hunt1989; Heckert & Lucas Reference Heckert, Lucas, McCord and Boaz2000; Parker & Martz Reference Parker and Martz2011) of the Chinle Formation of Arizona; Bull Canyon (Hunt Reference Hunt, Lucas and Ulmer-Scholle2001), Rock Point (Lucas & Hunt Reference Lucas, Hunt, Lucas, Kues, Williamson and Hunt1992), and Redonda (Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993) Formations of north-central and eastern New Mexico; Cooper Canyon Formation of West Texas (Lehman & Chatterjee Reference Lehman and Chatterjee2005; this study).
Machaeroprosopus lottorum sp. nov.
(Figs 2–5, 7, 11–15, 18, 20–23)
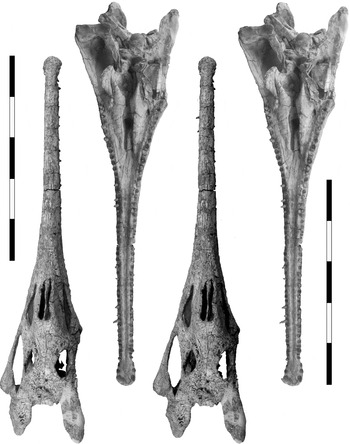
Figure 2 Stereopairs of the dorsal and ventral aspects of the skull of Machaeroprosopus lottorum TTU-P10076. Scale bars=100 mm.

Figure 3 Machaeroprosopus lottorum TTU-P10076: reconstruction of the skull in (A) ventral, (B) dorsal, (C) lateral and (D) occipital views. Scale bars=100 mm.

Figure 4 Stereopair of the dorsal aspect of the skull of Machaeroprosopus lottorum n. sp. TTU P-10077. Scale bar=100 mm.
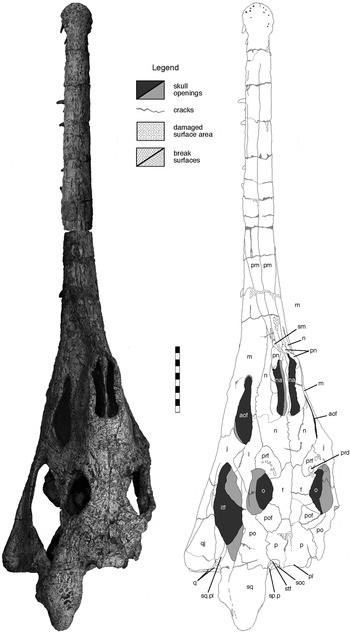
Figure 5 Machaeroprosopus lottorum TTU-P10076, skull in dorsal view. Scale bar=10 cm. Abbreviations: aof=antorbital fenestra; f=frontal; itf=infratemporal fenestra; j=jugal; l=lacrimal; m=maxilla; n=nasal; na=naris; o=orbit; p=parietal; pl=parietal ledge; pm=premaxilla; pn=paranasal; po=postorbital; pof=postfrontal; prd=preorbital depression; prf=prefrontal; q=quadrate; qj=quadratojugal; sm=septomaxilla; soc=supraoccipital; sp.p=squamosal process of parietal; sq=squamosal; sq.pl=squamosal platform; stf=supra-temporal fenestra.
Holotype. TTU-P10076, cranium.
Paratype. TTU-P10077, cranium, only referred specimen.
Type locality. TTU Vertebrate Paleontology Locality 3870 (exact locality data reposited at TTUP), 13 km South of Post, Garza County, Texas.
Type horizon. Upper part of the Cooper Canyon Formation, Dockum Group, Upper Triassic.
Age. Norian, Late Triassic.
Distribution. Restricted to type locality
Diagnosis.Machaeroprosopus lottorum is diagnosed by four characters which we interpret as autapomorphic for the taxon: (1) lateral rim of naris broad, flat, and rugose; (2) supratemporal fenestra fully closed in dorsal aspect, forming a shallow semicircular indentation into the skull roof (parietal, postorbital and squamosal) with strongly bevelled rim that continues onto the parietal; (3) free section of postorbital–squamosal bar (equalling in Machaeroprosopus the length of the dorsal surface of squamosal) short; (4) strongly developed horizontal medial laminae of palatines, that almost close the posterior section of the palatal vault in ventral view.
Differential diagnosis. In addition to the autapomorphic features, M. lottorum is distinguished from other species referred here to Machaeroprosopus by a number of characters that we assess as systematically important. The systematic validity of other variable features (see Table 1 and Appendix) is problematic because they differ in referred specimens from the type specimens (e.g., naris elevation: Camp Reference Camp1930; Zeigler et al. Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas, Heckert, Heckert and Lucasb, Reference Zeigler, Lucas and Heckert2003b) or they evidently vary within the hypodigms of other pseudopalatine taxa (see Hungerbühler Reference Hungerbühler1998). Machaeroprosopus lottorum differs from all species of Machaeroprosopus except M. gregorii by a comparatively broad dorsal surface of the squamosal; from both M. buceros (Cope Reference Cope1881), in which we include M. pristinus (Mehl Reference Mehl1928) and M. tenuis Camp Reference Camp1930, and from M. mccauleyi (Ballew Reference Ballew, Lucas and Hunt1989) by (1) the absence of a prenarial groove; (2) the lateral ridge of the squamosal being absent on the posterior process i.e., the dorsal and lateral surfaces of the process grade into each other; (3) a short and low rather than long and extensive flange of the squamosal; and (4) a rectangular rather than round top of the parietal–supraoccipital complex; in addition, from M. mccauleyi by (1) a shorter antorbital fenestra; (2) a parietal ledge that is twice as wide; and lateral walls of the parietal–supraoccipital complex that are (3) thick-based and (4) low; in addition, from M. buceros by a gently sloping rather than sinuous outline of the rostral crest in the morph with extended rostral crest; from both M. gregorii (Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993) and M. bermani (Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993) by a shorter and higher posttemporal fenestra; in addition, from M. gregorii by (1) a high posterior process of the squamosal with elevated dorsal surface; (2) a bulging lateral surface of this process; (3) a deep, rather than shallow supraoccipital shelf framed by squamosal processes of the parietals that are (4) sloping rather than vertical in their ventral section; and (5) the squamosal forming the lateroventral rim of the posttemporal fenestra by developing of a lamina onto the paroccipital process. M. lottorum differs from M. jablonskiae by having a lateral ridge on the squamosal, a knob-like tip on the squamosal, a rectangular outline of the parietal–supraoccipital complex, low lateral walls of the parietal–supraoccipital complex, and a beveled rim of the supratemporal fenestra that continues onto the parietal. A distinction from M. andersoni Mehl, 1922 is more difficult because of the incompleteness of the holotype of this species, but M. andersoni does not show the autapomorphic characters 1, 2, and 4 of M. lottorum, possesses a rounded, un-crested palatal ridge, and the maxillary furrow is absent.
Table 1 Character-taxon matrix used in the analysis of the phylogenetic position of Machaeroprosopus lottorum and TTU P10074. 0=primitive character state; 1, 2, 3=derived character states; A=character states 0 or 1; B=character states 0 and 1; ?=missing data.

Etymology. In honur of John Lott and Patricia Lott Kirkpatrick for their continuous support of this project.
2.1. Material and preservation
2.1.1. TTU-P10076 (Figs 2, 3)
Complete, 945 mm-long, slender-snouted and gracile cranium, including the most complete and informative dentition of a specimen of Machaeroprosopus known to date. The specimen was found lying on the left side and slightly tilted onto the skull roof, with the right posterior portion of the postorbital region broken up and scattered down the hill slope. Compaction resulted in a shearing of the skull to the right side, which affected in particular the narial region and the right postorbital region. The left side of the skull is thus flattened and appears more extended visually, the right side is verticalised. The right paroccipital process is crushed and was pushed dorsally. The right postorbital region was not reassembled with the cranium until after the description, to allow easy access to the braincase; it is not included in the figures. The cranium including the dentition is entirely freed of matrix.
2.1.2. TTU-P10077 (Fig. 4)
Almost complete, 1040 mm-long, robust cranium with a massive snout (altirostral sensu Hunt Reference Hunt, Lucas and Hunt1989, Reference Hunt1994). The skull was found dorsal side up and the occipital aspect partially was exposed, which explains the loss of the left squamosal and parts of the paroccipital processes and quadrates. The posterior portion of the narial region and the skull roof are crushed, and the entire skull is sheared to the right side. The skull deck was removed in pieces, prepared, and reassembled to its original shape. Similarly, the articular parts of both quadrates are kept separate from the skull. With exception of the rostrum, the skull is in parts strongly fragmented and the fragments are friable, the damage having been enhanced by root penetration. The fragments had to be fixed in their present distorted position before the matrix could be removed from the dorsal surface. On the ventral side, both tooth rows were fully exposed, but the palatal area is only prepared in broad outlines.
Machaeroprosopus sp.
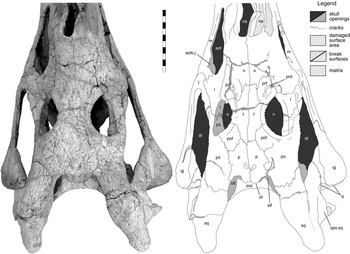
Figure 6 Machaeroprosopus sp. TTU-P10074, postnarial part of the skull in dorsal view. Scale bar=10 cm. Abbreviations: aof=antorbital fenestra; aofo.j=jugular antorbital fossa; f=frontal; itf=infratemporal fenestra; j=jugal; l=lacrimal; m=maxilla; n=nasal; na=naris; o=orbit; opo.sq=opisthotic process of squamosal; p=parietal; pl=parietal ledge; po=postorbital; pof=postfrontal; prd=preorbital depression; prf=prefrontal; q=quadrate; qj=quadratojugal; soc=supraoccipital; sq=squamosal; stf=supra-temporal fenestra.
2.1.3. TTU-P10074
Almost undistorted slender-snouted cranium (dolichorostral sensu Hunt Reference Hunt, Lucas and Morales1993, Reference Hunt1994), lacking the anteriormost section of the rostrum because of surface exposure. A section of a right premaxilla including the alveoli pm4 (4th premaxillary alveolus) through pm6 was recovered later at the site and most likely belongs to this specimen. The cranium is entirely freed of matrix. The length is 815 mm from the anterior break of the premaxillae to the tip of the sqamosals; the total length is estimated to be 900 mm.
2.2. Description of Machaeroprosopus lottorum sp. nov.
The description of the cranial anatomy of Machaeroprosopus lottorum focuses on TTU-P10076, one of the best preserved skulls of this genus available to date. Additional information from TTU-P10077 is inserted at the appropriate places. The shape of a phytosaur skull traditionally is the more important part of its anatomy for comparative and phylogenetic purposes, and most previous studies focused on characters of the general skull morphology rather than on osteological details. For this reason, we feel justified in giving separate accounts of the external skull morphology, the major skull openings and important external structures that extend over several cranial elements. Structures that are restricted to single skull elements are described with the individual bones.
2.3. External skull morphology
2.3.1. Rostrum
In phytosaurs, the prenarial area includes the rostrum with the highly variable prenarial crest, if present. The area is shaped differently in the two specimens of M. lottorum, and both differ from that in TTU-P10074. In the slender-snouted (dolichorostral) TTU-P10076 (Fig. 5) and TTU-P10074 (Fig. 6), the rostrum is wider than high, flattened semicircular in outline, and narrows only insignificantly in anterior direction in the premaxillar section. The rostrum is almost imperceptibly curved upwards. In dorsal view, the outline is triangular with concave ventral rims of the maxillaries. A slight constriction marks the articulation of the premaxilla with the maxilla on the rostrum.
In TTU-P10076 (Fig. 7), the prenarial area drops steeply over a distance of 35 mm at an angle of 75° from the nares. A kink in the profile marks the point where the slope starts to level out in a gradual steady way, being level at 210 mm in front of the nares. The posterior one fourth of the slope is rounded, but narrow in cross-section, and visually demarcated from the ventral part of the rostrum by two lateral depressions below the narial cone. Anteriorly, the top surface increasingly grades into the rostrum, but is still distinguished visually by narrow extensions of this depression. It is only in the anterior one fourth that the slope merges with the rostrum to a semicircular cross-section as in TTU-P10074.

Figure 7 Machaeroprosopus lottorum TTU-P10076, skull in left lateral view. Scale bar=10 cm. Abbreviations: aof=antorbital fenestra; ect=ectopterygoid; f=frontal; itf=infratemporal fenestra; j=jugal; l=lacrimal; lsp=laterosphenoid; m=maxilla; n=nasal; n(r)=right nasal; opo.sq=opisthotic process of squamosal; pl=parietal ledge; pm=premaxilla; pn=paranasal; pn(l)=left paranasal; pn(r)=right paranasal; po=postorbital; pof=postfrontal; pop.opo=paroccipital process of opisthotic; pp.sq=posterior process of squamosal; prf=prefrontal; pt=pterygoid; q=quadrate; qj=quadratojugal; sm=septomaxilla; sm(l)=left septomaxilla; sm(r)=right septomaxilla; sopo.sq=subsidiary opisthotic process of squamosal; sq=squamosal; sq.pl=squamosal platform.
The left side of the prenarial area of TTU-P10077 (Fig. 4) is dislocated ventrally for 12 mm in front of the nares, but is undistorted otherwise. It slopes from the elevated anterior rim of the naris, situated 205 mm above the alveolar plane, at the gentle angle of 20° in a slightly undulating line to about the midpoint of the rostrum. Here, at a height of 80 mm, the slope changes fairly abruptly to continue at 3–4° to a point 140 mm in front of the tip of the rostrum, where the now 50 mm-high crest ends in a step. In the anterior half of the crest, the flanks are constricted by two large, triangular depressions, which separate the vertical top of the crest from the steeply sloping alveolar portions of the maxilla and premaxilla. The outline of the depression corresponds exactly with the prenarial section of the nasal, and is characterised furthermore by the deepest sculpture of the entire skull. In the anterior half, the crest grades into the steeply sloping flanks of the rostrum, resulting in a triangular cross-section. The top of the crest is narrow and sharp in front of the nares, but expands and becomes more rounded anteriorly.
2.3.2. Infraorbital area
The area just ventrally to the orbits, formed by the orbital process of the postorbital, the posterior section of the lacrimal behind the preorbital ridge and the orbital process of the jugal and leading into the preinfratemporal shelf, is distinguished in TTU-P10077 from the surrounding surface by the different surface sculpture. Most of the area is smooth, and forms a conspicuous strip set deeper than the flank of the naris and the preorbital area. A distinct feature is a narrow and deep groove along the rim of the infratemporal fenestra. In TTU-P10077, the posterior rim of the groove is drawn out to an extensive, laterally projecting flange. The groove is bounded posteriorly by a series of three ridges, which extend oblique to the long axis of the groove. Each of the ridges is separated by a distinct groove running into the infratemporal fenestra. TTU-P10076 lacks a differentiated infraorbital area (Fig. 7) and shows only a very faint groove.
2.3.3. Parietal–supraoccipital complex. (Supraoccipital shelf, Figs 9, 10)
In posterior view, the shape of the parietal–supraoccipital complex in both TTU-P10076 and TTU-P10077 is rectangular, with a broad, horizontal parietal ledge. The lateral wall of the supraoccipital shelf, formed by the descending squamosal processes of the parietals, are vertical and, only in the ventral third of the complex they turn to the horizontal in a posterolateral direction in a gentle, steady curve. The supraoccipital shelf, including the supraoccipital and two lamella of the parietals, slopes continuously downward and posteriorly at an angle of 45°; the shelf is thus an inclined plane, neither curved and levelling out terminally as in Nicrosaurus and most specimens of Machaeroprosopus, nor vertical as in Mystriosuchus, Machaeroprosopus bermani and Machaeroprosopus gregorii.
2.3.4. Palate. (Fig. 11.)
On the ventral surface of the rostrum of TTU-P10076, the raised alveolar ridges enclose a flat, wide interpremaxillary groove. About 50 mm anterior to the choana, the surface of the groove widens gradually and deepens to form the rounded concave prechoanal area. In the centre of the prechoanal area, the prechoanal section of the vomers is raised slightly to a broad elevated structure. The alveolar ridge parallels the tooth row medially, so closely that, in parts of the premaxilla, a concave indentation is formed opposite each alveolus. The ridge is asymmetrical, with vertical lateral and sloping medial rims. On the premaxilla and the anterior part of the maxilla, the ridge is broad, reaching a maximum width of 15 mm opposite maxillary alveolus 9 (m9). Here, an outward kink in the anterior third of the maxillary tooth row (opposite m12) effectively splits the alveolar ridge in two structures. The alveolar ridge continues along the diverging tooth rows, standing out as a much more prominent structure on the palate because of its now much diminished width. The height of the alveolar ridge also decreases rapidly, and it merges with the palatal plane in the posterior quarter of the maxilla (roughly opposite m19). A second sharp, but narrow and low ridge, on or parallel to the premaxilla–maxilla suture splits from the alveolar ridge and runs straight across the slope of the prechoanal area, converges slightly with the midline and disappears towards the raised prechoanal area.
The choanal and postchoanal palate is subdivided into two tiers: the lower tier of the horizontal palatal plane (including the palatine and the palatal parts of maxilla and ectopterygoid); and the dorsally arched, higher tier, which includes the choana (the choanal process of the maxilla, the interchoanal septum i.e., the anterior part of the vomers and the vertical dorsal ramus of the palatine), followed by the palatal vault (the posterior part of the vomer and the arched anterior ramus of the pterygoid).
The palatal plane is morphologically dominated by two elevations: the fading alveolar ridge and the raised ridge on the centre of the palatine. The palatine ridge extends anteriorly into the convex round, tapering tip of the element, and posteriorly into the overhang over the palatal vault to merge with the medial side of the pterygoid flange. These elevations frame an elongated depression, in which lies the suborbital fenestra. The midline of the semi-cylindrical palatal vault is elevated for 50 mm above the crest of the palatal ridge. The opening between the two halves of the palatal plane is 22–26 mm wide below the choana and the anterior half of the palatal vault. Because of the extensive overhang of the palatines, the palatal vault is actually much wider (60 mm). Posteriorly, the exposure of the palatal vault is constricted severely in TTU-P10076. The width increases posteriorly to 65 mm because of the divergence of the palatines. With the emergence of the pterygoid flange (the ventral ramus of pterygoid and ectopterygoid, plus the posterior tip of the palatine) out of the walls of the palatal vault and the horizontal overhang of the palatine, both horizontal tiers merge into a unified vertical structure.
2.4. Skull openings
2.4.1. Nares and associated structures
The nares of all specimens are considerably elevated above the centre of the skull roof (for 15 mm in TTU-P10076). The septomaxilla forms most of the internarial septum and the anteromedial rim of the naris; an additional bony element, named here the paranasal, occupies the anterolateral border to a variable extent (see below), and the nasal forms most of the lateral and the entire posterior rim.
The subrectangular naris of TTU-P10076 (Fig. 12C) is long (83–85 mm), but narrow, reaching a maximum width of 15 mm close to the posterior border. It extends about 10 mm anterior to the level of the anterior rim of the antorbital fenestra. Anteriorly, the naris is indented as a distinct, 20 mm-deep and 5 mm-broad, V-shaped outlet (Fig. 12C, ‘nou’). The anteriormost section of the lateral rim is thin and sharp. The lateral rim is straight and slopes slightly in anterior direction, the anterior rim being 11 mm lower than the posterior. In both TTU-P10076 and TTU-P10077, most of the rim is squared in dorsal aspect and deeply rugose by predominantly transverse, irregular grooves, suggesting the presence of a cartilaginous structure (Fig. 12). The narial cone seems to be slightly constricted at midlength, and shows two symmetrical, flat and broad depressions below the anterior third, which enhances the cone-like appearance of the narial bulb. The internasal septum is thin, posteriorly with a sharp edge that grades into a smoothly rounded but still narrow rim anteriorly. The dorsal rim is straight throughout and situated a few mm below the lateral rims of the nares.
A raised central prenarial hump anterior to the nares, including parts of the internarial septum, or a pair of paramedian prenarial grooves extending onto the rostrum out of the narial outlets, are absent in M. lottorum and TTU-P10074.
2.4.2. Orbit and preorbital ridge
The orbit is surrounded by the prefrontal anterodorsally, the frontal dorsally, the postfrontal posterodorsally, the postorbital posteriorly, and the lacrimal ventrally (Fig. 13). The jugal is excluded from the orbit in both specimens of M. lottorum and TTU-P10074. The undeformed orbit is oval, with a conspicuous straight and flat ventral rim and a long axis directed anteroposteriorly (Fig. 2). The plane circumscribed by the opening faces laterally and, for about 20 to 30 degrees, upward; the orbits are not turned anteriorly to any degree, and thus there is no indication of binocular vision. With to exception of the broad and smooth ventral rim, the orbital rim is sharp and raised above the surrounding skull surface, forming a conspicuous, 10 mm-broad, less sculptured zone around the opening. The elevation is particularly strong at the posterodorsal corner, in the form of an extensive hump, especially pronounced in TTU-P10074, and along the anterior rim. This anterior orbital ridge causes the preorbital depression (Fig. 6, ‘prd’), here, in contrast to the pseudopalatine phytosaur Mystriosuchus (GPIT 261/001; Hungerbühler Reference Hungerbühler2002), actually a series of smaller, deep pits separated by bridges of the surface sculpture. The ridge continues ventrally to the anterior corner of the antorbital fenestra onto the lacrimal, where it marks the anterior border of the less sculptured infraorbital area.
2.4.3. Antorbital fenestra and antorbital fossa
In TTU-P10076, the antorbital fenestra is surrounded anterodorsally by the dorsal, and ventrally by the ventral processes of the maxilla, the lacrimal posterodorsally, and the jugal posteriorly and posteroventrally (Figs 7, 12A, B, 13). The opening is oval, with an angular rather than round posterior corner. In the undeformed openings, the long axis, which is positioned at a low angle with the axis of the skull, measures 106 mm, the short axis 43·5 mm; the size is comparatively small for phytosaurs, with reference to naris length. The ventral edge is prominent and sharp, whereas the dorsal rim is broadly rounded and deep. TTU-P10076 (as is Machaeroprosopus sp. TTU-P10074) is among the few specimens of pseudopalatine phytosaurs that show remnants of an antorbital fossa. The anterodorsal part of the fossa appears as a small depression on the lateral face of the maxilla (Figs 7, 8, 12A; ‘aofo.ml’). Further posteriorly, the dorsal process of the maxilla and the anterior process of the lacrimal are folded over, forming the broad dorsal rim of the antorbital fenestra, but continue as a steeply medioventrally sloping plane, a 15 mm-deep maxillo–lacrimal antorbital fossa as overhang over the antorbital fenestra proper (Fig. 12A, C). In addition, there is a distinct jugular fossa in the posteroventral corner extending along the posterior rim of the antorbital opening, in the form of a narrow, thin lamella medial to the lacrimo–jugular rim of the antorbital fenestra, both of which enclose a deep, V-shaped recess (Fig. 12C, ‘aofo.j).

Figure 8 Machaeroprosopus sp. TTU-P10074, postnarial part of the skull in left lateral view. Scale bar=10 cm. Abbreviations: aof=antorbital fenestra; aofo.ml=maxillo-lacrimal antorbital fossa; ect=ectopterygoid; f=frontal; itf=infratemporal fenestra; j=jugal; l=lacrimal; m=maxilla; n=nasal; opo=opisthotic; opo.sq=opisthotic process of squamosal; pm=premaxilla; pn=paranasal; po=postorbital; pof = postfrontal; pp.sq = posterior process of squamosal; ppr.sq=parietal process of sqamosal; prf=prefrontal; pt=pterygoid; q=quadrate; qj=quadratojugal; sm=septomaxilla; sm(l)=left septomaxilla; sm(r)=right septomaxilla; sq=squamosal.
2.4.4. Supratemporal fenestra
The supratemporal fenestra is bounded by the parietal anteriorly, the postorbital and the medial lamella of the squamosal laterally, the parietal ledge medially, and the sloping squamosal process of the parietal and the horizontal parietal process of the squamosal posteriorly (Fig. 13). Because of the elongated squamosal, the posterior rim is actually positioned posterolaterally, and it is depressed on the undeformed right side for 36–42 mm below the level of the skull roof, or for about 30% of the skull height. In all specimens, the anterior rim of the fenestra is shallow, but broad compared to other species previously referred to Pseudopalatus. It forms a semicircular, 13 mm-deep excavation in TTU-P10076 (Fig. 5) that hardly indents the skull roof proper, i.e., the parietal. The excavation is, rather, caused by the projection of the parietal ledge to the rear. In both TTU-P10076 (Figs 5, 9) and TTU-P10077, the rim is distinctly bevelled, the ventral edge almost spanning the entire indentation. Thus, the supratemporal opening is in M. lottorum and TTU-P10074 effectively closed on the skull roof, and all three specimens are more derived than the deeper, slit-like, straight or medially curved openings seen in the type specimens of M. buceros, M. mccauleyi, M. jablonskiae and M. andersoni. The maximum length of the opening to the posterior extent of the paroccipital process is 85 mm, of which the posterior 50 mm are completely recessed below the horizontal surface of the squamosal (i.e., under the dorsal sulcus; Figs 9, 10) and are not visible in dorsal view. The supratemporal fenestra opens thus almost exclusively to the rear.

Figure 9 Machaeroprosopus lottorum TTU-P10076, skull in occipital view. Scale bar = 50 mm. Abbreviations: boc = basioccipital; bpt.bsp = basipterygoid process of basisphenoid; bpt.pt = basipterygoid process of pterygoid; bsp = basisphenoidal part of parabasisphenoid; cr = central ridge of quadrate; ds = dorsal sulcus; ect = ectopterygoid; eoc = exoccipital; fl.sq = squamosal flange; fm = foramen magnum; f.pat = facet for proatlas; fq = quadrate foramen; itf = infratemporal fenestra; j = jugal; la.p = lamina of parietal; lc = lateral condyle of quadrate; m = maxilla; mc = medial condyle of quadrate; nc = narial cone; opo.sq = opisthotic process of squamosal; p = parietal; pl = parietal ledge; po = postorbital; pop.opo = paroccipital process of opisthotic; pp.sq = posterior process of squamosal; ppr.sq = parietal process of squamosal; pro = prootic; ptf = posttemporal fenestra; ptof = pteroccipital fenestra; ptr.q = pterygoid ramus of quadrate; q = quadrate; qj = quadratojugal; qr.pt = quadrate ramus of pterygoid; r = recess; soc = supraoccipital; sopo.sq = subsidiary opisthotic process of squamosal; sp.p = squamosal process of parietal; stf = supra-temporal fenestra; vr.pt = ventral ramus of pterygoid; vs = ventral sulcus.

Figure 10 Machaeroprosopus sp. TTU-P10074, skull in occipital view. Scale bar = 10 mm. Abbreviations: boc = basioccipital; bpt.bsp = basipterygoid process of basisphenoid; cr = central ridge of quadrate; eoc = exoccipital; fl.sq = squamosal flange; fm = foramen magnum; fq = quadrate foramen; la.p = lamina of parietal; lc = lateral condyle of quadrate; l.sq = lamella of squamosal onto paroccipital process of opisthotic; mc = medial condyle of quadrate; opo.sq = opisthotic process of squamosal; p = parietal; po = postorbital; pop.opo = paroccipital process of opisthotic; pp.sq = posterior process of squamosal; ppr.sq = parietal process of squamosal; ptf = posttemporal fenestra; ptof = pteroccipital fenestra; ptr.q = pterygoid ramus of quadrate; q = quadrate; qj = quadratojugal; qr.pt = quadrate ramus of pterygoid; soc = supraoccipital; sopo.sq = subsidiary opisthotic process of squamosal; stf = supra-temporal fenestra; vr.pt = ventral ramus of pterygoid; vs = ventral sulcus.
2.4.5. Infratemporal fenestra
The infratemporal fenestra is bounded by, clockwise from the orbit, the postorbital, squamosal, quadratojugal and jugal (Fig. 13). The opening roughly forms a parallelogram with a concave dorsal rim, the anterodorsal corner being located below the centre of the orbit. The fenestra extends far forward in TTU-P10077 (and TTU-P10074; Fig. 8), the anteroventral corner being 10 mm in front of the anterior orbital rim, but terminates level with the orbit in TTU-P10076 (Fig. 7).
2.4.6. Posttemporal fenestra
The posttemporal fenestra of TTU-P10076 is short and oval, with a flat dorsal rim, 27 mm by 11 mm wide (Fig. 9). The opisthotic–squamosal suture is located in the angular medial corner of the fenestra. The parietal process of the squamosal forms the entire dorsal rim of the posttemporal fenestra, the paroccipital process of the opisthotic two thirds and an extension of the squamosal the lateral third of the concave ventral rim. The fenestra is set somewhat oblique with respect to the axis of the paroccipital process: the lateral corner is located deeper, and the lateral section of the fenestra is thus strongly roofed over by the parietal process of the squamosal. The rounded lateral rim grades smoothly into the ventral sulcus.
2.4.7. Foramen magnum. (Fig. 9.)
The foramen magnum is a transversely oval opening (TTU-P10076: 21 mm by 15·5 mm). The supraoccipital shelf overhangs the foramen magnum by 15 mm in both specimens. The condylar parts of the exoccipitals extend posteriorly, resulting in a concave posterior edge of the exoccipital pillars, and thus forms a distinct, slightly concave platform in front of the foramen.
2.4.8. Choana. (Fig. 11.)
The undistorted subrectangular left choana is 73 mm in TTU-P10076, and thus subequal in length with the naris. The posterior rim is exactly below the posterior rim of the naris, but the anterior rim extends further forward. Because of the vaulting of the prechoanal palate, the anterolateral rim of the choana formed by the maxilla is curved upward and, at the same time, widens in a broad lateral recess, being slightly overhung by the anterior section of the palatine. In the posterior two thirds, an extensive vertical lamella of the palatine forms a lateral wall. The palatine and the vomers also form the posterior rim. The interchoanal septum formed by the vomers is a 9–11 mm-broad, flat or slightly convex ventral plane, the ascending medial walls of the choana converge at each other and the septum becomes much narrower internally. At midlength, the septum narrow considerably and curves upward in a step-like fashion to level out on the deeper level of the palatal vault. Thus, the choana expands from 9 mm to 15 mm in width, and the plane of the opening is bent in the vertical plane.
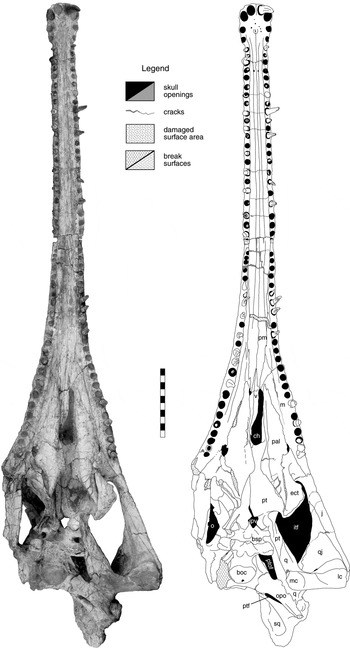
Figure 11 Machaeroprosopus lottorum TTU-P10076, skull in ventral view. Scale bar=10 cm. Abbreviations: boc=basioccipital; bsp=basisphenoidal part of parabasisphenoid; ch=choana; ect=ectopterygoid; ipv=interpterygoid vacuity; itf=infratemporal fenestra; j=jugal; lc=lateral condyle of quadrate; m=maxilla; mc=medial condyle of quadrate; o=orbit; opo=opisthotic; pal=palatine; pm=premaxilla; pt=pterygoid; ptf=posttemporal fenestra; ptof=pteroccipital fenestra; q=quadrate; qj=quadratojugal; sq=squamosal; v=vomer.
2.4.9. Suborbital fenestra
In TTU-P10076, the area of the suborbital fenestra is fractured on the right side and the left side is cracked and the palatine telescoped underneath the palatal section of the maxilla. The suborbital fenestrae are well-delineated (Fig. 11). The suborbital fenestra lies on the palatal plane between the maxilla and the palatine, about 10 mm behind the choana (Fig. 14, ‘sf’). The opening is reduced compared to that of non-pseudopalatine phytosaurs, and appears as a narrow oval fenestra 17 mm by 7 mm wide, with pointed anterior and posterior corners. The maxilla forms the anterior and the ectopterygoid the posterior half of the lateral rim, while the palatine borders the opening laterally.
2.4.10. Subtemporal fenestra. (Fig. 11)
According to the less deformed right side, the subtemporal fenestra of TTU-P10076 is the large ventral, anteroposteriorly elongated opening of the adductor chamber. The shape is subrectangular with an oblique, posteromedially trailing margin and posteriorly slightly converging walls. It is bordered anteriorly by the broadly rounded palatal part of the ectopterygoid, and medially by the ventral surface of the pterygoid–quadrate plate (predominantly the pterygoid ramus of the quadrate). The quadrate forms the posterior margin and the jugal the lateral margin, with the quadratojugal contributing to the posterolateral corner.
2.4.11. Interpterygoid vacuity
The interpterygoid vacuity is reduced to a roughly 35 mm-long, roughly heart-shaped opening (Fig. 14). On the roof of the palatal vault, the vacuity indents the pterygoids in a 10-mm long, tapering slit. The concave lateral rims are formed by the basipterygoid processes of the pterygoids. The notch between the basipterygoid processes of the parabasisphenoid marks the posterior limit.
2.4.12. Pteroccipital fenestra. (Fig. 9)
The pteroccipital fenestra is a narrow, predominantly ventrally- and somewhat posteriorly-facing passage between the pterygoid–quadrate plate and the braincase. It is crescentic in outline, with straight lateral and convex medial rims. As an estimate, the length is 70 mm and the maximum width is 20 mm, although the latter is somewhat exaggerated by deformation in all specimens. The passage is framed laterally by the dorsal rim of the pterygoid–quadrate plate, anteriorly by the lateral edge of the basipterygoid process of the parabasisphenoid, and medially by the anterior wall of the stapedial groove and the base of the paroccipital process.
2.5. Individual elements
2.5.1. Premaxilla
The elongated subrectangular terminal rosette of the premaxillae is longer than wide, terminating by a slight constriction (Fig. 11). It is only moderately expanded laterally, being 47 mm wide, with a width of the rostrum of 31 mm at the constriction. The premaxillae curve ventrally to a moderate degree in front of pm3, the tip being situated 24 mm below the level of the alveolar plane (Fig. 7). In anterior view, the total height of the terminal rosette is subequal to the width (44·5 mm). The slender rostrum widens gradually from 33 mm at pm5 to 46 mm at the end of the premaxillae; there is an almost imperceptible lateral convexity along the last five premaxillary alveoli.
The suture between the premaxillae is present as a groove on the anterior surface of the rosette and along most of the dorsal surface of the rostrum, but the bones are fused without an external trace over the distance of 90 mm in the anterior section above pm3 to pm9 (Fig. 5). A well developed longitudinal groove with a rectangular cross-section is present on the lateral surface of the rostrum between pm10 and pm18, about 10 mm above the alveolar plane (Fig. 7). Anteriorly and posteriorly, the groove is dissociated into an array of short grooves or depressions. In a somewhat irregular pattern, foramina in the grooves and depressions enter into the rostrum, on average at every second tooth position. Some foramina take on the shape of fairly long slits. Because of the dorsal expansion of the maxillae, the posterior section of the premaxillae is developed as a narrow extension on the top of the rostrum. Posteriorly, each element splits into two tapering processes: a broader and shorter medial process along the median plane that is wedged for 13 mm between the septomaxillae, and a larger, narrower process that extends for about 20 mm on the lateral side of the crest between the septomaxilla and the nasal (Fig. 12A, C). The opposite is the case in TTU-P10077 (Fig. 15). The dorsal (medial) process is narrower, non-tapering, and 37 mm long, of which 20 mm are represented by a thin prong along the midline. The stout ventral (lateral) process is twice as broad, subequal in length, extends horizontally below the septomaxilla, and forms an anteroventrally sloping suture with the nasal. Moreover, a deep and narrow furrow, in parts resembling an almost closed channel, extends for 75 mm on both sides on the premaxilla, in extension of the ventral septomaxilla–premaxilla contact. Such a groove has not been identified before in, and is not homologue with, the prenarial groove of other phytosaurs, which extends out of the nares on the septomaxillae and nasals.

Figure 12 Machaeroprosopus lottorum TTU-P10076: details of the narial area in (A) lateral left view; (B) lateral right view; (C) anterolateral left view. Scale bars=50 mm. Abbreviations: aof=antorbital fenestra; aofo.j = jugular antorbital fossa; aofo.ml=maxillo–lacrimal antorbital fossa; f=frontal; f.pn=facet for paranasal; g=groove; itf=infratemporal fenestra; j=jugal; l=lacrimal; m=maxilla; n=nasal; nou=narial outlet; o=orbit; pis=preinfratemporal shelf; pm=premaxilla; pn=paranasal; po=postorbital; prd=preorbital depression; prf=prefrontal; sm=septomaxilla.
The interpremaxillary groove (Fig. 11), starting between both pm5, is rounded in cross-section anteriorly, but rapidly develops into a flat-bottomed channel posteriorly. The unpaired foramen incisivum is located on a central mound between pm3 and pm4. Anteriorly and laterally, there is a series of three (left) and two (right) nutritious foramina. A conspicuous pair of foramina that open posteriorly is situated in the bottom of the interpremaxillary groove at the level of pm8 (Fig. 11). The palatal surface of the premaxilla extends into a thin, tapering palatal process that runs on the maxilla (illustrated in TTU-P10074, Fig. 16A, ‘pap.pm’) and reaches the tip of the choana, overlapping the vomer medially (Fig. 11).
2.5.2. Maxilla
The maxilla appears on the lateral surface, forming two sharp, anteriorly-pointing, subequally long prongs that interdigitate with the premaxilla (Fig. 7). Posteriorly, the suture curves more and more dorsally, until the maxilla extends over two thirds of the height of the prenarial area and levels out into the narrow dorsal process above the antorbital fenestra, firmly sutured by a strongly serrated suture with the nasal. In TTU-P10077 (Fig. 15), the maxilla bulges distinctly along the posterior part of the premaxilla and the nasal in a strongly convex suture. This section of the element is extremely thin, being 2 mm thick medio-laterally in TTU-P10074 (Fig. 16A). The base of the dorsal process is marked by a concavity of the nasal-maxillary suture, and the process continues with a restricted width of 11 mm. TTU-P10077 (and Machaeroprosopus sp. TTU-P10074) show a peculiar sturdy prong of the maxilla into the premaxilla in front of the apex of the suture. The prong is absent in TTU-P10076, and the nasal–maxillary suture is almost straight (Fig. 12). The surface of the much higher ventral process is vertical and extends far posteriorly, forming two thirds of the antorbital rim, and meets the jugal in a subvertical suture, the ventral section of which trails obliquely toward the jugal notch.
On the ventral surface, the suture between maxilla and premaxilla appears laterally to alveolus m1, curves closely in front around the alveolus and extends posteriorly in the lateral edge of the alveolar ridge (Fig. 11). It crosses the ridge obliquely opposite to alveoli m4 to m12 and heads obliquely on a ridge over the prechoanal depression, straight towards the anteromedial corner of the choana. This choanal process extends, sloping to the midline and posteriorly because of the concavity of the prechoanal palate, between the tip of the palatine laterally and the palatal process of the premaxilla and the vomer medially. It forms the anterior rim of the choana, and continues below the palatine on the lateral side of the opening for 21 mm; i.e., for one fourth of the choana length. The contribution of the maxilla to the choanal rim is thick and rounded, and the anterolateral rim is concave, resulting in a broadened lobate expansion of the anterior part of the choana below the tip of the palatine. The suture with the palatine runs parallel to the alveolar rim in the elongate depression of the palatal plane to the suborbital fenestra. The maxilla continues along the base of the ectopterygoid, overlapping the bone, straight towards the anterolateral corner of the subtemporal fenestra, and ends in a blunt tongue lying ventrally on the ectopterygoid–jugal complex.

Figure 13 Machaeroprosopus lottorum sp. nov.: schematic reconstruction of the skull and the configuration of the cranial elements in (A) ventral, (B) dorsal, (C) lateral and (D) occipital views. Scale bars=100 mm.

Figure 14 Machaeroprosopus lottorum TTU-P10076: (A) right palate in ventral view; (B) with pterygoid flange removed, showing the steinkern of the sinus between ectopterygoid and palatine. Scale bars=50 mm. Abbreviations: ch=choana; ect=ectopterygoid; g=groove; ipv=interpterygoid vacuity; m=maxilla; pal=palatine; pt=pterygoid; sf=suborbital foramen; si=sinus; vr.ect=ventral ramus of ectopterygoid; vr.pt=ventral ramus of pterygoid.

Figure 15 Machaeroprosopus lottorum TTU P-10077, left anterial narial and prenarial region. In the interpretive drawing, the right side is not shown. Scale bar=50 mm. Abbreviations: m=maxilla; n=nasal; pm=premaxilla; pn=paranasal; sm=septomaxilla.

Figure 16 Machaeroprosopus sp. TTU-P10074: (A) cross-section through the rostrum 75 mm in front of the nares, view in anterior direction. Transverse break-surfaces unshaded, arrows indicate position of sutures on the outside; (B) counterpart of the right dorsal part of the same section to show the sutural configuration, view in posterior direction; (C) right prenarial area in lateral view. Scale bars = 10 mm. Abbreviations: ac = alveolar canal; ar = alveolar ridge; c.pm = premaxillary cavity; cv.m = maxillary cavern; cv.n = nasal cavern; m = maxilla; m12 = alveolus 12 of maxilla; m14 = alveolus 14 of maxilla; n = nasal; pap.pm = palatal process of premaxilla; pm = premaxilla; sm = septomaxilla; sm(l) = left septomaxilla; sm(r) = right septomaxilla.

Figure 17 Machaeroprosopus sp. TTU-P10074, cross-section through the rostrum 195 mm in front of the nares, view in anterior direction. Scale bar = 10 mm. Abbreviations: ac = alveolar canal; ar = alveolar ridge; c.pm = premaxillary cavity; ipmg = interpremaxillary groove; m = maxilla; m2 = alveolus 2 of maxilla; pm = premaxilla.
2.5.3. Septomaxilla
The sutural configuration of the immediate prenarial area is difficult to interpret in the slender-snouted TTU-P10076 because of fractures, intense superficial damage, the sculpturing and the lamellate interdigitation of the constituent elements. The septomaxilla forms much of the internarial septum and extends forwards, wedged between the paranasal, the nasal and the premaxilla.
The septomaxillae are asymmetrically developed on the internasal septum of TTU-P10076 (Fig. 12C). Thirty mm behind the anterior rim of the naris, the suture between the septomaxillae drops vertically on the right flank of the septum, which is, from this point backwards formed by the left septomaxilla alone. The suture with the nasal appears on the septum 27 mm in front of the posterior rim, running horizontally, and ascends to the dorsal surface right at the posterior rim of the opening.
In the anteriormost section of the naris, the septomaxilla forms a laterally concave flange, the floor of the narial outlet, and abuts against the paranasal. Externally, the septomaxilla extends as a thin strip forwards along the midline, apparently intensively covered by the paranasal. The element triples in width in front of the paranasal and seems to terminate 70 mm in front of the naris, at about the level of the anterior tip of the nasal (TTU-P10076, right side; Fig. 12C). Surface damage to the bone on the left side reveals several parallel suture lines (Fig. 12A, C) that mark the intensive interdigitation with the underlying premaxilla, as in Smilosuchus gregorii UCMP 27200 (Camp Reference Camp1930). On both sides, the sutures between the septomaxilla and the paranasal extend out of the naris through the narial outlets, separating two 4 mm narrow elements. The septomaxillae widen to 9 mm at 75 mm in front of the nares (gradually on the right side, abruptly at a point 40 mm in front of the naris on the left side), extending further forward than the nasal. The length of the septomaxilla of TTU-P10077 compares with that of TTU-P10076, but the element is much broader. It is still 14 mm wide at the anterior end, where it broadly interdigitates with the dorsal process of the premaxilla, and a bulge of the lateral rim down the flank of the rostral crest results in a maximum width of 28 mm.
2.5.4. Paranasal
An additional discrete skull element is outlined clearly in the anterior lateral corner of the nasal opening of TTU-P10076 (Fig. 12) and TTU-P10077 (Fig. 15). We refer to this element as the paranasal bone. It forms the thick, pillar-like anterior rim of the naris and the anteriormost part of the narial flank. The element is triangular in outline, with a blunt, ventrally pointing tip. The medial side lies on the septomaxilla and forms the lateral wall of the narial outlet. Laterally and posteriorly, it is overlapped by the nasal, forming a short squamal suture, as evidenced by the partially exposed sutural contact on the left side of TTU-P10076, and the oblique course over the dorsal rim of the naris (Fig. 12C). The posterior extent is restricted to the anterior one fourth of the side of the naris in TTU-P10076. In TTU-P10077, the nasal is detached from the paranasal along the posterior section of the suture line and crushed down, and extends externally along the anterior third of the naris. A cross-section of the narial rim reveals (Fig. 15, inset) that the nasal underlies the paranasal and reaches forward to a point 15 mm behind the anterior corner of the naris. This suggests that the extent of the paranasal may vary by expanding over the nasal. The element seems to be present also in TTU-P10074, according to a sigmoidal suture line on the left side that separate the anterior third of the lateral narial rim from the nasal (Fig. 8).
2.5.5. Nasal
The nasal is an extensive element that forms much of the interorbitonasal area and the narial cone. Anteriorly, the nasal is wedged as a thin, tapering prong between the convex maxilla and the posteriormost section of the premaxilla, terminating 75 mm in front of the naris (Fig. 7). The bone expands, however, much deeper internally below the premaxilla and the dorsal process of the maxilla, meets its counterpart along the midline, and forms much of the roof of the large antorbital cavity here (Fig. 12B); similarly, a thin wedge of the nasal is exposed in TTU-P10076 between the septomaxilla and the posterior lateral process of the premaxilla (Fig. 12A, C). In this region, the nasal contains a broad, dorsoventrally-compressed cavern that extends into the base of the premaxilla. The cavern extends posteriorly at least to a point 75 mm in front of the naris, and communicates with the antorbital cavity by means of narrow canals (Fig. 16A, B). The nasal forms the posterior three fourths of the lateral rim and the entire posterior rim of the naris, but seems to contribute little to the internasal septum. Posterior to the nares, the width of the nasal is restricted by the lacrimal, but in particular by the prefrontal posterolaterally to two processes that contact the frontal 10 mm in front of the orbit.
2.5.6. Frontal
Both frontals, are cross-shaped, as is typical for phytosaurs (Fig. 5). The subequally long anterior and posterior processes are wedged between the nasals and prefrontals, and the parietals and postfrontals, respectively. There is a remarkable left–right asymmetry in TTUP-P10076 (as is in TTU-P10074) regarding the length of these posterior processes. The orbital wing of each element expands laterally and dorsally to the centre of the raised dorsal rim of the orbit; both frontals thus enclose a depressed concave interorbital area. The interfrontal suture is located on a low and broad elevation along the midline of the skull.
2.5.7. Prefrontal
The prefrontal is about 1·5 times longer than the postfrontal. The element restricts the width of the posteriormost section of the nasal anteromedially and the anterior process of the frontal medially on the dorsal plane of the skull (Fig. 5), and extends onto the lateral side to form the anterodorsal and entire anterior rim of the orbit. At the level of the ventral rim of the orbit, the prefrontal meets the lacrimal; the suture is convex in TTU-P10076 (Fig. 7).
2.5.8. Lacrimal
The lacrimal is an elongated, but low element. The high anterior process forms the posterior section of the dorsal antorbital rim, including the inclined dorsal antorbital fossa (Fig. 12B). The process tapers slightly anteriorly to meet and interdigitate with the dorsal process of the maxilla at about midlength of the antorbital fenestra. The lacrimal is bounded dorsally by the posterior section of the nasal, and the prefrontal in a distinctly convex suture at the level of the ventral rim of the orbit, which results in a dorsally-directed prong between the nasal and the prefrontal. The lacrimal forms the anterior half (TTU-P10076) of the ventral orbital rim, and does not contribute to the anterior rim. Because of extensive fragmentation, no information is preserved regarding the opening of the lacrimal canal, which in phytosaurs is usually placed internally in the anteroventral corner of the orbit, at or close to the joint of lacrimal and prefrontal (Case Reference Case1929; Camp Reference Camp1930; Case & White Reference Case and White1934; Witmer Reference Witmer1997; Senter Reference Senter2002). The ventral process is absent in TTU-P10076 (Fig. 7), and does not contribute to the antorbital fenestra.
2.5.9. Jugal. (Fig. 7)
The maxillary process of the jugal is short and high. There is no jugal notch developed at the maxilla–jugal suture, but only a slight concavity of the ventral rim. The jugal contributes only to about one fourth of the ventral rim of the antorbital fenestra. The posteroventral antorbital fossa is exclusively in the jugal (Fig. 12C). The jugal forms here a thin lamina, 8 mm medial to the the posterior rim of the antorbital fenestra, that ascends parallel to the rim. The maximum width of the lamina is 7 mm. With the outer rim of the antorbital fenestra, it encloses a recess that leads into an ascending groove. A preinfratemporal shelf is present, but poorly defined in TTU-P10076. In the anteroventral corner of the infratemporal fenestra, the jugal holds a deep, posteriorly-facing oval recess enclosed by the bases of the quadratojugal process and the jugal process of the ectopterygoid. The orbital process is slender, and extends along the lacrimal in an ascending, slightly serrated suture to contact the postorbital about 25 mm below the orbit in a posterodorsally trailing suture. The jugal is thus excluded from the orbit. Posteromedially, the orbital process forms much of an extensive, 25 mm-broad medially-directed flange along the postorbital–jugal bar, and thus a deep, posteriorly-facing area developed as the anterior wall of the infratemporal fenestra. The jugal continues high up to about mid-height of the orbit, overlapping the postorbital in a broad tongue.
The quadratojugal process is low, being 24 mm high at its base, and only expands a little on the ventral side until it is enclosed, and topped, by the lateral and medial lamina of the quadratojugal. The ventral rim sharpens posteriorly and almost reaches the lateral condyle of the quadrate (Fig. 11). The dorsal rim is flattened and merges anteriorly with the area of the preinfratemporal shelf; posteriorly, the anterior process of the quadratojugal rides on the flattened surface of the jugal.
2.5.10. Postorbital
In dorsal view, the postorbital appears stout and truncated in comparison to other phytosaur taxa (Fig. 5). This is because of the short squamosal process of the postorbital that forms the anterolateral rim of the supratemporal fenestra. The process extends over only one fourth of the length of the postorbital–squamosal bar, which is mainly formed by the elongated squamosal. The postorbital reaches forwards along the parietal and the postfrontal to form the centre of the posterior rim of the orbit. The crescentic orbital process extending downward and forward is low, but strongly expanded transversally, being twice as broad as high. It is wedged as a thin but long prong between the lacrimal and the jugal (Fig. 7), rather than extending along the infratemporal fenestra and meeting the lacrimal anteriorly and the jugal ventrally. The postorbital also contributes little to the anterior rim of the infratemporal fenestra externally, although it extends downward on the medial rim of the postorbital flange to about mid-height of the infratemporal fenestra (Fig. 7).
2.5.11. Postfrontal
The postfrontal is the smallest element exposed on the skull roof (Fig. 5). It is almost semicircular in shape. The convex edge is surrounded, from front to back, by the frontal, the parietal and the postorbital, and the concave edge forms the posterodorsal rim of the orbit. A conspicuous broad hump on the postfrontal marks the highest point of the orbital rim (Fig. 8), and thus of the skull roof.
2.5.12. Parietal
The parietals extend forward to the level of the posterior rim of the orbits, forming a short transverse interdigitating suture with the frontals (Fig. 5). Anterolaterally, the parietal meets the postfrontal in a concave suture. Laterally, the bone is in contact with the postorbital, and it forms the entire anterior rim of the strongly reduced supratemporal fenestra posteriorly. In TTU-P10074 (left side, Fig. 6), it extends slightly farther posterior along the medial rim of the postorbital–squamosal bar. In the centre of the parietals, there are two oval depressions situated symmetrically along the midline, posterior to a marked central hump on the interparietal suture. In Mystriosuchus westphali GPIT 261/001 and Smilosuchus gregorii UCMP 27200, such an elevation marks a dome-like excavation of the internal side of the parietals that is confluent with the brain cavity (Camp Reference Camp1930; Hungerbühler Reference Hungerbühler1998). This structure has been interpreted as evidence for a pineal organ (Camp Reference Camp1930; Langston Reference Langston1949) or, alternatively, as the cartilaginous tips of the supraoccipital in the otherwise ossified skull roof (Roth & Roth Reference Roth, Roth, Thomas and Olson1980). The central skull roof terminates posteriorly with the conjoined parietal extensions that form a rectangular parietal ledge overhanging the supraoccipital shelf for 25 mm (Fig. 9). The corners of the ledge are extended into two posteriorly-pointing prongs, which are dorsoventrally flattened. In the cavity below the ledge, the parietals bear two symmetrical pits close to the midline, probably the origin of strong tendons of parts of the epaxial musculature (Anderson Reference Anderson1936). The corners of the cavity below the parietal ledge are deeply sculptured with ridges, protuberances and grooves, which provide further insertion points for musculature.
The squamosal processes of the parietals form almost vertically descending walls around the supraoccipital, then curve horizontally while diverging posterolaterally, and the pointed tips finally meet and overlap the parietal processes of the squamosals. An exception is the left side of TTU-P10077, in which the squamosal process of the parietal does not reach the squamosal on the dorsal face of the opisthotic process, but terminates in a broad end sutured to the supraoccipital (Fig. 20). The squamosal processes that form the lateral walls of the supraoccipital shelf and ride on the supraoccipital are low and stockily built, with a broad base that narrows rapidly to a sharp rim. They send a thin crescentic lamina onto the supraoccipital, both laminae together covering about one quarter of the surface of the shelf. The descending flange of the parietal anterior to the squamosal process extends forwards, forming about half of the lateral surface of the braincase, and meets the laterosphenoid, epiotic, and opisthotic in a straight oblique suture (Fig. 18).

Figure 18 Machaeroprosopus lottorum TTU-P10076, left side of the braincase. For details of the otic region, see Figure 24. Scale bar=50 mm. Abbreviations: cp.ps=cultriform process of parasphenoid; ept=epipterygoid; eptp.pt=epipterygoid process of pterygoid; g=groove; hypg=hypophyseal gap; pqg=palatoquadrate groove; ptr.q=pterygoid ramus of quadrate; qr.pt=quadrate ramus of pterygoid; V=trigeminal foramen; VII=foramen for facial nerve.
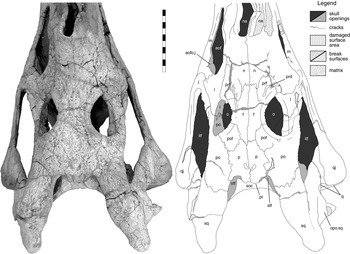
Figure 19 Machaeroprosopus sp. TTU P-10074, postnarial part of the skull in dorsal view. Scale bar=10 mm. Abbreviations: aof=antorbital fenestra; aofo.j=jugular antorbital fossa; f=frontal; itf=infratemporal fenestra; j=jugal; l=lacrimal; m=maxilla; n=nasal; na=naris; o=orbit; opo.sq=opisthotic process of squamosal; p=parietal; pl=parietal ledge; po=postorbital; pof=postfrontal; prd=preorbital depression; prf=prefrontal; q=quadrate; qj=quadratojugal; soc=supraoccipital; sq=squamosal; stf=supra-temporal fenestra.

Figure 20 Machaeroprosopus lottorum TTU P-10077, left bases of parietal–squamosal bar and paroccipital process: (A) dorsolateral view; (B) section along indicated crack. Scale bar=10 mm. Abbreviations: epi=epipterygoid; opo=opisthotic; p=parietal; pro=prootic; ptf=posttemporal fenestra; soc=supraoccipital; sq=squamosal.
2.5.13. Squamosal
A major distinction between M. lottorum TTU-P10076 and TTU-P10077 on the one hand, and all other nominal species of Machaeroprosopus including Machaeroprosopus sp. TTU-P10074 on the other is the shorter length of the free postorbital–squamosal bar i.e., the distance between the tip of the squamosal and the anterior rim of the supratemporal fenestra, which corresponds largely to the maximum length of the squamosal, in proportion to the distance between the anterior rim of the supratemporal fenestra and the orbit. The relative (and absolute) maximum width of the squamosal in the anterior third to midlength of the element is also larger in M. lottorum than in TTU-P10074 and in other species of Machaeroprosopus, with the exception of M. gregorii and M. bermani. The width increase is due to the expansion of the medial lamella of the squamosal. In TTU-P10076, the posterior section of the rim of the medial lamella is squared and the anterior section is rounded (Fig. 9), in contrast to a thin and sharp rim in TTU-P10077. Both specimens show a natural depression on the central medial part of the free postorbital–squamosal bar (e.g., Fig. 6), which enhances the elevated appearance of the posterior process.
The posterior process of the squamosal projects far beyond the extremity of the paroccipital process. Although almost identical in absolute length, the length of the posterior process of the squamosal in proportion to postorbital length is moderate in the larger skull TTU-P10077 (39 mm) than in TTU-P10076 (45 mm), which falls in the same longer relative size class as TTU-P10074. The overall shape of the process is, however, similar in all specimens. It forms a massive, broad, but in particular vertically expanded structure called the terminal knob of the squamosal by Ballew (Reference Ballew, Lucas and Hunt1989). The process is considerably narrower as the parietal–squamosal bar, which leads to a sinuous (in other specimens angular) curvature of the medial rim of the postorbital–squamosal bar (Figs 5, 19; indeterminate in TTU-P10077 because of incompleteness). The medial side of the knob is flattened and confluent with the flat medial rim of the parietal–squamosal bar, the lateral side is convex and bulgy. The verticalisation of the terminal knob results in a dorsal surface sculptured with faint pits and grooves that is significantly raised above the level of the postorbital–squamosal bar. The knob forms the posterior border of a lanceolate area on the horizontal face of the squamosal that show the typical sculpture of anastomosing blunt ridges and elongated grooves in TTU-P10074 and TTU-P10076 (Fig. 5), but is almost unsculptured in TTU-P10077 (Fig. 4). Both TTU-P10076 and TTU-P10077 show large, irregularly shaped pits, suggestive of insertions of tendons, on the lateral face of the ventral base of the posterior process (Fig. 7).
The opisthotic process (Figs 7, 9; ‘opo.sq’) of the squamosal (descending process in Long & Murry 1995; hook-like process in Mehl 1928), the origin of the m. depressor mandibulae profundus (Anderson Reference Anderson1936), projects downward for about 30 mm. The vertical, slightly sinuous anterior margin forming the posterior rim of the otic notch ends in two unequally sized prongs set transversally. The ascending posterior margin shows several protuberances and encloses an angle of 40° with the anterior margin (less steep with 55° in TTU-P10074), resulting in a triangular rather than hook-shaped outline of the process. The smooth lateral surface is confluent with the area for the m. depressor mandibulae superficialis. The dorsal half of the medial face is braced against the extremity of the anterior side of the paroccipital process of the opisthotic. The posterior surface and the lateral end of the paroccipital process merge to a single strongly rugose area that faces posteriorly.
The thin parietal process extends exactly anteromedially (Fig. 9, ‘ppr.sq’). The lateral half is expanded posteriorly by a thin lamella that forms a sharp posterior ridge running parallel to and below the medial rim of the postorbital–squamosal bar. A cross-section of the process is here teardrop shaped, with an extended sharp posterior rim. The broad base of the parietal process, the ventral and the medial surface of the squamosal body enclose the dorsal sulcus (‘ds’), which is the area of the adductor chamber that is open posteriorly because of the depression of the parietal–squamosal bar. The medial section of the bar is twisted for about 90° around the long axis. The posterior ridge is gradually reduced in size leading to an oval cross-section, shifts onto the dorsal surface, and merges with the descending rim of the supraoccipital shelf. Medially, the parietal process enters the corner of the supraoccipital shelf and interdigitates with the supraoccipital, excluding this element from the posttemporal fenestra. The anteromedial extent of the parietal process is largely destroyed on the right side of TTU-P10076 and difficult to access on the left side as well as in TTU-P10074, and accessible in TTU-P10077 only (Fig. 20). Here, the squamosal sends a narrow, 2 mm, thin extension that lies on the opisthotic onto the medial side of the braincase. The extension forms a marked ridge in continuation with the anterior edge of the parietal–squamosal bar. It continues below the anterior face of the squamosal process of the parietal, lying on the descending flange of the parietal, and makes contact with the prootic. The parietal process forms the entire upper rim of the posttemporal fenestra. The ventral surface of the base of the parietal process roofs over the paroccipital process of the opisthotic, and forms with the squamosal flange the ventral sulcus. The squamosal is firmly sutured with the paroccipital process along the distinct lateral edge of the ventral sulcus (Fig. 9, ‘vs’). On the lateral side, a shallow triangular and rugose pit, probably the origin of a tendon, is separated from the ventral sulcus by a broad, ventrally narrowing ridge. Anteromedially, the squamosal develops a thin lamella that overlies the posterior face of the paroccipital process forming the bottom of the vental sulcus. The lamella extends upon the dorsal surface of the paroccipital process and extends forward covering the opisthotic/quadrate suture. Thus, the squamosal does not only form the lateral corner of the posttemporal fenestra, but also the lateral one third of the ventral rim. It continues as a flat, broad process (pterygoid process sensu Mehl Reference Mehl1916) for a short distance anteromedially on the dorsal face of the paroccipital process along the inner side of dorsal rim of the pterygoid–quadrate plate, i.e. the quadrate ramus of the pterygoid.
The quadratojugal process is a broad vertical extension that articulates with the quadrate head medially. At the base of the upper one third of the infratemporal fenestra, the quadratojugal process juts out to form a laterally facing facet for the quadratojugal, which results in a narrow horizontal platform (Fig. 7). The quadratojugal overlaps the quadratojugal process, leaving a thin tapering prong visible along three quarters of the posterior rim of the infratemporal fenestra. Internally, the process extends broadly downward and covers a large area of the quadratojugal.
The sharp lateral edge of the postorbital–squamosal bar continues as a prominent ridge beyond the infratemporal fenestra, and outlines the base of the quadratojugal process as a depressed, smooth area, bordered ventrally by the horizontal platform of the squamosal. This area includes a funnel-shaped depression on the squamosal in extension of the infratemporal fenestra, which was interpreted as the origin of the m. adductor externus superficialis in Smilosuchus gregorii UCMP 27200 by Anderson (Reference Anderson1936). In TTU-P10076, this depression extends into a very faint groove that is cut off by the sculptured dorsal part of the lateral face of the squamosal. The ridge of the postorbital–squamosal bar is reduced to broad and low area, and the dorsal and lateral surface of the squamosal grade smoothly into each other in the centre of the bone and on the posterior process. The smooth area for the m. depressor mandibulae superficialis above the opisthotic process of the squamosal is poorly defined in TTU-P10076. It terminates posteriorly in a concave vertical edge that connects the posterior process with the opisthotic process of the squamosal, here called the squamosal flange (Fig. 9, ‘fl.sq’). The flange is comparatively small and thick, but quickly thins out to a sharp edge that extends only over a short distance on the ventral side of the posterior process. Ventrally, above the opisthotic process, this edge is drawn out into a short, knobby posteroventral projection; the subsidiary opisthotic process of the squamosal (‘sopo.sq’).
2.5.14. Quadratojugal
The quadratojugal has a triangular outline (Fig. 7). Dorsally and posteroventrally, the quadratojugal articulates with the quadrate in two extensive joint surfaces above and below the quadrate foramen; the bone does not, however, contribute to the horizontal platform that is exclusively formed by squamosal and quadrate. The dorsal border with the quadratojugal process of the squamosal has a sinuous outline and, below the quadratojugal, borders the ventral third of the infratemporal fenestra. Anteriorly, the element is drawn out into an outer tapering sharp process that extends along the posterior half of the ventral rim of the fenestra. The anterior process forms a well-marked offset ridge riding on the dorsolateral edge of the jugal. Ventrally to the anterior process, the quadratojugal encloses the posterodorsal part of the jugal with a lateral and a more extensive medial lamina. The external suture between the quadratojugal and the jugal runs from the tip of the anterior process posteroventrally step-like towards the lateral condyle of the quadrate. The thin tapering medial lamina extends along the centreline of the jugal over the posterior third of the infratemporal fenestra. The ventral edge of the quadratojugal is a broad, 25 mm-long rugose area lateral to the posteroventral tip of the jugal and the medial condyle of the quadrate (illustrated also in TTU-P10074, Fig. 8). The contribution of the quadratojugal to the rim of the subtemporal opening is a 2 mm narrow strip that connects the rugose area with the medial lamina.
Posteriorly, the quadratojugal is drawn out into a deep and vertical, but blunt posterior ridge that defines both the lateral edge of the skull and the recess for the quadrate foramen (Fig. 9). The posterior ridge curves outward, as does the entire ventral section of the quadratojugal, and disappears towards the ventrolateral corner of the skull. The central area of the quadratojugal is sculptured with ridges, humps and even spike-like protuberances, but the peripheral areas are smooth or undulating. There is a deep, vertical depression anterior to the posterior ridge and the anteroventral part of the quadratojugal shows a larger, but shallower horizontal depression which extends onto the jugal.
Medial to the posterior ridge, the quadratojugal is wrapped around the side of the quadrate and appears along the side on the posterior face of the skull. Here, the element forms much of the quadrate recess, but actually contributes only ventrolaterally to the rim of the quadrate foramen proper. A groove situated almost entirely on the quadratojugal extends ventrally out of the quadrate foramen between the posterior ridge and the suture with the quadrate.
2.5.15. Quadrate
The quadrate is firmly inserted in the ventral side of the squamosal. The quadrate head curves backward almost to the base of the paroccipital process, and the head is visible in lateral view as semicircular posterior extension of the squamosal (Fig. 7). Lateral to the head, a 40 mm-high vertical extension projects outward for 12 mm to form the dorsal articulation facet for the quadratojugal. The projection forms a horizontal platform of the cheek that is covered by the quadratojugal process of the squamosal dorsally. Medially, the head articulates with the paroccipital process of the opisthotic in a deeply serrated, obliquely transverse suture on the posterior wall of the otic notch; thus, most of the external auditory canal lies within the quadrate.
The neck of the quadrate is the narrowest part of the bone medial to the quadrate foramen (Fig. 9). The foramen itself is framed by the process for the quadratojugal articulation dorsally and dorsolaterally, the neck medially, and the massive articular part of the quadrate ventrally. A marked dorsomedially to ventrolaterally trailing ridge borders the recess of the quadrate foramen posteriorly and continues in a sinuous curve downward, forming the faint medial rim of the shallowing groove between the quadrate and the quadratojugal. Towards the lateral condyle of the quadrate, the groove deepens to a sharp slit exactly on the quadrate–quadratojugal suture. Between the ridge and the neck of the quadrate, a broad shallow groove leads out of the quadrate recess and ventrally and medially in the direction of the medial condyle. Below the level of the quadrate foramen, the quadrate flares out laterally, but in particular medially (Fig. 11), to form the wide but narrow condylar area (TTU-P10076: 84 mm by 20 mm). The medial condyle is almost flat and projects with a sharp rim laterally and anteriorly. The lateral condyle is less extensive, more bulbous and exposed in lateral view. The area in between is anteroposteriorly constricted and bears an indistinct, obliquely oriented central ridge, which is represented by a less prominent, blunt third condyle in TTU-P10074.
The pterygoid ramus of the quadrate is a thin, vertical, triangular plate with truncated anterior apex that is closely applied to the lateral side of the quadrate ramus of the pterygoid (Fig. 18), forming a single unit, the pterygoid–quadrate plate. The plate is triangular with a horizontal ventral rim and an anteromedially sloping dorsal edge (Fig. 9). The ventral part of the pterygoid–quadrate plate is folded medially to form a horizontal ledge 25 mm deep at the base of the pterygoid process, which diminishes to 15 mm anteriorly. At the base of the pterygoid ramus, at the level of the quadrate foramen, is a deep vertical recess enclosed by the horizontal and vertical parts of the pterygoid ramus, and the neck of the quadrate. The suture between the quadrate and the pterygoid on the medial face of the pterygoid–quadrate plate forms a broad triangle extending over the posterior half of the plate. On the ventral surface of the ledge, the suture runs obliquely on a marked ridge from the medial to the lateral edge at about midlength of the plate (Fig. 11). The pterygoid ramus of the quadrate covers, as a triangle with truncated anterior apex, about three quarters of the lateral face of the pterygoid–quadrate plate facing the adductor chamber, the suture between quadrate and pterygoid running atop the dorsal edge.
Internally, a concave lateral flange extends forward onto the inner lamina of the quadratojugal. The quadrate bears four potential attachment areas for the adductor muscles: (1) the blunt, vaulted anterior edge of the flange running from the medial rim of the quadrate foramen downward and outward, plus the steeply inclined plane of the flange that faces medially and slightly anteriorly; (2) a marked oblique ridge in extension of the lateral edge of the ventral surface of the pterygoid–quadrate plate towards the lateral condyle, and the area below this and the horizontal ridge; (3) a horizontal ridge on the central axis of the pterygoid–quadrate plate in extension of the lateral edge of the quadrate ramus of the pterygoid, and the area below, terminating on the lateral edge and the concavity below the medial condyle: and (4) a central ridge on the ventral face of the pterygoid–quadrate plate, and a concave area on the quadrate ramus in front of it.
2.5.16. Vomer
The vomers form the interchoanal septum (Fig. 11). The vomers are extraordinary broad in the anterior section (11·4 mm in TTU-P10076), but become narrower with the dorsal curvature of the septum at mid-length of the choana and are narrow and sharp in the anterior third. The sharp edge of the vomers continues as a crest for a short distance on the palatal vault. The contribution of the vomers to the palatal vault is minute and restricted to a rectangular, 7 mm-long area along the midline and the posteromedial corner of the choana. Anteriorly, both vomers form a simple, 33 mm long prong onto the prechoanal depression and are overlapped by the premaxillae.
2.5.17. Palatine
The ventrally exposed area of the palatine forms the medial section of the palatal plane along the choana and the palatal vault (Fig. 11). The anterior portion of the palatine lateral to the choana is tapering and strongly vaulted. The apex of the convexity faces ventrally, forming an elevated medial part of the palatal plane and a verticalised area that overhangs slightly the anterior section of the choana. Anteriorly, the palatine encroaches upon the maxilla, terminating in a blunt prong about 10 mm in front of the anterior rim of the choana. The section below the choana and the anterior half of the palatal vault is distinguished by a prominent, medially inclined, ragged ridge. It demarcates sharply the now less convex, horizontal palatal section from a vertical lamella, the dorsal ramus of the palatine forming the lateral wall of the choana. The ventral area of the vertical and the entire horizontal lamella is covered with longitudinal ridges, oblique short pits and rugosities. In the postchoanal section (Fig. 14), the palatine thins out distinctly, the ridge merges into the ventral surface, and the vertical section is flexed ventrally, so that the bone is transformed into a horizontal, slightly convex structure with a sharp median edge. This horizontal palatine lamella is extensive, both sides forming two lobes over the palatal vault that leave a narrow, 12 mm-wide gap in between. The palatines thus cover here about 80% of the width of the vault, the closest approach to a fully ossified secondary palate that has been observed in phytosaurs. In its posteriormost section, the palatine narrows and the medial rim curves outward. The element is applied as a thin rod to the medial side of the vertical pterygoid flange. A thin lamella extends dorsally onto the palatal vault; i.e., onto the pterygoid. The exact extent of the lamella on the palatal vault is indeterminable, but seems to be restricted to the ventral parts of the wall, and is largely concealed under the overhang. A narrow posterior extension of the bone is wedged between the pterygoid and the ectopterygoid and almost reaches the rim of the pterygoid flange; this extension is absent in TTU-P10074. The palatine also forms the posterolateral corner of the choana. The suture with the pterygoid is vertical and disappears ventrally under the palatine overhang.
Posterolaterally, a deep groove is developed between the palatine and the ectopterygoid, ventrally along the insertion of the bone with the pterygoid flange. The lateral rim of the broadening palatine becomes sharp and lies on the ectopterygoid, both bones closing the groove and separating it from the suborbital fenestra. Anterior to the suborbital fenestra, the palatine overlaps the maxilla in a suture that leads straight parallel the alveolar rim into the tip of the bone.
2.5.18. Pterygoid
The pterygoid may be subdivided into four areas. Because of the development of a secondary palate, the anterior or palatal ramus is verticalised and consists of (1) the arched ventral section with a medial lip forming with its counterpart the dorsally closed palatal vault; and (2) a vertical extension on top, the dorsal lamella, which is largely hidden from view between the palatal plane and the orbitonasal area. The anterior process grades posteroventrally into (3) the ventral ramus (part of the pterygoid flange); and (4) the quadrate ramus extends posteriorly and laterally.
The palatal ramus extends anteriorly, forming the roof and much of the lateral wall of the pterygoid vault to reach the posterior rim of the choana between the vomer and the palatine. This includes a vertical strut with a concave medial rim 10 mm posterior to the choana that merges dorsally with the wall of the palatal vault and supports the overhanging horizontal palatine like a shelf brace. Both pterygoids send out a medial lip, and they meet at the midline closing the roof of the palatal vault. In the posterior half of the vault, the rims of the lips are turned ventrally, and both are closely applied to form a marked ridge. The vaulted rims diverge about 10 mm in front of the posterior rim of the vault, framing the anterior tip of the interpterygoid vacuity, and curve steeply downward to form the basipterygoid process and grade into the posterior rim of the ventral ramus. The dorsal laminae are largely concealed below the orbitonasal region and only the posterior section is open to description. This section includes an extensive, but only a fraction of a millimetre-thin lamina that extends slightly inclined to the medial side upward, sheathing the base of the cultriform process of the parabasisphenoid. The vertical posterior rim extends in alignment with the posterior rim of the palatal vault and levels out horizontally at a level of about one-third of the space between the palatal vault and the skull roof. In the dorsal one-third, the rims of both laminae diverge in their course forward to enclose the cultriform process. As in Mystriosuchus westphali GPIT 261/001(Huene 1911, fig. 4), the dorsal laminae leave a narrow gap compared to that in Smilosuchus gregorii UCMP 27200 (Camp Reference Camp1930, fig. 34) and Brachysuchus megalodon UMMP 10336 (Case Reference Case1929, fig. 17) that can be traced to at least the posterior end of the choana.
In extension of the posterolateral wall of the palatal vault, the ventral ramus extends downward onto the pterygoid flange (Fig. 14A). The broad and comparatively thick base of the flange above the level of the palatine is formed largely by the pterygoid (Fig. 11), except for the anterolateral rim (ectopterygoid) and parts of the medial surface (lamina of the palatine). Below, the ventral ramus extends into a thinner, vertical lamina that overlies the ventral process of the ectopterygoid, facing posterolaterally. This surface is smooth, and projects up to several millimetres over the rim of the ectopterygoid all around the pterygoid flange. In TTU-P10074, this lamina is particularly thin with sharp edges. The outline of the flange is angular rather than rounded, forming an oblique (anterolateral to posteromedial) subrectangular base with an unequilateral triangular ventral apex. The anterolateral and posteromedial rims are subvertical, and the longer anteroventral and shorter posteroventral rims are inclined towards each other and meet in a blunt corner.
The basipterygoid process of the pterygoid is a cup-shaped posterior projection, attached with a broad base to the vaulted posterior rim of the palatal vault. The concave articular face is directed laterally, the slightly convex inner surface medially. The base of the quadrate ramus is stout and low. Here, the ventromedial rim is raised to form a second, vertical articulation with the basipterygoid process of the parabasisphenoid. Above this joint, a plate-like vertical extension, the epipterygoid process, extends dorsally. The process is largely broken off in TTU-P10074, but shows ragged edges in TTU-P10076 (Figs 18, 21, ‘ept.pt’), and must have been of considerable extent, because the remnants are 20 mm high and 15 mm long. On the lateral side is a broad and shallow groove framed by well-developed ridges (Fig. 18, ‘g’). It curves from the base of the quadrate ramus in extension of the epipterygoid forward and downward towards the ventral ramus and opens up to a broad funnel (‘pqg’). Walker (Reference Walker1990) concluded that a similar palatoquadrate groove in the sphenosuchian Sphenosuchus most likely received the posterior extension of the cartilaginous palatoquadrate, which continued as epipterygoid ossification. Posteriorly, the quadrate ramus expands dorsally, lying medially on the pterygoid ramus of the quadrate, and is folded over ventrally to participate in the horizontal platform of the pterygoid–quadrate plate (Figs 9, 21). The vertical and the horizontal surfaces enclose a deep, cone-shaped recess on the medial face of the base of the quadrate ramus. On the medial face of the pterygoid–quadrate plate, the quadrate ramus develops a dorsal process along the ascending dorsal rim, and a ventral process that largely overlies the quadrate on the horizontal platform and extends into the recess at the base of the quadrate. By contrast to the morphology of the pterygoid ramus of the quadrate, the quadrate ramus is grooved on the ventral face by a raised medial and a ridge-like lateral rim, terminated posteriorly by the oblique ridge of the pterygoid–quadrate suture (Fig. 11). At the base of the quadrate ramus, the grooved ventral face turns ventrally and laterally to merge with the pterygoid flange. This groove probably delimits an attachment area for the jaw adductors in addition to those described on the quadrate.
2.5.19. Ectopterygoid
The ectopterygoid consists of the prominent ventral ramus directed posteromedially onto the pterygoid flange, and the laterally directed palatal part (Fig. 11).
The ventral ramus is verticalised by a twist along the long axis of the ectopterygoid, and underlies the ventral ramus of the pterygoid. The ectopterygoid contribution of the pterygoid flange is considerably thicker than the ventral ramus of the pterygoid, and effectively the ectopterygoid represents the main structural support for the pterygoid flange. At the anterolateral base of the pterygoid flange in extension of the anterior rim of the subtemporal fenestra, the ectopterygoid wraps over the ventral ramus of the pterygoid, and appears as a broad and rugose elevation on the lateral face of the pterygoid flange (Fig. 14A). The ventral and posterior rims of the ectopterygoid on the pterygoid flange are thickened and rugose, and recessed below the pterygoid, indicating the presence of a cartilaginous cap around the edges of the flange. The broad, ridge-like anteromedial rim of the bone, exposed on the left side because of disarticulation of the palatine, forms the lateral wall of a groove between both elements (‘g’). The groove is 45 mm long and less than 5 mm broad. It leads into an elongated, 16 mm-deep sinus preserved as a steinkern in TTU-P10076 (Fig. 14B). The recess is enclosed between the ectopterygoid ventrally and the palatine part of the palatal vault medially and dorsally, and opens anterolaterally into the antorbital cavity. Anteriorly, the rim of the ectopterygoid sharpens, is extended medially and extensively overlapped by the palatine. The ectopterygoid is only about 7 mm thick in the area between the groove and the suborbital fenestra, and this might represent the roofed-over posterior section of the slit-like suborbital fenestra, the plesiomorphic character state. The ectopterygoid expands both vertically and horizontally into the stout and compact palatal part that forms the robust posterior rim of the subtemporal fenestra, and meets the maxilla laterally and the jugal posterolaterally. The jugal process of the ectopterygoid extends for at least 20 mm upward onto the jugal on the medial edge of the bone.
The following account of the braincase elements, their connectivities and structures is largely based on the accessible right side of TTU-P10076. For this section, if not indicated otherwise we refer to Figure 18 for illustration, and present a reconstruction of the braincase in left lateral view in Figure 22.
2.5.20. Epipterygoid
The ventral parts of both epipterygoids are preserved in articulation in TTU-P10076 (Figs 21, 18). The element appears as an anteroposteriorly compressed rod that is 2·9 mm long and 5 mm broad. However, cross-sections of the left epipterygoid show the rod to be a largely hollow tube (Fig. 23). The base is curved anteriorly and merges with the base of the quadrate ramus of the pterygoid to become the dorsal edge of the funnel-like depression. Because no suture is evident, both elements are most likely fused together. The epipterygoid encloses a laterally facing, vertical recess with the epipterygoid process of the pterygoid situated posteriorly to the element, but does not touch the process or if so only with the medial edge at the base.

Figure 21 Machaeroprosopus lottorum TTU-P10076, right side of the braincase with right temporal region removed. Scale bar = 50 mm. Abbreviations: boc = basioccipital; bpt.bsp = basipterygoid process of basisphenoid; bsp = basisphenoidal part of parabasisphenoid; cp.lsp = capitate process of laterosphenoid; cp.psp = cultriform process of parasphenoid; dl.pt = dorsal lamina of pterygoid; ect = ectopterygoid; eo = epiotic; eoc = exoccipital; ept = epipterygoid; eptp.pt = epipterygoid process of pterygoid; f = frontal; hypg = hypophyseal gap; ic = foramen for cerebral branch of internal carotid; j = jugal; jg = jugular groove; (l) = left; lsp = laterosphenoid; mc = medial condyle of quadrate; mdf = median foramen (of laterosphenoid); opo = opisthotic; opo.sq = opisthotic process of the squamosal; p = parietal; pal = palatine; pl = parietal ledge; po = postorbital; ppr.sq = parietal process of squamosal; pro = prootic; ps = presphenoid; ps.p = parasphenoid process; pt = pterygoid; ptf = parasphenoid process; ptr.q = pterygoid ramus of quadrate; q = quadrate; qj = quadratojugal; qr.pt = quadrate ramus of pterygoid; (r) = right; r = recess; rg = rugosity (of laterosphenoid); sg = stapedial groove; stf = supra-temporal fenestra; vp.lsp = ventral process of laterosphenoid with projection; III = foramen for n. oculomotorius; IV = foramen for n. trochlearis; V = trigeminal foramen for n. trigenimus; VII = foramen for n. facialis.

Figure 22 Machaeroprosopus lottorum n. sp., reconstruction of the braincase in right lateral view. Scale bar = 50 mm. Abbreviations: ap.pro = anterior projection of prootic; boc = basioccipital; bsp = basisphenoidal part of parabasisphenoid; cr.pro = crista prootica; dl.pt = dorsal lamina of pterygoid; eo = epiotic; eoc = exoccipital; f = frontal; fo = foramen ovalis; hyp.g = hypophyseal gap; ic = foramen for cerebral branch of internal carotid; jg = jugular groove; l = lacrimal; lsp = laterosphenoid; mdf = median foramen (of laterosphenoid); mf = metotic foramen; opo = opisthotic; p = parietal; po = postorbital; pof = postfrontal; pro = prootic; ps = presphenoid; psp = parasphenoid; psp.p = parasphenoid process; r.lsp = recess in laterosphenoid; sq = squamosal; stg = stapedial groove; tg.lsp = triangular gap of laterosphenoid; vp.lsp = ventral process of laterosphenoid with projection; III = foramen for n. oculomotorius; IV = foramen for n. trochlearis; V = trigeminal foramen for n. trigeminus; VII = foramen for n. facialis; XII = foramen for n. hypoglossus.

Figure 23 Machaeroprosopus lottorum TTU P-10076, cross-sections of left epipterygoid: (A) 14 mm above base; (B) 28 mm above base. Arrows point in anterior direction. Scale bar=1 mm.
2.5.21. Parabasisphenoid and hypophyseal area
The parasphenoid is not clearly distinguishable from the basisphenoid. Only on the right side of TTU-P10076 (Fig. 21) does an ascending suture seem to be present close to the proximal part of the basipterygoid process, trailing toward the hypophyseal gap. The entire area is also marred by cracks, but it seems that the parasphenoid extends as a thin sheet of bone onto the basisphenoid, and is partially broken away here. Otherwise, both elements may be fused over large parts of the braincase, and the complex is best regarded as a parabasisphenoid.
On the exposed left side of TTU-P10076, the cultriform process appears to be composed of two individual structures, separated by a horizontal suture, somewhat similar to the initial description for Mystriosuchus westphali GPIT 261/001 by Huene (Reference Huene1911), subsequently found to be unsubstantiated (Hungerbühler Reference Hungerbühler2002). However, the left side, and the intact condition in TTU-P10074 indicates that this is a fracture that caused a slight ventral displacement of the upper section of the cultriform process. The entire structure projects anteriorly from the base of the braincase in front of the sella turcica. The cultriform process forms a narrow, parallel-sided rod bearing a horizontal, trough-like excavation with very thin walls dorsally and a sharpened rim, which results in a ‘pointed U’-shaped (rather than the usual ‘V’-shaped) cross-section ventrally. The height decreases gradually as the result of a concave ventral rim, and the process is enclosed and concealed anteriorly by the dorsal processes of the pterygoids. Impressions under the dorsal process suggest that the cultriform process extends as a low but broad rod close to the dorsal rim of the processes into the choanal region. At midheight of the cultriform process, there is a blunt, ridge-like expansion below a narrow and faint groove. Posteriorly, the trough of the cultriform process is closed against the sella turcica by a hook-shaped dorsal projection formed by the parasphenoid processes (Fig. 24, ‘psp.p’). The delicate walls of the trough are extended dorsally here, curve inward and are closely applied and meet in a sharp posterior edge, forming a flat, vertical, recurved hook. The parasphenoid processes are inserted between two anteroventral projections of the presphenoid, but leave narrow gaps to each projection. The upper rim approaches the floor of the optic nerve foramen. It is possible that a narrow fissure remains between the ventral sections of both halves of the projection through which the trough on the cultriform process communicates with the sella turcica, but the structure is too delicate to safely allow further mechanical preparation. In front of the base of the parasphenoid process is a notch in the wall of the parasphenoid trough that expands into a large, round foramen (this area is crushed on the left side). Ventrally, the base of the cultriform process expands posterolaterally on each side onto the basisphenoidal portion of the braincase in a sharp and high, curved ridge that contacts the anterior pillar of the prootic and frames the sella turcica laterally.

Figure 24 Machaeroprosopus lottorum TTU-P10076, otic area of the braincase in right lateral view. Scale bar = 50 mm. Abbreviations: boc = basioccipital; bsp = basisphenoidal part of parabasisphenoid; d = depression; eoc = exoccipital; fm = foramen magnum; fo = foramen ovalis; f.pat = facet for proatlas; jg = jugular groove; lr = lateral ridge sensu Gower & Walker (Reference Gower and Walker2002); mf = metotic foramen; ol = opisthotic lamella; pit = pit sensu Camp (Reference Camp1930) in stapedial groove; plt = platform of anterior wall of stapedial groove; p.psp = parasphenoid process; pr = projection; pr.lsp = anteroventral process of the laterosphenoid; prs = presphenoid; r = recess; stg = stapedial groove; v = trigeminal foramen.
The hypophyseal area has been freed from matrix, but is still difficult to access because of the parasphenoid processes of the cultriform process. The sella turcica is a conical pit deeply excavated into the basisphenoidal portion of the parabasisphenoid, opening dorsally, below a comparatively high, vertical, anteriorly concave dorsum sellae. A subtriangular hypophyseal gap lies between the parabasisphenoid and the cultriform process ventrally and anteriorly, the presphenoid dorsally, and the prootic posteriorly. As it most likely represents a cartilaginous zone of the anterior portion of the braincase, it is a preservational gap rather than a true fenestra as designated in Camp (Reference Camp1930) and Chatterjee (Reference Chatterjee1978). The plane of the gap faces anterolaterally, and exposes the dorsum sellae and the uppermost part of the sella turcica. The prootic–parabasisphenoid suture continues horizontally over the dorsum sellae, and therefore much of the dorsum is formed by the prootic. The paired cerebral branches of the internal carotids enter the bottom of the sella turcica through the parabasisphenoid. The hypophyseal cavity communicates with the braincase cavity via a round central foramen in the dorsum sellae just below the presphenoid.
The basipterygoid process is a column with a triangular cross-section, the blunt apex of which lies level with the plane of the tubera (TTU-P10074) or slightly below (TTU-P10076). Two vertical, flat facets occupy the entire height of the process: a lateral one with the base of the quadrate ramus of the pterygoid, and an anterolateral one with the cup-shaped basipterygoid process of the pterygoid. The pterygoid thus enwraps the basipterygoid process of the parabasisphenoid on two sides, with only the convex posteromedial face of the process being free. Ventral displacement of the right basipterygoid process of the parabasisphenoid relative to the left one in TTU-P10074 indicate that the joints were not fused, but potentially mobile. The shape of the articulation allows only vertical sliding movements along the joint surfaces, provided that movement of both components was not restricted by other means. In ventral view, the base of each basipterygoid process is defined by a crescentic, marked ridge on the postero-medial side. On both sides, the ridges converge from the apices of the recesses between the basipterygoid processes and the basioccipital tubera toward the ventral extent of the parabasisphenoid below the cultriform process. These ridges also outline the anterior extent of the central, shallow, triangular median pharyngeal recess between both processes. Each ridge forms the ventral rim of a groove that curves upward and forward around the base of the basipterygoid process toward the lateral face of the cultriform process.
The foramen for the cerebral branch of the internal carotid is situated in the parabasisphenoid high up on the anterior face of the base of the tuber, anterior to the ridge in continuation of the anterior wall of the stapedial groove. It opens into a posterodorsally ascending, short groove that crosses this ridge.
2.5.22. Basioccipital and basioccipital tubera
The basioccipital condyle is hemispherical (Figs 9, 10). The central normal axis of the condylar surface points somewhat more ventrally than posteriorly, being located a few millimetres below the notochordal pit, but the condylar surface is far from facing posteroventrally as in most phytosaurs. Analysis of the bone texture indicates that the bases of the exoccipitals are functionally integrated into the articular surface. The exoccipital–basioccipital sutures on the condyle are marked by two deep, slightly concave grooves. A deep notochordal pit faces exactly posteriorly. From the pit, a deep vertical groove trails dorsally toward the joint exoccipital suture. The basioccipital neck is stout, relatively broad, and smooth ventrally, i.e. without a central ridge.
The basioccipital tubera extend ventrally, but predominantly laterally to a width of 80 mm as opposed to 27 mm at the basioccipital neck. The rugose tuberal surface is directed ventrally. The tubera form a confluent unit, the left and right side of which is separated by a short and sharp ridge. A groove on the lateral edge distinguishes the basisphenoidal contribution from the basioccipital contribution of the tubera. The parabasisphenoid forms about two thirds and the basioccipital one third of the ventral surface of the tubera. A central tongue of the basioccipital, bearing the midline ridge, reaches the anterior rim of the tubera and separates both parabasisphenoidal halves. The basioccipital portion of each tuber projects farther laterally than the parabasisphenoidal portion to a blunt point, and is in addition even slightly recurved in TTU-P10074. The parabasisphenoidal portion is larger anteroposteriorly, with a rounded rectangular outline, and more rugose. The well distinguished muscle scars of the m. basioccipito-vertebralis (Anderson Reference Anderson1936; in crocodylians, m. longus colli, Frey Reference Cope1988) are oblong oval, positioned slightly oblique, and meet anteriorly at the midline ridge.
2.5.23. Laterosphenoid
TTU-P10076 preserves the complete anterior processes of the laterosphenoids, which enclose the olfactory tract of the brain that give rise to the olfactory nerve (I). The laterosphenoid is here sutured to a sharp, prominent ventral ridge on the frontal, which continues beyond the laterosphenoid forming the anterior internal rims of the orbit. The laterosphenoids extend as an oval tube along the posterior three quarters of the orbit, but do not reach the prefrontals as in Smilosuchus gregorii UCMP 27200 (Camp Reference Camp1930). The ventral symphysis of the anterior processes is somewhat broadened and rugose, indicating the attachment of the interorbital septum, and becomes a more prominent ridge toward the optic foramen. There is no external evidence that the anterior process of the laterosphenoid consists of several sphenoidal elements, as postulated for Smilosuchus gregorii UCMP 27200 (Camp Reference Camp1942), nor is an ossification in the dorsal section of the interorbital septum (as in Smilosuchus gregorii UCMP 27200: Camp, Reference Camp1930; or as in M. jablonskiae PEFO 31207: Parker & Irmis Reference Parker and Irmis2006) detectable. With the divergence of the internal orbital ridge in the anterior section of the orbit, the symphysis is separated by a narrow cleft, and the olfactory tube opens anteriorly as a large round funnel. Just above the optical nerve is a single median foramen on a mound-like elevation, in a similar position as one in Smilosuchus (Camp Reference Camp1930, figs 25, 37, “r”). The anterior third of the anterior process of the laterosphenoid, as well as a narrow zone below the frontal–laterosphenoid suture and the symphyseal area are slightly rugose to finely pitted, which may indicate that the ossification was not fully completed.
The prominent capitate process of the laterosphenoid arches upward and outward and meets the frontal at the anterior base of the process, the postfrontal and the postorbital in a central triple point, and the tip of the parietal posteriorly. At the extremity, there is a shallow triangular gap between the laterosphenoid on the medial side, and the prefrontal, postorbital and parietal laterally (as in TTU-P10074; also noted in modern crocodylians: Iordansky Reference Iordansky, Gans and Parsons1973), suggesting that the capitate process was not fully ossified and the extremity was represented by a tongue of cartilage. The axis of the capitate process is marked by a blunt subvertical ridge, the cotylar crest, aligned ventrally with the anteroventral process of the bone. Between the olfactory tube, the capitate process, the frontal, and the postfrontal is a considerably large, but shallow recess (also present, but much deeper in TTU-P10074) that continues forward as a groove along the frontal–laterosphenoid suture. A foramen at the bottom of the recess most likely represents the exit of the vena cerebralis anterior (cf. Janensch Reference Janensch1936; Walker Reference Walker1990).
The posterior section of the laterosphenoid is wedged between the descending wall of the parietal posterodorsally and the prootic ventrally, and contacts the epiotic posteriorly. The horizontal suture with the prootic runs on a broad ridge that terminates anteriorly in a small, pyramidal, upward pointing projection, situated entirely on the laterosphenoid. The suture is indistinct and appears to be largely fused.
Anteriorly, the laterosphenoid forms the roof of the transversely kidney-shaped optical foramen (II). Paired crescentic grooves dorsally to the lateral lobes of the foramen probably received the epipterygoid (see below). The exit for the trochlearis nerve (IV) is in a small depression laterally and slightly above the optical foramen. A small foramen of unknown affinities lies in the laterosphenoid just above the tip of the presphenoid in both TTU-P10076 and TTU-P10074. A narrow ventral process of the laterosphenoid extends between the presphenoid and the prootic into the roof of the trigeminal foramen (V). A distinguished feature of this process is an anteroposteriorly flattened, rectangular projection above the trigeminal foramen that is directed anterolaterally and ventrally (Fig. 24). The ventral rim of the process extends as a faint ridge onto the roof of the trigeminal foramen, separating two passages out of the opening. As interpreted by Camp (Reference Camp1930) for Smilosuchus gregorii, the ventral section of the laterosphenoid–prootic suture probably runs in the deep, V-shaped cleft behind this projection. The anterior border of the ventral process with the pre-trigeminal part of the prootic is marked by a vertical suture from mid-height of the presphenoid to the rim of the trigeminal foramen. However, the suture cannot be traced onto the inner wall of the opening, and the laterosphenoid and the prootic seem to be partially fused.
2.5.24. Presphenoid
The presphenoid appears as a stout, triangular element. It is unclear whether the element represents an unpaired ossification or two fused symmetrical elements, although the strongly damaged bone in TTU-P10074 does not reveal a median suture and favours the first interpretation.
The body of the element, a central, anteriorly sloping platform, forms the floor of the optic foramen. Two wing-like, extremely thin dorsal processes, largely lying on the laterosphenoid (TTU-P10076, left side), contribute to the lateral rims of the optic foramen. Two stubby, rectangular processes point ventral and anterior, and terminate in small facets that face slightly outward (Fig. 24). These processes receive the dorsal projection of the cultriform process in between them. Two small ventral processes extend along the laterosphenoid and the hypophyseal section of the prootic in the direction of the anterior pillar of the prootic. The oculomotorius foramen (III) is situated between this process and the prootic.
2.5.25. Prootic
The trigeminal foramen (V) is bounded largely by the prootic, separating the anterior pillar of the element from the posterior capsular portion, with a contribution of the laterosphenoid to the roof. The foramen, which also gave exit to the vena cerebralis medialis and most likely the abducens nerve (VI) (see discussion), is a large, vertical oval opening. It is 20 mm deep, with a shelf-like, concave ventral rim and overall converging walls. The actual foramen into the braincase cavity is about half as large as the external opening, and the deep walls might have enclosed and protected the trigeminal (Gasserian) ganglion. Two passages out of the trigeminal foramen can be reconstructed by osseous correlates and are indicated in Figure 21: one passage leading dorsally through the cleft between the anteroventral process of the laterosphenoid and the ridge-like continuation of the external posterior wall of the foramen. It probably transmitted the ophthalmic branch of the trigeminal nerve (V1), similar to the interpretation by Camp (Reference Camp1930) for Smilosuchus gregorii. The ophthalmic branch most likely crossed the ridge of the capitate process through a groove between the base of this process and the anterolateral process of the laterosphenoid, to continue in a fainter groove upward and forward along the curvature of the laterosphenoid. A second broader, anterodorsal exit is marked by a notch between the projection of the anteroventral process of the laterosphenoid and the anterior pillar of the prootic. We speculate that this notch indicates the course of the abducens nerve (VI).
The anterior pillar of the prootic is stout, rises in a dorsal direction lateral to the hypophyseal area to the level of the top of the dorsum sellae, and forms the expanded dorsal flange of the dorsum. On the inner side, a curved ridge extends to the base of the presphenoid as the dorsolateral rim of the dorsum sellae. The oblique suture with the clinoid process of the parabasisphenoid enters the hypophyseal gap at midheight (Fig. 21). On the base of the pillar, close to the ventral rim of the trigeminal foramen, is a finely ridged area from which the m. retractor pterygoidei arose (Walker Reference Walker1990). A peculiar feature so far not recognised in phytosaurs is an anterior triangular, flat projection at the top of the anterior pillar. It is largely lost on the right side of TTU-P10076 (Fig. 21), but present on the left side. In TTU-P10074, the size of a similar projection is considerable, with a length of 12 mm and a base height of 9 mm.
The capsular (post-trigeminal) exposure is large and extends below the laterosphenoid, as described above, meets the epiotic posterodorsally, and extends ventrally and posteriorly along the parietal and opisthotic, forming a thin superficial splint on the anterior edge of the base of the paroccipital process. On its ventral face, the prootic–opisthotic suture trails medially into the stapedial groove and towards the opisthotic lamella, but is cut off by a large crack. Ventrally, the prootic–parabasisphenoid suture crosses the extension of the stapedial groove between the foramen ovale and the base of the tuber and turns dorsally onto the anterior face of the opisthotic lamella; the further course is obscured by damage.
The crista prootica runs subhorizontally parallel to and above the anterior edge of the paroccipital process towards the posteroventral rim of the trigeminal foramen. The posterior section of the crista prootica is extended into a marked horizontal flange that is thick and blunt in TTU-P10076, but much sharper in TTU-P10074. The posterior section of the flange terminates abruptly, and the crista prootica faints quickly in anterior direction in some distance behind the trigeminal foramen, so that the structure resembles more a short overhang rather than a proper crest. The singular foramen for the facial nerve (VII) is a horizontal slit in the centre of the space between the edge of the anterior section of the crista prootica and the anterior lamella of the stapedial groove. It is overhung by a convex ridge. This ridge provides a short groove for the palatine branch of the facial nerve in anteroventral direction, and a much more pronounced and longer channel for the hyomandibular branch of the facial nerve, running in a posterior direction above the anterior wall of the stapedial groove and overhung by the flange of the crista prootica.
2.5.26. Structures of the otic region
The complex morphology of the region of the stapedial and the jugular grooves is separately described and illustrated in Figure 24. The configuration of the elements is only in parts determinable, and includes the posteriormost section of the prootic, the dorsal parts of the parabasisphenoid and the basioccipital, the anterior section of the exoccipital, and the base of the paroccipital process and presumably the ventral ramus of the opisthotic. The foramen ovale and the metotic foramen lie in the lateral halves of these grooves, which continue anteromedially onto the base of the basioccipital tuber.
The stapedial groove is twice as broad as the jugular groove, and not divided into a stapedial and a tympanic fossa as figured for Smilosuchus gregorii UCMP 27200 (Camp Reference Camp1930, fig. 37). The foramen ovale is small and oval (long axis is 6 mm), situated close to the posterior rim opposite a small recess in the anterior wall. Anteromedially, the groove passes over a small, round depression and leads into a narrow passage caused by a platform-like bulge of the anterior wall with the traversing prootic–parabasisphenoid suture. It widens abruptly to a broader medial depression. Here, an elongated recess, which could also be a foramen, opens into the anterior wall (Camp Reference Camp1930, fig. 37 indicates a ‘pit’ in similar position for Smilosuchus gregorii). The anterior wall formed by the prootic is the most prominent elevation. It shows a marked step above this ‘pit’ to continue as a lower, but sharp ridge forward and downward, until it curves laterally to merge with the lateral face of the basisphenoidal portion of the tuber.
It cannot be demonstrated for TTU-P10076 (or for TTU-P10074) that the opisthotic forms a ventral ramus separating the stapedial from the jugular groove. This ramus, termed the opisthotic lamella (Walker Reference Case and White1990; Gower Reference Gower1997, Hungerbühler Reference Hungerbühler2002; crista paroccipitalis of Chatterjee (Reference Chatterjee1978); descending ramus of Stocker (Reference Stocker2010)), is thin and sharp and continues anteromedially as a marked edge of the basisphenoidal portion of the tuber. The suture between the parabasisphenoid and the basioccipital runs parallel to this ridge in the anterior portion of the stapedial groove.
The jugular groove is much narrower and somewhat deeper than the stapedial groove. The metotic foramen is a large and long oval (15 mm by 5 mm) and occupies the entire mid-section of the jugular groove. The jugular groove continues as a slowly widening cleft onto the basioccipital. The posterior wall formed by the exoccipital and the basioccipital (the equivalent of the lateral ridge (Gower & Walker Reference Gower and Walker2002) or the subvertical crest (Gower Reference Gower2002) of the exoccipital in pseudosuchians) is low, broad and blunt. A broad anterior projection narrows the jugular groove below the metotic foramen, but the opening is clearly not subdivided into a foramen pseudorotunda and a vagus foramen. Lateral to this projection and close to the metotic foramen, a single hypoglossal foramen (XII) exits the exoccipital in the dorsal portion of the posterior wall of the jugular groove. The hypoglossal nerves leave the braincase thus in an anterior, rather than a lateral direction.
2.5.27. Opisthotic
The cochlear part of the opisthotic is treated above. The complex internal configuration of the opisthotic here (see Camp Reference Camp1930) and the external appearance on the base of the paroccipital process is largely indeterminable in the specimens described here.
The paroccipital process of the opisthotic (Fig. 9) extends in a posterolateral direction from the midline, both enclosing an angle of about 45–50°. The base of the paroccipital process is exceptionally broad. In ventral view, the process expands to the rear in a sinuous curve, resulting in the oblique position of the posttemporal fenestra with respect to the axis of the paroccipital process of the opisthotic and the parietal–squamosal bar, respectively and, in the exoccipital pillar, being situated in extension of the midline of the paroccipital process.
The lateral half of the paroccipital process is expanded in a significantly ventral direction and the process is transformed into a comparatively narrow, vertical structure. In M. lottorum TTU-P10076 (Fig. 9) and TTU-P10077, it is abrupt with a kink marked by a ventral prong, followed by a level ventral rim (oar-shaped paroccipital process sensu Ballew Reference Ballew, Lucas and Hunt1989), whereas in TTU-P10074, the expansion is gradual (Fig. 10). The anterior face is braced against the quadrate and the entire dorsal face slots into the squamosal and is covered anteromedially by a lamina of this element. Posterodorsally, the narrow lateral face abuts against the subsidiary opisthotic process of the squamosal.
2.5.28. Epiotic
The epiotic is a separate ossification in the dorsal part of the otic capsule that later in ontogeny fuses with the supraoccipital (e.g., Rieppel Reference Rieppel1993). Because such a fusion cannot be demonstrated in the specimens described, in contrast to the condition in M. jablonskiae PEFO 31207 (Parker & Irmis Reference Parker and Irmis2006), the epiotic is treated here as separate entity. The epiotic is exposed as a small, roundish area in the centre of the lateral face of the braincase. It is distinguished from the surrounding parietal (posterodorsally), laterosphenoid (anteriorly) and prootic (anteroventrally) by well-developed sutures. The epiotic is at the bottom of a well-defined, subrectangular depression in an oblique position, much deeper and better defined than in any other phytosaur. The depression opens anteroventrally onto the prootic by means of a broad channel. It almost certainly housed the exit of the vena capitis dorsalis, although there is no foramen visible in TTU-P10076, TTU-P10077 or TTU-P10074. M. jablonskiae has a vena capitis dorsalis foramen in the laterosphenoid close or at the junction with the supraoccipital in a similar depression (Parker & Irmis Reference Parker and Irmis2006). Thus, the presence of this foramen is variable within two species of Machaeroprosopus, but the taxonomic usefulness of the character needs to be explored over a wider range of taxa.
2.5.29. Exoccipital
The exoccipitals meet along the midline and thus form most of the periphery of the foramen magnum and the cavity of the medulla oblongata. The exoccipital pillars are stout and broad, as the lateral face arches outward to merge with the paroccipital process. The element is fused to the opisthotic, and even the contact with the supraoccipital above the foramen magnum seems to be fused in TTU-P10076. The exoccipital–opisthotic complex has a broad exposure on the lateral face of the braincase, with a slightly ascending suture with the basisphenoid.
2.5.30. Supraoccipital
The outline of the supraoccipital on the centre of the supraoccipital shelf (Figs. 9, 10) is triangular, covered by concave sutures with the laminae of the squamosal processes of the parietals and truncated above by the parietal, near the apex of the shelf 10 mm below the parietal ledge. The inclined plane of the supraoccipital bears a faint central ridge in its lower half. Laterally, the supraoccipital narrows into a triangular prong toward the corners of the shelf, and is truncated by strongly serrated sutures with the squamosal. In TTU-P10076 and TTU-P10077, the element extends close to the posttemporal fenestra, but fails to reach this opening. An unusual modification is present on the left side of TTU-P10077 (Fig. 20): the supraoccipital appears on the dorsal surface of the base of the parietal–squamosal bar, separating the squamosal from the parietal by deeply interdigitating sutures on this surface.
The suture with the exoccipital/opisthotic complex is present on the edge below the extremity of the shelf, trailing toward the posttemporal fenestra. In TTU-P10076, the supraoccipital and exoccipital are fused above the foramen magnum. Two transversely elongated, smooth humps above the foramen magnum represent the facets for the proatlantes (Fig. 24). The condition in TTU-P10074, in which the proatlas articulations are rugose projections with a central concave facet, suggests that the medial half of each facet is situated on the supraoccipital, the lateral half on the exoccipital.
2.6. Dentition
TTU-P10076 is one of the very few specimens referable to Machaeroprosopus that provides substantial primary data on the dentition. There are 48 teeth preserved in situ, out of which 12 are fully erupted and complete enough for an identification of the dental characteristics. Although there are some uncertainties because of gaps in the tooth row, this specimen allows the determination of the diversity of the tooth morphologies and the onset of dental features within the premaxilla and maxilla series. The dental nomenclature follows the suggestions of Smith & Dodson (Reference Smith and Dodson2003).
The premaxilla contains 27 and the maxilla 24 tooth positions, resulting in a total of 51. The spacing of the alveoli is regular; premaxillary teeth are set in a distance of half an alveolar diameter, which decreases to one third between the larger pm24 to pm26. There is a distinct, 12 mm-long diastema anterior to m1, which is largely on the premaxilla. M1 through m3 are very close together, m4 through m16 are separated by half an alveolar diameter, and posteriorly the interalveolar space gradually decreases, being very close again between the last seven maxillary alveoli. With the exception of pm1 and pm2, the alveoli are circular and differ only in size. Alveolus pm1 is labio-lingually oval (15 by 11·5 mm). The round, 10 mm-wide alveoli pm3 and pm4 face ventrally, rather than ventrolaterally. Posteriorly, the alveolar plane on the premaxilla and the anterior section of the maxilla is inclined for 30° relative to the horizontal plane (Fig. 17). Alveoli pm5 through pm22 are even sized, with diameters of between 7 mm and 8 mm. The alveolar width increases to 9 mm in pm23 through pm25, pm26 being again noticeably smaller (the size of pm27 is indeterminate). Alveoli m1 through m3 get rapidly larger from 5 mm to 7 mm, followed by a gradual size increase to 10·5 mm in alveolus m23. The two last alveoli are significantly smaller.
Regardless of their position, the teeth have several features in common. In fully erupted teeth, the crown-root line is consistently situated 4–6 mm above the alveolar plane and in some specimens such as pm1 even substantially more. Hence, the gum line must have been at about the level of the alveolar ridges. Secondly, all teeth are smooth, without ornamentations seen in comparable phytosaur dentitions, such as wrinkles in Rutiodon carolinensis (Emmons Reference Emmons1856) or fluting by widely spaced ridges in Mystriosuchus planirostris (McGregor Reference McGregor1906; Hungerbühler Reference Hungerbühler1998). Visible pseudostriae in the teeth of TTU-P10076 are actually dense discontinuous arrays of cracks in the enameloid.
The first of the two enlarged premaxillary teeth (pm1 left) extends for 39 mm out of the alveolus, but the actual tooth crown height (TCH) is 27·6 mm. The crown is steadily curved lingually. The cross-section is oval (fore–aft basal length (FABL): 9·5 mm; basal width (BW): 10·1 mm) with a rounded labial and a more pointed lingual side. A faint, unserrated lingual carina extends over the entire crown height. The premaxilla teeth posterior to position pm4 are uniform in shape. Tooth crowns are acarinate, slender and conical with an oval, labiolingually extended base (in average, BW is 1·5 times as wide as FABL) that taper gradually to a round cross-section towards the apex. The maximum crown heights are 24 mm (pm7) and 20 mm (pm12). The basal part of the crown is straight. In the anterior positions, the apical half to one third of the crown is decidedly curved in lingual direction; in more posterior positions the curvature is less over only the apical one-third. This gives the premaxillary teeth a kinked appearance. There are no teeth preserved in the posteriormost enlarged premaxillary alveoli or in the anterior maxilla positions. Teeth in mid-position of the maxilla resemble premaxillary teeth, but are stouter (m7: TCH 16 mm; FABL 6·6 mm; BW 7·4 mm) and less curved. A faint, unserrated distal carina appears first in position m8. Starting with m11, the maxillary teeth are curved in a distal direction. M15 begins the transition to posterior maxillary teeth with a serrated distal carina over the entire crown, and a mesial carina over the apical fourth of the crown. The cross-section is rounded with a distal edge, but the lingual side is flatter, although still decidedly convex. One of the posteriormost maxillary tooth, m23, is stout (TCH 8 mm; FABL 9·4 mm; BW 5·9 mm), with a triangular outline with two fully serrated carinae. Because the mesial carina is more convex than the distal, the crown axis is slightly recurved. In all four posterior maxillary teeth in which the serrations are preserved, the density amounts to 4–4·5 denticles per mm on both carinae, measured at midheight of the crown. In contrast to teeth from similar positions in strongly heterodont phytosaur dentitions (e.g., Nicrosaurus kapffi SMNS 5727, Hungerbühler Reference Hungerbühler2000), posterior maxillary tooth crowns are biconvex, although the labial side is much more vaulted, the carinae do not form distinct blades that are morphologically set apart from the central labial surface of the crown, and these teeth do not show an imbricating pattern.
The nature of the dentition of TTU-P10077 must be largely inferred from the alveoli. There are 24 premaxillary and 23 maxillary teeth, totaling 47. The terminal rosette contains two strongly enlarged teeth followed by two round, 11 mm-wide alveoli which are widely spaced and set high up on the lateral side of the constriction facing ventrolaterally. Posterior to pm5, all alveoli are very closely spaced; being separated by thin interalveolar septa with a maximum thickness of 3 mm. Pm 5 is round, with a diameter of 5 mm, which increases to an 11·5-mm labio–lingual width in pm8. From here, the shape of the premaxilla alveoli is transversely subrectangular, and there are no size differences up to position pm23. The alveolar size changes abruptly to large dimensions in pm 23 and 24 (labio–lingual 17 mm, mesio–distal 16 mm), pm 24 is somewhat smaller (17mm by 13·5 mm). This suggests an abrupt transition to three large posteriormost premaxillary teeth. There is no diastema between premaxilla and maxilla. M1 to m7 are strongly mesio-distally compressed, extremely closely spaced, and show a transverse subrectangular outline with the transverse axis pointing laterally and somewhat anteriorly. The dimensions increase gradually from 11 mm by 5 mm in m1 to 13·5 mm by 10·5 mm in m7, which is the first oval maxillary alveolus. There is a greater size increase between m7 and m13 (17 mm by 15·5 mm). The alveoli are oval with the long axis oriented transversally. M14 to m21 are the largest alveoli in the dentition with no size differences in the series, m20 being 18 mm by 15 mm long. The shape is broad oval, and the long axis is directed posteriorly and laterally. Alveoli m22 and m23 are smaller, the last one with an 11·5-mm short axis and a 14-mm long axis that points directly posteriorly.
2.7. Skull morphology of Machaeroprosopus sp. TTU-P10074
TTU-P10074 differs in a number of character states from Machaeroprosopus lottorum TTU-P10076 and TTU-P10077, and some structures are preserved in more detail than in these specimens.
2.7.1. External skull morphology
Two features on the rostrum distinguish the dolichorostral TTU-P10074 from TTU-P10076: (i) the posterior section of the prenarial area slopes less steeply, at an angle of about 50° (Fig. 8). The slope is somewhat steeper in front of the prenarial protuberances to the left and right of the midline, thus enhancing the pillar-like appearance of these structures, but not along the midline; (ii) the prenarial area is generally rounded, and the top surface grades indistinguishably into the tooth-bearing section of the rostrum.
The smooth intraorbital area of TTU-P10074 is demarcated posteriorly by a blunt ridge pointing laterally and posteriorly, which is an intermediate stage between the faint ridge of TTU-P10074 and the projecting flange of TTU-P10077.
TTU-P10074 differs from M. lottorum in having a considerably narrower parietal ledge (26 mm versus 39 mm in TTU-P10076, at identical overall width of the skull table). The squamosal processes of the parietals converge slightly towards the parietal ledge, resulting in an ‘inverted-U-shaped’ (sensu Ballew Reference Ballew, Lucas and Hunt1989) frame around the supraoccipital shelf. In addition, the slope of the squamosal processes of the parietals is only vertical close to the ledge, but slopes continuously at a steep angle below.
The exposure of the palatal vault is parallel-sided in TTU-P10074, rather than severely constricted as in TTU-P10076, and the undistorted choana is 83 mm longer than the naris. The suborbital opening seems to have been much narrower in TTU-P10074 , and a lobate rim of the right palatine may indicate that the suborbital fenestra was subdivided into several smaller foramina, as in the types of the pseudopalatine phytosaurs Mystriosuchus westphali (Huene Reference Huene1911) or Machaeroprosopus andersoni (Mehl Reference Mehl1922).
TTU-P10074 differs from both specimen of M. lottorum in several features of the narial area (Fig. 6): The centre of the narial cone is deformed to the right side, and thus the narial rims appear to be angular and the narial openings constricted at midlength; in life, the shape of each naris was subrectangular with a more pointed anterior rim. The anterior rim is closed, although two narrow and shallow grooves extend out of the nasal openings. The sharp anterior narial rims are crescentic pillar-like protuberances, between which the narial grooves and the median suture of the septomaxillae pass onto the prenarial area. The smoothly rounded lateral rim of the naris in general slopes anteriorly; the rim is sinuous, the central section of the rim being levelled out, and thus the posterior rim is elevated for 10 mm above the anterior (Fig. 8). A similar condition in Mystriosuchus was termed a narial wing by Hungerbühler (Reference Hungerbühler2002). The median suture on the septomaxilla runs in a deep and narrow groove, present to about midlength of the internarial septum. The internarial septum is narrow and sharp, without any significant increase in width anteriorly.
2.7.2. Internal structures of maxilla and septomaxilla
An extensive canal runs in the maxilla dorsal to the alveoli, probably transmitting blood vessels and nerves to supply the teeth (Figs 16A, 17, ‘ac’). In the proximity of the choana, the palatal section of the maxilla is extensively pneumaticised, showing a central large cavern that is further subdivided by thin laminae and an array of accompanying smaller caverns or canals (Fig. 16A, ‘cv.m’). In TTU-P10074, the median suture between the septomaxillae and, in continuation, the premaxillae, is developed as a deep central groove all along the internarial septum and over a distance of 145 mm anterior to the nares. Figure 16B shows details of the internal structure: a vertical lamellae of the septomaxilla interdigitates with the underlying premaxilla, among them a very thin, sheet-like lamella along the sagittal plane that reaches the deeper parts of the nasals. Internally, the septomaxilla contributes extensively to the roofs of the nasal and premaxillary cavities in Smilosuchus gregorii UCMP 27200 (Camp Reference Camp1930, figs 22, 23, 25), but the cross-sections of TTU-P10074 (Fig. 16A, B) are too far forward to show the ventral extent of the element for Machaeroprosopus.
2.7.3. Jugal (Fig. 8)
TTU-P10074 has a broad and stout ventral process of the lacrimal that contacts and forms a short distance of the ventral rim of the antorbital fenestra. A further distinction from M. lottorum is that a well-defined, crescentic, bumpy ridge surrounds the preinfratemporal shelf. This large area is 35 mm by 17 mm and opens dorsally into the postorbital depression.
2.7.4. Parietal
In TTU-P10074, the parietal ledge shows irregular vertical ridges rather than two distinct prongs (Reference ChatterjeeFig. 10). Two symmetrically placed, posteriorly directed rugose knobs are situated on the inner side of the squamosal processes of the parietal of TTU-P10074 just below the parietal ledge. Similar structures are absent in TTU-P10076 and P10077, but were reported for Mystriosuchus westphali (Hungerbühler Reference Hungerbühler2002).
2.7.5. Squamosal
In accordance with the total length of the squamosal, the posterior process of the squamosal on the undeformed right side of TTU-P10074 projects far beyond the extremity of the paroccipital process (58 mm). In contrast to M. lottorum, the ridge of the postorbital–squamosal bar in TTU-P10074 becomes broader and blunter, but continues posteriorly onto the posterior process (Fig. 8). Thus, the dorsal and lateral surfaces of the central section of the squamosal are set at a distinct angle in this specimen. A narrow but distinct groove extends in TTU-P10074 from the external adductor depression backward. It peters out above the level of the opisthotic process. This groove demarcates an elevated zone below, which merges with the bulging lateral surface of the posterior process. This comparatively broad zone is reminiscent of the narrow, well-developed lateral ridge of the squamosal typical of M. pristinus and M. buceros but, in contrast to these, the elevation does not extend onto the posterior process in TTU-P10074. The smooth area for the m. depressor mandibulae superficialis above the opisthotic process of the squamosal is distinctly demarcated by a horizontal bumpy overhang in TTU-P10074. The parietal process of the squamosal of TTU-P10074 extends farther medially on the posterior face of the supraoccipital shelf, and expands dorsoventrally onto the shelf (Fig. 10). In medial aspect, the entire rim of the medial lamella of the squamosal is developed as a vertical plane that is demarcated dorsally and ventrally by prominent blunt ridges (Fig. 10). This contrasts with TTU-P10076, in which the rim is vertical in the posterior section only, and with the thin and sharp rim in TTU-P10077. The medial lamella in TTU-P10074 is also much more thickened by a prominent ventral bulge along the entire medial rim of the postorbital–squamosal bar.
2.7.6. Palatine
Although the same structures of the palatal section of the palatine are principally present in TTU-P10074, they are differently developed. The vault of the anterior section of the bone is less marked, its apex points ventromedially and the palatine terminates level with the choana. The palatine ridge is lower, points ventrally and is restricted to the central area of the bone. The postchoanal parts of the palatines are parallel-sided, showing a much smaller palatine lamella that is curved upward rather than horizontal and thus overhangs only about two third of the width of the palatal vault. The palatine lamella is significantly less sculptured.
2.7.7. Supraoccipital (Fig. 10)
A central ridge on the supraoccipital shelf is absent in TTU-P10074 and the lateral tips of the supraoccipital terminate more medial to the postorbital fenestrae. The sutures between the supraoccipital and the exoccipitals cross the edge of the suparoccipital shelf and pass downward and inwards into the foramen magnum, forming a 4–6 mm thin strip on the roof of the canal for the medulla oblongata.
3. Discussion of anatomical observations
Narial and rostral crest. Given the high variability in the shape of phytosaur rostra, it is necessary to define criteria for identifying a crest and to classify crests to eliminate a potential source of confusion. For example, Zeigler et al. (Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b) distinguish, in the Canjilon Quarry phytosaur assemblage, between a morph with a tall, longer and robust crest (M. buceros) and a morph with an abrupt, distinct narial crest (M. pristinus). Most previous authors (e.g., Camp Reference Camp1930; Gregory Reference Gregory1962; Ballew Reference Ballew, Lucas and Hunt1989; Long & Murry Reference Long and Murry1995) regarded specimens of M. pristinus as crest-less phytosaurs. Stocker (Reference Stocker2010) categorised phytosaur crests as narial and rostral crests, a crest being any elevation that rises above the area anterior and posterior to it. Narial crests were defined as dorsal elevations both anterior and posterior to the nares. Narial crests can be in form of a narial cone raised above the level of the remainder of the snout, but also include prenarial crests, redefined as “no or very little change in dorsal elevation of the area of the skull roof posterior to the nares, but a decrease in dorsal elevation of the rostrum anterior to the nares” (Stocker Reference Stocker2010, supplementum, unpaginated). She subdivided rostral crests into four types: (1) premaxillary crests; (2) dorsoventrally undulating crests including ‘partial’ crests that do not extend anteriorly to the terminal rosette of the premaxillae; (3) crests that are continuous, sloping steeply to the terminal rosette; and (4) crests that are continuous at a level from the nares to the terminal rosette.
We adopt here Stocker's (Reference Stocker2010) classification of phytosaur crests. According to her criteria, all specimens described here have narial crests because there are elevated areas anterior and posterior to the nares or, in other words, the nares are situated in a cone that is elevated above the surface of the snout. Both TTU-P10076 and TTU-P10077 have partial rostral crests, i.e., elevations that rise above the surrounding rostral surface but do not reach the anterior tip of the premaxillae. However, Stocker's categories do not describe the entire morphological variation seen in pseudopalatine phytosaurs. Characters such as visual demarcation from the tooth-bearing part of the rostrum in the form of a longitudinal furrow, depression, or a flattened ‘shelf’-area lateral to the crest, crest profile, or mediolateral thickness are not considered. TTU-P10076 has a very short partial rostral crest below the narial cone, which is outlined by two lateral depressions. TTU-P10077 shows a much extended, undulating and, in the anterior portion, sharp partial rostral crest, outlined laterally by large depressions. Stocker (Reference Stocker2010) probably defined the categories largely to accommodate leptosuchomorph phytosaurs, the primary subjects of her study, and she discussed other variable crest features in the description without incorporating them in the classification. We do not wish to modify or expand Stocker's classification here; this should be left to a comprehensive study of the crest morphology in all phytosaur taxa. In this study, we prefer a reductive approach to character construction (Wilkinson Reference Wilkinson1995), and compartmentalise rostral crests into four discrete characters (Table 1: characters 1, 3, 4, 5) that adequately and objectively describe the variation in crest morphology in Pseudopalatinae.
Septomaxilla and paranasal. Two discrete ossifications are identified in the narial and immediate prenarial regions of TTU-P10076, TTU-P10077 and, possibly, in TTU-P10074. Paired separate elements anterior to the nasal openings that incorporate most of the internasal septum were first mentioned by Meyer (Reference Meyer1865) for Nicrosaurus meyeri NHMUK 42745, later identified as septomaxillae by Huene (Reference Huene1909) in Mystriosuchus westphali GPIT 261/001 and, since then, confirmed as being present in every phytosaur taxon. The topology and the relationships with the premaxilla and nasal show that the medial element in the specimens described here is the same element, the septomaxilla. Sereno (Reference Sereno1991), Senter (Reference Senter2002) and Stocker (Reference Stocker2010) questioned the homology of this bone because of topological, functional and phylogenetic incongruence with the septomaxilla. The arguments were that the septomaxilla in lepidosauromorphs (1) is not exposed on the skull surface; (2) is located on the floor rather than the roof of the nasal passage; and (3) does contact the maxilla, unlike the septomaxilla in phytosaurs. We wish to point out that the external nasal openings of phytosaurs are highly modified with respect to shape, orientation and position, as is the configuration of the nasal bones, the choana and the palate. It seems rather unlikely that details of the topological relationships of a bone present in this region are not affected. The septomaxilla also undergoes substantial evolutionary modifications in synapsids, including a facial exposure (Wible et al. Reference Wible, Miao and Hopson1990). Senter (Reference Senter2002) argued that (4) the septomaxilla provides support for the vomeronasal (Jacobson's) organ, for which there is no independent evidence in phytosaurs. The presence of a septomaxilla, however, is not necessarily correlated with the development of the vomeronasal organ in, for example, urodeles (Trueb Reference Trueb, Hanken and Hall1993, table 6.2) and it may have acquired this structurally supportive function only secondarily in squamates (Wible et al. Reference Wible, Miao and Hopson1990, fig. 1). Finally, (5) a septomaxilla is apparently absent in any other archosauromorph except the much more basal Proterosuchus (Senter Reference Senter2002); i.e., there is no phylogenetic continuity via Erythrosuchidae and Euparkeria to phytosaurs. With respect to this argument, we wish to point out that a usually delicate intranasal element in groups in which the narial area has received little attention could have been easily lost by taphonomic processes, or overlooked. Hence, we see no sufficient reason to doubt the homology of the bone in question with the septomaxilla in other tetrapods. We interpret the expansion of the septomaxilla, including an exposure of the bone on the prenarial surface and the exclusion of the premaxilla and the nasal from the internasal septum, as an apomorphic development of Phytosauria.
The lateral ossification situated in the anterolateral corner of the naris between the nasal and premaxilla has no precedence in amniotes. It certainly represents a neomorphic bone that evolved within Phytosauria. We introduce the name paranasal (from Greek ‘para’ – beside or alongside of) for this element. All other descriptions of phytosaur crania agree in that the same space is occupied by the nasal. There is no evidence for a paranasal in any specimen of Nicrosaurus (Hungerbühler Reference Hungerbühler1998), or Mystriosuchus (McGregor Reference McGregor1906; Hungerbühler Reference Hungerbühler2002), and we are confident that such an ossification is absent in the immediate outgroups of Machaeroprosopus hypothesised by Hungerbühler (Reference Hungerbühler2002). As described for TTU-P10074, the prenarial area is difficult to interpret in many specimens of Machaeroprosopus, and the sutural configuration was only confirmed in excellently preserved specimens. We suspect that a reexamination will reveal that the paranasal is much more common in Machaeroprosopus. For this reason, we do not suggest the paranasal as diagnostic for M. lottorum. In all three specimens under study, the paranasal is correlated with a columnar appearance of the anterolateral rim of the nares, and such morphology may represent an indicator for the presence of the element in other specimens.
Braincase
Several features of the braincase and related structures of Machaeroprosopus are described here for the first time, or previous description are confirmed for this taxon. Many of these features are, however, difficult to evaluate critically with respect to homology and phylogenetic significance, in the absence of detailed studies in most phytosaurs.
Contribution of squamosal to lateral wall of braincase
An anterior lamella of the parietal process of the squamosal, that extends far onto the lateral wall of the braincase and contacts the prootic in M. lottorum (Fig. 22), invalidates this feature as an autapomorphy for Mystriosuchus westphali (Hungerbühler Reference Hungerbühler2002). Parker & Irmis (Reference Parker and Irmis2006) confused this character with the ‘squamosal flange’ (see description) and its distribution (char. 26), but correctly stated that the anterior lamella is present, but does not reach the prootic in M. jablonskiae and in an isolated pseudopalatine braincase they referred to M. cf. pristinus. This indicates that the anterior extent of the squamosal on the braincase is indeed variable within Machaeroprosopus and might be a taxonomically informative feature.
Epipterygoid. Among phytosaurs, the morphology of the epipterygoid is known in sufficient detail only for Smilosuchus gregorii (UCMP 27200) and Mystriosuchus westphali (GPIT 261/001). The epipterygoid of Machaeroprosopus differs in general shape and, in particular, in the mode of articulation with the pterygoid. As described by Camp (Reference Camp1930) and Huene (Reference Huene1909, Reference Huene1911) and based on own observations (AH pers. obs. 2000, 2002), the epipterygoid is a much thinner element in Smilosuchus and Mystriosuchus, with sharp lateral and medial edges. As far as preserved, the epipterygoid of Mystriosuchus is a largely antero–posteriorly elongated rod, and only the dorsal section is twisted and faces anterolaterally. Smilosuchus and Machaeroprosopus share a derived feature, the transverse orientation of the epipterygoid with an anteriorly-facing side. In the ventral section, the epipterygoid of Smilosuchus is distinctly twisted along the vertical axis and develops a thin, posteriorly-directed lateral flange, resulting in a deeply grooved posterior side of the element. The base of the epipterygoid is bifurcated and wraps around the epipterygoid process of the pterygoid, the groove receiving the anterior edge of that process. The epipterygoid process thus forms a firm buttress for the epipterygoid. In all epipterygoids except Machaeroprosopus, a process extends posteriorly along the quadrate wing of the pterygoid, and the epipterygoid and the pterygoid are clearly unfused, separate elements.
Primitively, the epipterygoid articulates dorsally with the parietal close to the skull roof, and this condition is retained in the archosauriform Proterosuchus (Broili & Schröder Reference Broili and Schröder1934; Clark et al. Reference Clark, Welman, Gauthier and Parrish1993). In more derived archosaurs, the dorsal insertion of the epipterygoid, when ossified and preserved, probably shifted onto the laterosphenoid (Gower & Sennikov Reference Gower and Sennikov1996). The exact location of the dorsal articulation of the epipterygoid with the braincase in Machaeroprosopus is uncertain. Chatterjee (Reference Chatterjee1978) interpreted a pit in the dorsal section of the laterosphenoid behind the capitate process as the epipterygoid facet in Parasuchus hislopi; a comparable recess in P-10076 is here identified as the exit of a blood vessel. In Smilosuchus gregorii, the epipterygoid inserts in a pit on the anterolateral face of the laterosphenoid at the base of the capitate process (Camp Reference Camp1930, left side; in contrast to his figures 36, 38 and 39, only the base of the right epipterygoid is now preserved). There is a shallow rugose depression in P-10076 in a similar position as in Smilosuchus gregorii. Were the epipterygoid to insert here, the element would have been considerably recurved (unlike any known epipterygoid in phytosaurs) and, for the spatial conditions, would have to have been buttressed by the anteroventral process of the laterosphenoid, thereby closing the exit of the cleft behind this process to a foramen. An alternative insertion is the crescentic groove on the ventral side of the anterior process of the laterosphenoid, above the optical foramen, which is exactly in extension of the preserved base of the epipterygoid, as it is in Mystriosuchus westphali (Hungerbühler Reference Hungerbühler2002, fig. 8). Gower (Reference Gower1997) suggested a pit in a similar position as an articulation of the epipterygoid of Erythrosuchus. We think this is the most likely insertion of the epipterygoid. The posterior shift of the epipterygoid on the laterosphenoid in Smilosuchus gregorii might be connected to the strong anteroposterior compression of the braincase in this taxon.
In comparison to other phytosaurs, the epipterygoid of Machaeroprosopus shows several differences. First, it forms a thick, rather than blade-like rod. The preservation as a hollow tube in TTU-P10076 suggests that only the peripheral parts of the epipterygoid were ossified, and the core may have been infilled with matrix after decay of the cartilage. Secondly, the entire transversely expanded side of the epipterygoid faces anteriorly and, in conjunction with the different orientation, the posterior basal process is reduced. Thirdly, the ossified epipterygoid is spatially decoupled from the epipterygoid process of the pterygoid. The ragged outline of the free epipterygoid process suggests that the structure was topped by a significant cartilaginous extension, and it might have taken the place of the epipterygoid in closing the primary skull wall. It might even represent an ossification within the palatoquadrate, rather than a primary part of the pterygoid, which became fused with this dermal bone.
Parasphenoid process. TTU-P10076 and P-10074 are the first phytosaur skulls to preserve this delicate structure (Fig. 22, ‘psp.p’). Remnants seems to be also present in Smilosuchus gregorii UCMP 27200 (AH pers. obs.), but were apparently omitted in Camp's (Reference Camp1930, fig. 37) braincase reconstruction. From its shape and position, it is likely that at least the top part of the projections represents ossifications embedded in the interorbital septum.
Gower & Walker (Reference Gower and Walker2002, p.13) described a “subvertical prow of the parabasisphenoid” in front of the hypophyseal fossa for the aetosaur Stagonolepis robertsoni. They interpreted tentatively a pair of foramina in the front section of the ‘prow’, associated with horizontal ledges as exits of the orbital arteries out of the hypophyseal fossa. The description and illustration (Gower & Walker Reference Gower and Walker2002, fig. 3) is essentially similar to the parasphenoid processes and the associated foramen in TTU-P10076, although there is no horizontal ledge. It is conceivable that the orbital arteries emerged from the hypophyseal fossa between the parasphenoid processes and travelled into the orbital cavities via foramina in the ossified walls of the cultriform process (but see discussion in Gower Reference Gower2002). Otherwise, parasphenoid processes among archosaurs have only been noted for dromaeosaurid and troodontid dinosaurs (Colbert & Russell Reference Colbert and Russel1969; Currie Reference Currie1985).
Presphenoid and orbitosphenoid. Two elements in addition to the laterosphenoid and the prootic in the anterior braincase have been reported as present for some phytosaur taxa.
A discrete ossification below the optical foramen in phytosaurs was first recognised and described as presphenoid by Camp (Reference Camp1930) in the type specimen of Smilosuchus gregorii (UCMP 27200). The presphenoid illustrated for the basal phytosaur Parasuchus hislopi in Chatterjee (Reference Chatterjee1978) is not preserved in the specimen, but was postulated on the basis of sutural facets on the laterosphenoids in accordance with Camp's (Reference Camp1930) interpretation in UCMP 27200 (S. Chatterjee, pers. comm. 2002). A presphenoid is, however, present in another basal form, Paleorhinus scurriensis TTU-P8090 (Simpson Reference Simpson1998, named ‘orbitosphenoid’; AH, pers. obs.). So far, most studies of the braincase in other taxa have failed to identify the element (Lees Reference Lees1907; Mehl Reference Mehl1928; Case Reference Case1929; Case & White Reference Case and White1934; Colbert Reference Colbert1947), and one of us questioned the existence of this bone altogether (Hungerbühler Reference Hungerbühler2002). TTU-P10076 and TTU-P10074 are, to our knowledge, the only other phytosaur skulls that show well-preserved presphenoids.
Frequently, this area of the braincase is not ossified in archosaurs, leaving a gap (e.g., Euparkeria; Clark et al. Reference Clark, Welman, Gauthier and Parrish1993), or a substantially enlarged ‘optical foramen’ (e.g., Plateosaurus, Galton Reference Galton1985). Reports of a presphenoid in other pseudosuchian archosaurs are questionable. After repreparation of the type specimen, the presence of a presphenoid in the rauisuchid Postosuchus as reconstructed by Chatterjee (Reference Chatterjee1985) is unsubstantiated, and should be regarded as a part of the anterodorsal process of the prootic (Weinbaum Reference Weinbaum2011). The evidence for a presphenoid in Saurosuchus (Alcober Reference Alcober2000) was considered doubtful by Gower (Reference Gower2002) because the demarcation from the laterosphenoid seems to be a fracture, and the postulated element borders, unlike a presphenoid, the trigeminal foramen. In addition, the ‘presphenoid’ of Saurosuchus (Alcober Reference Alcober2000, figs 7, 8) shows a process of the same shape and in the same position as the anteroventral process in TTU-P10076, which is definitely part of the laterosphenoid (Fig. 22, ‘vp.lsp’). For ornithodiran archosaurs, the only presphenoid described is in Ceratosaurus (Madsen & Welles Reference Madsen and Welles2000), but there authors present too little information to allow a comparison. An element called the presphenoid in troodontid theropods (Currie Reference Currie1985) more likely represents, according to its spatial relationships, an ossification of the ethmoidal series.
On the basis of three specimens of Smilosuchus, Camp (Reference Camp1942) postulated that the area of the laterosphenoid as described for other phytosaurs is actually subdivided into a laterosphenoid sensu stricto, an anteriorly placed orbitosphenoid, and a ‘septosphenoid’ enclosing the anterior part of the olfactory tract. He suggested that these three elements fuse through ontogeny into a single element, and claimed that sutures only persist externally in juveniles, but are obscured by superficial bone layers in adult specimens. An orbitosphenoid in a corresponding position to that of Smilosuchus was also identified in Mystriosuchus westphali (Hungerbühler Reference Hungerbühler2002).
For other archosaurs, an overview is given in Hungerbühler (Reference Hungerbühler2002) and we restrict our comments to some supplementary information. An orbitosphenoid ossification in front of the laterosphenoid is well established for plateosaurid (Galton Reference Galton1985) and a variety of theropod dinosaurs (e.g. Madsen Reference Madsen1976; Currie Reference Currie1985; Currie & Zhao Reference Currie and Zhao1993; Madsen & Welles Reference Madsen and Welles2000). This is a paired ossification, with the centre of the bone dorsal to the optical nerve. A medially curved, thin ventral process usually extends downward to a certain degree, forming the lateral rim of the optical foramen. The process may meet its counterpart at the midline (e.g., Allosaurus fragilis, Madsen 1978; confirmed by AH pers. obs.), and the optical foramen is completely surrounded by the orbitosphenoids. Among pseudosuchian archosaurs, only Parrish (Reference Parrish1994) reconstructed an orbitosphenoid in the braincase of the aetosaur Longosuchus, but his evidence could not be confirmed by Gower & Walker (Reference Gower and Walker2002), nor was an orbitosphenoid identified in other taxa (Small Reference Small2002).
This survey shows that two elements may ossify in the anterior section of the braincase of archosaurs around the optical nerve, in addition to and substituting the prootic or the laterosphenoid in this region. These bones are not homologue, as indicated by their topological incongruence. A dorsal element, usually called the orbitosphenoid, is paired and centred above the optical nerve, although it may extend downward along the sides of the foramen. A ‘slender’ or ‘ventral process’ of the laterosphenoid in Euparkeria (Clark et al. Reference Clark, Welman, Gauthier and Parrish1993) is topologically consistent with the ventral process of the orbitosphenoid in dinosaurs. A ventral ossification, the presphenoid, may ossify below the optical nerve, although a dorsal process may occupy the same position lateral to the optical nerve as the process of the orbitosphenoid (or laterosphenoid). Whether these elements are equivalent ossifications of the orbitosphenoid and presphenoid (both names are based on ossifications in the mammalian skull; Romer Reference Romer1956) is in need of further study, but we retain these names, which are well embedded in the literature, for the time being.
TTU-P10076 and TTU-P10074 demonstrate the ossification of a distinct presphenoid in Machaeroprosopus lottorum. The occurrence of this element across various phytosaur clades (Paleorhinus, Smilosuchus and Machaeroprosopus) suggests that this element regularly ossified in phytosaurs. According to Camp (Reference Camp1930), the presphenoid forms much of the anterior inner wall of the braincase, and is thus a much more extensive element than it appears to be externally. Because there is no unambiguous evidence for this element in other archosaurs (with the possible exception of the theropod dinosaur Ceratosaurus; Madsen & Welles Reference Madsen and Welles2000), the ossification of a presphenoid is most likely an autapomorphy of Phytosauria. The evidence for a discrete orbitosphenoid in phytosaurs (and pseudosuchian archosaurs) is meagre, in contrast to a more widespread occurrence in dinosaurian archosaurs. Camp's (Reference Camp1942) suggestions are intriguing and offer an explanation for the erratic observations of a discrete orbitosphenoid ossification in phytosaurs (and, more generally, in other archosaurs), but his evidence for a subdivision of the laterosphenoid needs to be confirmed in his specimens and across more taxa.
Abducens (VI) foramen. In Parasuchus hislopi, the abducens nerve (VI) exits on the anterior face of the braincase on the suture between the prootic and the parabasisphenoid, at the level of the lower one-third of the hypophyseal gap to the medial side and the trigeminal foramen to the rear (Chatterjee Reference Chatterjee1978, fig. 5a). There is a broader matrix-filled notch in the same position on the right side of TTU-P10076 (Fig. 21), but such a notch is clearly absent on the well-preserved left side (Fig. 18). The structure on the right side is more likely to be the result of a partial separation of the prootic from the parabasisphenoid along their suture. Camp (Reference Camp1930) could not identify a separate abducens foramen for Smilosuchus gregorii either, and assumed that the abducens nerve exited together with the trigeminal nerve through the trigeminal foramen. This could indicate that in derived phytosaurids (Smilosuchus, Machaeroprosopus) the course of the abducens nerve out of the braincase has been modified relative to the plesiomorphic pattern in basal phytosaurs.
4. Phylogenetic evaluation of Machaeroprosopus lottorum
4.1. Design and purpose of the parsimony analysis
We assess the phylogenetic position of Machaeroprosopus lottorum (TTU-P10076 and TTU-P10077) and Machaeroprosopus sp. TTU-P10074 by parsimony analysis in the framework of the systematics of North American pseudopalatine phytosaurs, as suggested in the reviews of Ballew (Reference Ballew, Lucas and Hunt1989), Hunt & Lucas (Reference Hunt, Lucas, Lucas and Morales1993) and Parker & Irmis (Reference Parker and Irmis2006). We emphasise that we do not attempt a revision of the North American pseudopalatine phytosaurs at this stage, for reasons given below. Consequently, we rely on previous studies for the validity of the OTUs, and the data matrix does not include shared derived characters which exclusively support the monophyly of the ingroup, but are uninformative with respect to ingroup interrelationships. The conclusions of our analysis presented here are tentative, and a more comprehensive analysis is in preparation.
4.2. Choice of outgroups
The trees are rooted and the ancestral states of the characters are identified by comparison with Mystriosuchus planirostris (Fraas Reference Fraas1896) and Mystriosuchus westphali Hungerbühler & Hunt, Reference Hungerbühler and Hunt2000, which most parsimoniously form the sister-group to the ingroup (Hungerbühler Reference Hungerbühler2002). However, several features of Mystriosuchus, in connection with the supratemporal fenestra and the temporal region, most likely do not represent ancestral states, but are reversals or independent developments in this highly derived lineage (see Hungerbühler Reference Hungerbühler2002). The close relationship with the North American pseudopalatine phytosaurs has also been contested (e.g., Hunt Reference Hunt1994; Long & Murry Reference Long and Murry1995). For this reason, we include Nicrosaurus, represented by the type species Nicrosaurus kapffi (Meyer Reference Meyer1860), as an additional outgroup that currently is well-supported as the most basal pseudopalatine phytosaur (Ballew Reference Ballew, Lucas and Hunt1989; Hunt Reference Hunt1994; Long & Murry Reference Long and Murry1995; Hungerbühler Reference Hungerbühler2002).
4.3. The ingroup
Apart from the skulls described in this study, the ingroup includes the six North American pseudopalatine species that are regarded as valid in the most recent reviews (Ballew Reference Ballew, Lucas and Hunt1989; Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993; Long & Murry Reference Long and Murry1995, in parts; Hungerbühler Reference Hungerbühler2002; Parker & Irmis Reference Parker and Irmis2006). Character states were coded by examining the holotype of each OTU only, to avoid subjectivity that might result from including specimens a priori in the hypodigm.
Machaeroprosopus pristinus (Mehl Reference Mehl1928)
Holotype. UMo 525, complete cranium
Type horizon. Petrified Forest Member, Chinle Formation
Type locality. unknown locality in Painted Desert, Petrified Forest National Park, Apache County, AZ (Irmis Reference Irmis2005)
M. pristinus is the type species of Pseudopalatus Mehl, Reference Mehl1928. Personal inspection by one of us (AH 2000) revealed that the specimen falls rather short from being “of better than average preservation” (Mehl Reference Mehl1928, p. 7). The postorbital region of the skull has evidently been assembled from numerous fragments, and the shape and the proportions of some features in the temporal region seem to be distorted. The nasal area and the left squamosal are in parts, and the top of the parietal–supraoccipital complex fully, reconstructed in plaster. This raises the question of whether the morphology of these features in the holotype is real, deformational or artificial, and we regard some of the data taken from Mehl's (Reference Mehl1928) drawings, or from subsequent photographs (Long & Murry Reference Long and Murry1995, figs 40B, 41A, B), as unreliable. Some characters of the holotype are at variance with those seen in other specimens that have been included in the hypodigm (Ballew Reference Ballew, Lucas and Hunt1989; Long & Murry Reference Long and Murry1995; Zeigler et al. Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b). What remains of the original palate does not support Mehl's (1928) reconstruction of a fully ossified secondary palate, as already suspected by Camp (Reference Camp1930). The characters for M. pristinus are scored ‘as is’ in the type specimen (Table 1), and those from reconstructed areas are scored as ‘indeterminate’. It might be necessary to reconsider some of the scores that might have been affected by deformation (e.g., characters 8, 17, 18) pending the results of a thorough reassessment of the specimen. Zeigler et al. (Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b) regarded M. pristinus as the female morph of M. buceros and thus a junior synonym of M. buceros. We chose the conservative approach to retain M. pristinus for the analysis, also in order to test their interpretation as sexual morph by conducting a parsimony analysis. M. pristinus was diagnosed by Ballew (1989) by five autapomorphies: (1) proportionally very long rostrum; (2) mound-shaped, convex and thickened nasals around nares; (3) sharp descent of nasals on the rostrum; (4) homodont dentition; and (5) subcircular antorbital fenestra.
Machaeroprosopus buceros (Cope Reference Cope1881)
Holotype. AMNH 2318, cranium
Type horizon. upper Petrified Forest Formation (Lucas et al. Reference Lucas, Heckert, Zeigler, Hunt, Heckert and Lucas2002)
Type locality. near Orphan Mesa, Rio Arriba County, NM (Lucas et al. Reference Lucas, Heckert, Zeigler, Hunt, Heckert and Lucas2002)
AMNH 2318 is a poorly preserved skull (see Lucas et al. Reference Lucas, Heckert, Zeigler, Hunt, Heckert and Lucas2002), which accounts for the amount of missing data. Originally erected as Belodon buceros, over time the species was assigned to no less than five different genera until Parker et al. (Reference Parker, Hungerbühler and Martz2013, this volume) determined that M. buceros is the type species of Machaeroprosopus. Long & Murry (Reference Long and Murry1995) designated Belodon buceros as the type species for their genus Arribasuchus. Hunt (Reference Hunt, Lucas and Morales1993, Reference Hunt1994; Hunt in Hunt et al. Reference Hunt, Lucas, Heckert, Heckert and Lucas2002) considered the taxon a species of Nicrosaurus, because the type specimen shows a dorsolaterally compressed posterior process of the squamosal, slit-like supratemporal openings, and the narial openings are situated below the level of the skull roof. However, the squamosals show numerous similarities with those of other species of Machaeroprosopus (see Table 1), including the dorsoventrally expanded posterior process, and we regard the shape of the supratemporal fenestrae as ambiguous because of damage. The position of the nares was correctly identified by Hunt, although the narial rims are severely abraded and parts of the narial area are crushed down. This represents, however, the plesiomorphic state of this character. The type specimen shares no derived feature with the type species of Nicrosaurus, N. kapffi (Meyer Reference Meyer1860) to the exclusion of the other pseudopalatine taxa, and consequently we reject this assignment. Ballew (Reference Ballew, Lucas and Hunt1989) identified three autapomorphies, which are posttemporal fenestra extending laterally above the opisthotic, labio–lingually compressed posterior maxillary teeth with a mesial ridge, and a V-shaped rostral crest.
Machaeroprosopus mccauleyi (Ballew Reference Ballew, Lucas and Hunt1989)
Holotype. UCMP 126999, cranium with mandible
Type horizon. Petrified Forest Formation
Type locality. Billings Gap, Apache County, AZ
Ballew (Reference Ballew, Lucas and Hunt1989) lists five autapomorphic characters in support of the species: (1) squamosal with distinct triangular outline without knob-like process; (2) lateral portion of opisthotic thin and elongate; (3) posttemporal fenestra large because of a medial extension; (4) basioccipital head relatively large; (5) basioccipital neck relatively short. Hunt et al. (Reference Hunt, Lucas and Spielmann2006) suggested two additional diagnostic characters: the lateral margins of the skull flare at 60° in posterior view and the presence of a ‘complete’ rostral crest.
Machaeroprosopus jablonskiae (Parker & Irmis Reference Parker and Irmis2006)
Holotype. PEFO 31207, posterior part of skull roof and braincase
Type horizon. Jim Camp Wash beds of Sonsela Member, Chinle Formation
Type locality. near Mountain Lion Mesa, Petrified Forest National Park, Apache County, AZ (Parker & Martz Reference Parker and Martz2011).
Parker & Irmis (Reference Parker and Irmis2006) identified a bevelled edge on the antero-medial edge of the postorbital–squamosal bar as a single autapomorphic character in support of the species.
Machaeroprosopus gregorii (Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993)
Holotype. YPM 3294, cranium
Type horizon. Duke Ranch Member, Redonda Formation
Type locality. Shark Tooth Hill, Quay County, NM (Spielmann & Lucas Reference Spielmann and Lucas2012).
M. gregorii is the type species of Redondasaurus. Long & Murry (Reference Long and Murry1995) did not accept the validity of Redondasaurus and referred M. gregorii to Pseudopalatus pristinus. In the type specimen, the entire narial area, the left side of the snout, the anterior two thirds of the right premaxilla, and most of the palate are missing. M. gregorii was diagnosed as a species of Redondasaurus, therefore with dorsally concealed supratemporal fenestrae, that is characterised by the absence of a ‘rostral’ crest (Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993, p. 193). Recently, Spielmann & Lucas (Reference Spielmann and Lucas2012) rediagnosed the species (in which they include M. bermani) by, in addition to the supratemporal fenestra concealed in dorsal view: (1) reduced antorbital fenestra; (2) a prominent pre-infratemporal shelf; (3) the septomaxilla forming the anterolateral half of the external naris; (4) rim of the orbit thickened; (5) inflated posterior part of nasal; and (6) thickened dorsal osteoderms.
Machaeroprosopus bermani (Hunt &Lucas Reference Hunt, Lucas, Lucas and Morales1993)
Holotype. CM 69727, cranium with strongly deformed mandible lacking the anterior half of the symphysis
Type horizon. ‘siltstone member’ of Chinle Formation (Nesbitt Reference Nesbitt2011) or Rock Point Formation (Spielmann & Lucas Reference Spielmann and Lucas2012)
Type locality.Coelophysis Quarry, Rio Arriba County, NM; the skull was recovered above the actual Coelophysis bonebed when the quarry was reopened in the early1980s and the overburden was removed (D. Berman pers. comm. 2001).
The type specimen is only superficially prepared, and the sutural configuration and details of some structures, such as the posttemporal fenestra and the parietal–supraoccipital complex, are currently obscured. M. bermani was originally diagnosed a species of Redondasaurus in possession of a ‘partial’ rostral crest. Spielmann & Lucas (Reference Spielmann and Lucas2012) regarded the holotype as the male morphotype of M. gregorii, and consequently M. bermani as a junior synonym of that species.
4.4. Methods and analytical procedures
The character-taxon matrix and a list of the character states are presented in Table 1 and the Appendix, respectively. The data were subjected to a parsimony analysis using the software program PAUP 3.1.1 (Swofford & Begle Reference Swofford and Begle1993), run on a PowerMac computer to produce the most parsimonious branching pattern. Multistate characters were left unordered, and all characters were initially treated as equally weighted. The analyses employed the ‘exact’ branch-and-bound algorithm (addition sequence ‘furthest’ and the option ‘collapse zero-length branches’ activated), which guarantees finding all most parsimonious trees (MPTs). The data were organised and tree statistics were calculated in MacClade 3.07 (Maddison & Maddison Reference Maddison and Maddison1997); TL increased by one step relative to PAUP because C21 for Machaeroprosopus bermani is treated as ‘polymorphism’. To evaluate the support of the individual branches of the MPTs, three methods were employed. First, a bootstrap analysis was performed (10000 replications; search settings as above) and, secondly, Bremer support was computed for each node using the software Autodecay 4.0 (Eriksson Reference Eriksson1998). Thirdly, a number of subsequent runs were performed, in which several constraints were enforced upon the MPTs retained by the parsimony analysis. The constraints included clades of terminal taxa that were not present in the MPTs from the initial analysis, but have been hypothesised by previous workers. The minimum number of additional steps required to produce such suboptimal trees when compared to the MPTs without the constraint is then used as a relative measurement for the probability, how far the constraint clades represent a reasonable option to be considered in reconstructing the interrelationships of the taxa and specimens.
The initial run recovered six MPTs with tree lengths (TL) of 127 steps, consistency indices (CI) of 0.5, retention indices (RI) of 0.51, and rescaled consistency indices (RCI) of 0.25. An Adams consensus tree (Fig. 25A) shows that there is little congruence among the trees. The analysis indicates that not only M. mccauleyi and M. jablonskiae, but also M. bermani, are basal pseudopalatine taxa, whereas M. pristinus, M. buceros, M. gregorii and the specimens under study are positioned more crownwards.

Figure 25 (A) Adams consensus tree of six most parsimonious trees (MPTs) obtained by parsimony analysis of character-taxon matrix, Table 1. (B) Strict consensus tree of two MPTs after exclusion of characters 1, 3, 4 and 5. (C) Single MPT after successive approximate character weighting (SAWC). Left values represent bootstrap percentages >50; right values Bremer support.
In an attempt to remove one potential source of non-taxonomic variation, we tested the suggestions of sexual dimorphism proposed by Zeigler et al. (Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b) and excluded the characters 1, 3, 4, and 5, which together describe the presence and the shape of a rostral crest, and reran the analysis with the same settings. This resulted in a drop of the number of the MPTs to two (Fig. 25B) with a TL of 109 steps and slightly improved statistical values (CI 0.52; RI 0.55; RCI 0.29). For further improvement, a series of runs were undertaken with successive approximations character weighting (SACW; see Wilkinson & Benton Reference Wilkinson and Benton1996). In this method, the characters are re-weighted a posteriori according to a measurement of fit as determined by the previous run, and the data is then re-analysed. Re-weighting and re-analysing continues until the weight no longer changes, or identical trees are found in a consecutive run (Swofford & Begle Reference Swofford and Begle1993). Thus, character incongruences that result in multiple MTPs are expected to be resolved by successively low-weighting of homoplastic characters. As a measurement of fit, the RI was chosen and the baseweight was set to 1000. SACW recovered a single tree, which was identical to one of the two MPTs retrieved in the previous run (Fig. 25B). A bootstrap analysis was undertaken and Bremer support was calculated for the less inclusive data matrix, and mapped onto the SACW tree (Fig. 25C).
Both MPTs now show M. pristinus+M. buceros, as well as TTU-P10076+TTU-P10077, as sister-taxa. (TTU-P10076+ TTU-P10077)+M. gregorii, the type species of Redondasaurus, form a clade. TTU-P10074 is recovered as immediate outgroup to this clade, followed by (M. pristinus+M. buceros) closer to the root of the tree.
5. Conclusions on sexual dimorphism
The exclusion of a set of characters (rostral crest presence and morphology) that has been suggested to represent sexually dimorphic features in pseudopalatine phytosaurs resulted in an improvement of the resolution of the phylogenetic relationships. The initial hypothesis of Zeigler et al. (Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b), on the basis of 11 skulls from the Canjilon quarry, and similar evidence from a phytosaur assemblage from the time-equivalent Snyder quarry (Zeigler et al. Reference Zeigler, Heckert, Lucas, Zeigler, Heckert and Lucas2003a) in north-central New Mexico was based on three lines of reasoning. First, taphonomic studies of the Machaeroprosopus assemblages from the Canjilon and Snyder quarries (Hunt & Downs Reference Hunt, Downs, Heckert and Lucas2002; Zeigler Reference Zeigler, Zeigler, Heckert and Lucas2003) found that both represent a snap-shot of a contemporaneous population. Secondly, a morphological comparison of the Machaeroprosopus assemblages suggested to the present authors that the only distinction between the specimens lies in the shape of the rostral crest. Lastly, Zeigler et al. (Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b) argued that dimorphic structures in fossil vertebrates that were interpreted as sexually dimorphic are cranial structures suitable for visual display, and phytosaur crests fit this pattern. An analysis of the Canjilon Quarry data, however, did not produce a statistically significant signal, because of the small sample size (K. Zeigler pers. comm. 2002). Irmis (Reference Irmis2005) pointed out weaknesses of the sexual dimorphism hypothesis: other possible explanations of dimorphism such as sympatric closely related species or resource polymorphism have not been ruled out; the hypothesis does not test apparent dimorphism in other phytosaur genera; and does not address the presence of third morphs such as M. mccauleyi and Smilosuchus gregorii, with shorter, much broader, fully-crested snouts (brachyrostral type of Hunt Reference Hunt, Lucas and Hunt1989). Hunt et al. (Reference Hunt, Lucas and Spielmann2006) suggested that sexual dimorphism is expressed in M. mccauleyi by an increased robustness of the rostrum in males, but because their two ‘male’ voucher specimens are significantly larger than the ‘female’ holotype, the more massive snout can be similarly well explained as an advanced stage of ontogeny.
Our analysis lends further support for Zeigler et al.'s (Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b) hypothesis. It produced a similar result on the basis of a parsimony analysis of a different data set, although M. pristinus and M. buceros differ evidently more than only in rostral crest characters (see Appendix; the pairwise distance between both specimens corrected for missing data is 0·393, and M. pristinus and M. buceros are more distant to each other than, e.g., M. bermani from the crested skull TTU-P10077 [0·389]). Moreover, the data is exclusively derived from the actual name-bearing rather than from referred specimens. We thus conclude that it is more likely that M. pristinus and M. buceros are two morphotypes of the same species than two separate species. This hypothesis is even better supported for M. lottorum TTU-P10076 and TTU-P10077.
European and North American Upper Triassic deposits have yielded contemporaneous phytosaurs that share numerous characters, and differ in overall appearance mainly by the absence, presence and the nature of a rostral crest and the general build of the skull (see Hungerbühler Reference Hungerbühler2002, table 1). We restrict our conclusion on sexual dimorphic characters explicitly to M. buceros+M. pristinus and M. lottorum. Our main concern is that the explanation as sexual morphs as suggested here, and by Zeigler et al. (Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas and Heckert2003b), might be less critically and universally applied to all phytosaurs, as it was the case with poorly-founded previous suggestions of sexual dimorphic features in phytosaurs (Camp Reference Camp1930; Colbert Reference Colbert1947; for a critique see Ballew Reference Ballew, Lucas and Hunt1989), Indeed, Kimmig (Reference Kimmig2009) presented results of a principal component analysis in support of sexual dimorphism for M. buceros and M. pristinus, but also extended that interpretation onto the genera Leptosuchus (including Smilosuchus and Pravusuchus sensu Stocker Reference Stocker2010), Nicrosaurus, and Mystriosuchus. A brief survey indicates that the situation in other phytosaur taxa may be much more complex than it appears to be in Machaeroprosopus.
(1) There are evidently no crested variants known for basal phytosaurs (“Paleorhinus”) and no fully crested morph is known for Angistorhinus (Rutiodon sensu Hungerbühler & Sues Reference Hungerbühler and Sues2001). The only morph within this clade with a short rostral crest is Angistorhinus talainti (Dutuit Reference Dutuit1977). In addition, both the basal phytosaurs and Angistorhinus include specimens of ‘gracile’ and ‘robust’ built, which are traditionally separated at species level (e.g. Paleorhinus bransoni vs. Paleorhinus sawini; Long & Murry Reference Long and Murry1995) or even as distinct genera (Angistorhinus vs. Brachysuchus; Hunt Reference Hunt1994; Leptosuchus vs. Smilosuchus; Pseudopalatus vs. Arribasuchus; Long & Murry Reference Long and Murry1995). A similar distinction is seen in Machaeroprosopus lottorum, with the ‘gracile’ TTU-P10076 with a short rostral crest and the much more ‘robust’ TTU-P10077 with a long rostral crest. Thus, ‘gracile’ and ‘robust’ morphs might be interpreted as sexual variants of one taxon. However, we advise caution, because robustness is difficult to define objectively and, also, and perhaps predominantly, subject to growth variation. Note here that the Canjilon phytosaur assemblage includes both ‘gracile’ smaller specimens (UCMP 34250) and substantially more ‘robust’ larger specimens (UCMP 34256) of the crested morph Machaeroprosopus buceros (Zeigler et al. Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, figs 3D, E, 4D, E).
(2) The genera Leptosuchus and Smilosuchus do not include a clearly uncrested, or dolichorostral morph, with the exception of some smaller and probably juvenile specimens (Camp Reference Camp1930). In those species within both genera which can be reliably differentiated by characters of the temporal region (e.g., by several features of the squamosal and the configuration of the septomaxillae: Stocker Reference Stocker2010; A. Hungerbühler unpublished data), the pattern of crest development is highly variable and does not show a conclusive pattern. In most cases it is hard to see a dimorphic crest development, and we agree with Stocker's (Reference Stocker2010) findings that both the pattern of cresting and the robust or gracile built of skulls are incongruent with the distribution of character states that are currently deemed taxonomically significant within these taxa.
(3) Nicrosaurus and Mystriosuchus both include a crested and partially crested or uncrested (again, broadly, but not absolutely correlated with ‘robust’ and ‘gracile’) morph. First, in contrast to Machaeroprosopus pristinus/buceros and M. lottorum, these morphs differ evidently by more than merely the development of a rostral crest or by crest length. Contra Kimmig (Reference Kimmig2009), we find it difficult to attribute all observed character differences (which also include, for example, the development of the narial openings and the shape of the squamosal; Hungerbühler Reference Hungerbühler1998, Reference Hungerbühler2002) to intraspecific variation within a population. Secondly, there is evidence for the dimorphic development of various cranial characters, both within the hypodigms of the fully crested Nicrosaurus kapffi and in Nicrosaurus meyeri without or with small rostral crests (Hungerbühler Reference Hungerbühler1998). Thirdly, in the case of Mystriosuchus, there is a marked discrepancy in the numerical proportion of the two ‘morphs’: the uncrested, ‘gracile’ Mystriosuchus planirostris is known from at least one dozen skulls, not counting numerous additional cranial fragments, whereas the ‘robust’ Mystriosuchus westphali with a premaxillary crest is represented by the type skull, a partial rostrum, and some jaw fragments only (Hungerbühler Reference Hungerbühler2002).
In conclusion, we strongly urge restraint from rash nomenclatural acts on the basis of our and Zeigler et al.'s (Reference Zeigler, Lucas, Heckert, Heckert and Lucas2002a, Reference Zeigler, Lucas, Heckert, Heckert and Lucasb, Reference Zeigler, Heckert, Lucas, Zeigler, Heckert and Lucas2003a, Reference Zeigler, Lucas and Heckertb) findings without substantial supporting evidence.
6. Systematic conclusions
6.1. Machaeroprosopus lottorum
TTU-P10076 and TTU-P10077 are identified as more closely related to each other than to any other specimen or taxon included in the analysis. This is the strongest phylogenetic signal in the matrix, because the clade was found five out of six times from the initial run of the analysis, and it is supported by the highest bootstrap and decay values in the ingroup (Fig. 25).
6.2. TTU-P10074
Despite the close temporal proximity and the occurrence at the same site, TTU-P10074 is consistently confirmed as not closely related to TTU-P10076 or TTU-P10077, both by parsimony analysis as well as by traditional comparative anatomy (Table 2). Because the range of variation for a given species of Machaeroprosopus is currently unknown, we do not expand the diagnosis of either M. lottorum or M. buceros to incorporate TTU-P10074, and refer to this specimen as Machaeroprosopus sp. This study also demonstrates that pseudopalatine phytosaurs from the same location within a very close temporal range are not necessarily conspecific, but may exhibit a wealth of morphological differences when examined in detail.
Table 2 Assessment of characters of Machaeroprosopus lottorum n. sp. and Machaeroprosopus sp. TTU P-10074 in comparison with the type species of Pseudopalatus and Redondasaurus. Numbers in square brackets refer to characters listed in the Appendix.

6.3. Machaeroprosopus
The diagnosis of Redondasaurus is given as “supratemporal fenestra that are essentially concealed in dorsal view and whose anterior margin only slightly emarginates the skull roof” and “wide squamosal-postorbital bars” (Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993, p. 193). The first diagnostic character is here subdivided in three distinct states (C27[2–4]), which can be unambiguously distinguished in all specimens under study. Merging C27[(2-4] into one character state and re-analysing the data does not change the number and topologies of the MTPs obtained. We quantified the second character to score it objectively. Machaeroprosopus lottorum corresponds to the original diagnosis of Redondasaurus, and could be referred to this genus. However, we propose that Redondasaurus should be regarded as a junior synonym of Machaeroprosopus for the following reasons:
(1) A comparison and assessment of cranial characters in Table 2 shows that Machaeroprosopus lottorum bridges the morphological gap between Redondasaurus (as defined by the type species Redondasaurus gregorii) and Machaeroprosopus (as defined by the type species Machaeroprosopus buceros; the species that were formerly included in the junior synonym Pseudopalatus) with respect to the supratemporal fenestra, the parietal–supraoccipital complex, and some features of the squamosal. In addition, M. lottorum shows a combination of characters that are exclusively expressed either in Machaeroprosopus (=Pseudopalatus) (e.g., C25[2], a laterally expanded terminal knob of the squamosal) or in Redondasaurus (e.g., C18[0], a wider postorbital-squamosal bar). Thus, the existence of a taxon like Machaeroprosopus lottorum makes a distinction between the genera Redondasaurus and Machaeroprosopus (=Pseudopalatus) de facto arbitrary.
(2) Machaeroprosopus (=Pseudopalatus) sensu Ballew (Reference Ballew, Lucas and Hunt1989) constitutes a paraphyletic group in all trees. We explored the possibility of Machaeroprosopus (=Pseudopalatus) being monophyletic by rerunning the analysis under the constraint that only trees were withheld in which the type species Redondasaurus gregorii formed the sister-group to all other taxa. The shortest suboptimal trees that comply with the constraint all show a clade of both species of Redondasaurus as a sister-group of the rest of the ingroup, and are five additional steps (TL 109) or 5% longer than the MPTs; i.e., they contain a considerable amount of additional homoplasies. Therefore, we consider a basal position of Redondasaurus with respect to all other North American pseudopalatine phytosaurs (=Machaeroprosopus) an unlikely option.
(3) Machaeroprosopus gregorii and Machaeroprosopus bermani do not form a monophyletic group in any MPT. Even when the rostral crest features are not considered, it required three additional steps to force M. gregorii with M. bermani into a sister-taxon relationship (=Redondasaurus). This is particularly striking because, according to the original descriptions of Redondasaurus gregorii and Redondasaurus bermani, the presence or absence of a crest is the only distinction between both species, and M. bermani is often regarded as the ‘male’ morph of M. gregorii. This is not reflected in our data, and Machaeroprosopus gregorii is consistently interpreted as more derived than Machaeroprosopus bermani.
In contrast to Long & Murry (Reference Long and Murry1995), we concur with Hunt & Lucas (Reference Hunt, Lucas, Lucas and Morales1993) and Spielmann & Lucas (Reference Spielmann and Lucas2012) that Machaeroprosopus gregorii (Hunt & Lucas Reference Hunt, Lucas, Lucas and Morales1993) represents a valid species. The validity of the taxon is supported by three characters: (1) complete absence of an indentation of the supratemporal fenestra into the skull roof (C27[4], shared with the type of Redondasaurus bermani); (2) dorsoventrally compressed posteriormost portion of the squamosal with lateral rims that strongly converge to each other (C24[0]); and (3) vertical rims of the parietal–squamosal complex surrounding a shallow, vertical supraoccipital shelf (C31[3]). The latter two traits are not present in any other nominal species of the ingroup.
6.4. Arribasuchus
Long & Murry's (Reference Long and Murry1995) subdivision of North American pseudopalatine phytosaurs into a gracile, crest-less Pseudopalatus, and Arribasuchus with massive, crested skulls is unsubstantiated by our data. A minimum of nine additional steps (increase by 7%; TL 136), and even 15 steps when the rostral crest characters are included (increase by 14%; TL 123) is required to force the OTUs into a dichotomy of Pseudopalatus sensu Long & Murry (Reference Long and Murry1995) (Pseudopalatus pristinus, Redondasaurus gregorii, TTU-P10074 and TTU-P10076) on the one hand, and Arribasuchus (Pseudopalatus mccauleyi, Redondasaurus bermani and TTU-P10077) on the other. Hence, we regard Arribasuchus as a junior synonym of Machaeroprosopus.
7. Prospectus
The comparatively poor support for all nodes in the MPTs recovered in the analysis indicates that the data matrix still contains a significant amount of ‘noise’. We suspect that, like those describing the rostral crest morphology, a number of characters do not contain a phylogenetic signal, but represent ontogenetic, sexual or even individual variation. These are difficult to identify at present. An examination of a wider range of specimens and the correlation of character states with those suggested as sexually dimorphic, or with those correlated with a proxy for individual age of the specimens, may help to identify and eliminate phylogenetically uninformative characters. In addition, this study revealed a wealth of cranial characters with potential taxonomic significance, but only few of them could be included in the analysis and evaluated, either because the character states are unknown for the majority of specimens, or because the distribution is uninformative for the small number of OTUs of this analysis.
A factor contributing to the uncertainty is the missing or ambiguous data for several species because of the incompleteness of their type specimens (Redondasaurus gregorii) or poor preservation (Pseudopalatus pristinus, Pseudopalatus buceros). It became evident during the preparation of the data matrix that the majority of the type specimens among Machaeroprosopus are in need of reinvestigation, both to substantiate the current alpha-taxonomy of Machaeroprosopus and to obtain reliable data for a phylogenetic analysis.
Our analysis did not include three additional nominal taxa of North American pseudopalatine phytosaurs. Two of them were not considered because they are generally accepted as junior synonyms of taxa included in the analysis: Machaeroprosopus tenuis Camp, 1930 as Machaeroprosopus pristinus (Ballew Reference Ballew, Lucas and Hunt1989; Long & Murry Reference Long and Murry1995), and Machaeroprosopus andersoni Mehl, 1922 as Machaeroprosopus buceros (Hunt Reference Hunt, Lucas and Morales1993; Long & Murry Reference Long and Murry1995). However, these suggested synonymies were never substantiated by a thorough character analysis.
8. Acknowledgements
First and foremost, we are grateful to John Lott and Patricia Kirkpatrick, for unlimited access to their land, generous financial support and their interest in our work. Without their support and continuing enthusiasm, this project would not have taken place. Jeffrey Martz skillfully executed Figures 3 and 13. AH wishes to thank the staff of the following institutions for help during procuring data used in this study: Mark Norell, Peter Makovicky, and Alejandra Lara (AMNH), David Berman (CM), Spencer Lucas, Andy Heckert, and Alan Lehner and his family (NMNNH), Kevin Padian, Mark Goodwin, and Pat Holroyd (UCMP), Ray Ethington (UMo), and Lyn Murray and Mary Ann Turner (YPM). He also gratefully acknowledges financial support by a HCM fellowship of the European Union (grant no. ERBCHBICT 930521), the Doris and Samuel Welles Fund (Berkeley), and research funds awarded to Hans-Dieter Sues (formerly Royal Ontario Museum, Toronto). We wish to thank Richard Butler and in particular Michelle Stocker for their thorough and constructive reviews.
9. Appendix. List of characters used in the phylogenetic analysis
Character states are polarised with respect to Nicrosaurus kapffi as outgroup. Character states scored for TTU P-10074, TTU P-10076 and TTU P-10077 are described in more detail in the descriptive part.
(1) Shape of anterior half of rostrum: semicircular (0); triangular (1).
(2) Ventral bulge of posteriormost section of premaxillae: absent (0); small bulge to marked dorsally convex expansion (1).
(3) Length of elevated crest: crest absent according to criteria given in text (0); crest over less than half of rostrum (1); crest over at least half of rostrum length (2).
(4) Sagittal profile of prenarial area: steep to moderately steep slope (0); very gentle slope (1); sinuous, i.e., horizontal or convex outline with anterior slope (2).
(5) Individuality of rostral crest: crest absent or crest flanks grade into rostrum (0); crest morphologically set apart from rostrum by a depression, a paramedian groove, or a horizontal expansion of the ventral part of the rostrum (1).
(6) Ventral outline of maxillary rim: straight or concave (0); distinctly convex (1).
(7) Prenarial groove: present (0); absent (1).
(8) Elevation of nares: below or at level with skull roof (0); above level of skull roof (1).
(9) Position of anterior rim of nares relative to anterior rim of antorbital fenestra: anterior (0); at level with (1); posterior (2).
(10) Narial outlets: present (0); absent (1).
(11) Narial wing: present (0); absent, narial opening closed anteriorly (1).
(12) Lateral rim of naris: narrow, rounded (0); flattened, rugose (1).
(13) Length of antorbital fenestra, relative to length of naris: large, >2 (0); moderate to small, <2 (1).
(14) Depression and flange in postorbital bar: absent (0); vertically elongated depression, posterior rim of postorbital bar may form a rounded narrow strut (1); vertically elongated depression, posterior rim of postorbital bar forms a distinct flange (2).
(15) Anterior extent of infratemporal fenestra relative to orbit: anteroventral corner below anterior half of orbit (0); anteroventral corner distinctly in front of anterior rim of orbit (1).
(16) Length of posterior process of squamosal in relation to postorbital length: absent or very small (0); moderate, >4·5 (1); long, <4·5 (2).
(17) Length of free postorbital–squamosal bar: long, >2 (0); moderate, between 1·5 and 2 (1); short, <1·5 (2). Measured as the index of the distance between the level of the edge of the parietal ledge and the tip of the squamosal along the longitudinal axis of the postorbital–squamosal bar (to account for the variable length of the supratemporal fenestra; for the ingroup, the distance is largely identical to squamosal length) to the distance between the parietal ledge to the posteriormost point of the orbit.
(18) Width of squamosal: very broad, <2 (0); broad to narrow, >2 (1). Measured as the index of the distance between the level of the parietal ledge and squamosal tip to the maximum width of squamosal in dorsal view.
(19) Lateral ridge of postorbital–squamosal bar continues as ridge onto posterior process of squamosal: absent (0); present (1). The apomorphic state is a correlated with an angular separation of the dorsal and lateral surfaces of the squamosal on the posterior process.
(20) Ridge on lateral, vertical surface of squamosal: absent (0); present (1).
(21) Sculpture on dorsal surface of squamosal: fully sculptured (0); lanceolate shape, faint sculpture on terminal knob (1); lanceolate shape, terminal knob smooth (2).
(22) Outline of medial rim of squamosal along supratemporal fenestra and posterior process: sinuous (0); angular (1); straight (2).
(23) Face of medial rim of squamosal along supratemporal fenestra and posterior process: entire rim rounded or sharp (0); squared in posterior section (1); entire rim squared (2).
(24) Dorsoventral expansion of posterior process of squamosal: strongly compressed (0); expanded, dorsoventrally high, but at level with surface of postorbital–squamosal bar (1); strongly expanded, distinctly raised over surface of postorbital–squamosal bar (2).
(25) Mediolateral expansion of posterior process of squamosal (terminal knob sensu Ballew, 1989): knob absent, strongly converging medial and lateral rim of squamosal (0); knob present, with parallel or slightly tapering sides (1); knob present, with strong lateral bulge.
(26) Squamosal flange: absent (0); shallow, does not extent along entire length of posterior process of squamosal (1); extensive vertical area, along entire ventral rim of posterior process of squamosal (2).
(27) Supratemporal fenestra on skull roof: fully open, about as broad as long (0); narrow straight or medially curved slit (1); short, semicircular indentation into skull roof, relatively broader than state 1 (2); fully closed, no indentation into skull roof (3).
(28) Rim of supratemporal fenestra on skull roof: rounded (1); bevelled (1).
(29) Outline of top of parietal–supraoccipital complex: angular, ‘inverted V-shaped’ sensu Ballew (1989) (0); rounded, ‘inverted U-shaped’ sensu Ballew (1989) (1); rectangular (2).
(30) Width of parietal ledge: acute (0); horizontal ledge, narrow (1); horizontal ledge, broad (2).
(31) Lateral prongs on parietal ledge: absent (0); present (1).
(32) Slope of lateral walls of parietal–supraoccipital complex: constant moderate (0); constant steep (1); dorsal part vertical (2); entire walls vertical (3).
(33) Shape of supraoccipital shelf: walls grade smoothly into bottom of shelf (0); walls set at marked kink to bottom of shelf (1).
(34) Height of lateral walls of parietal–supraoccipital complex: high (0); low (1).
(35) Thickness of lateral walls of parietal–supraoccipital complex: constantly thin (0); thick base converges rapidly to sharp rim (1).
(36) Posttemporal fenestra, width to height relationship: relatively short and high, 2 to 3 times wider than high (0); wider and lower, about 6 times wider than high (1); very short, slit-like (2).
(37) Postemporal fenestra, ventrolateral border: parietal process of squamosal and paroccipital process merge, paroccipital process forms the entire ventral rim and the medial corner of the fenestra (0); vertical lamina of squamosal below the parietal process forms lateral corner of fenestra (1); lamina of squamosal extends onto paroccipital process, forming ventrolateral border of fenestra (2).
(38) Postemporal fenestra, medial extent: extends medially indenting the triangular extremity of the union of the supraoccipital shelf (supraoccipital) plus the parietal–squamosal bar, and the paroccipital process (exoccipital, opisthotic) (0); fenestra extends less medially, marked groove indents paroccipital process below extremity of supraoccipital shelf (1); parietal–squamosal bar is fused with paroccipital process over a longer distance, fenestra extends only under the flat horizontal part of the parietal–squamosal bar (2).
(39) Oblique furrow crossing alveolar ridge between premaxilla and maxilla absent (0); present, as shallow, but distinct furrow or deep and broad trench (1).
(40) Palatal ridge: low, rounded, longitudinal elevation (0); prominent, sharp, ventrally to ventromedially directed crest (1).
(41) Medial edge of palatine below posterior part of palatal vault: sloping or vertical (0); horizontal flange, may restrict the opening of the palatal vault significantly (1).



























