

Epiphytes grow upon other plants and derive their moisture and nutrients from rain, air, leachates and detritus of the host and other epiphytes, material transported by ants and termites, and excreta of animals, which are effectively distributed by stemflow (Johansson Reference Johansson, Lieth and Werger1989). Epiphytic plants and lichens may comprise a remarkable proportion of the forest flora and biomass, and they are ecologically important in intercepting and retaining atmospheric moisture, providing habitat and a food source for various invertebrates and, in some cases, also contributing fixed nitrogen into the ecosystem (Nash Reference Nash and Nash2008; Hietz Reference Hietz, Bruijnzeel, Scatena and Hamilton2010; Köhler et al. Reference Köhler, Hölscher, Bruijnzeel, Leuschner, Bruijnzeel, Scatena and Hamilton2010; Leuschner Reference Leuschner, Bruijnzeel, Scatena and Hamilton2010). Vascular plants, such as ferns, bromeliads and orchids, form an important proportion of the epiphytic community (Hietz & Hietz-Seifert Reference Hietz and Hietz-Seifert1995; Krömer et al. Reference Krömer, Kessler, Gradstein and Acebey2005; Hietz Reference Hietz, Bruijnzeel, Scatena and Hamilton2010); however, non-vascular cryptogams such as mosses and liverworts can make up more of the photosynthetically active epiphytic biomass in these forests than all the other plant groups put together (Hofstede et al. Reference Hofstede, Wolf and Benzing1993). The overall prominence and species richness of non-vascular epiphytes, including bryophytes and lichens, increase with humidity (Gradstein & Pócs Reference Gradstein, Pócs, Lieth and Werger1989; Hietz Reference Hietz, Bruijnzeel, Scatena and Hamilton2010). Moist tropical forests harbour the most diverse epiphytic communities, although epiphytic bryophytes and lichens occur in almost all biomes, interacting together as a functional epiphytic community (Cornejo & Scheidegger Reference Cornejo and Scheidegger2016).
Dominican amber has preserved a plethora of Miocene species such as arthropods, plants and sometimes also fungi and lichens (reviewed in Penney Reference Penney and Penney2010; Heinrichs et al. Reference Heinrichs, Vitt, Schäfer-Verwimp, Ragazzi, Marzaro, Grimaldi, Nascimbene, Feldberg and Schmidt2013b, Reference Heinrichs, Kettunen, Lee, Marzaro, Pócs, Ragazzi, Renner, Rikkinen, Sass-Gyarmati, Schäfer-Verwimp, Scheben, Solórzano Kraemer, Svojtka and Schmidt2015a; Lóriga et al. Reference Lóriga, Schmidt, Moran, Feldberg, Schneider and Heinrichs2014). The mines in the northern, eastern and western parts of the Dominican Republic yield ambers with an estimated age of 15–20 million years (Iturralde-Vinent & MacPhee Reference Iturralde-Vinent and MacPhee1996; Penney Reference Penney and Penney2010). This fossil resin derives from extinct species of the extant leguminous tree genus Hymenaea Linnaeus, Reference Linnaeus1753 (Langenheim Reference Langenheim1966; Hueber & Langenheim Reference Hueber and Langenheim1986; Poinar Reference Poinar1991; Poinar & Brown Reference Poinar and Brown2002). These trees grew in tropical forests with open areas, and were rich in epiphytes (Grimaldi Reference Grimaldi1996).
Based on its animal and plant inclusions, the Dominican amber forest has been characterised as a moist tropical forest, which was not remarkably different from the modern Neotropical lowland rainforest (Grimaldi Reference Grimaldi1996; Poinar & Poinar Reference Poinar and Poinar1999). Three species of lichens, 22 species of liverworts and 29 species of mosses have so far been described from Dominican amber, of which the liverworts clearly represent extinct taxa (Poinar et al. Reference Poinar, Peterson and Platts2000; Rikkinen & Poinar Reference Rikkinen and Poinar2008; Heinrichs et al. Reference Heinrichs, Kettunen, Lee, Marzaro, Pócs, Ragazzi, Renner, Rikkinen, Sass-Gyarmati, Schäfer-Verwimp, Scheben, Solórzano Kraemer, Svojtka and Schmidt2015a). In addition, the fossil lichens have been described as representatives of extinct species; however, the absence of several important chemical and apothecial characters hinders a reliable differentiation between the fossil and morphologically similar extant species (Poinar et al. Reference Poinar, Peterson and Platts2000; Rikkinen & Poinar Reference Rikkinen and Poinar2008). It has even been suggested that some moss fossils from Dominican amber represent extant species (Frahm & Newton Reference Frahm and Newton2005), but recent analyses strongly suggest that assignments of bryophyte fossils from Cenozoic ambers to extant species should only be conducted if a congruent morphology is present, and if the DNA sequence variation of extant species provides certain likelihood for a species age that is in accordance with the age of the amber (Heinrichs et al. Reference Heinrichs, Vitt, Schäfer-Verwimp, Ragazzi, Marzaro, Grimaldi, Nascimbene, Feldberg and Schmidt2013b, Reference Heinrichs, Scheben, Lee, Váňa, Schäfer-Verwimp, Krings and Schmidt2015b; Villarreal & Renner Reference Villarreal and Renner2014; Váňa et al. Reference Váňa, Schäfer-Verwimp, Bechteler, Schmidt and Heinrichs2015).
Here, we describe a piece of Dominican amber that provides a window on a Miocene Caribbean epiphyte community. The virtually in situ preserved cryptogamic community includes a new species of the extant lichen genus Phyllopsora Müller, Reference Müller1894, a moss that is similar to, but clearly distinguished from, the modern genus Aptychella (Brotherus) Herzog, Reference Herzog1916 and, therefore, assigned to the new fossil genus Aptychellites; as well as three extinct species of the extant leafy liverwort genera Cheilolejeunea (Spruce) Stephani, Reference Stephani1890, Lejeunea Libert, Reference Libert1820 and Radula Dumortier, Reference Dumortier1822.
The members of this community are distinct from extant Caribbean species, further substantiating the notion of a stasis in morphotype diversity, but a certain turnover of species, in the Caribbean since the early Miocene, as recently suggested by Schneider et al. (Reference Schneider, Schmidt, Nascimbene and Heinrichs2015). It is also in line with the hypothesis of Poinar & Poinar (Reference Poinar and Poinar1999) that many of the plants and animals preserved in the Miocene Dominican amber do not have immediate extant descendants.
1. Material and methods
The fossilised epiphytes are preserved in a c.3 cm×2 cm×0.8 cm-sized piece of Dominican amber which is housed in the amber collection of the American Museum of Natural History in New York (AMNH DR-15-3). The specimen originated from the amber mines in the Cordillera of the Dominican Republic. This amber piece contains five cryptogam species: a lichen, a moss and three leafy liverworts (Fig. 1).

Figure 1 Cryptogamic community preserved in Dominican amber specimen AMNH DR-15-3: (A) overview of the inclusions; (B) overview with inclusions indicated: AF = Aptychellites fossilis (holotype circled); CA = Cheilolejeunea antiqua; LM = Lejeunea miocenica (holotype only visible from the opposite side of the amber piece); PM = Phyllopsora magna (holotype circled); RI = Radula intecta (holotype circled); (C) Cheilolejeunea antiqua. Scale bars = 1 mm (A, B); 100 µm (C).
The amber surface was ground and polished manually with a series of wet silicon carbide abrasive papers (grit from FEPA P 600–4000; i.e., 25.8 μm to 5 μm particle size, Struers) to minimise light scattering during the investigation. Portions of the lichen thallus reaching the amber surface were further polished to visualise the thallus section (Fig. 2D). The prepared specimen was placed on a glass microscope slide with a drop of water applied to the upper surface of the amber, and covered with a glass coverslip. The inclusions were examined under a Carl Zeiss Stemi 2000 dissection microscope and a Carl Zeiss Axio Scope A1 compound microscope equipped with Canon 5D digital cameras. In most instances, incident and transmitted light were used simultaneously. Oblique incident light was obtained using a goose-neck light guide of a Carl Zeiss CL 1500 Eco cold light source. All presented illustrations are digitally stacked photomicrographic composites of up to 110 individual focal planes, obtained by using the software package HeliconFocus 5.0.

Figure 2 Phyllopsora magna sp. nov. in Dominican amber (AMNH DR-15-3): (A) portion of thallus with fabellate to elongate ascending squamules. The large thallus portion represents the holotype (upper half of image shows shoot systems of Radula intecta); (B) ascending squamule tip revealing the pseudocortex on the lower side; (C) prominent isidia developing from the tips of the squamules; (D) cross-sections of the thallus showing the thin cortex, medulla and loose pseudocortical hyphae on the lower side. Scale bars = 1 mm (A); 100 µm (B–D).
2. Systematic palaeontology
2.1. Lichens
Phylum Ascomycota Cavalier-Smith, Reference Cavalier-Smith1998
Class Lecanoromycetes Eriksson & Winka, Reference Eriksson and Winka1997
Order Lecanorales Nannfeldt, Reference Nannfeldt1932
Family Ramalinaceae Agardh, Reference Agardh1821
Genus Phyllopsora Müller, Reference Müller1894
Phyllopsora magna Kaasalainen, Rikkinen &
A. R. Schmidt, sp. nov.
(Fig. 2)
Holotype. American Museum of Natural History, New York, USA, AMNH DR-15-3; MycoBank number MB 818898. The large thallus portion shown in Fig. 2A (middle) represents the holotype.
Type locality. Dominican Republic, Santiago area.
Age and stratigraphic position. Early Miocene, about 15 to 20 million years old.
Etymology. Latin for large. The species epithet refers to the exceptionally large size of the thallus lobes of this Phyllopsora species.
Diagnosis. Lobes subfoliose, large, up to 3.1 mm wide; prothallus imperceptible or lacking; upper cortex poorly developed, <5 µm thick; lower side possessing a pseudocortex; isidia marginal, cylindrical to clavate.
Description. Prothallus not visible. Thallus squamulose; with foliose-like lobes but without the lower cortex (Figs 1, 2A, B). Squamules ascending, large, up to 3.1 mm wide and 3.8 mm long, and approximately 80 µm thick (Fig. 2A, B, D). Shape of the squamules varying from wide flabellate to elongate, with undulating to slightly lobate margins (Fig. 2A–C). Upper surface glabrous and brown, paling towards the margins (Fig. 2A). Cortex very thin (2–4 µm; Fig. 2D). Medulla composed of pale hyphae (Fig. 2D). Lower surface with a pseudocortex, formed by white (occasionally with a hue of rusty-brown), loosely and mainly longitudinally organised hyphae (Fig. 2B, D). Isidia elongate, cylindrical to clavate, developing horizontally from the margins of the squamules (Fig. 2C). Ascomata or pycnidia not detected.
2.2. Liverworts
Phylum Marchantiophyta Stotler & Crandall-Stotler, Reference Stotler, Crandall-Stotler, Shaw and Goffinet2000
Class Jungermanniopsida Stotler & Crandall-Stotler, Reference Stotler and Crandall-Stotler1977
Order Porellales Schljakov, Reference Schljakov1972
Family Lejeuneaceae Cavers, Reference Cavers1910
Genus Cheilolejeunea (Spruce) Stephani, Reference Stephani1890
Cheilolejeunea antiqua (Grolle) Ye & Zhu, Reference Ye and Zhu2010
(Fig. 1C)
Description. Sterile, unbranched shoot, c.1.6 mm long, 650–700 µm wide. Stem straight, c.30–50 µm in diameter, with a c.3 cells-wide ventral merophyte; cortical cells rectangular, walls thin- to slightly thick-walled. Rhizoids in bundles at base of underleaves. Leaves remote to weakly imbricate, insertion J-shaped; lobes longer than wide, slightly falcate, orbicular to oblong-ovate, apex broadly rounded, margins entire. Median leaf cells +/− regular, 15–20 µm×18–27 µm, thin-walled to slightly thick-walled, with weak trigones, occasionally weak intermediate thickenings present; cells towards leaf margins smaller. Cuticle smooth. Lobules well-developed, inflated, ovoid to ovoid-triangular, c.2/5–1/3 of leaf length, free margin often +/− straight, with a short apical tooth. Underleaves [poorly preserved and often somewhat damaged] remote, entire (unlobed), broadly orbicular to orbicular reniform, c.150–200 µm wide, c.3.5–4.5 times as wide as shoot.
Genus Lejeunea Libert, Reference Libert1820
Lejeunea miocenica Heinrichs, Schäf.-Verw.,
M. A. M. Renner & G. E. Lee sp. nov.
(Fig. 3)
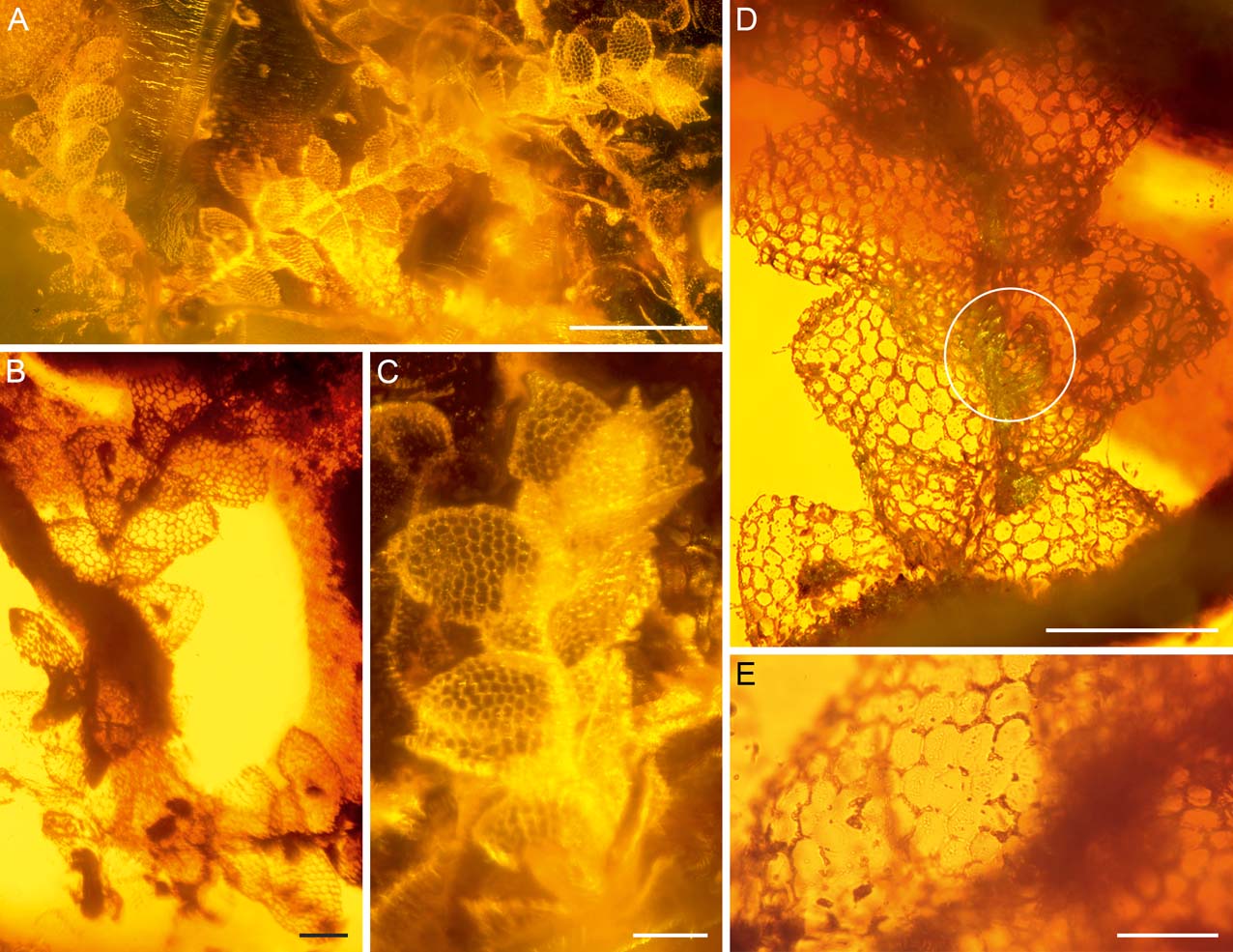
Figure 3 Lejeunea miocenica sp. nov. in Dominican amber (AMNH DR-15-3): (A–C) gametophytes in dorsal view. The gametophyte fragment shown in (A) (left), (B) and (D) represents the holotype; (D) holotype in ventral view, one underleaf circled; (E) close-up of the ventral leaf surface of the holotype, showing asperulate cuticle. Scale bars=500 µm (A); 100 µm (B–D); 50 µm (E).
Holotype. American Museum of Natural History, New York, USA, AMNH DR-15-3. The gametophyte fragment shown in Figure 3A (left), B and D represents the holotype.
Type locality. Dominican Republic, Santiago area.
Age and stratigraphic position. Early Miocene, about 15 to 20 million years old.
Etymology. The species epithet refers to the Miocene age of the fossil.
Diagnosis. Gametophytes with two cells-wide ventral merophytes, complicate bilobed, incubously inserted leaves with (triangularly) ovate dorsal lobes with obtuse to apiculate apices, a papillose cuticle and well developed ventral lobules, as well as bifid underleaves with often blunt lobes and without lateral appendages.
Description. Sterile shoots up to c.2.5 mm long, 340–490 µm wide. Stems straight, ±25–40 µm wide, with a two cells-wide ventral merophyte; cortical cells rectangular. Leaves distant to contiguous, widely to obliquely spreading. Lobes variable in shape, ovate to triangularly ovate, apex obtuse to subacute to apiculate; margin entire to somewhat crenate. Median leaf cells regular, sometimes +/− irregular, c.15–25 µm×20–25 µm, thin-walled, with weak trigones and intermediate thickenings, marginal cells quadrate to rectangular, 10–15×10–27 µm. Cuticle finely papillose. Lobules well-developed, c.2/5 (–1/2) of leaf length, rounded-rectangular to asymmetrically triangular, somewhat inflated, free margin often +/− straight, involute, apical tooth hidden or sometimes seen in situ. Underleaves distant, appressed to stem, c.60–100 µm long, 60–95 µm wide, rounded, c.1.7–2.5×the stem width, margins entire, 30–50 % bifid, sinus V-shaped, lobes erect, narrowly obtuse to (sub-)acute, (3–)4–5 cells wide at the base and 1–2 cells wide at the apex, no rhizoids seen. Vegetative distribution possibly by caducous branches. Androecia and gynoecia unknown.
Remarks. Several isolated fragments which are considerably smaller than the main shoots may represent detached branches. These fragments have more remotely inserted leaves, reduced lobules and underleaf lobes that are only 1–2 cells wide at the base.
Family Radulaceae Müller, Reference Müller1909
Genus Radula Dumortier, Reference Dumortier1822
Radula intecta M. A. M. Renner, Schäf.-Verw. &
Heinrichs, sp. nov.
(Fig. 4)
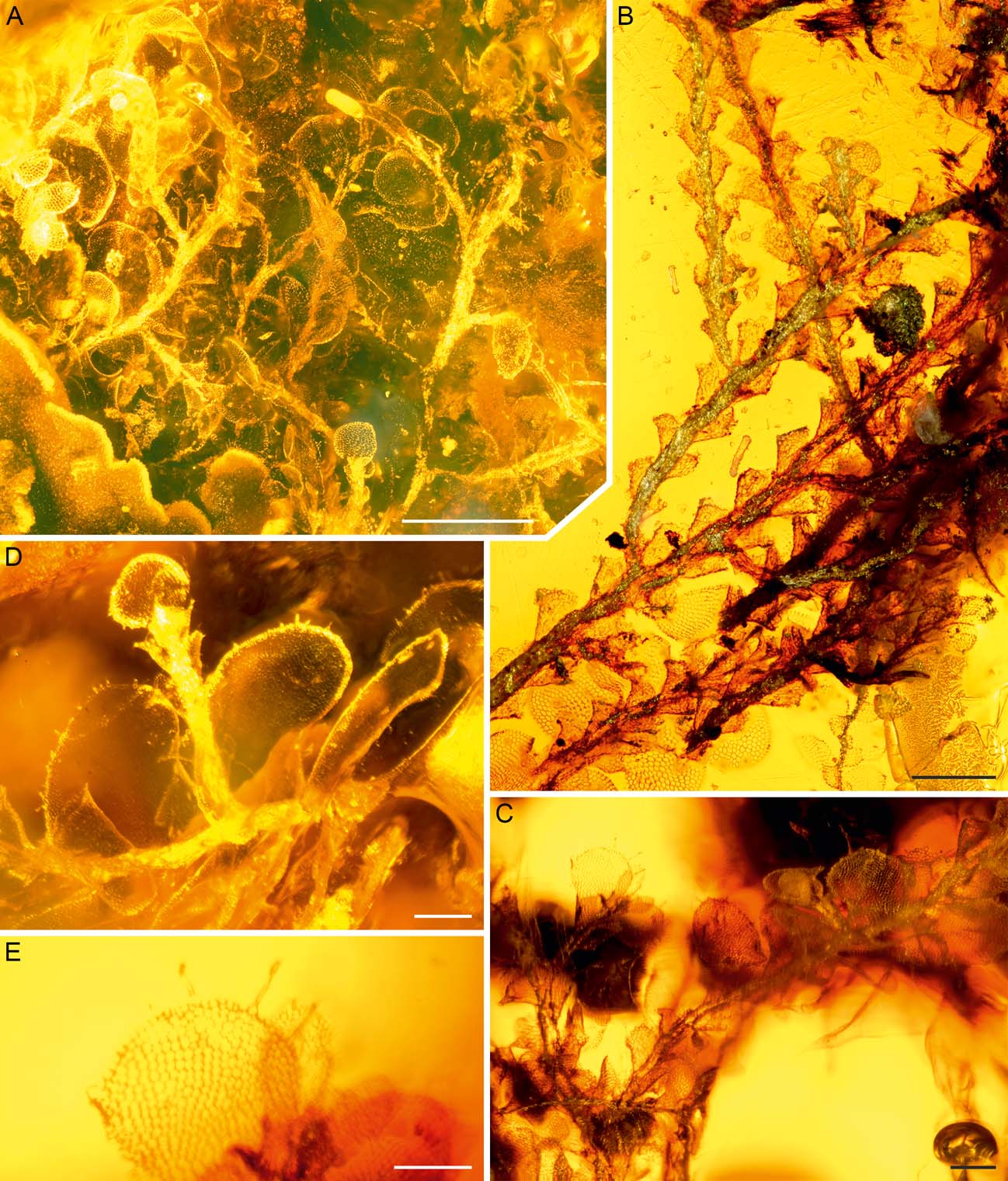
Figure 4 Radula intecta sp. nov. in Dominican amber (AMNH DR-15-3): (A) shoot system in ventral view. The bright shoot fragment at the left margin of the image belongs to Lejeunea miocenica; (B) shoot system with detached lobes. The plant fragments with complete leaves in the lowermost portion of the image belong to Cheilolejeunea antiqua and Lejeunea miocenica; (C–E) portions of shoots with partly detached leaf lobes. Some lobe margins with ciliate outgrowths. The gametophyte fragment shown in (D) represents the holotype. Scale bars = 500 µm (A); 200 µm (B–D); 100 µm (E).
Holotype. American Museum of Natural History, New York, USA, AMNH DR-15-3. The gametophyte fragment shown in Figure 4D represents the holotype.
Type locality. Dominican Republic, Santiago area.
Age and stratigraphic position. Early Miocene, about 15 to 20 million years old.
Etymology. Latin for uncovered, exposed, naked. The species epithet refers to the frequently caducous leaf lobes which are often detached in the fossil.
Diagnosis. Gametophytes with incubously inserted, complicate-bilobed, smooth, broadly elliptic leaves with frequently caducous dorsal lobes having ciliate marginal outgrowths, and subquadrate, small ventral lobules; underleaves completely lacking.
Description. Plants sterile, stems 80–120 µm in diameter, indistinctly pinnately branched, branching exclusively of Radula-type, secondary shoots of smaller stature than primary shoots. Leaves imbricate, slightly obliquely patent, broadly elliptic with rounded apex, those of primary shoots c.650–900 µm long and 340–580 µm wide, margins entire, interior of dorsal margin ampliate but not auriculate, extending over the stem surface in dorsal view and completely obscuring the stem, frequently caducous and older stem sectors frequently denuded of leaf lobes; those that remain attached sometimes producing rhizoids from leaf lobe marginal cells. Keel straight to weakly convex, at an angle of 40–60° to the stem, junction between keel and leaf lobe flush, at that point keel turns to 45° at the keel-lobe-junction. Postical lobe margin straight, otherwise continuously curved. Leaves of secondary shoots possibly rotund, completely caducous, dislocating as a whole along a curved to L-shaped line from top of stem insertion to lobe-lobule junction (two putatively mature lobes from secondary shoots seen). Lobule width 120–160 µm, length 160–180 µm, subquadrate, exterior margin inclined towards stem straight, apex obtuse, antical margin straight to slightly concave, carinal region indistinctly inflated along keel, insertion longitudinal. Leaf surface smooth, cells isodiametric to slightly elongate, not particularly regular in shape, 15–25 µm in diameter, towards margin quadrate to rectangular, 7–10 µm×10–30 µm, thin-walled, toward leaf margin continuously thickened except some exterior walls, trigones small, triangular.
2.3. Mosses
Phylum Bryophyta Schimper, Reference Schimper and von Zittel1879
Class Bryopsida Pax, Reference Pax1900
Order Hypnales W. R. Buck & Vitt, Reference Buck and Vitt1986
Family Pylaisiadelphaceae Goffinet & Buck, Reference Goffinet and Buck2004
Aptychellites fossilis Schäf.-Verw., Hedenäs, Ignatov & Heinrichs gen. & sp. nov.
(Fig. 5)

Figure 5 Aptychellites fossilis gen. et sp. nov. in Dominican amber (AMNH DR-15-3): (A) overview; (B) portion of a shoot system showing apical gemmae clusters (arrows); (C) holotype with apical gemmae cluster. The inclusions above the gemmae clusters are detritus and do not belong to the moss; (D, E) portions of leafy shoots; (F) apical gemmae cluster; (G) leaf in lateral view with group of alar cells. Scale bars = 1 mm (A); 100 µm (B–F); 50 µm (G).
Holotype. American Museum of Natural History, New York, USA, AMNH DR-15-3. The plant shown in Figure 5C represents the holotype.
Type locality. Dominican Republic, Santiago area.
Age and stratigraphic position. Early Miocene, about 15 to 20 million years old.
Etymology. The genus name points to the similarities of the fossil to the extant genus Aptychella. The species epithet emphasises the presence of an extinct taxon.
Diagnosis. Gametophytes with denticulate, spirally inserted, acute, concave leaves without or with a very short costa, prosenchymatous leaf cells and groups of alar cells, and apiculate clusters of filamentous, uniseriate gemmae.
Description. At least eight branches or secondary stems with branches present. Stems ca 50–80 µm in diameter, contrastingly darker than leaves, glossy, well visible between leaves, robust, without hyalodermis, fasciculately branched and without pinnate branching in the subapical 5 mm parts of stems. Leaves spirally arranged, c.20 per 1 mm of stem, erect to spreading on the larger gametophyte portions, patent on the weaker branch portions and towards the attenuate branch ends. Leaves mostly ovate, sometimes obovate-oblong and widest shortly below the constriction to acumen, gradually tapered to the base, above suddenly narrowed to distinct set-off acumen, strongly concave. Largest leaves c.1 mm long (acumen 0.2 mm) and 0.4 mm wide. Alar region not auriculate. At least branch leaves not decurrent. Costa not visible; i.e., very short or absent. Leaf margin serrate in acumen and above the widest level of the leaf, serrulate to subentire below, plane. Median leaf lamina cells narrowly linear, incrassate or at least slightly so and smooth, cells in acumen shorter and wider, alar cells (few seen in one leaf) rectangular or shortly rectangular, inflated and incrassate. Leaf lamina cells in central portions of largest leaves c.38–54 µm x 4–5 µm or longer, towards leaf apex becoming shorter and often only 15–25 µm long. Apical clusters of filamentous, up to at least 16 cells-long uniseriate gemmae frequently present. Gemmae originating from a ‘basal’ cell that is c. twice as long as the gemmae cells and narrows towards its insertion on the stem. Cells of gemmae rectangular, shortly rectangular or (rarely) quadrate, c.20–25 µm in diameter, towards apical region sometimes somewhat swollen, then with a diameter of up to c.40 µm. Leaves becoming smaller towards gemmae clusters.
3. Discussion
The five epiphyte species preserved in specimen AMNH-DR-15-3 were part of a corticolous community of lichens, mosses and liverworts in a Miocene Caribbean forest ecosystem. Previous records of cryptogams from Dominican amber included an exceptional finding of seven species of bryophytes in a single piece of amber (Frahm & Newton Reference Frahm and Newton2005), providing evidence that the bark of the resinous Hymenaea trees was covered by diverse cryptogamic communities. However, it is still unclear if the Hymenaea bark was generally covered by cryptogamic communities or if only some parts of the tree, or single individuals, hosted cryptogams.
3.1. Systematic assignment of the cryptogamic inclusions
3.1.1. Lichens
The fossil lichen has a squamulose thallus consisting of relatively wide lobes without a lower cortex. Similar structures are produced by many lichen genera, including Phyllopsora and Cladonia Browne, Reference Browne1756, which are both likely candidates for preservation in amber. The fragile habit and very thin upper cortex of the thallus lobes more resemble those in some extant Phyllopsora species (e.g., Timdal Reference Timdal2008, Reference Timdal2011) than in the primary squamules of Cladonia species, which tend to be more robust and have a thick upper cortex (Osyczka & Rola Reference Osyczka and Rola2013). Additionally, the ventral pseudocortex formed by longitudinally organised hyphae, together with clearly corticated, clavate marginal isidia, indicate that the fossil lichen belongs to Phyllopsora. The genus includes more than 100 extant, crustose to squamulose species, of which 34 are present in the West Indies (Timdal Reference Timdal2011). In addition, one fossil species, Phyllopsora dominicanus Rikkinen (Rikkinen & Poinar Reference Rikkinen and Poinar2008), has been previously described from Dominican amber (Rikkinen & Poinar Reference Rikkinen and Poinar2008).
The most distinctive feature of Phyllopsora magna are the large squamules, which give it an almost foliose appearance. Most extant Phyllopsora species and the fossil P. dominicanus, have squamules less than one millimetre wide (Swinscow & Krog Reference Swinscow and Krog1981; Brako Reference Brako1991; Timdal & Krog Reference Timdal and Krog2001; Upreti et al. Reference Upreti, Divakar and Nayaka2003; Elix Reference Elix2006; Rikkinen & Poinar Reference Rikkinen and Poinar2008; Mishra et al. Reference Mishra, Upreti, Nayaka and Haridas2011; Timdal Reference Timdal2008, Reference Timdal2011; McCarthy Reference McCarthy2016). Some of the larger species may have lobes up to two millimetres wide (Brako Reference Brako1991; Timdal Reference Timdal2008, Reference Timdal2011), which are still notably smaller than the over 3 mm-wide lobes observed in Phyllopsora magna.
The fossil lichen does not seem to have had a prothallus, a characteristic structure of many extant Phyllopsora species. However, the base of the thallus is covered by bryophytes and thus not clearly visible. Some rusty hued hyphae occur among the otherwise white pseudocortex hyphae, possibly indicating the presence of a poorly developed prothallus, which is reddish brown in the extant taxa. However, several extant, especially Neotropical, Phyllopsora species also do not seem to produce prothalli (Timdal Reference Timdal2008, Reference Timdal2011). The structure of the upper cortex is an important feature used to discriminate between extant Phyllopsora species (Swinscow & Krog Reference Swinscow and Krog1981). The cortex structures of most taxa can be classified into one of two main types, but intermediate forms are also common. In one type, the cortex hyphae are highly gelatinised and their lumina are reduced to narrow thread-like channels resembling a network; whilst in the other type the hyphae are much less gelatinised and the cortex is formed by isodiametric cells (Swinscow & Krog Reference Swinscow and Krog1981). The thickness of the cortex may vary from five to 60 micrometres and seems to be related to the size of the squamules (Swinscow & Krog Reference Swinscow and Krog1981; Brako Reference Brako1991). The cortex of Phyllopsora magna is very thin and not assignable to the main structure types. Similar, poorly-developed upper cortices are known from some extant Neotropical Phyllopsora species (Timdal Reference Timdal2008, Reference Timdal2011).
The combination of characters observed in Phyllopsora magna is unique and the species thus seems to represent an extinct, fossil taxon. The previously known Phyllopsora dominicanus better resembles extant species and cannot be confidently separated from all of them on morphological grounds (Rikkinen & Poinar Reference Rikkinen and Poinar2008). Two fossil species of Parmelia Acharius, Reference Acharius1803, previously described from Dominican amber, were only tentatively placed in that genus, and may well represent other parmelioid genera (Poinar et al. Reference Poinar, Peterson and Platts2000).
3.1.2. Liverworts
All observed liverworts belong to Porellales, a largely epiphytic lineage of leafy liverworts characterised by exclusively lateral branching, incubous foliation, rhizoids in bundles, frequent occurrence of complicate bi- or trilobed leaves, endosporous protonemata and a complete lack of mycorrhiza-like mutualisms (Heinrichs et al. Reference Heinrichs, Gradstein, Wilson and Schneider2005; Feldberg et al. Reference Feldberg, Schneider, Stadler, Schäfer-Verwimp, Schmidt and Heinrichs2014). With more than 1000 species, Lejeuneaceae is the most speciose family of the Porellales (He & Zhu Reference He and Zhu2011). Their centres of diversity are the humid tropics, and especially tropical lowland forests, where Lejeuneaceae can account for 70 % of the species diversity of liverworts (Cornelissen & Ter Steege Reference Cornelissen and Ter Steege1989; Gradstein Reference Gradstein2006). Some 70 genera of Lejeuneaceae are currently accepted (Bechteler et al. Reference Bechteler, Lee, Schäfer-Verwimp, Pócs, Peralta, Renner, Schneider and Heinrichs2016; Wang et al. Reference Wang, Zhu and Gradstein2016) of which Lejeunea is one of the largest, including some 300 extant species (Lee Reference Lee2013). Lejeunea is a hyper-diverse genus which differs from the closely related genera Microlejeunea (Spruce) Stephani, Reference Stephani1898 and Harpalejeunea (Spruce) Schiffner, Reference Schiffner, Engler and Prantl1893 by a lack of ocelli (Dong et al. Reference Dong, Schäfer-Verwimp, Pócs, Feldberg, Czumaj, Schmidt, Schneider and Heinrichs2013); its morphology-based supra-specific classification is not supported by molecular data (Heinrichs et al. Reference Heinrichs, Dong, Schäfer-Verwimp, Pócs, Feldberg, Czumaj, Schmidt, Reitner, Renner, Hentschel, Stech and Schneider2013a). The fossil, however, resembles members of the polyphyletic Lejeunea subgenus Crossotolejeunea Spruce, 1884 in the circumscription of Reiner-Drehwald & Goda (Reference Reiner-Drehwald and Goda2000), in having bifid underleaves with blunt lobes and variably shaped leaf lobes with obtuse to acute apices, an asperulate cuticle and sometimes crenate margins. Lejeunea is a Cenozoic genus with several species lineages dating back to the early Miocene (Heinrichs et al. Reference Heinrichs, Scheben, Bechteler, Lee, Schäfer-Verwimp, Hedenäs, Singh, Pócs, Nascimbene, Peralta, Renner and Schmidt2016). Several Crossotolejeunea species occur in modern Caribbean rainforests; however, taxonomically relevant characters of extant species, such as perianth shape, oil body type and branching pattern, are not preserved in the fossil. We therefore abstain from assigning the fossil to an extant species and describe it as a fossil taxon.
Another sterile Lejeunea fossil from Dominican amber was identified to genus level only (Reiner-Drehwald et al. Reference Reiner-Drehwald, Schmidt and Heinrichs2012). It differs from Lejeunea miocenica in having entire lobe margins with rounded apices. Since the Mexican amber fossil Lejeunea palaeomexicana Grolle, Reference Grolle1984 has recently been transferred to Ceratolejeunea (Spruce) Jack & Stephani, Reference Jack and Stephani1892 (Lee et al. Reference Lee, Schäfer-Verwimp, Schmidt and Heinrichs2015), only two fossils of Lejeunea are currently known.
Another sterile inclusion inside specimen AMNH DR-15-3 matches the morphology of the Dominican amber species Cheilolejeunea antiqua, although the leaf lobules are somewhat smaller than in the original description (Grolle Reference Grolle1983). This species has already been observed several times and seems to be a rather common element of the liverwort flora of the Dominican amber forest.
Lejeuneaceae are the most diverse family in Dominican amber; however, other Porellales families have also been reported, namely representatives of the monogeneric families Frullaniaceae and Radulaceae (Heinrichs et al. Reference Heinrichs, Kettunen, Lee, Marzaro, Pócs, Ragazzi, Renner, Rikkinen, Sass-Gyarmati, Schäfer-Verwimp, Scheben, Solórzano Kraemer, Svojtka and Schmidt2015a). The genus Radula includes at least 200 extant species (Yamada Reference Yamada, Gradstein and Costa2003) and has an almost world-wide distribution, with the majority of species occurring in humid, tropical or warm-temperate regions (Devos et al. Reference Devos, Renner, Gradstein, Shaw, Laenen and Vanderpoorten2011a). Most species are obligate or facultative epiphytes on bark or leaves, and they exist in a variety of habitats varying from rainforests to alpine tundra (Devos et al. Reference Devos, Renner, Gradstein, Shaw, Laenen and Vanderpoorten2011a). The genus can easily be recognised by the total lack of underleaves and the presence of rhizoid bundles on the lobule surface rather than on the stem. The caducous leaves which produce marginal rhizoids, the distinct trigones of the leaf cell walls and the pinnately branched dimorphic shoot systems of Radula intecta are consistent with the extant Radula subgenus Radula (Devos et al. Reference Devos, Renner, Gradstein, Shaw and Vanderpoorten2011b), but not necessarily diagnostic of that lineage. In particular, the apparent absence of ornamentation from the leaf-cell surfaces suggests that R. intecta may not belong to the subgenus Radula, whose crown-group species mostly have ornamented surfaces (Renner Reference Renner2014). Species with dimorphic, pinnately branched shoot systems, caducous leaves and smooth cell surfaces also occur in the subgenera Odontoradula Yamada, Reference Yamada1979 and Volutoradula Devos et al., Reference Devos, Renner, Gradstein, Shaw and Vanderpoorten2011b; thus it is not possible to assign the fossil to a subgenus with confidence.
Three fossil Radula species are currently accepted, the Paleogene Baltic and Bitterfeld amber taxa Radula sphaerocarpoides Grolle, Reference Grolle1980 and Radula oblongifolia Caspary, Reference Caspary1887 (Grolle & Meister Reference Grolle and Meister2004), and the Miocene Dominican amber species Radula steerei Grolle (Grolle Reference Grolle1987). Radula intecta differs from these species in the frequent presence of caducous leaf lobes.
Among extant Radula species, Radula intecta resembles the Neotropical Radula cubensis Yamada (Yamada Reference Yamada1983) and Radula schaefer-verwimpii Yamada (Yamada Reference Yamada1990) in having caducous leaves; however, the leaf margins of both extant species lack the ciliate outgrowths of R. intecta. Such cilia are present in some forms of the polyphyletic Radula javanica Gottsche in Gottsche et al., Reference Gottsche, Lindenberg and Nees1845 (Renner Reference Renner2014); however, this extant species has a puncticulate leaf lobe cuticle and is more robust than R. intecta. The combination of subquadrate lobules whose lamina is plane (not reflexed) and its exterior margin straight and inclined toward the stem, the dislocation of nearly all leaves on both primary and secondary shoots, and the production of rhizoids from leaf lobe marginal cells, is distinctive for the new fossil.
3.1.3. Mosses
Several pleurocarpous families of mosses include species with filamentous, uniseriate gemmae. These gemmae usually cluster in leaf axils and/or on specialised brood branches; e.g., in Crossomitrium Müller, Reference Müller1874 (Pilotrichaceae); Jaegerina Müller, Reference Müller1876 (Pterobryaceae); Clastobryopsis Fleischer, Reference Fleischer1923 (Pylaisiadelphaceae); Pylaisiadelpha Cardot, Reference Cardot1912 (Pylaisiadelphaceae) (Buck Reference Buck1998; Goffinet & Buck Reference Goffinet and Buck2004; Tixier Reference Tixier1977); Leptotheca Schwaegrichen, Reference Schwägrichen1823–1827 (Rhizogoniaceae) (Churchill & Buck Reference Churchill and Buck1982); and Gammiella Brotherus, Reference Brotherus, Engler and Prantl1908 (Pylaisiadelphaceae) (Tan & Jia Reference Tan and Jia1999).
Amongst pleurocarpous mosses, terminal clusters of filamentous gemmae are typical for the Central American species Brymela crosbyi (Allen) Allen, Reference Allen2010 and the Pylaisiadelphaceae genus Aptychella. Brymela crosbyi, however, has lanceolate to elongate-triangular, long acuminate, strongly plicate leaves and faint costae that extend more or less two-thirds the leaf length (Allen Reference Allen2010) and is thus not closely related to the moss fossil. Aptychella occurs with some nine extant species in Asia and includes a single extant Neotropical species, Aptychella proligera (Brotherus) Herzog, Reference Herzog1916. The latter species may have Asian ancestors and has possibly colonised the Neotropics in rather recent times (Akiyama et al. Reference Akiyama, Schäfer-Verwimp, Printarakul, Suleiman, Tan, Goffinet, Yong and Müller2015). It is a montane rather than a lowland species and occurs from Mexico southwards to Bolivia, in Brazil and in the Caribbean (Buck Reference Buck1998). Although the numerous gemmae clusters of the Dominican amber fossil suggest a relationship with Aptychella, the strongly concave and rather densely denticulate, spreading leaves are untypical for this genus (Tixier Reference Tixier1969). The concave, denticulate leaves are more indicative of Wijkia Crum, Reference Crum1971 (Pylaisiadelphaceae); however, this genus has gemmae on flagellate branches. A single extant species (Wijkia flagellifera (Brotherus) Crum, Reference Crum1971) occurs in montane forests of the Dominican Republic.
As a consequence of the somewhat deviant morphology of the fossil and extant taxa, we prefer to place the fossil in a new genus, Aptychellites, and name it A. fossilis. This taxon may have also been observed by Frahm (Reference Frahm1993) in the Dominican amber collection of the Naturkundemuseum Stuttgart; however, Frahm identified it as “Clastobryum spec.” Beside the terminal clusters of gemmae, we observed a single gemma in the axil of a leaf; however, we felt unable to decide if this gemma originated at the leaf base or, in fact, represents a detached gemma from a terminal cluster. We were also not able to decide on the number of individuals preserved in the piece of amber, since it is difficult to see if some of those that appear connected are, in fact, separate pieces.
3.2. Palaeoecology
The taxonomic diversity of organisms and the habit of the bryophytes preserved in specimen AMNH DR-15-3 closely resemble extant epiphytic communities living on trunks and branches in lowland to submontane and relatively humid Caribbean tropical forests. Extant Phyllopsora species grow primarily on bark in humid forests of the tropics and subtropics, but sometimes also on bryophytes (Swinscow & Krog Reference Swinscow and Krog1981; Brako Reference Brako1991; Timdal & Krog Reference Timdal and Krog2001; Mishra et al. Reference Mishra, Upreti, Nayaka and Haridas2011; Timdal Reference Timdal2011). The poorly developed upper cortex and the relatively large, ascending lobes without proper lower cortex enable efficient water evaporation and suggest a moist and shaded environmental preference. Coexistence with the moisture-binding bryophytes might have facilitated the maintenance of the favourable microclimatic conditions. Pleurocarpous mosses and Porellalean liverworts are known to include numerous epiphytes and are common elements of the Dominican amber forest (Frahm & Newton Reference Frahm and Newton2005; Heinrichs et al. Reference Heinrichs, Vitt, Schäfer-Verwimp, Ragazzi, Marzaro, Grimaldi, Nascimbene, Feldberg and Schmidt2013b, Reference Heinrichs, Kettunen, Lee, Marzaro, Pócs, Ragazzi, Renner, Rikkinen, Sass-Gyarmati, Schäfer-Verwimp, Scheben, Solórzano Kraemer, Svojtka and Schmidt2015a). Many of these species are exposed to direct sunlight and thus need to cope with periodical dehydration. The water lobules and sacs in Frullaniaceae, Lejeuneaceae and Radulaceae have been interpreted as adaptations to a drier environment (Cornelissen & Ter Steege Reference Cornelissen and Ter Steege1989; Kürschner et al. Reference Kürschner, Frey and Parolly1999; Acebey et al. Reference Acebey, Gradstein and Krömer2003) and may assist the plants in surviving daily periods of desiccation. It is considered that the preserved cryptogam community once grew on the bark of a Hymenaea tree in a rather open environment. However, we cannot exclude an occurrence in a dense forest; though in this case the cryptogamic community was likely situated in the light-permeated forest canopy.
Acknowledgements
We would like to thank David A. Grimaldi and Paul C. Nascimbene (New York) for providing the amber specimen for study. We thank Bruce Allen (St. Louis, USA) for comments on the moss inclusion. This study was supported by the Alexander von Humboldt Foundation (grants to UK and GEL).





