

With regard to the fauna of the British coal measures, Ward (Reference Ward1875, Reference Ward1890, Reference Ward1905) presented a description, illustrations and the distribution of many of the genera from the North Staffordshire coalfield, which he believed would also be found in other coalfields throughout the British Isles. In fact, most of the genera described from North Staffordshire by Ward are also to be found in the Lower Coal Measures of the Midland Valley of Scotland. There is, however, a discrepancy of complete and articulated specimens of Coal Measure actinopterygians from the Pennsylvanian in Scottish collections, as compared with English collections from the same period. This discrepancy is probably due to the mode of fossil preservation in Scottish areas, and also the result of material from the coal and ironstone seams of the Scottish coalfields having been given less attention and not collected to the same degree as they were in the larger English coalfields. Moreover, no extensive work on the Scottish coal measure fauna was undertaken during the period when the coal industry was at its height, whereas many scientific works were available regarding the English and Welsh coalfields. Binney (Reference Binney1841) described many fossil fishes, including Elonichthys egertoni Agassiz, from the Pendleton Coal Field; Hancock & Atthey (Reference Hancock and Atthey1868) described the teeth of Elonichthys egertoni from the coal measures of Newsham, and in a later paper (Hancock & Atthey Reference Hancock and Atthey1872) they described a further three new species of Actinopterygii, also from Newsham; Davis (Reference Davis1878) gave an account of the fish remains from the Yorkshire Coal Measures; Wellburn (Reference Wellburn1899, Reference Wellburn1900) described and illustrated material from Lower Coal Measures of the Halifax and Littleborough Districts, and included a description of the remains of Elonichthys binneyi Traquair and Rhadinichthys monensis Traquair; Bolton (Reference Bolton1904) gave an account of the palaeontology of the Lancashire Coal Measures; and Wood (Reference Wood1936) discussed the remains and distribution of a number of fossil genera, including actinopterygians, from the North Wales Coalfield.
Grossart (Reference Grossart1868), however, described the geology and stratigraphy of the workable coals and ironstones from the Upper Coal Measures of Lanarkshire and included a comprehensive list and brief description of the biota; nonetheless, there was no descriptive information regarding the Actinopterygii, as he referred only to their stratigraphical occurrence. The first major work on the description of Carboniferous Actinopterygii was undertaken by Traquair (Reference Traquair1877–1914), with a monograph on British Carboniferous ganoids. In this work, he described and illustrated much of the genera from the Mississippian Period, with special reference to specimens from the Calciferous Sandstone Series of Scotland. He also included a few genera from the English Coal Measures with descriptions and illustrations (Traquair Reference Traquair1886, Reference Traquair1888), but these are rather scant and incomplete. Traquair's report (Reference Traquair1881) on the fossil fishes from Eskdale and Liddlesdale described in some detail many of the fossil actinopterygii recovered from that area. Moy-Thomas & Dyne (Reference Moy-Thomas and Dyne1938) revised much of that report, and included a revision of the Rhadinichthyidae. In consequence, Gardiner (Reference Gardiner1963) revised much of the genera described by Traquair, except those which were sufficiently covered by Moy-Thomas & Dyne (Reference Moy-Thomas and Dyne1938) and, in a later paper, Gardiner (Reference Gardiner1967) included and redescribed the genera Gonatodus and proposed the new taxon Pseudogonatodus. More recently, the rhadinichthyids were further investigated by Lund & Poplin (Reference Lund and Poplin1997), with a revised definition of the family.
This paper is based on fossil material which was recovered from several coal waste tips from the Central Coalfield of the Midland Valley of Scotland. The material, being of a disarticulated or isolated nature, includes not only the dermal head bones and scales of actinopterygian fishes but also their fossilised neurocrania. These neurocrania, including the associated parasphenoids, are not a common find; they are however, frequently preserved in the shales at Greenrigg, although the neurocrania are usually found compressed laterally or dorsoventrally.
Indeed, several papers have been written on the anatomy and morphology of the actinopterygian neurocrania. Day (Reference Day1915), for example, described at great length the parasphenoid of Rhadinichthys monensis, a fossil actinopterygian which is well distributed throughout the British Coal Measures and is the most frequently encountered fossil fish in North Lanarkshire material. Watson (Reference Watson1925, Reference Watson1928), nonetheless, presented the first detailed descriptions of the palaeoniscoid actinopterygian neurocranium from specimens from the coal measures of Kansas, USA. He continued by describing the palate and associated palatoquadrate apparatus of fossil specimens from Scotland and a number of specimens from the coal measures of Derbyshire and Staffordshire in England, taxa which are also found in North Lanarkshire material: Elonichthys binneyi, Mesonichthys aitkeni, Elonichthys semistriatus, Elonichthys caudalis and Platysomus parvulus. Worthy of note is that the scales of Elonichthys semistriatus have a characteristic ornament on the exposed surface and are occasionally encountered in North Lanarkshire coal tip material. Up until now, these scales have only been found at Calderhead, whereas the remains of M. aitkeni, P. parvulus and scales believed to be those of Elonichthys caudalis are found at all the coal tip locations, especially in the stratigraphically younger shales at Longriggend and Wester Bracco. Furthermore, Poplin and Veran (Reference Poplin, Veran and Milner1996) redescribed the endocranium and visceral skeleton of Coccocephalus wildi from the Lower Coal Measures of Lancashire and Coates (Reference Coates1999) wrote extensively on the neurocranium of Mesopoma planti and included a phylogenetic study of the interrelationships of certain lower Actinopterygii from the coalfields of Lancashire and Staffordshire.
This paper attempts to further our understanding of the Pennsylvanian fossil record of the early actinopterygian fishes from the Central Coalfield of the Midland Valley of Scotland. Moreover, due to the available material being disarticulated or isolated, a study was undertaken which collated information from the reconstruction of various bones, mostly dermal head material. This information was then used to erect ten new taxa, to study possible relationships with those taxa and to further our knowledge of two previously known but insufficiently documented species.
1. Geology and stratigraphy
All samples were collected over a period of several years from five localities in the Midland Valley of Scotland (Figure A). The collection sites are all North Lanarkshire coal shale tips: Greenrigg [NS 9146 6259]; Calderhead (near Shotts) [NS 8774 6164]; Dewshill [NS 8531 6385]; Ardenrigg (near Wester Bracco) [NS 8264 6570] and Longriggend [NS 8171 6917]. Stratigraphically, the Greenrigg shale tip contains the oldest material and is found geologically very close to the base of the Scottish Coal Measures Formation. The base of this formation is the Lowstone Marine Band (or local equivalent) (Forsyth et al. Reference Gardiner1996) and is at a slightly higher stratigraphical level than the base of the Subcrenatum Marine Band of the Pennine Coal Measures Group of England and Wales (Powell et al. Reference Powell, Chisholm, Bridge, Rees, Glover and Besly2000).

Figure A Map of southern Scotland showing the Midland Valley study area.
Fossils studied were from the Carbonicola communis chronozone of the Westphalian A stratigraphic range (Bashkirian: 323.2–315.2 Ma). This is Pennsylvanian strata which were deposited in a fluvio-deltaic environment and which incorporated a rock sequence consisting of a repetitive sedimentary cycle, encompassing a mudstone or fireclay at the base and a root-bed and coal at the top. These were laid down in equatorial conditions in a wet tropical climate, which resulted in peat swamps and bogs and which eventually produced extensive deposits of coal and the associated ironstones. Fossil material was collected from finely laminated carbonaceous shales from the Drumgray coal waste tips. These shales would normally have formed from layers of clay or mudstones below the coal and ironstone bands; however, in this case the shales were formed above the bands of the Drumgray coal seam. As with many coal seams, the Drumgray coal was given the name by the mining company who first wrought this coal at Drumgray near Airdrie, North Lanarkshire. A list of the flora and fauna assemblage for this facies is given in Table 1.
Table 1 List of faunal material collected from North Lanarkshire coal waste tips
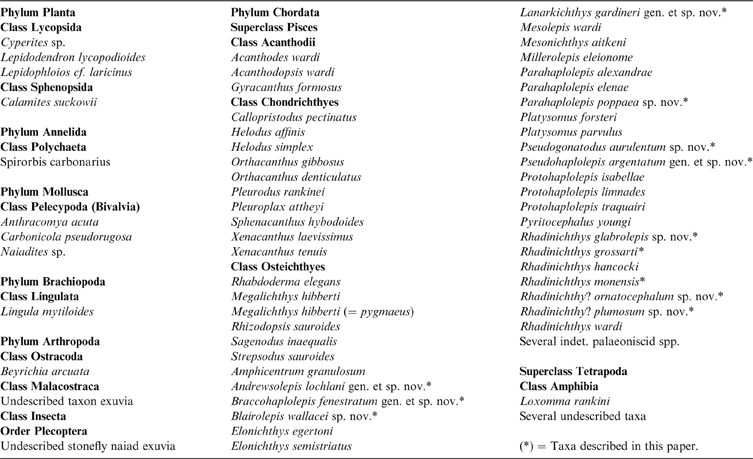
Plant material is not particularly common in the shale, but Lepidophloios and Calamites are relatively abundant in some parts. The commonest fossils are the bivalves, with the non-marine mussels Anthracomya acuta, Carbonicola pseudorugosa and Naiadites sp. the most abundant. Anthracomya is often found in bands 200–300 mm in thickness; these mussels are invaluable for identifying stratigraphical sequences, hence the geological age of the strata (Trueman & Weir Reference Trueman and Weir1946; Calver Reference Calver1969). Lingula mytiloides and the teeth of the chondrichthyans Helodus simplex and Callopristodus pectinatus are often encountered where there are abundant ‘palaeoniscoid' actinopterygian fossils, and in addition, with respect to the rocks which contain haplolepid material, the ostracod Beyrichia arcuata is always found in large numbers. In the case of the osteichthyans, Rhabdoderma elegans and Rhizodopsis sauroides are by far the most abundant of the larger fishes, with Rhizodopsis the only vertebrate to be found, so far, as articulated specimens in the coal shales. A variety of mandibles and other bones of small tetrapods are frequently found at the Greenrigg spoil tip but less often at the other locations.
Table 2 shows the stratigraphic distribution and relative abundance of the early actinopterygian material under study, collected from the North Lanarkshire coal spoil tips. The most abundant species encountered at these tips is Rhadinichthys monensis, the remains of which are found at all the locations and account for about 95 % of the specimens collected at Greenrigg, with Rhadinichthys glabrolepis sp. nov. occupying most of the remaining 5 %. Remains of Rhadinichthys grossarti are found at all locations except for the slatyband ironstone areas and this is the most frequently encountered species at Calderhead and Longriggend, accounting for about 70 % of the material from each tip. At Calderhead, specimens of R. monensis form the bulk of the remainder of the material; whereas at Longriggend the remains of Mesonichthys aitkeni and the eurynotiform Platysomus parvulus tend to form the bulk. The most abundant species encountered at Dewshill and Ardenrigg (Wester Bracco) are the eurynotiform Amphicentrum granulosum, R. monensis and species of Haplolepidae, with material from Ardenrigg having the greatest diversity in actinopterygian material as compared with the other locations.
Table 2 Stratigraphic distribution and relative abundance of basal actinopterygian remains collected from North Lanarkshire coal waste tips

Interpretation of chart: light grey = rare; dark grey = relatively abundant; black = abundant; dotted line = distribution uncertain.
A comparison of flora and fauna from the open water brackish facies of Joggins, Nova Scotia, a facies that is also of Bashkirian age (Falcon-Lang et al. Reference Falcon-Lang, Benton, Braddy and Davies2006, table 1), contains a fossil assemblage similar to that of the Lower Coal Measure Formation of Scotland. The table of biota from Joggins material, albeit more complete due to the long collection period, illustrates that a similar non-marine faunal assemblage existed.
2. Material, methods and sample collection
The material used for this study was obtained from slabs of finely laminated carbonaceous shale containing incomplete and disarticulated specimens in situ, and includes isolated skull roofs and head dermal elements and scales preserved en masse. Actinopterygian specimens were usually preserved showing excellent detail and possess a metallic sheen, and if undamaged, these specimens exhibit three distinct lamina of different material, a bony layer sandwiched between an outer brown-coloured one, most likely ganoine, and a thin lower layer, which in some specimens was all that remained. Moreover, the metallic sheen is highly lustrous and this quality remains even when the material has been exposed to normal atmospheric conditions for an indefinite period.
Slabs of laminated shale which contain the remains of one or two individual specimens only were selected for investigation; this was to simplify identification, as some slabs comprise a complex mass of the disturbed remains of many taxa. Also required for the reconstruction of the dermal skulls of the individual fish is familiarisation with the properties of the fossils; i.e., sculpture, texture, colour, etc. The association of fossils within the matrix is also important and the arrangement or disposition of the material throughout was noted. Also worth noting is the distribution (or absence) of specimens from other collection locations.
The small size of the specimens required the use of a Wild stereomicroscope throughout this study. For photographic material, a Sony DSC-W17 digital camera fitted with a Unilink digital camera adapter was attached to the stereomicroscope for all close-up images. In addition, a Swann-Morton scalpel with a No. 25 blade was employed, essentially to aid in the separation of the finely laminated layers of material but also to assist in the removal of unwanted matrix from the specimens. Dilute hydrochloric acid could not be used to etch the fossils due to the harsh reaction which would cause blackening and disintegration of the specimens and therefore, as an alternative, a solution of hydrogen peroxide (100 vol.) was used and, although slower in removing the matrix, was found to be both effective and less corrosive on most specimens.
The line drawings of the skull of preferred taxa were produced from the reconstructions of the dermal head bones. This was accomplished by first collecting the appropriate bones from the available material, photographing this material and then tracing the photograph to the same scale onto acetate sheet. The acetate tracing of the skull roof was then selected and, by considering the general shape or outline of the bones, the overlap areas and the relative position of the sensory canals, each bone was carefully placed into as near a natural position relative to the skull roof as possible, thus creating a more or less complete dorsal reconstruction of the dermal skull.
A similar procedure was then applied to the cheek bones, although this time the maxilla is used as the principal element, to create the lateral reconstructions of the dermal skull. The dorsal and lateral reconstructions were then cut, bent into shape and joined together. This was achieved by first connecting the dorsal reconstruction with the upper parts of the cheek area of the lateral reconstructions, using the alignment of the sensory canals and any overlap area as a guide. This was followed by connecting the nasal area with the ramus of the maxilla, linking the bases of the clavicles together and then joining both sides of the mandibles at the symphysis. The complete assembly thus produced a more-or-less three-dimensional shape for the reconstruction of the line drawings of the dermal skull bones.
The respective line drawings were then produced first by hand and then refined using Microsoft Paint or Paint.NET. All dermal skull reconstructions are shown in a horizontal direction, from right to left; drawings or images of skull roofs are vertically orientated (i.e., the anterior margin is to the top of the figure), as this direction is more appropriate for their study. All morphometric measurements are taken from actual specimen material and considered to be from adult fish, unless stated otherwise. The approximate total length of each taxon was calculated from descriptive measurements and illustrations given by Traquair (Reference Traquair1877–1914) and Westoll (Reference Westoll1944).
Reconstructing natural subjects in this way does, of course, produce inherent inaccuracies. Nonetheless, the main point is that there is a large amount of specimen material available at some of these coal waste tips and, due to the disjointed mode of preservation, reconstructing by this method is probably the only means of illustrating how the animal may appear in life.
3. Referred specimens
Table 3 contains a record of specimens examined or consulted for the construction of this paper. The first part of this list encompasses ten newly erected taxa and includes three named taxon which were further investigated as a result of the new information. The second part of the list comprises 22 actinopterygians, for which information was collated from various sources concerning each taxon and used for a comparison.
Table 3 Specimens referred to in this paper

A list of specimen material discussed and erected, including museum catalogue numbers, can be found in Appendix 1 (section 9).
4. Systematic palaeontology
Osteichthyes Huxley, Reference Huxley1880
Class Actinopterygii Woodward, Reference Woodward1891
Subclass Actinopteri Cope, Reference Cope1871
Family Rhadinichthyidae Romer, Reference Romer1945
Genus Rhadinichthys Traquair, Reference Traquair1877
Diagnosis. (After Traquair 1897; limited modification after Moy-Thomas & Bradley Dyne Reference Moy-Thomas and Dyne1938). Body fusiform, often elongated: caudal prolongation weakly developed; body scales rhombic; principal rays of the pectoral fin only articulated distally: dorsal fin far back with its anterior edge slightly in front of that of anal, both fins being triangular and acuminate, the latter being the larger: caudal fin deeply cleft and inequilobate: all fins have well-developed fulcra, and lepidotrichia bifurcating distally. Skull with rather prominent rostrum, fairly large orbit, wide gape and very oblique suspensorium: opercula oblong: subopercular smaller and squarish: nasals meet the dermosphenotics: two suborbitals present: head bones ornamented with ridges and tubercles; jaws armed with conical incurved laniaries and numerous smaller teeth.
Revised diagnosis. (After Lund & Poplin Reference Lund and Poplin1997). Prominent snout and subterminal mouth; rostropostrostral; absence of premaxillae resulting in a rostral notch below the rostropostrostral and between the antorbitals; reverse-L shaped antorbital; no supraorbital, tectal, and pineal bones, nor pineal foramen; lack of a profundus canal; three infraorbitals, the third (postero-dorsal) one is T-shaped and contacts the nasal; two suborbitals; operculum higher but shorter than suboperculum; presence of an anamestic anocleithrum; elongate clavicles in contact along midline, without an interclavicle; dorsal fin triangular, posteriorly placed, its origin hardly in front of that of the anal fin, or opposed to it; caudal fin deeply cleft and inequilobate; fin rays distally bifurcated in all fins; low ventrolateral abdominal scale rows.
Type species. Rhadinichthys canobiensis Traquair.
Remarks. In 1850, Sir Philip de M. Grey-Egerton first described Rhadinichthys monensis as Palaeoniscus monensis with the figured specimen of a detached scale. Traquair (Reference Traquair1878) added a further description of the fish, especially the body form and the disarticulated rays of the pectoral fin. Additional information was then added by Wellburn (Reference Wellburn1900), by including the arrangement of the fins in general. However, as far as the present author is aware, a formal diagnosis or indication of a specimen having been designated as the holotype has never been presented of this abundant fossil fish from the British Coal Measures.
Palaeoniscus monensis Egerton, Reference Egerton1850
Rhadinichthys monensis Traquair, Reference Traquair1878
(Figs 1, 2a, 3, 4, 5, 7, 8, 9a, 10a)
Diagnosis. (Original). Ornamentation of the skull roof is of coarse, occasionally broken ridges and rounded tubercles, with the frontals having a chevron pattern of ridges which are directed posteriorly; dermopterotics exceptionally long, extending from base of parietals to approximately half the length of the frontals; middle pit-line conspicuous and of an unusual V-shape; cleithrum with a long ventral pit-line; scales with distinctive ornamentation of six to eight fine striae traversing the anterior margin and with four or five broad, oblique ridges commencing about midscale and terminating at the posterior margin as long denticulations.
Locality. Remains of Rhadinichthys monensis are found at all coal shale tips throughout the Scottish Lower Coal Measures where ichthyofauna are encountered (see Table 2). This species accounts for about 95 % of the specimens collected from the Slatyband Ironstone facies.
Material. About a dozen fragments of slate containing disarticulated specimens in situ and a large amount of material, including isolated skull roofs and endoskeletal elements (specimens GLAHM 152825 to GLAHM 152830), from the above shale tips.
Description. A tentative reconstruction of the complete skull of Rhadinichthys monensis is shown in Figure 1.

Figure 1 Rhadinichthys monensis, tentative reconstruction of the dermal head bones: (a) left lateral view and (b) dorsal view. Heavy dotted lines indicate path of sensory canals; lighter dotted lines indicate possible arrangement of some smaller elements. Abbreviations: ang = angular; ant = antorbital; br = branchiostegal plates; cl = cleithrum; clv = clavicle; den = dentary; dhy = dermohyal; dpt = dermopterotic; dsph = dermosphenotic; esc = extrascapular; ethc = ethmoid commissure; fr = frontal; ioc = infraorbital canal; ju = jugal; lac = lacrymal; mdc = mandibular canal; mpl = median pit-line; mx = maxilla; na = nasal; op = operculum; pa = parietal; pcl = postcleithrum; pop = preoperculum; popc = preopercular canal; pt = posttemporal; ptc = posttemporal sensory canal; ro = rostrum; scl = supracleithrum; sclc = supracleithrum sensory canal; slp = sclerotic plates; soc = supraorbital canal; sop = suboperculum. Scale bar = 2 mm.
Skull roof. A dorsal view of the skull roof (GLAHM 152825) is shown in Figure 2a. The form approximates an isosceles triangle; the frontals are sub-rectangular, twice as long as broad; the parietals are trapezoid in form, about half the size of the frontals; the dermopterotics extend from base of parietals to approximately half the length of the frontals in the form of an elongate quadrilateral; the transverse suture is weakly sinuous; the middle pit-line is very strong and V-shaped, proximal to mid-sagittal suture, extending across the parietal and terminating briefly in the dermopterotic region; anterior and posterior pit-lines have not been observed; the anterior margin of the skull roof with median recess for articulation with the rostrum and lateral notches for articulation with posterior limbs of the nasals; the lateral margin of the frontals slightly concave; the posterior margin of the skull roof is lobed; the supraorbital canals are observed as gentle sinusoid curves along lateral margins of frontals and terminating mid-parietal region; sensory canals are clearly discernible along the lateral margins of dermopterotics; a pineal foramen absent. The ornamentation of the skull roof is of coarse, occasionally broken ridges anteriorly, passing into rounded tubercles posteriorly especially at dermopterotic and parietal regions; the ornamentation of the anterior region of the frontals is of a well-developed chevron pattern of ridges directed posteriorly; the median area of the frontals with less sculpture or smooth.

Figure 2 Diagrams of centre part of skull roof of basal actinopterygians showing pineal foramen, pit-lines and oblique/transverse sutures. Dotted lines indicate path of sensory canals: (a) Rhadinichthys monensis; (b) Rhadinichthys glabrolepis sp. nov.; (c) Rhadinichthys grossarti; (d) Rhadinichthys? ornatocephalum sp. nov.; (e) Pseudogonatodus aurulentum sp. nov.; (f) Pseudohaplolepis argentatum gen. nov.; (g) Lanarkichthys gardineri gen. nov. Not to scale.
Snout. The snout encompasses a single median rostral element, paired nasals and paired antorbitals. No dentigerous premaxillae present. The rostrum is keyhole-shaped, with a rounded anterior region occupying about one-third of its length, the ethmoidal commissure crosses dorsally and small, spiked tubercles are found ventrally; the remaining portion is broad, ornamented anteriorly with a few ridges and the posterior margin embayed for contact with the frontals; laterally there is a deep restriction between the two regions for the nasal openings. Nasal with a long, narrow limb posteriorly which abut with the dermosphenotic; anteriorly the region is broad with an almost straight terminal edge which articulates with the antorbital; medial margin with contour for attachment to the lateral margin of the rostrum and a notched area for the nasal opening; the lateral margin is gently curved with embayed area and forms the anterodorsal margin of the orbit; supraorbital canal visible along the whole length of the bone. Antorbitals approximately polygonal in form, with the infraorbital canal joining the supraorbital canal from the nasal and with the ethmoidal commissure extending medially, and when connected with the rostrum and paired nasals dorsally a rostral notch would most likely be produced.
Neurocranium. Specimens of neurocrania are not uncommonly found in material from the ironstone waste tips located around the Fauldhouse moor area. These fossils are usually encountered compressed dorsoventrally or laterally and in a rather friable carbonaceous material high in sulphur content. The parts of the specimens which are preserved include the occipital, otic, orbital and ethmoid regions and, in many specimens, the parasphenoid remains attached to the neurocranium.
In the neurocranium of Rhadinichthys monensis (GLAHM 152826; Fig. 3), the openings for the foramen magnum and notochordal canal are just visible near the posterior margin of the occipital region. The anterior part of the otico-occipital fissure is observed to intersect with the somewhat elongate vestibular fontanelles which appear to be closed, as in Howqualepis Long (Reference Long1988). The dorsal aorta is seen as a wide ridge which bifurcates anteriorly, forming two lateral dorsal aortae, and emerging mid-length of the right lateral dorsal aorta is the first epibranchial artery. However, unlike actinopterygians, in which the dorsal lateral aortae are widely spaced (such as in Cheirolepis Giles et al. (Reference Giles, Coats, Gaewood, Brazeau, Atwood, Johanson and Friedman2015), Woodichthys Coates (Reference Coates1998), Howqualepis, Kentuckia Rayner (Reference Rayner1951) and Cosmoptychius Schaeffer (Reference Schaeffer1971)), in R. monensis these features are closely spaced and near parallel. The craniospinal process is not strongly developed, but is directed posteriorly on each side of the posterior margin of the occipital. The postorbital process is much pronounced and directed anteriorly, as in Cheirolepis, Woodichthys, Howqualepis and possibly Lawrenciella Poplin (Reference Poplin1984). However, as in Leuderia Schaeffer & Dalquest (Reference Schaeffer and Dalquest1978), the ventral otic fissure of R. monensis cannot be ascertained, due to the overlapping by the large posterior lamina of the parasphenoid. Additionally, the jugular canal can be seen to traverse the most anterior region of a large jugular depression and the articulation facet for the hyomandibular is visible as an indentation along the left margin of the occipital.

Figure 3 Rhadinichthys monensis, GLAHM 152826: (a) ventral view of neurocranium with attached parasphenoid; (b) interpretive drawing of same. Abbreviations: asc.pr = ascending process of parasphenoid; bpt = basipterygoid process; crsp = craniospinal process; da = dorsal aorta; dent = denticle field of parasphenoid; epi.1 = first epibranchial artery; fm = foramen magnum; hmf = articulation facet for hyomandibular; jc = jugular canal; jd = jugular depression; lda = lateral dorsal aorta; not = notochordal canal; oof = otico-occipital fissure; por = postorbital process; psp = parasphenoid; v.fon = vestibular fontanelle. Scale bar = 2 mm.
Figure 4 shows the dorsal aspect of the skull roof of Rhadinichthys monensis (GLAHM 152827) in which skull roof material has been removed in a natural manner and allows inspection of the underlying dorsal surface of the parasphenoid. This specimen displays a complete parasphenoid in the correct orientation relative to the dermal skull roof. The medial surface of the bone in dorsal aspect is rather course in texture, though some specimens do exhibit a smoother surface; and unlike the ventral surface it lacks the denticles along its length. In most specimens of R. monensis, the buccohypophyseal canal is usually clearly visible as a minute aperture situated between the arms of the ascending processes of the parasphenoid when the skull is viewed dorsally, as with Cheirolepis and Cosmoptychius; although in the specimen illustrated the canal aperture is barely discernible. Furthermore, although the buccohypophyseal canal is only visible on the ventral surface of the parasphenoid in R. monensis, in other actinopterygians it may be observed on the dorsal surface, or on both dorsal and ventral surface, as in Cheirolepis. The articular facet for the autopalatine is shown as a large plated area with several distinct notches which mark the articular surface of the ethmoid region. The ascending process of the parasphenoid is shown to be long and directed posteriorly. The basipterygoid process of the parasphenoid is about half the length of the ascending process and is also directed posteriorly, unlike that of Woodichthys, Lawrenciella, Leuderia and Cosmoptychius, which are directed anteriorly, or are absent and replaced with short lateral stubs or extensions, as in Cheirolepis, Howqualepis and Coccocephalus Poplin & Veran (Reference Poplin, Veran and Milner1996).

Figure 4 Rhadinichthys monensis, GLAHM 152827: (a) dorsal view of skull roof of showing parasphenoid in situ; (b) interpretive drawing of same. Abbreviations: apal = articular facet for autopalatine; asc.pr = ascending process of parasphenoid; bhc = buccohypophyseal canal; bpt = basipterygoid process; dpt = dermopterotic; fr = frontal; infc = infraorbital canal; pa = parietal; por = postorbital process; psp = parasphenoid; soc = supraorbital canal. Scale bar = 2 mm.
Morphological detail is much more difficult to differentiate with specimens preserved laterally, due to the two-dimensional mode of compression. Figure 5 shows a more or less lateral aspect of the neurocranium of Rhadinichthys monensis (GLAHM 152828), with the orientation of the skull dermal bones relative to the neurocrania such as the frontals, parietals and dermopterotics and the sensory canals clearly discernible. The otic and orbital regions are relatively well preserved, with the most prominent features being the strong postorbital process; the craniospinal process; the almost elliptical infraorbital septum; the very conspicuous basipterygoid process; and the parasphenoid.

Figure 5 Rhadinichthys monensis, GLAHM 152828: (a) lateral view of neurocranium. (b) interpretive drawing of same. Abbreviations: asc.pr = ascending process of parasphenoid; bpt = basipterygoid process; crsp = craniospinal process; dpt = dermopterotic; dptc = dermopterotic sensory canal; fr = frontal; ios = infraorbital septum; pa = parietal; por = postorbital process; psp = parasphenoid; soc = supraorbital canal; v. fon = vestibular fontanelle. Scale bar = 2 mm.
Line drawings of the parasphenoids from different actinoperygians from the Lower Coal Measures of Scotland are shown in Figure 6 (specimens GLAHM 152835–152842).

Figure 6 Parasphenoids from various coal waste tips of North Lanarkshire (Scottish Lower Coal Measure Formation (Pennsylvanian)). Scale bar = 2 mm.
Palatoquadrate. Figure 7 shows the mesial surface of the palatoquadrate of Rhadinichthys monensis with associated maxilla and preoperculum (GLAHM 152829). The bone is long and slender, narrowing to almost a point anteriorly and planar posteriorly with a small, deep posteroventral extension. The different elements of the palatoquadrate are not discernible as the whole entity is fused as a single unit though the ventral margin of the palatoquadrate can be seen to be edentate. The articulation for the endocranial basipterygoid process is in the form of a small notch situated along the dorsal margin about one-third of the way from the anterior margin.

Figure 7 Rhadinichthys monensis, GLAHM 152829, mesial view of palatoquadrate with associated preoperculum and maxilla attached. Abbreviations: bp = basal process; max = maxilla; pop = preoperculum; pop.c = preopercular canal; pq = palatoquadrate; sy = symplectic. Scale bar = 2 mm.
Hyomandibular. A mesial view of the right hyomandibular of Rhadinichthys monensis (GLAHM 152830) is shown in Figure 8. This bone has the form of an inverted ‘V' and its natural orientation is tilted slightly forward as shown in the illustration. The two shafts of the ‘V' form an obtuse inner angle are about equal in length with the anterior shaft paddle-shaped and the width of the posterior shaft parallel along its whole length. A groove runs anteroposteriorly along the medial surface of the hyomandibular and mesial side of the bone and close to mid-point of the dorsal margin there is a small opercular process. The shape of this bone is such that it fits into a groove close to the dorsal margin of the palatoquadrate (Poplin & Veran Reference Poplin, Veran and Milner1996), with the anterior margin planar to enable articulation with the endocranium. There is also a small hole or slit at the distal end of the paddle-shaped shaft, which could be interpreted as a hyomandibular foramen. On occasion, pieces of dermohyal are found still fused to the lateral surface. In GLAHM 152829 (Fig. 7), the remains of what appears to be a symplectic (or interhyal) lies close to the posterior part of the palatoquadrate complex; this small bone would have connected with the posterior shaft of the hyomandibular as part of the hyoid arch.

Figure 8 Right hyomandibular of Rhadinichthys monensis, GLAHM 152830. Scale bar = 2 mm.
Cheek, orbital and lower jaw regions. This area includes the maxilla, sclerotic bones, infraorbital bones (jugal, lacrymal and dermosphenotic), suborbitals, preoperculum, quadratojugal, dermohyal and dentary. The maxilla has a long, narrow, upward curved suborbital ramus and expanded posterior plate approximating an isosceles trapezoid: posterior plate planar dorsally with posteroventral elongation; ornamentation of short tubercles along the complete length of the suborbital ramus and fine, near straight ridges running obliquely from posterior region of plate to the posteroventral extension; mesial surface with a curved horizontal furrow along most of its length; dentary margin of maxilla curved and armed with teeth of two sizes, moderately large incurved teeth and smaller ones about half the size as far back as the posteroventral elongation. Small, curved and unornamented bones of varying thickness constitute the sclerotic ossicles, the thicker bones probably being located dorsally (White Reference White1939, p. 45); individual sclera are a relatively common find with four the maximum found still connected. The lacrymal is a thin bone and ventral to the orbit with the infraorbital canal running along its whole length; the jugal, is a large crescentic bone, rounded dorsally and located at the posteroventral part of the orbit, with the sensory canal running along its length with about eight branches radiating from the canal; the dermosphenotic (posterodorsal infraorbital) is a crescent-shaped bone which tapers anteriorly to abut with the narrow posterior margin of the nasal, the broad posterior margin of the dermosphenotic is embayed for connection with the dorsal margin of the jugal (posterior infraorbital) and the complete length of the mesial margin in contact with the dermosphenotic, the infraorbital canal intersects posteriorly, passes briefly across the bone and emerges at the medial margin where it enters the anterior margin of the dermopterotic. Suborbitals have not been identified so far and, considering the outline and shape of other cheek bones, they may or may not exist. The preoperculum is the shape of an inverted ‘V' with the two limbs forming an obtuse angle between, this angle agreeing with the angle of the posterosuperior margin of the maxilla; the anterior limb is longer than the posterior one and the preopercular canal runs along the complete length of the dorsal margin of the bone. A quadratojugal has not been observed. Although hyomandibulars of this species are frequently found, dermohyals (see Fig. 8) are only rarely preserved and usually not in a condition suitable for description; pieces of dermohyal are, however, occasionally found still fused to the upper limb of the hyomandibular and the general form of the remains indicates a narrow, triangular bone located between the anterodorsal margin of the preoperculum and the anterosuperior margin of the operculum. The dentary is a slender bone, deeper posteriorly with the mandibular sensory line clearly visible along the ventral border of the outer surface only; the angular which makes up the posterior margin is only occasionally preserved. However, the surangular is well represented as a triangular or rectangular extension to the posterior margin; the teeth of the dentary are identical to those of the maxilla, that is, of two sizes, moderately large incurved teeth and smaller ones about half the size of the lanaries and extending as far back as the dorsal margin of the surangular; ornamentation of the inner and outer surface is of long, fine ridges radiating in herring-bone like fashion from the posteroventral region and terminating in an acute angle along the dentary margin or more or less confluent with the ventral border.
Operculogular. The operculogular system comprises the operculum, the suboperculum, the branchiostegals and the gulars. The operculum is approximately rectangular and twice as long as broad; anterodorsal and posteroventral angles acute; ventral margin sinusoid; anterior margin embayed along upper third of the margin, the remainder almost straight with an unornamented overlap for attachment of the preoperculum; ornamentation is of small tubercles anterodorsally changing to low ridges and following the contours of the bone along most of the posterior and ventral regions. The suboperculum is about half the size of the operculum, fan-shaped with both dorsal and ventral margins sinusoid, anterior margin planar with unornamented overlap for attachment of the preoperculum; posterior margin convex; the dorsal margin also with overlap for the ventral margin of the operculum; ornamentation of low tubercles along anterior margin, the remaining surface with low ridges running dorsoventrally.
The branchiostegals are slender, tapered at both ends and ornamented with small, pointed tubercles directed posteriorly; the exact number of branchiostegals is not known. The gulars (Fig. 9a) consist of two lateral (possibly asymmetrical) plates and one median plate; the median gular is pear-shaped, with a V-shaped pit-line slightly anterior of mid area, and ornamented with a few broken ridges which follow the contours of the anterior and posterior margins, the remaining area with a sparse amount of posteriorly directed small tubercles; the lateral paired gulars with anteromedial margin sigmoid for coupling with the median gular, and with the lateral and posterior margins angled; ornamentation of sparse amount of posteriorly directed small tubercles. A number of isolated gulars are preserved in slab GLAHM 152829.

Figure 9 Reconstruction of median and lateral gulars and first paired branchiostegals of (a) Rhadinichthys monensis and (b) Rhadinichthys glabrolepis sp. nov. from the Greenrigg coal tip. Dotted line indicates actual shape of lateral gulars. Scale bar = 2 mm.
Extrascapulars and posttemporals. Extrascapulars have not been identified so far. The single paired posttemporals are large, triangular elements, joined mid-line and spanning the posterior margin of the skull roof; there is a large overlap on the embayed anterior margin; the lateral and posterior margins are curved and the inner (medial) margin is linear; the anterolateral corner also incorporates the posttemporal sensory canal; ornamentation is of coarse ridges running in a lateroposterior direction and directed along the acute angle of the posterior margin.
Pectoral girdle. The pectoral girdle comprises the supracleithrum, cleithrum, postcleithrum and clavicle. The supracleithrum is a large, slender bone which tapers dorsoventrally and has a large dorsal triangular protuberance which is overlapped by the posttemporal; the path of the supracleithrum sensory canal is clearly seen to traverse the bone from the dorsal protuberance to the mid posterior margin; ornamentation is of fine ridges running in an oblique manner from the lateral line to the posterosuperior margin terminating as acute denticulations, and as long, fine, curved ridges following the contours across the medial face. The cleithrum is a large inverted S-shaped bone with an expanded posterosuperior region and a squared ventral region; a vertical ridge runs medially from the anterodorsal tip to the squared ventral area; a long ventral pit-line can be seen to bisect this plate and there is a deep posterior embayment for the pectoral fin; ornamentation of fine vertical ridges anteriorly, more widely spaced ridges posteriorly and fine straight ridges running across the ventral plate. The clavicle is a lanceolate bone with a tall dorsal spur; ornamentation is of fine ridges running anteroposteriorly. A postcleithrum has not yet been identified, although it is very probable that one exists. The pectoral fin endoskeleton has not, as yet, been found or identified as belonging to R. monensis.
Squamation. The scales (Fig. 10a) are square to slightly rhomboidal in form, the strength of ornamentation being very variable with location and within individual specimens; the dorsal peg is strong, and the mesial articulation for the inferior scale is also strong but the central keel is weak or absent. Scales may be almost smooth or ornamented with six to eight fine striae running down the anterior margin and following the ventral margin, and with four or five broad, oblique ridges commencing about mid-scale and terminating at the posterior margin as long denticulations.

Figure 10 Typical flank scales of early actinopterygians referred to in this study: (a) Rhadinichthys monensis (upper and lower surface); (b) Rhadinichthys glabrolepis sp. nov. (upper and lower surface); (c) Rhadinichthys grossarti (upper surface); (d) Rhadinichthys? ornatocephalum sp. nov. (upper surface); (e) Rhadinichthys? plumosum sp. nov. (upper surface); (f) Pseudogonatodus aurulentum sp. nov. (upper surface); (g) Pseudohaplolepis argentatum gen. nov. (upper surface); (h) Lanarkichthys gardineri gen. nov. (upper surface). Scale bars = 0.5 mm.
Length of fish is up to 90 mm.
Rhadinichthys glabrolepis sp. nov. (Figs 2b, 9b, 10b, 11, 12)
Holotype. GLAHM 152831, skull roof.
Diagnosis. Skull roof with long, broken ridges or irregular tubercles, with median area of frontals lacking any form of ornamentation; anterior margin of skull roof with distinct median recess for articulation with the rostrum; anterior limb of preoperculum with projecting anterodorsal spur; dentary with row of circular marks along ventral rim; scales generally without ornamentation.
Remarks. Following Lund and Poplin's revised diagnosis on the Rhadinichthyidae (Lund & Poplin Reference Lund and Poplin1997), this taxon has been placed in that family as a result of the following observations: (i) a reconstruction of the dermal head bones show the snout to be prominent, there are paired antorbitals, and the premaxillaries are absent; the dermosphenotics are more or less T-shaped; (ii) pineal foramen absent; (iii) clavicles are elongate and probably connect midline as interclavicles have never been identified; (iv) the caudal fin is deeply cleft and inequilobate with the fin rays distally bifurcated. Additionally, due to the arrangement of the bones of the skull roof following a very similar pattern to that of Rhadinichthys monensis, the taxon has been given the generic title of Rhadinichthys.
Type locality. Specimens were collected from slatyband ironstone shale at the Greenrigg coal waste tip [NS 9146 6259], the only known locality.
Material. Two pieces of slate containing parts of disarticulated specimens in situ (GLAHM 152851 and GLAHM 152844) and a large amount of isolated dermal elements collected from the above shale tip.
Etymology. The species name refers to the almost completely smooth sculpture of the scales: glabrous = smooth and lepis = scale. This is the only species encountered so far where the scales lack a strong surface sculpture.
Description. A tentative reconstruction of the complete skull of Rhadinichthys glabrolepis is shown in Figure 11.

Figure 11 Rhadinichthys glabrolepis sp. nov., tentative reconstruction of the dermal head bones: (a) left lateral view; (b) dorsal view. Heavy dotted lines indicate path of sensory canals; lighter dotted lines indicate possible arrangement of some smaller elements. Abbreviations: ang = angular; ant = antorbital; br = branchiostegal plates; cl = cleithrum; clv = clavicle; den = dentary; dhy = dermohyal; dpt = dermopterotic; dsph = dermosphenotic; esc = extrascapular; ethc = ethmoid commissure; fr = frontal; hpl = horizontal pit-line; ioc = infraorbital canal; ju = jugal; lac = lacrymal; mdc = mandibular canal; mesc = median extrascapular; mpl = median pit-line; mx = maxilla; na = nasal; op = operculum; pa = parietal; pcl = postcleithrum; pop = preoperculum; popc = preopercular canal; pt = posttemporal; ptc = posttemporal sensory canal; ro = rostrum; scl = supracleithrum; sclc = supracleithrum sensory canal; slp = sclerotic plates; soc = supraorbital canal; sop = suboperculum. Scale bar = 2 mm.
Skull roof. The skull roof (Fig. 2b) has the form of an isosceles triangle; the frontals are approximately quadrilateral, twice as long as broad; the parietals more or less the form of a trapezoid and about one-third the size of the frontals; the dermopteroptics are near ovoid except for the irregular anterior margin which extends only as far as the strongly sinuous transverse suture; the middle pit-line runs from about the mid parietal region and terminates as it briefly enters the dermopterotic, anterior and posterior pit-lines are not observed; the anterior margin of skull roof with median recess for articulation with the rostral and lateral notches for articulation with posterior limbs of the nasals; the lateral margins of the frontals are slightly concave; the posterior margin of the skull roof lobed; the supraorbital canal extends as a rather weak, almost straight sinusoidal curve from anterior margin of frontals and terminating at mid parietal region; the sensory canals clearly discernible along lateral margins of dermopterotics; pineal foramen absent. Ornamentation of skull roof of long, occasionally broken ridges or irregular tubercles running in anteroposterior direction with median area of frontals lacking any form of ornamentation.
Snout. The snout comprises a single median rostral bone, paired nasals and paired antorbitals. No dentigerous premaxillae are present. The rostral is keyhole-shaped with a rounded anterior region occupying about one-third of its length and with the ethmoidal commissure crossing anterodorsally with the remaining portion rectangular; the posterior margin embayed for contact with the frontals and ornamented with short, flat, irregular tubercles covering most of the anterior region; the rounded anterior region of the rostral is ornamented with longer, flat tubercles; the lateral margin with deep restrictions between the two regions for the nasal openings. The nasal is broad, with a long, narrow limb posteriorly which abuts with the dermosphenotic; anterior area is broad, with an almost straight terminal edge which connects with the antorbital; the medial margin is quite straight, but angled for attachment to the lateral margin of the rostral and with deeply notched area for the nasal opening; the lateral margin of nasal is gently curved and forms the anterodorsal margin of the orbit; the supraorbital canal is visible along the whole length of the bone. Antorbitals are approximately polygonal in form, with the infraorbital canal joining the supraorbital canal from the nasal and with the ethmoidal commissure extending medially; in the event of the antorbitals being connected with the rostrum and paired nasals, a rostral notch would most likely be produced.
Cheek, orbital and lower jaw regions. Maxilla with long, narrow, upward curved suborbital ramus and expanded posterior plate; posterior plate planar dorsally with deep posteroventral elongation; suborbital ramus twice as long as the posterior plate; ornamentation of moderately coarse ridges along the suborbital ramus and fine, near straight ridges running obliquely from posterior region of plate to the posteroventral extension; mesial surface with a curved horizontal furrow along most of its length; dentary margin of maxilla strongly curved and armed with teeth of two sizes, large conical teeth and smaller ones about half the size as far back as the posteroventral elongation. The sclerotic bones are small, curved and unornamented ossicles, similar to those of R. monensis but slightly thinner; there are probably four in number, with the thicker bones located dorsally. The infraorbital bones are the lacrymal, a thin bone ventral to the orbit with the infraorbital canal running along its whole length; the jugal, a large more or less crescentic bone with a planar dorsal margin and located at the posteroventral part of the orbit with the infraorbital canal running along its length, this canal has about seven radiating branches; the dermosphenotic (posterodorsal infraorbital) is a crescent-shaped bone which tapers anteriorly to abut with the narrow posterior margin of the nasal, the broad posterior margin is embayed for connection with the dorsal margin of the jugal (posterior infraorbital) and the complete length of the mesial margin in contact with the dermopterotic and the frontal; the infraorbital canal intersects posteriorly, passes briefly across the bone, emerges at the medial margin where it enters the lateral margin of the dermopterotic. Suborbitals have not been identified so far and, considering the outline and shape of other cheek bones, they possibly do not exist. The preoperculum has two limbs, an anterior limb and a posterior limb which form a 90° angle between them, and which agrees with the angle of the posterosuperior margin of the maxilla; the anterior limb is slightly longer and much wider anterolaterally than the posterior one and has a projecting anterodorsal spur; the preopercular canal runs along the complete length of the dorsal margin of the bone. A quadratojugal has not been observed and hyomandibulars do not seem to be preserved. The dentary is a thickset bone, deep posteriorly and tapering sharply anteriorly; the lateral surface has a row of circular marks along a broad ventral rim running in an anteroposterior direction, these rings are faint or absent on the mesial side; the mandibular sensory line is medial to the circular marks and clearly visible ventrally; the angular which makes up the posterior margin is only occasionally preserved, however the surangular is well represented as a triangular or rectangular extension to the posterior margin; the teeth of the dentary are identical to those of the maxilla, that is, of two sizes, large conical teeth and smaller ones about one-third the size of the laniaries and extending the length of the dentary margin but stopping short of the dorsal margin of the surangular; ornamentation on both sides of long, fine ridges running horizontally but slightly inclined upwards anteriorly from the posteroventral region and terminating at an angle along the dentary margin.
Operculogular. The operculum lies at an oblique angle posterior to the preoperculum as in R. monensis, it is approximately sub-ovate, rounded dorsally with the ventral and posterior margins convex; ornamentation is of a few broken ridges dorsally with low ridges following the contours of the bone along most of the posterior and inferior areas. The suboperculum is a little more than half the size of the operculum, more or less rectangular; ornamentation as operculum with low ridges following the contours of the bone along the posterior and inferior areas. The branchiostegals are slender, tapered at both ends and ornamented with a few lines directed posteriorly, the exact number is not known. The gulars consist of two lateral (possibly asymmetrical) plates and one median plate (Fig. 9b); the median gular is diamond-shaped, with two parallel pit-lines slightly anterior of mid area and ornamented with a few concentric lines along the lateral and posterior margins; the lateral paired gulars with an acute lateroposterior angle with a rectangular tab anteriorly; ornamentation is of fine lines directed in a posterior direction.
Extrascapulars and posttemporals. Paired extrascapulars have not been identified, although a possible median extrascapular exists. The single paired posttemporals are very similar in structure to those of R. monensis; they are large, triangular elements, joined midline and span the posterior margin of the skull roof with the anterior margins curved anteromesially; anterolateral corner with posttemporal sensory canal; the ornamentation is of low ridges following the contour of the lateral, mesial and posterior margins.
Pectoral girdle. The supracleithrum is the usual large bone with a posterosuperior protuberance tapering from about mid-bone with the remaining posterior region inflated; the path of the supracleithrum sensory canal is clearly seen to traverse the bone from the dorsal protuberance to the mid posterior margin; the ornamentation is of broad ridges at the posterior inflated region running in an oblique manner mostly from the anterior border to the ventral margin. The cleithrum is an inverted S-shaped bone with an expanded posterosuperior region and a large, squared ventral region; a strong vertical ridge runs medially from the anterodorsal tip to the squared ventral area; a ventral pit-line is absent; ornamentation may be absent or there may be a few very fine vertical ridges anteriorly and a fine straight line separating the ventral plate from the dorsal region. The clavicle is a broad lanceolate bone with a tall dorsal spur and a long posteriorly directed ventral spur; ornamentation seems to be absent. A postcleithrum has not yet been identified though it is very probable one exists.
Caudal fin. Caudal fin (Fig. 12) incorporates about 50 rays, with dorsal and ventral fin lobes separated by a conspicuous medial cleft at posterior margin. Dorsal lobe rounded posteriorly and longer than ventral lobe, dorsal margin of lobe with about 18 lanceolate ridge scales; ventral lobe triangular with fin rays bifurcated distally; fulcra evident along ventral margin; caudal basal region with several large basal fulcra and most scales on the axial lobe rhombic and smooth.

Figure 12 Rhadinichthys glabrolepis, GLAHM 152851: (a) caudal skeleton, ventral lobe detached and rotated a few degrees anticlockwise; (b) interpretive drawing of specimen. Abbreviations: axf = axial fringing fulcra; axl = axial lobe; cb = caudal basal fulcra; cp = caudal peduncle; dcr = dorsal caudal lobe fin rays; ff = fringing fulcra; vcr = ventral caudal lobe fin rays. (Nomenclature after Sallan & Coates Reference Sallan and Coates2013). Scale bar = 2 mm.
Squamation. The scales (Fig. 10b) are square and more or less without ornamentation; dorsal peg tall and thin, ventral articulation for inferior scale also strong but central keel weak or absent; anterior margin with a few fine striae occasionally turning ventrally; posterior margin entire or with a few coarse denticulations. This is the only known genera so far found at the locations described in this study in which the scales are almost completely without ornamentation; all other genera have the surface of the scales adorned with some form of ornamentation and often this sculpture is in the form of an intricate pattern, which makes the different species easily recognisable.
Approximate length of fish is 90 mm or more.
Rhadinichthys grossarti Traquair, Reference Traquair1878
(Figs 2c, 10c, 13, 14, 15, 16)
Type specimen. An almost complete specimen (NMS G.1950.38.90.), but without the head, is kept in the National Museums of Scotland.
Diagnosis (after Traquair). Ornamentation of the skull roof with tubercles or long posteriorly directed spines which may be extremely profuse in some specimens; parietals diminutive and irregular in form; no pit-lines are observed; premaxillaries present: cleithrum large and of an unusual shape; about six large, rounded-cordate ridge scales anterior to the dorsal fin; scales with four to six ridges which terminate posteriorly as sharp denticulations and anterior striae faint or absent.
Remarks. Traquair mentions the specific characters of this small fish using the type specimen, two other specimens and fragmentary material:
“The shape is slender, its depth at the ventral fin being not quite ¼ inch, from which it tapers to a very delicate tail-pedicle. The ventral fin is small, and consists of a few delicate and rather disturbed rays. The dorsal commences very slightly in front of the anal, and consists of fourteen or fifteen rays; the anal fin with only ten. All fins delicate, with slender distantly articulated rays; minute fulcra are observable on the anterior margins of at least the dorsal and anal; the caudal body prolongation is very slender and attenuated. The scales are more strongly marked than in R. monensis, their ornament consisting of from four to six sharply defined straight ridges passing from before backwards, with a slight obliquity over nearly the whole exposed surface, and terminating in denticulations of the posterior margin; in some cases feeble traces of the delicate vertical grooves seen in R. monensis are visible just in front of the commencement of the longitudinal ridges”. (Traquair Reference Traquair1874–1878, p. 244)
A formal diagnosis has not been previously presented.
Locality. Found in coal shale tips throughout the Scottish Lower Coal Measures, although remains are more frequently found in localities at younger stratigraphic levels.
Material. Pieces of slate containing the disarticulated dermal head bones in situ (GLAHM 152852); the endoskeletal remains of a juvenile specimen showing the parasphenoid (GLAHM 152834); and a well-preserved maxilla with associated preoperculum and palatoquadrate still attached. All specimens from Wester Bracco. Also examined was a large amount of isolated material, including skull roofs and other dermal elements, from other shale tips from the Scottish Lower Coal Measures.
Description. A tentative reconstruction of the complete skull of Rhadinichthys grossarti is shown in Figure 13 (left lateral and dorsal views), and a dorsal aspect of disarticulated head bones in situ is shown in Figure 14.

Figure 13 Rhadinichthys grossarti, tentative reconstruction of the dermal head bones: (a) left lateral view; (b) dorsal view. Heavy dotted lines indicate path of sensory canals; lighter dotted lines indicate possible arrangement of some smaller elements. Abbreviations: ang = angular; br = branchiostegal plates; cl = cleithrum; clv = clavicle; den = dentary; dhy = dermohyal; dpt = dermopterotic; dsph = dermosphenotic; esc = extrascapular; fr = frontal; ju = jugal; lac = lacrymal; mdc = mandibular canal; mesc = median extrascapular; mx = maxilla; na = nasal; op = operculum; pa = parietal; pcl = postcleithrum; pmx = premaxilla; pop = preoperculum; popc = preopercular canal; pt = posttemporal; ptc = posttemporal sensory canal; ro = rostrum; scl = supracleithrum; sclc = supracleithrum sensory canal; slp = sclerotic plates; soc = supraorbital canal; sop = suboperculum. Scale bar = 2 mm.

Figure 14 Rhadinichthys grossarti, GLAHM 152852: (a) dorsal aspect of disarticulated specimen in situ, showing dorsoventrally compressed dermal head bones; (b) interpretive drawing of specimen. Abbreviations: den = dentary; fr = frontal; mx = maxilla; pa = parietal; pq = palatoquadrate; scl = supracleithra. Scale bar = 2 mm.
Skull roof. The general form of the skull roof (Fig. 2c) is an isosceles triangle; the frontals are approximately quadrilateral, more than twice as long as broad; the parietals diminutive and irregular in form, about one-fifth the size of the frontals; the dermopterotics are rectangular, almost twice as large as the parietals and extending as far as the transverse suture margin; the transverse suture weakly sinuous; the anterior, middle and posterior pit-lines are not observed; the anterior margin of skull roof dentate for the articulation of the rostrum and lateral notches for articulation with the posterior limbs of the nasals; the lateral margins of the frontals are slightly concave with a thin rim along the whole length; the posterior margin of skull roof is lobed; the supraorbital canal extends from the anterior margin of the frontals and terminates at the posterior margin of these bones in the form of a rather strong sinusoid curve; the dermopterotic sensory canals are only discernible at the posterolateral margins of the bones; a pineal foramen is absent. Ornamentation of the skull roof ranges from sparse short tubercles to long posteriorly directed spines, the spines may be so prolific in some specimens that other dermal features are obscure or completely hidden; the medial area of the frontals in some specimens may lack ornamentation, in others they are sculptured with posteriorly directed spines; in specimens which lack ornamentation of the frontals there are two to several small, paired depressions located along the length of the bone and close to the path of the supraorbital canal.
Snout. The snout is comprised of a single median rostral bone, paired nasals and dentigerous premaxillae. The rostrum is stubby and keyhole-shaped with a wide, rounded anterior region occupying about half of its length, ethmoidal commissure not evident; the posterior margin tapered for contact with the frontals; ornamentation of large, irregular tubercles covering most of the rostral region; lateral margins with deep restrictions between the anterior and posterior regions for the nasal openings. The nasals are stubby with a short, narrow, tapered limb posteriorly, the anterior region has an almost straight terminal edge which connects with the premaxilla; the medial margin is near straight but angled for attachment to the lateral margin of the rostral and there is a deeply notched area for the nasal opening; the lateral margin is gently curved and forms the anterosuperior margin of the orbit; the supraorbital canal not visible. The premaxillae are paired rectangular elements half as high as wide and each armed with small teeth along inferior margin.
Palatoquadrate. Figure 15 shows the lateral surface of the palatoquadrate of Rhadinichthys grossarti with associated maxilla and preoperculum (GLAHM 152865). The palatoquadrate is long, slender and almost the same depth along its complete length, rounded anteriorly and planate posteriorly with a small, deep posteroventral extension; sharp conical teeth, similar to those found on the maxilla but slightly smaller, are located along the ventral margin of the bone. As in R. monensis, the different elements of the palatoquadrate are not discernible as the whole entity is fused as a single unit. The articulation for the endocranial basipterygoid process is in the form of a large notch situated about midpoint of the dorsal margin. The parasphenoid GLAHM 152834 (Fig. 16) of Rhadinichthys grossarti is quite unlike that of actinopterygians such as Woodichthys, Lawrenciella, Leuderia and Cosmoptychius. It is similar in form to that of Rhadinichthys monensis, including the minute denticles which cover the medial area of the ventral surface. However, the ascending process is shorter and the basipterygoid process is also hidden slightly behind the ascending process when viewed dorsally as in that species. The buccohypophyseal canal is also clearly visible as a tiny aperture situated between the arms of the ascending processes of the parasphenoid.

Figure 15 Rhadinichthys grossarti, GLAHM 152865, lateral view of maxilla with associated preoperculum and palatoquadrate attached. Abbreviations: bp = basal process; max = maxilla; pop = preoperculum; pop.c = preopercular canal; pq = palatoquadrate. Scale bar = 2 mm.

Figure 16 Rhadinichthys grossarti, GLAHM 152834: (a) dorsal view of parasphenoid from a juvenile specimen; (b) interpretive drawing of specimen. Abbreviations: asc.pr = ascending process of parasphenoid; bhc = buccohypophyseal canal; bpt = basipterygoid process. Scale bar = 1 mm.
Cheek, orbital and lower jaw regions. The maxilla has a short and slightly up-curved suborbital ramus, which is about the same length as the expanded posterior plate; the posterior plate is somewhat rectangular, planar dorsally with a posteroventral elongation; the surface is smooth without ornamentation or there may be small tubercles along the suborbital ramus and with a few, minute, posteriorly directed spines on the posterior plate; the mesial surface is furnished with a horizontal furrow close to the inferior margin and traverses the complete length of the bone; the dentary margin of the maxilla is almost straight and armed with small conical teeth of two sizes as far back as the posteroventral elongation. Fortunately, dermal elements of individual specimens of R. grossarti are frequently encountered en masse and very occasionally an odd sclerotic ossicle can be found among the bones; these bones are small, curved, unornamented and of varying thickness; thus, by inference there are probably four in number, with wider bones being located dorsally, as in R. monensis. The lacrymal is thin and ventral to the orbit; the jugal is more or less a crescentic bone with the dorsal margin planar and located at the posteroinferior part of the orbit; the dermosphenotic (posterodorsal infraorbital) does not connect with the limb of the nasal, it is near tear-shaped, slightly curved and tapered anteriorly, the broad posterior margin is rounded for connection with the dorsal margin of the jugal (posterior infraorbital) and the complete length of the mesial margin in contact with the dermopterotic and the frontal; the infraorbital canal is not visible on any of the infraorbital bones. Suborbitals have not been identified so far. The preoperculum is often found with the maxilla still attached in a natural position; this bone has two limbs, an anterior limb and a posterior limb which form an obtuse angle between, and which agree with the angle of the posterosuperior margin of the maxilla; the anterior limb is slightly longer and much wider anterolaterally than the slim posterior one; the preopercular canal runs along the complete length of the medial region of the bone. A quadratojugal has not been observed and hyomandibulars do not seem to be preserved. The dentary is slender, deeper posteriorly and tapering sharply anteriorly; there is row of elongate marks along the ventral rim running in an anteroposterior direction; mesial to the elongate marks the mandibular sensory line which is clearly visible; the posterior region of the dentary is somewhat extended at the angular and the surangular is well represented as a rectangular extension to the posterosuperior margin; the teeth of the dentary are identical to those of the maxilla; that is, small conical teeth of two sizes and extending the length of the dentary margin, but stopping short of the posterosuperior margin of the surangular. Ornamentation on both surfaces of dentary consists of a few long, coarse, occasionally broken ridges running anteroposteriorly in a horizontal direction with a slight upward inclination.
Operculogular. The operculum lies at a very oblique angle posterior to the preoperculum, more-or-less trapezoid in form with a tapered anterosuperior angle, dorsal margin rectilinear, ventral and posterior margins convex; ornamentation consists of a few low ridges following the contours of the posterior and ventral margins and a number of round markings which follow closely the length of the anterior margin. The suboperculum is approximately half the size of the operculum, fan-shaped with the dorsal margin more curved than the inferior margin; ornamentation as on operculum, with low ridges following the contours of the bone along the posterior and ventral areas. The exact number of branchiostegals is not known, but they are stubby with a tapered anterior region and convex posterior one; ornamentation seems to be of long, course ridges running almost the complete length of the bone in an anteroposterior direction. It is not known whether or not gulars exist.
Extrascapulars and posttemporals. Paired lateral extrascapulars have not been identified so far; however, due to a faint shape having often been observed in a number of skull roofs in that area, a median extrascapular most probably exists. The single paired posttemporals are large, rounded-triangular elements, joined mid-line and spanning the posterior margin of the skull roof; the anterior margin is rectilinear, with the anterolateral corner incorporating the posttemporal sensory canal; ornamentation of long, coarse ridges originating from close to the anterior margin and terminating before entering the posterior region or terminating as spine-like processes along the margin.
Pectoral girdle. The supracleithrum is a large bone which tapers dorsally, forming a triangular protuberance which articulates with the mesial surface of the posttemporal; the anterior and posterior margins are near parallel and the inferior margin convex; the path of the supracleithrum sensory canal is clearly seen to traverse from the dorsal protuberance to close to the mid posterior margin; the ornamentation is of low ridges following the contours of the anterior and inferior margins, with long spines covering most of the bone and directed towards the posterior and posteroinferior margins. The cleithrum is large, with an unusual shape in being much restricted in the middle with a triangular upper portion and an inflated, triangular lower portion; the middle constricted region has a large, deep embayment for insertion of the pectoral fin and the anteroinferior margin has a small recess for the attachment of the clavicle; a strong vertical ridge runs medially from the anterosuperior tip to the end of the triangular lower portion; a ventral pit-line is absent; ornamentation consists of low ridges following the contour of the dorsal margin and posteriorly directed spines covering the surface of the lower portion. The clavicle is a lanceolate bone without the tall dorsal spur found in other rhadinichthyids of the Scottish Lower Coal Measures such as R. monensis and R. glabrolepis; it is also without ornamentation. A postcleithrum has not yet been identified, although it is very probable one exists.
Squamation. Ornamentation of the scales as described by Traquair with additional notes that some flank scales (Fig. 10c) are a little higher than wide; the articulation peg is very strong for such small scales relative to the size of the fish; and that the ridges are long-cuspidate as they pass well beyond the posterior margin of the scale. There are also about six large, round-cordate ridge scales anterior to the dorsal fin present on the type specimen, which were not mentioned by previous authors.
Length of fish is 55–70 mm.
Rhadinichthys? ornatocephalum sp. nov.
Holotype. GLAHM 152853, shale slab containing disarticulated specimen.
Diagnosis. Skull roof sculptured with an intricate pattern of irregular-shaped ridges and tubercles; posteroventral elongation of the maxilla deeply rounded; anterior margin of the skull roof with a median cleft for articulation with the rostrum; ornamentation of the anterior region of frontals of a strong chevron pattern directed posteriorly; scales with five or six broad, flat ridges commencing about midscale and terminating as sharp posterior denticulations.
Remarks. As with Rhadinichthys glabrolepis mentioned earlier, the result of the following observations place this taxon in the Rhadinichthyidae: (i) a reconstruction of the dermal head bones show the snout to be prominent, though not so much as that species; (ii) paired antorbitals may exist as neither they nor the premaxillaries have been identified; (iii) the dermosphenotics are more or less T-shaped; (iv) a pineal foramen is not recorded; (v) clavicles and interclavicles have also not yet been identified, though by inference, they would most likely connect along the midline. Moreover, owing to the form and disposition of the operculae and cheek bones and the arrangement of the bones of the skull roof which follow a similar pattern to that of Rhadinichthys monensis, this taxon has also been given the generic title of Rhadinichthys? Fossils of this little fish are immediately recognised by the skull roof sculpture and by the long spines along the margins of some of the dermal bones.
Locality. Found in all localities where Drumgray shales are exposed.
Type locality. Wester Bracco, North Lanarkshire [8264 6570].
Material. Piece of laminated shale from Wester Bracco shale tip (GLAHM 152853) containing the associated remains of a disarticulated specimen showing most of the head dermal bones, including scales but devoid of any fin material. In addition, the study material includes a large amount of isolated dermal elements and a dozen or so complete or near-complete skull roofs.
Etymology. The epithet refers to the individual skull dermal bones, which are ornamented with an elaborate sculpture of contorted, raised ridges and tubercles.
Description. A tentative reconstruction of the complete skull of Rhadinichthys? ornatocephalum is shown in Figure 17 (left lateral and dorsal views), and the dorsal aspect of disarticulated head bones in situ is shown in Figure 18.

Figure 17 Rhadinichthys? ornatocephalum sp. nov., tentative reconstruction of the dermal head bones: (a) left lateral view; (b) dorsal view. Heavy dotted lines indicate path of sensory canals; lighter dotted lines indicate possible arrangement of some smaller elements. Abbreviations: ang = angular; ant = antorbital; br = branchiostegal plates; cl = cleithrum; clv = clavicle; den = dentary; dhy = dermohyal; dpt = dermopterotic; dsph = dermosphenotic; esc = extrascapular; fr = frontal; ju = jugal; lac = lacrymal; mdc = mandibular canal; mesc = median extrascapular; mpl = median pit-line; mx = maxilla; na = nasal; op = operculum; pa = parietal; pcl = postcleithrum; pmx = premaxilla; pop = preoperculum; popc = preopercular canal; pt = posttemporal; ptc = posttemporal sensory canal; ro = rostrum; scl = supracleithrum; sclc = supracleithrum sensory canal; soc = supraorbital canal; sop = suboperculum. Scale bar = 2 mm.

Figure 18 Rhadinichthys? ornatocephalum sp. nov., holotype GLAHM 152853: (a) dorsal aspect of disarticulated specimen in situ, showing dorsoventrally compressed dermal head bones; (b) interpretive drawing of specimen. Abbreviations: den = dentary; dpt = dermopterotic; dsp = dermosphenotic; fr = frontal; ju = jugal; mx = maxilla; pa = parietal; pop = preoperculum. Scale bars = 2 mm.
Skull roof. The skull roof (Fig. 2d) is the form of an isosceles triangle; the frontals approximately quadrilateral, twice as long as broad; the parietals trapezoidal, about one-third the size of the frontals; the dermopterotics extend from the base of the parietals to almost half the length of the frontals approximating the form of a circular segment; the transverse suture is strongly sinuous; the middle pit-line is clearly observed at the mid parietal region; the anterior margin of the skull roof with a median cleft for articulation with the rostrum; the lateral margin of frontals slightly concave; the posterior margin of the skull roof is lobed; the supraorbital canal extends as a rather conspicuous, strong sinusoid curve from anterior margin of frontals and terminating at mid parietal region; the dermopterotic sensory canals are clearly visible along the lateral margins of these bones; a pineal foramen is absent. The ornamentation of the skull roof is an intricate pattern of irregular ridges and tubercles running mostly in an anteroposterior direction; the ornamentation of the anterior region of the frontals is of a strong chevron pattern directed posteriorly; the median area of the frontals with less sculpture.
Snout. The snout, as usual, comprises the single median rostral with a single nasal bone attached laterally. It is not known whether paired antorbitals or a dentigerous premaxillae are included, as neither has been identified so far. The rostrum is rounded anteriorly, with a tapered posterior margin for contact with the anterior margin of the frontals; the median portion is rectangular and ornamented with large irregular tubercles aligned more or less anteroposteriorly; the anterior region is smooth and unornamented and the lateral margin has deep constrictions between the two regions for the nasal openings. The nasals are broad, with a short limb posteriorly; their anterior areas are broad with an almost straight terminal edge, which may have connected with an antorbital or premaxillary; the medial margin has a deeply notched area for the nasal opening and is slightly curved for attachment to the lateral margin of the rostral; the lateral margin of the nasal isgently curved to form the anterodorsal margin of the orbit; the supraorbital canal is not visible along this bone. The rostral region is unlikely to protrude very far, as a result of the rostrum and nasals being short, stout bones.
Cheek, orbital and lower jaw regions. The maxilla has a long, narrow, upward curved suborbital ramus and expanded posterior plate; the posterior plate is more or less rectangular, with a deeply rounded posteroventral elongation; the suborbital ramus slightly longer than the posterior plate; ornamentation is of elongate tubercles along the dentary margin of the ramus, with several well-spaced ridges running along the dorsal and anterior region of the posterior plate, with a few of the ridges continuing to the margin of the posteroventral elongation, occasionally bifurcated; the mesial surface of maxilla has a curved horizontal furrow along most of its length; the dentary margin is strongly curved and armed with small, tightly packed teeth as far back as the posteroventral elongation. The sclerotic bones, which would most likely be similar to those of other species found in these coal measures, have not yet been identified. The infraorbital bones are not particulary well preserved in this fish; however, the lacrymal is the usual thin bone ventral to the orbit, with the infraorbital canal running along its whole length; the jugal is large and crescentic, located at the posteroventral part of the orbit. The infraorbital canal is not observed on this bone; however, three or four well-spaced ridges run along its length; the dermosphenotic (posterodorsal infraorbital) is the usual crescentic shape, tapering anteriorly; however, as it does not articulate with the posterior margin of the nasal, a small supraorbital bone may lie between the two bones. The broad posterior margin of the dermosphenotic is recessed for connection with the dorsal margin of the jugal (posterior infraorbital) and the complete length of the mesial margin is in contact with the dermosphenotic and the frontals; however, the sensory canal cannot be seen to cross the bone. Suborbitals have not been identified so far, and again, considering the outline and shape of other cheek bones, they possibly do not exist. The preoperculum has two limbs, an anterior inflated limb and a smaller posterior limb, with each limb forming an obtuse angle of about 135° between and which agree with the angle of the dorsal margin of the maxilla; the preopercular canal extends along the complete length of the dorsal margin of the bone. A quadratojugal has not been observed and hyomandibulars do not seem to be preserved. The dentary is a thickset bone, deep posteriorly and tapering along its axis with a rounded anterior margin; there is a reticulated ornamentation along a broad ventral rim running in an anteroposterior direction; the mandibular sensory line is clearly visible ventrally and mesial to the reticulated sculpture; the angular which makes up the posterior margin is preserved and the surangular is well represented as a triangular or rectangular extension to the posterior margin; the teeth of the dentary are identical to those of the maxilla, that is, armed with small, tightly packed teeth extending the length of the dentary margin but stopping short of the dorsal margin of the surangular; the ornamentation of both inner and outer surface of the dentary is of long ridges or elongate tubercles running in a more or less horizontal direction from the posteroventral corner.
Operculogular. The operculum seems to lie at an oblique angle posterior to the preoperculum, approximately rectangular and rounded dorsally; the ornamentation is unclear but seems to be of a few low ridges following the contours of the bone along most of the posterior and ventral areas. The suboperculum is a little less than half the size of the operculum, more or less rectangular, with an upward curved posterosuperior angle; the ornamentation is as the operculum with low ridges following the contours of the bone along the posterior margin and with a few scattered tubercles close to anterior margin. The branchiostegals are slightly stout, tapered at both ends and ornamented with several wide ridges which traverse almost the complete length of the bone and terminate along the posterior margin as sharp denticulations. The exact number of branchiostegals or gulars is not known.
Extrascapulars and posttemporals. Paired lateral extrascapulars have not been identified so far, although traces of a median extrascapular are visible on some specimens. The single paired posttemporals are similar in form to those of R. glabrolepis, appearing as large, triangular elements, joined midline and spanning the posterior margin of the skull roof; the anterior margins are curved anteromesially; and the anterolateral corner incorporates the posttemporal sensory canal; the ornamentation is of flat ridges following the contour of the lateral, mesial and posterior margins and terminating as sharp denticulations along the posterior margin.
Pectoral girdle. The supracleithrum is a large, elongate bone tapering sharply from about the upper two-thirds of the bone, with the path of the supracleithrum sensory canal clearly seen to traverse the dorsal protuberance; the ornamentation is of low ridges following the contours of the anterior margin, with a few rather feeble ones along the posterior region; there are about five very sharp spines directed posteriorly along the posterior margin. The cleithrum is the usual inverted S-shaped bone, with an expanded posterodorsal region and a large, expanded ventral region; a strong vertical ridge extends medially from the anterodorsal tip to the expanded ventral area; a ventral pit-line is absent from this bone; the ornamentation may consist of a few very fine ridges following the inferior margin of the ventral plate, or a few faint lines along the anterior and posterosuperior angle, or it may be absent altogether. The clavicle and a postcleithrum have not yet been identified.
Squamation. The scales (Fig. 10d) are large for the size of the fish, almost square or slightly rhomboid, with some flank scales a little higher than wide and with a strong peg and socket articulation; the anterior margin of the scales has four or five closely-set striae, which normally follow the contour of the ventral region and may have a straight or sinusoid margin; the remainder of the scale has five or six broad, flat ridges commencing about midscale and terminating as sharp denticulations in the posterior half of the scale, or along the posterior margin. The strong fin fulcra of this species are frequently encountered among the scales.
Approximate length of fish is about 40 mm.
Rhadinichthys? plumosum sp. nov.
Holotype. GLAHM 152845, slab of shale containing disarticulated specimen.
Paratype. GLAHM 152846, slab of shale showing maxilla and frontal bone.
Diagnosis. Ornamentation of skull dermal bones of large, round or near-round and elongate tubercles; frontals almost three times as long as broad; dentary and maxilla armed with strong conical laniaries with smaller teeth between the larger ones; scales have a feather-like sculpture of fine, closely set, oblique ridges.
Type locality. Specimens were collected from light-coloured silty, carbonaceous shale at Wester Bracco coal waste tip [NS 8264 6570] in North Lanarkshire, the only known locality.
Remarks. Up until now, there has been insufficient material suitable for an analysis of characters which would help determine the familial level of Rhadinichthys? plumosum. Nonetheless, an appreciable amount of dermal material belonging to this taxon has been found to be similar in morphological detail as that found in at least two of the Rhadinichthyids from the Scottish Lower Coal Measures, Rhadinichthys monensis and Rhadinichthys ornatocephalum: the path and form of the supraorbital canals on the frontals; the pattern of ornamentation of the tubercles along the maxilla; and the arrangement of conical teeth with smaller teeth between the larger ones are characters which are easily recognised as those of Rhadinichthys monensis. Furthermore, the anterior margin of the skull roof of Rhadinichthys? plumosum exhibits a similar median recess for articulation with the rostrum to that of Rhadinichthys? ornatocephalum; this also may suggest that posterior margin of the rostrum is also similar. For those reasons, Rhadinichthys? plumosum has been tentively given the generic title of Rhadinichthys? until a more complete study can be undertaken.
Rhadinichthys? plumosum is one the larger rhadinichthyids and is usually found in close association with smaller actinopterygians such as Pseudogonatodus aurulentum. Isolated remains of Rhadinichthys? plumosum are often found among a mass of scales and bones of Pseudogonatodus. The disposition and condition of this material, however, does not indicate that it was originally gut content. Nevertheless, the large voracious teeth of Rhadinichthys? plumosum do suggest it may have been the main predator of these smaller fish.
Material. Material consists of two pieces of finely laminated shale containing part and counterpart of a disarticulated specimen in situ (GLAHM 152845), a small piece of shale containing the dentary and frontal (GLAHM 152846) and a large amount of isolated scales, all collected from Wester Bracco shale tip.
Etymology. The species name refers to the feather-like sculpture of the body scales. The fine ridges which cross obliquely across the surface of the scale resemble part of the vein with its close arrangement of barbs and rachis of a plume or semi-plume.
Description. A reconstruction of the complete head of Rhadinichthys? plumosum is not possible, due to the lack of many of the key dermal elements of the skull; however, a dorsal aspect of a disarticulated specimen in situ is shown in Figure 19.

Figure 19 Rhadinichthy? plumosum sp. nov., holotype GLAHM 152845: (a) aspect of specimen in situ, showing disarticulated dermal head bones; (b) interpretive drawing of specimen. Abbreviations: cra = cranium; den = dentary; mx = maxilla; op = operculum; pmx = premaxilla; pop = preoperculum; sop = suboperculum. Scale bars = 2 mm.
Skull roof. The only bone of the skull roof found in good condition so far is the right frontal (specimen GLAHM 152846), which is approximately quadrilateral, almost three times as long as broad and slightly wider across the posterior region; the transverse suture is similar to, but not as strongly sinuous, as in Rhadinichthys monensis and Rhadinichthys? ornatocephalum; the anterior margin of skull roof is similar to that of Rhadinichthys? ornatocephalum in the form of the median recess which articulates with the rostrum; the lateral margins of frontals are slightly concave; supraorbital canal comparable to that of Rhadinichthys monensis, extending the full length of the frontal and close to the lateral margin as an almost straight sinusoidal curve, though closer to the margin than Rhadinichthys monensis; a pineal foramen is absent; ornamentation of the frontals is of large, round or near-round tubercles, the tubercles are flat over most of the surface of the frontals, except along the lateral margins where they are more pronounced and the area proximal to the mid-sagittal margin where it is devoid of ornamentation.
Snout. A median rostral bone and paired nasals have not been identified so far, and due to the fact that the shape of the anterior margin of the frontals is very similar to that of Rhadinichthys? ornatocephalum, it can be assumed that the posterior margin of the rostrum of Rhadinichthys? plumosum also has a similar shaped posterior margin. Furthermore, a small bone which is partly hidden by the large cranial plate could be interpreted as a premaxilla, as this bone appears to show what seem to be two or three small conical teeth. However, as the bone is only partly exposed in the shale, a proper identification is not possible.
Cheek, orbital and lower jaw regions. The maxilla has a long, thick and straight suborbital ramus; the expanded posterior plate is more or less in the form of an isosceles trapezoid and is planar dorsally, with deep posteroventral elongation; the ornamentation of the maxilla is similar to that of Rhadinichthys monensis, with large, oblique tubercles located along the dentary margin of the suborbital ramus. At the expanded plate, the tubercles are round or near-round and across the posterior region of the expanded plate the ornamentation is of fine, near straight ridges crossing in an oblique direction to the posteroventral extension. The dentary margin of maxilla is armed with strong, regularly-spaced conical laniaries, with several smaller teeth occupying the space between the laniaries. Infraorbital bones have not been identified so far. The preoperculum is the shape of an inverted ‘V', with the two limbs forming an obtuse angle – as in other species this angle roughly corresponds with the angle of the posterosuperior margin of the maxilla; the anterior limb is twice as long as the posterior one, with the preopercular canal traversing the complete length of the dorsal margin of the bone. The dentary is a thickset bone, deep posteriorly and tapering anteriorly; the mandibular sensory line is clearly visible and a row of circular indentations is located along the ventral margin; the angular is only partially preserved at the posterior margin; the teeth of the dentary are identical to those of the maxilla, that is, regularly-spaced conical laniaries with several smaller teeth occupying the space between the laniaries; the ornamentation of the dentary is of fine ridges running in a horizontal direction, commencing close to the posterior margin and changing to small, round or near-rounded tubercles as the ornamentation approaches the anterior margin, the mesial surface is almost devoid of any form of sculpture except for a horizontal furrow close to the ventral margin along most of length.
Operculo-gular apparatus. The operculae are too poorly preserved for a precise description. However, the suboperculum of Rhadinichthys? plumosum does appear to be of similar form to that of Rhadinichthys monensis and both the operculum and suboperculum exhibit a similar characteristic ornamentation of low ridges, which appear to follow the contours of the bone.
Squamation. The scales (Fig. 10e) are rectangular, a little higher than wide with a rounded inferior margin; the dorsal margin is sigmoid with a tall and strong articulation peg; the anterior covered margin is wide and without vertical striae; fine, closely set, oblique ridges traverse the scale surface, occasionally bifurcating and terminating at the posterior margin as fine denticulations.
Approximate length of fish is about 65 mm or larger.
Family Gonatodidae Long, Reference Long1988
Genus Pseudogonatodus Gardiner, Reference Gardiner1967
Diagnosis (after Gardiner Reference Gardiner1967). Body fusiform; dorsal fin placed rather far back, partly in advance of, partly opposing the anal but giving appearance of being almost opposite it. Both fins of moderate size and triangular. Paired fins large, with stout rays, pelvics placed nearer to anal than to pectorals; caudal fin deeply cleft and inequilobate. All fins with prominent fulcra anteriorly and rays closely articulated and distally bifurcated. Skull with almost vertical suspensorium and rostrum not pronounced. Opercular less than one and three-quarter times as deep as subopercular. Suborbital and dermohyal series present. Branchiostegal rays numerous, skull roofing bones and cheek bones ornamented with coarse, stout ridges of enamel; teeth of variable size but in single series. Scales rhomboid, large and ornamented with fine striae and distinct punctations.
Type species. Gonatodus parvidens Traquair, 1882 (in Traquair Reference Traquair1877–1914).
Pseudogonatodus parvidens Gardiner, Reference Gardiner1967
Diagnosis (emended). Fishes not exceeding 22 cm in total length; length of head one fifth the body length and maximum depth of body about one quarter the body length. Suspensorium nearly vertical, head short and with bluntly rounded snout. Maxilla of distinctive shape, high posteriorly. Single large suborbital present. Teeth on jaws very small, arranged in single closely-set row. Scales large in proportion to body size, deeper than broad on the flank, feebly ornamented with few striae and exhibiting coarse punctations. Posterior borders of scales delicately serrated.
Pseudogonatodus aurulentum sp. nov.
Holotype. GLAHM 152849, skull roof.
Paratype. GLAHM 152850, disarticulated specimen.
Diagnosis. A small fish in which the form of the skull roof shows a marked morphological similarity to that of Canobius ramsayi Traquair, Reference Traquair1881, also incorporating other characteristics relating to genera in the Canobius–Gonatodus–Mesopoma complex (Traquair Reference Traquair1914; Moy-Thomas & Bradley Dyne Reference Moy-Thomas and Dyne1938). This similarity includes the pattern of the dermal skull bones of Pseudogonatodus aurulentum, which have a striking resemblance to those of Canobius elegantulus, and even more so with Pseudogonatodus parvidens. This resemblance also includes the short unprotruding rostral region, which has resulted in a rather familiar short, rounded head of this group of fishes. Other characteristic features worth considering are the high posterior maxillary plate; the form of the dentary bone, which is short, thickset, deep posteriorly and tapering sharply anteriorly; the slightly tilted preoperculum, which suggests an almost vertical suspensorium; and the large posterior spines of the supracleithrum. The large scales are also characteristic of the gonatodids, albeit the sculpture in P. aurulentum is of strong ridges and not pectinated as in P. parvidens. This disparity may be explained by resorption of the external scale surface (Gardiner Reference Gardiner1967), where the ornamentation of the scales in the posterior region is the first to undergo resorption and is smooth with distinct pectinations, whilst the anterior part of the fish shows scales with a complex ornamentation of ridges and striae.
Remarks. Although dermal bones and scales are occasionally found as isolated components, they are more usually encountered en masse in a 2–3 mm-thick fragment of laminated slate as fossilised material close to, but not in, the anthracitic shale from the Drumgray coal.
Locality. Found in all localities where Drumgray shales are to be met.
Type locality. Wester Bracco, North Lanarkshire. [NS 8264 6570].
Material. Fragments of slate collected from the above shale tips containing a large amount of isolated dermal elements [GLAHM 152150], including several dozen skull roofs of which GLAHM 152849 has been erected as the holotype.
Etymology. Fossilised material of this small fish is immediately recognised by its delicate gold tint; hence the specific title ‘aurulentum'.
Description. A tentative reconstruction of a complete skull of Pseudogonatodus aurulentum is shown in Figure 20 (left lateral and dorsal views).

Figure 20 Pseudogonatodus aurulentum sp. nov., tentative reconstruction of the dermal head bones: (a) left lateral view; (b) dorsal view. Heavy dotted lines indicate path of sensory canals; lighter dotted lines indicate possible arrangement of some smaller elements. Abbreviations: ang = angular; ant = antorbital; br = branchiostegal plates; cl = cleithrum; clv = clavicle; den = dentary; dhy = dermohyal; dpt = dermopterotic; dsph = dermosphenotic; esc = extrascapular; ethc = ethmoid commissure; fr = frontal; hpl = horizontal pit-line; ioc = infraorbital canal; ju = jugal; lac = lacrymal; mdc = mandibular canal; mesc = median extrascapular; mx = maxilla; na = nasal; op = operculum; pa = parietal; pcl = postcleithrum; pop = preoperculum; popc = preopercular canal; pt = posttemporal; ptc = posttemporal sensory canal; ro = rostrum; scl = supracleithrum; sclc = supracleithrum sensory canal; soc = supraorbital canal; sop = suboperculum. Scale bar = 2 mm.
Skull roof. The skull roof (Fig. 2e) is in the form of an isosceles triangle; the frontals are approximately triangular, half again as long as broad; the parietals are square and about one-third the size of the frontals; the dermopterotics are quadrilateral and roughly equal in size as the parietals; the transverse suture is W-shaped, though near horizontal along the anterior margin of parietals; the anterior margin of the skull roof is scalpriform (chisel-shaped) for articulation with the rostral and laterally notched for articulation with the posterior limbs of the nasals; the posterior margin of skull roof is slightly indented; the supraorbital canal extends as a sinuous curve from the anterior margin of the frontals and terminates at the anterolateral corners of the parietal; the dermopterotic sensory canals are clearly discernible along the lateral margins of the bones; pit-lines have not been observed and a pineal foramen is absent. The ornamentation of skull roof is of broad, flat, relatively long tubercles arranged in a strong regular pattern, which is directed mediolaterally across the frontals, parietals and dermopterotics.
Snout. The median rostral is a rather stout bone when compared with previously described species; it is also keyhole-shaped and with a rounded anterior region which occupies about one-third of its length; the ethmoid commissure is seen to traverse the anterodorsal region; the remaining portion of the bone is stout, widest at the mid-lateral margin; the posterior margin is straight, with two small posterolateral projections for contact with the chisel-shaped frontal anterior margins; the lateral margins of the rostrum show deep restrictions between the two regions for the nasal openings; the surface of the bone is smooth and without ornamentation. The nasals are broad, with a short posterior limb which articulates with the dermosphenotic; the anterior margin is broad with an irregular terminal edge which connects with the antorbital; the medial margin is curved for attachment to the lateral margin of the rostral and there is a deeply notched area for the nasal opening; the lateral margin of the nasal is gently curved and forms the anterodorsal margin of the orbit, with a notch opposite the nasal embayment; the supraorbital canal is visible along the whole length of the bone, of which the surface is smooth except for the ornamentation of irregular tubercles close to the path of the supraorbital canal; and as a result of the short rostral region a non-protruding rounded head is produced. The antorbitals are approximately polygonal in form, with the infraorbital canal running from the inferior margin and linking with the ethmoid commissure dorsally.
Cheek, orbital and lower jaw regions. The maxilla has a relatively short, straight suborbital ramus approximately equal in length to the posterior plate: the posterior plate is high and much expanded with an angled posterior margin; the ventral margin of the maxilla is straight along the whole length and edentulous; the ornamentation is of four or five slightly oblique, long, flat, lustrous tubercles running in a dorsoventral direction on the posterior plate and with short tubercles along the ramus. Four small, thin, curved and unornamented bones of varying thickness constitute the sclerotic ossicles, the thicker bones being most likely located dorsally. The lacrymal is a thin, tapered bone ventral to the orbit, with the infraorbital canal running along its whole length; the jugal is a large, long and thin crescentic bone located at the posteroventral part of the orbit, with the infraorbital canal running dorsoventral along its length; the dermosphenotic (posterodorsal infraorbital) is a thin crescentic bone tapering anteriorly to articulate with the narrow posterior margin of the nasal, the broad posterior margin connects with the dorsal margin of the jugal (posterior infraorbital) and the complete length of the mesial margin in contact with the dermopterotic and the frontal; the dermosphenotic sensory canal intersects posteriorly, passes across the bone, emerges at the medial margin, where it probably enters the lateral margin of the dermopterotic. A suborbital has not been identified and, considering the outline, shape and positions of the jugal, preoperculum and maxilla, and unless it is present as a very thin wedge of bone, it is unlikely one exists. The preoperculum is more-or-less sickle-shaped, with the two limbs of about equal size, the preopercular canal runs along the complete length of the dorsal margin of the bone and occasionally a horizontal pit-line can be seen; and due to the fact that the preoperculum is only slightly tilted, it may suggest an almost vertical suspensorium. A quadratojugal has not been observed. Dermohyals do not seem to be preserved. The dentary is a short, thickset bone, deep posteriorly and tapering sharply anteriorly; there is row of five or six spines along the ventral margin and there is no evidence of teeth; the mandibular sensory line is clearly visible and separated from the ventral margin by a wide rim; the angular is not usually preserved; the surangular is clearly seen as a rectangular extension to the posterior margin; both inner and outer surfaces of the dentary are smooth, lack ornamentation, and have no visible sutures.
Operculo-gular apparatus. The operculum lies at a slight angle posterior to the preoperculum and is approximately sub-ovate; it is bluntly pointed dorsally with a rounded posterior margin and planar ventral margin; the ornamentation is of a few broken lustrous ridges anterodorsally and low ridges following the contours of the posterior and inferior margins. The suboperculum is less than half the size of the operculum and trapezoid in form; the posterosuperior angle is drawn upwards forming a strong curve along its dorsal margin with the inferior margin sigmoid; ornamentation of the suboperculum consists of low ridges posteriorly, with a few irregular lustrous tubercles close to the anterior margin. The branchiostegals are slender, tapered anteriorly and more-or-less rounded posteriorly; ornamentation is of a few low ridges following the posterior margin and lateral margin with or without small spines directed posteriorly; the exact number of branchiostegals is not known. Gulars have not been identified.
Extrascapulars and posttemporals. Possible paired and median extrascapulars have been identified. The single paired posttemporals are large, triangular bones, joined midline and spanning the posterior margin of the skull roof; the anterior margin is curved anteromesially or sigmoid; the mesial and lateral margins are drawn to almost a point posteriorly and the posttemporal sensory canal traverses the anterolateral region. The ornamentation of the posttemporals is of flat, lustrous tubercles, with posteriorly directed points over the whole surface and with spines along the posterior margin, as in R. grossarti.
Pectoral girdle. The supracleithrum is a large, stout bone, tapering from the inflated mid-region to the dorsal and ventral margins; the path of the supracleithrum sensory canal is clearly seen originating from the dorsal protuberance and terminating near the middle of the posterior margin; the anterior margin is convex; the posterosuperior margin is planar where articulation of the bone is below the posttemporal; the posterior margin has four or five large, posteriorly directed spines; the ornamentation is reduced to broad ridges only in the inflated region and follows the anterior and posterior margins. The cleithrum is the usual inverted S-shaped bone, with an expanded posterosuperior region and a squared ventral region; a very strong vertical ridge runs medially from the anterodorsal tip to the squared ventral area of the bone; a ventral pit-line is absent; the surface of the cleithrum is smooth and without ornamentation. The clavicle has not yet been identified; however, its form is most probably a lanceolate bone with or without a dorsal spur. A postcleithrum has not yet been identified, though it is very probable one exists.
Squamation. The scales (Fig. 10f) are large for the size of the fish, almost square or slightly rhomboidal with some flank scales a little higher than wide; the peg and socket articulation is well-developed; and the anterior margin has five or six well-spaced striae that occasionally turn into the ventral margin or may be straight or convex; the remainder of the scale has six or seven broad, flat ridges, commencing about midscale and terminating as broad denticulations at the posterior margin. The strong fin fulcra of this species are frequently encountered among the scales.
Approximate length of fish is about 36 mm.
Family undesignated
Pseudohaplolepis gen. nov.
Diagnosis. As for the type species.
Etymology. The lustrous nature of the dermal ornamentation and the clearly visible pineal foramen could easily be confused with that of a species of haplolepid.
Remarks. Remains of this little fish are usually found in association with haplolepid material, with which it may easily be confused; however, the general shape of the skull roof and the lateral position of the nasals separate Pseudohaplolepis from the Haplolepidae. Furthermore, the course of the supraorbital canals, which can be clearly marked on the skull roofs of other palaeoniscoid actinopterygians, is also observed in Pseudohaplolepis. Up until now, this feature has never been observed on any haplolepid skull roofs recovered from the Pennsylvanian of North Lanarkshire and is not normally observed in haplolepid skulls from other locations unless they have been exposed by weathering or have had the surface layer removed by scraping. However, specimens from the English locations of Longton and Newsham tend to display the suborbital canals quite clearly, although these fossils may have been subjected to some form of abrasive technique to expose these features.
It should be worth noting that although Pseudohaplolepis possesses scales which are more or less typically actinopterygian in form, an examination of Lanarkshire material shows that the dentary, gular and the arrangement of the middle and especially the posterior pit lines require forethought. Both the dentary and gular in Pseudohaplolepis differ to those of other known actinopterygians found in the Scottish Lower Coal Measures, but are very similar in form to those of immature Megalichthys hibberti, which is often found in association with these small fishes; and furthermore, posterior pit-lines are not usually observed in Lanarkshire material except in immature M. hibberti and in some haplolepids; they are, however, located close to the posterior margin of the parietals in Pseudohaplolepis (compare Figures 2f and 27a).
Pseudohaplolepis argentatum sp. nov.
Holotype. GLAHM 152854, slab of shale containing disarticulated specimen.
Paratype. GLAHM 152855, skull roof.
Diagnosis. Ganoine surface of dermal bones intensely lustrous (more so than the associated and also lustrous haplolepid material); skull roof strongly tapered at frontals, lobed at the parietals and ornamented with posteriorly directed spines at the dermopterotics; pineal foramen strongly evident and located slightly posterior of midpoint of frontals; supraorbital canal clearly observed as an almost straight line; nasals rhomboidal in form, stubby and broad with a short, narrow posterior limb; jugal bone and cleithra adorned with long, sharp spines; preoperculum a crescentic bone with concave anterior margin and produced spur; dentary with inflated anterior portion with a row of minute teeth; outer margin of opercular bones with row of sharp spines; two large lateral gulars present; middle pit-line and posterior pit-lines pronounced; scales very small and usually much taller than wide.
Locality. Found in close proximity to, and within, the anthracitic shales of the Drumgray coal at Dewshill [NS 8531 6385] and Wester Bracco [NS 8264 6570].
Etymology. The specific name refers to the lustrous nature of the surface of the dermal bones, which resemble the metallic element silver.
Material. Fragment of slate collected from the Wester Bracco shale tip with the disarticulated part remains of a specimen in situ (GLAHM 152854), showing the skull roof, scales and several other dermal elements. Other material consists of pieces of a disarticulated specimen (GLAHM 152856) and a number of isolated dermal elements and scales which includes about 30 complete skull roofs, of which GLAHM 152855 has been erected as the paratype.
Description. A reconstruction of the dorsal aspect of the skull of Pseudohaplolepis argentatum is shown in Figure 21, with images of various other dermal bones, including a complete skull roof. A lateral aspect reconstruction is not possible, due to the lack of principal cheek dermal elements.

Figure 21 Pseudohaplolepis argentatum gen. nov. (a) Reconstruction of the dermal head bones in dorsal view. Heavy dotted lines indicate path of sensory canals. (b) Dermal elements: (i) skull roof, paratype GLAHM 152855; (ii) supracleithra; (iii) jugal; (iv) preoperculum; (v) operculum; (vi) dentary; (vii) left gular. Abbreviations: cl = cleithrum; dpt = dermopterotic; dsph = dermosphenotic; esc = extrascapular; fr = frontal; ju = jugal; mesc = median extrascapular; mpl = median pit-line; na = nasal; op = operculum; pa = parietal; pf = pineal foramen; ppl = posterior pit-line; pt = posttemporal; ro = rostrum; scl = supracleithrum; slp = sclerotic plates; soc = supraorbital canal. Scale bars = 2 mm (a); 1 mm (b).
Skull roof. The skull roof (Fig. 2f) is in the form of an isosceles triangle; the frontals are triangular, twice as long as broad; the parietals rectangular, slightly smaller than the frontals in length; the dermopteroptics approximate an isosceles trapezoid with the posterior margin spiked and the anterior margin extending only as far as transverse suture; the transverse suture is almost straight; the middle pit-line is pronounced, originating from lateral margin of parietal and terminating about mid bone; the posterior pit-line is also much pronounced, commencing close to ventrolateral margin at an angle of 45° and directed towards the mid-sagittal suture; the anterior margin of the skull roof strongly tapered and terminates as a minute median recess for articulation with the rostral; the lateral margins of the frontals are more or less planar, with the bones tapering towards the anterior margin; the posterior margin of the skull roof is gently lobed at the parietals and ornamented with posteriorly directed spines at the dermopterotics; the supraorbital canal can be observed as an almost straight line originating from about one-third the distance from the anterior margin of the frontal and terminating just short of the transverse suture, or extending passed the suture and briefly into the parietals; as a result of the very strong sculpture of the dermopterotics the sensory canals cannot be ascertained; the pineal foramen is clearly visible and located slightly posterior to midpoint of frontals. The surface of the skull roof is always highly lustrous, with the frontals being smooth and without sculpture; and the posterior areas are ornamented with flat rugae, which at times are shaped into long sharp spines and directed towards the posterior margin.
Snout. The rostral is without ornamentation, relatively small and slim with a rounded anterior region, with the remaining portion tapering to a point posteriorly. The nasals are rhomboidal in form, stubby and broad with a short, narrow limb posteriorly which articulates with the limb of the dermosphenotic; anteriorly, the terminal margin is almost straight for connection with either an antorbital or premaxilla; the medial margin almost straight for attachment to the lateral margin of the rostral and with a long notched area for the nasal opening; the lateral margin gently concave and forms the anterosuperior margin of the orbit; the supraorbital canal is not visible. Antorbitals or premaxillae have not been identified. Due to the small rostral and the nasals being small and stubby, the snout is unlikely to be pronounced but slightly rounded.
Cheek, orbital and lower jaw regions. Despite the fact that other skull dermal elements of this small fish are frequently found, the maxilla has yet been identified. The sclerotic ossicles also have not yet been identified; they would most likely be similar to those of other species found in these coal measures. The lacrymal is a large, slightly bent bone ventral to the orbit, with its width thickening anteriorly and there is a thin groove following the curved mesial margin, which may indicate the path of the infraorbital canal; the jugal is the usual large crescentic bone located at the posteroventral part of the orbit; however, the lateral margin is armed with about ten sharp spines located along its length, which follow the contour of the orbit and are directed in a posteroventral direction; the dermosphenotic (posterodorsal infraorbital) is long, curved and tapers anteriorly to abut with the narrow posterior margin of the nasal, with the length of the mesial margin in contact with the dermopterotic and more than half the length of the frontals. Suborbitals have not been identified so far. The preoperculum is a more-or-less crescentic bone with an expanded anterior region; the margin of the anterior region is concave; the curved inferior margin terminates with a produced spur anteriorly; the preopercular canal has not been detected. A quadratojugal has not been observed and hyomandibulars do not seem to be preserved. The dentary is a large odd-shaped bone with an inflated anterior portion. The anterior portion is twice as wide as the posterior part and has a row of minute teeth along the dorsal margin, commencing from the anterior tip to about half the length of the bone; the posterior margin is truncate, with a single posteroventral spine; a surangular is well-exposed in many specimens and an angular or prearticular is present; ornamentation is of lustrous rugae with occasional small indentations over the surface.
Operculogular. The operculae are very similar in size, form and ornamentation; both are semi-hemispherical, with sharp spines situated along outer curved margin. Nevertheless, the operculum and suboperculum may be separated by noting the orientation of the spines and ornamentation, and also by noting the shape of the anterior margins of the bones, as they are slightly different. The operculum has six to eight marginal spines and has a small anterosuperior section of the bone absent, possibly the site for the location of a hyomandibular, the suboperculum has eight to twelve spines; the ornamentation of both bones consists of lustrous rugae which often form small indentations over the complete surface. There are two large, spathulate lateral gulars with a V-shaped pit-line mid area of the bone; ornamentation is similar to that of the opercular bones.
Extrascapulars and posttemporals. Possible paired and median extrascapulars have been identified; however, the highly lustrous nature of the ornamentation of the skull roof makes it difficult to determine the actual extent of the anterior margin of the posttemporals. Nevertheless, the single paired bones are large, more or less triangular elements, joined midline and spanning the posterior margin of the skull roof; the ornamentation is as that of the skull roof, consisting of a smooth lustrous ganoine rugae.
Pectoral girdle. Up until now, the only element of the pectoral girdle to be identified is the cleithrum, which is a very slim and inverted S-shaped bone of which the superior region is narrow, with three posteriorly directed sharp spines and the inferior region spoon-shaped with the embayment for the pectoral fin shallow; a strong vertical ridge runs medially from the anterodorsal tip of the superior region to the mid-posterior margin of the spoon-shaped ventral area, which does not seem to have a ventral pit-line; the ornamentation of the usual smooth lustrous ganoine rugae is characteristic of this species.
Squamation. The scales (Fig. 10g) are small for the size of the fish, rectangular and up to three times taller than wide on some scales; the dorsal articulation peg is tall and thin, the ventral articulation has not been observed; the anterior margin has three or four fine striae, some of which follow the margin round to the ventral border; fine irregular striae traverse the surface of the scale at an oblique angle and terminate at the posterior margin as fine denticulations.
Approximate length of fish is 25 mm.
Family undesignated
Lanarkichthys gen. nov.
Diagnosis. As for the type species.
Etymology. The genus is named from the region of the Midland Valley of Scotland, where the remains of this species are found and which, up until now, is the only known location.
Remarks. These infrequently encountered remains are found only in close association with haplolepid material. More often than not, the dermal bones of individual Lanarkichthys are found almost exclusively within a small surface area of a fragment of shale, but distributed vertically in the laminated matrix to a depth of two or three millimetres. This is possibly the result of the dead fish falling through the water and being suspended for a period of time in brackish silt which has not quite settled. As the fish disintegrates, the various bones eventually settle, the depth of which may depend on the mass of the bone relative to the density of the brackish composite at the different depths. Furthermore, the body scales of Lanarkichthys resemble closely those of extant species of Diodontidae such as Chilomycterus sp. The long, pointed spines on the body scales of Chilomycterus, which are often employed as a defensive mechanism, may also have been used for the same purpose in Lanarkichthys; the spines may also have possessed similar properties to those of the spines and skin of the extant species. Furthermore, located directly above the suborbital ramus of the maxilla in Lanarkichthys is a long-elliptical area with numerous, evenly-spaced minute protuberances, which could be interpreted as denticles or possibly some form of sensory papillae. These denticles or papillae appear somewhat similar in form and arrangement to those found in the extant family of Perciformes, the Gobiidae, and the arrangement of sensory papillae of these fishes is very important for their classification (Hoese Reference Hoese1983). However, although the denticles or sensory papillae in Lanarkichthys have a similar configuration to that of the Gobiidae, they are bony structures and not the soft-tissue structures which develop from cells on the skin and which occasionally are erectile, as in the urogenital papilla between the anus and the base of the anal fin in Neogobius melanostomus.
Lanarkichthys gardineri sp. nov.
Holotype. GLAHM 152847, disarticulated specimen.
Paratype. GLAHM 152848, skull roof.
Diagnosis. The ornamentation of the skull roof is a lustrous sculpture of small, posteriorly directed spines fairly evenly distributed over the complete surface; the rostral bone is broad across the anterior margin and adorned with three large spines along the lateral margins; the maxilla with short, deep and more or less straight suborbital ramus with a long-elliptical area adjacent to the dorsal margin which displays minute protuberances; the cleithrum is slender, an inverted comma-shaped bone with a large, triangular ventral region; the clavicle is stout, triangular with a short dorsal spur and one or two posteriorly directed ventral spurs; each scale is equipped with a long, sharp spine close to the inferior margin; the length of the spine varies, but may be equal to the height of the scale; some flank scales are fused in groups of three, one scale on top of the other.
Type locality. Specimens were collected from light-coloured silty, carbonaceous shale at Wester Bracco, North Lanarkshire coal waste tip [NS 8264 6570], the only known locality.
Material. Small fragments of shale containing an isolated skull roof (GLAHM 152848), part skull roof and other isolated dermal elements (GLAHM 152847), all collected from the above shale tip.
Etymology. The epithet of this fish is named after the palaeontologist Brian G. Gardiner, in recognition for his work on basal Actinopterygii.
Description. A reconstruction of the complete skull of Lanarkichthys gardineri is not possible. due to the lack of key dermal elements of the skull. Figure 22 shows a near complete skull roof and other dermal elements; the high posterior plate of the maxilla, the long and slender cleithrum and supracleithrum and the form of the deep scales suggest a deep-bodied fish.

Figure 22 Lanarkichthys gardineri gen. nov., holotype GLAHM 152847, dermal elements: (i) skull roof with rostral attached; (ii) cleithrum; (iii) area of denticles above maxilla; (iv) clavicle; (v) conjoined scales; (vi) maxilla with preoperculum attached; (vii) denticle area enlarged. Scale bars = 1.0 mm.
Skull roof. A dorsal view of the skull roof of Lanarkichthys gardineri (reproduced from GLAHM 152847 and GLAHM 152848) is shown in Figure 2g. The form is of an isosceles triangle with the frontals triangular and roughly twice as long as broad, the lateral margins of which are more or less straight; posterior margin of skull roof concave and somewhat lobed. The paths of the transverse and parietal-dermopterotic sutures are unclear, making it difficult to determine the extent of these bones. However, the parietals seem to be more or less square and about half the size of the frontals; the middle pit-lines are clearly observed in the paratype, traversing the mid parietal regions; the dermopteroptics are roughly ovoid in shape and armed with posteriorly directed spines, a characteristic shown only in the holotype. The anterior margin of skull roof has a deep “V-” or “U-shaped” median recess for articulation with the rostral. The supraorbital canals are observed as near straight, parallel paths originating close to the transverse suture and extending in an anterior direction to just over half the length of the frontals. A pineal foramen is not observed. The ornamentation is lustrous and consists of minute, posteriorly directed spines covering the complete surface of the skull roof.
Snout. Only the median rostral bone is available and is keyhole shaped. It is widest across the anterior margin and adorned with three large spines along the lateral margins of the anterior part of the bone and occupies about half its length. The remainder of the bone is posteriorly obtuse for articulation with the recess at the anterior margin of skull roof. Dentigerous premaxillae have not been observed. Ornamentation is as that of the skull roof.
Cheek, orbital and lower jaw regions. The maxilla has a short, deep and more-or-less straight suborbital ramus; the ramus is roughly the same length as the expanded posterior plate. The expanded plate has a rounded posterosuperior margin, with the posteroventral elongation slightly drawn and rounded. The preoperculum is often still attached to the posterosuperior margin of the maxilla. The ornamentation is highly lustrous and consists of small posteriorly directed spines which adhere to a general pattern which follows the borders of the expanded plate. The dentary margin of the maxilla is edentulous. No infraorbital bones have been found so far; however, there is a long-elliptical area in the region of the jugal (posterior infraorbital) which is furnished with many small denticles. Moreover, on close examination of the suborbital ramus of the maxilla, these small denticles are concentrated within the area which appears to be a fenestration of the dorsal margin of this bone. Suborbitals have not been identified so far. The preoperculum is a deep bone anteriorly, short and curving in agreement with the dorsal margin of the maxilla, with a sharp taper towards the posterior margin. The ornamentation is as that of the maxilla, except that the pattern of the posteriorly directed spines is more haphazard and only partly follows the curvature of the bone. A dentary has not yet been identified.
Operculogular. Several unidentified plates have been found which, in all probability, are elements of the operculo-gular apparatus; the ornamentation is similar to that on the skull roof, but more lustrous.
Extrascapulars and posttemporals. Extrascapulars and posttemporals have not been identified so far.
Pectoral girdle. The supracleithrum is difficult to describe, due to a large bone (probably a branchiostegal plate) lying on top; however, it seems to be a large element which tapers from about mid-bone to the dorsal and ventral margins. The supracleithrum sensory canal is just visible, its path can be followed from the dorsal protuberance to the posterior margin; two spines also ornate the posterior margin, one is short and the other is very long, longer than the large spines on the scales; the ornamentation of the bone is as that of the skull roof and covers the posterior two-thirds, the remainder being devoid of ornamentation. The cleithrum is a slender and inverted comma-shaped bone with a large, triangular ventral region; the anterior margin of this region is sigmoid in form for articulation with the posterior margin of the clavicle; a shallow embayment for the pectoral fin is evident along the posterior margin and two or three strong vertical ridges run medially from the anterodorsal tip to the commencement of the ventral area; a ventral pit-line is absent; ornamentation is as that of the supracleithrum and covers the complete area posterior to the vertical ridges. The clavicle is a stout triangular bone with a short dorsal spur and one or two posteriorly-directed ventral spurs; ornamentation is as that of the rest of the pectoral girdle.
Squamation. The typical body scales (Fig. 10h) are ovoid, taller than wide, with a strong articulation peg at the dorsal margin. A sharp spine originates from about the midpoint of the scale and close to the inferior margin; the length of the spine varies, but may be equal to the height of the scale. The covered surface of the scale may vary from about one-quarter to half the area, with the exposed part of covered in a lustrous ornamentation of minute posteriorly directed spines. Some of the flank scales are fused together in groups of three, one scale on top of the other, with each scale exhibiting a backwards-directed spine.
Approximate length of fish is 30 mm.
Family Haplolepidae Westoll, 1944
Subfamily Parahaplolepinae Lowney, Reference Lowney1983
Genus Blairolepis Lowney, Reference Lowney1983
Diagnosis (after Lowney Reference Lowney1983). A haplolepid with the Parahaplolepis type skull roof; i.e., with posterolateral lappets and embayed parietals, but having separate dermopterotics and parietals.
Blairolepis wallacei sp. nov.

Figure 23 (a) Holotype of Blairolepis wallacei sp. nov., skull roof, GLAHM 152862. (b) Line drawing of same skull roof showing ornamentation. Reconstructed from paratypes GLAHM 152863 and GLAHM 152864. Scale bar = 2.0 mm.
Holotype. GLAHM 152862. Incomplete skull roof with anterior portion of frontals missing, but showing the left posterolateral lappet, embayed parietals and separate dermopterotics and parietals.
Paratype. GLAHM 152863. Incomplete skull roof showing most of frontals and parietals; dermopteroptics and posterolateral lappets missing.
Paratype. GLAHM 152864. Incomplete skull roof with anterior part of frontals and posterolateral lappets missing; position of pineal foramen indicated by weakly ossified area near middle of frontals;
Material. The above three incomplete specimens.
Type locality. Carbonaceous shale from the Drumgray coal waste tip at Wester Bracco, North Lanarkshire [NS 8264 6570].
Etymology. The species is named in honour of Alfred Russel Wallace (1823–1913), the British naturalist, explorer and geographer, best known for proposing a formulation of evolution due to natural selection.
Diagnosis. Skull roof with posterolateral lappets and embayed parietals, but having separate dermopterotics and parietals. Can be distinguished immediately by the peculiar ornamentation over most of the skull roof, the separate parietals and dermopterotics with posterolateral lappets, and the diminutive size of the parietals relative to the frontals.
Discussion. Skull roof (Fig. 27a) is in the form of an isosceles triangle; dermopterotics and parietals separate; posterolateral lappets present; anterior margin of frontals rounded, tapering at the shoulders; lateral margin with a marked concavity; posterior margin embayed; transverse suture with oscillating path; anterior pit-line does not cross into frontal; pineal foramen faint; skull roof ornamented with round tubercles anteriorly tending to irregular shaped tubercles posteriorly. Skull roof slender, outline similar to that of Parahaplolepis canadensis Baird (Reference Baird1978) and Parahaplolepis anglica Traquair (Reference Traquair1894 in Traquair Reference Traquair1877–1914), except that the shoulders of the anterior margin are tapered, whereas in those species they are rounded. In Blairolepis loanheadensis Lowney (Reference Lowney1983) the anterior margin is also tapered; however, this species can be distinguished by the longitudinal rugae on the frontals. Frontals of Blairolepis wallacei are more-or-less trapezoid in form, their length being more than twice their width with the lateral margins distinctly concave. Parietals are small and square-shaped, each of the sides fitting three and a half times into the length of the frontals. Dermopterotics are in the shape of an irregular pentagon, and one and a half times the size of the parietals. Posterolateral lappets are present, but are usually poorly preserved or missing altogether. Pineal foramen is extremely faint, located slightly behind the midpoint of the frontals. Mid-sagittal suture is present in some specimens, absent from others; the transverse suture is clearly seen to pass along the parietals at right angles from the mid-sagittal suture to about half its length, loop over the anterior pit-line then directed forward and outward in an undulating path separating the frontal from the dermopterotic. The anterior pit-line does not cross from the parietal to the frontal, as found in Parahaplolepis Westoll (1944), and both the middle and posterior pit-lines are clearly visible and in their customary positions. Sculpture of the skull roof is typical to that of most Parahaplolepis species and consists of large, round tubercles anteriorly, but tending to irregular shaped tubercles posteriorly as far as the posterolateral lappets, of which the margins are not usually found in good condition; ornamentation persists over most of the skull roof, except the median frontal area which is comparatively smooth along most of its length.
Approximate total length of fish is about 80 mm.
Genus Parahaplolepis Westoll, 1944
Diagnosis. A haplolepid in which the dermopterotics and parietals are not separate; the anterior pit line extends into the frontal from the parietal; posterior margin of skull roof deeply embayed for the reception of the extrascapulars and posttemporals, the latter two pairs of bones meeting in midline; skull roof ornament of broad raised rugae or low tubercles; maxilla (in the one species where it is known) with tall posterior expansion and a quadratojugal present; ventral lateral line present.
Parahaplolepis poppaea sp. nov.

Figure 24 (a) Holotype of Parahaplolepis poppaea sp. nov., skull roof, GLAHM 152859. (b) Line drawing of same skull roof showing ornamentation. Leading edge of left lappet reconstructed from paratype GLAHM 152860. Scale bar = 0.25 mm.
Holotype. GLAHM 152859. Near complete skull roof, with leading edge of left lappet damaged.
Paratype. GLAHM 152860. Near complete skull roof.
Locality and horizon. Found amongst the compressed fossilised stems of Calamites suckowii and other vegetation in a light-coloured carbonaceous shale from the Drumgray coal, Wester Bracco, North Lanarkshire [NS 8264 6570].
Etymology. The species is named after my granddaughter, Poppy Sophia Elliott, born in the same year this species was found.
Material. Two near complete skull roofs (GLAHM 152859 and GLAHM 152860), and one incomplete specimen.
Diagnosis. Dermopterotics and parietals not separate; posterolateral lappets essentially present, but with large distal expansion; anterior pit-line crosses well into frontal; pineal foramen very strongly marked and located close to anterior margin of frontals; ornamentation consists of long lateral rugae and small irregular tubercles on frontals and broad, flat ridges on parietals.
Discussion. The skull roof (Fig. 27b) is in the form of a rather squat, isosceles triangle; the anterior margin, which is not usually well preserved in this diminutive species, appears to be weakly rounded; the posterior margin is slightly embayed to receive the extrascapulars and the posttemporals, which may or may not meet midline. The complete frontal area of the skull roof of Parahaplolepis poppaea is in the form of an isosceles trapezoid; however, as the mid-sagittal suture is not visible, the medial margin of the frontals cannot be established. Nonetheless, it may be deduced that each frontal is trapezoid in form, their length being approximately equal to their greatest width and with the lateral margins somewhat concave; ornamentation of the frontals is of long, occasionally bifurcating, rugae running in an anteroposterior direction almost the full length of the bones. The pineal foramen, which is very strongly marked, is positioned very close to the anterior margin of the frontals. The parietals have the form of an irregular pentagon and are similar in shape and direction of ornamentation to that found in Parahaplolepis elenae Elliott (Reference Elliott2014). However, in P. elenae the parietals are slightly smaller than the frontals and in Parahaplolepis poppaea they are approximately the same size as the combined areas of the two frontal bones. Ornamentation of the parietals is not unlike that found in Haplolepis corrugata (Newberry) Westoll (1944) from Linton and consists of wide, flat ridges bordering the anterior, posterior and chisel-shaped posterolateral margin in a concentric manner, with the remaining area ornamented with irregular shaped tubercles; the leading anterior edge of the lappets have an undulating outline, remarkably similar to that found in some Protohaplolepis Lowney (Reference Lowney1983). The position of the mid-sagittal suture is not apparent; however, the transverse suture is clearly seen to be sinusoidal in form. The anterior pit line can be seen to cross the transverse suture well into the area of the frontal, and a long middle pit line is also present; however, a posterior pit line has not been observed in any specimens.
Approximate total length of fish is about 38 mm.
Genus Andrewsolepis gen. nov.
Diagnosis. Skull roof of Parahaplolepis type; that is, the dermopteroptics and parietals are not separate. However, the anterior pit-line does not cross into the frontal as in Parahaplolepis, and the posterior pit-line is located in an unusual position; the posterior margin of skull roof is also deeply embayed for the reception of the extrascapulars and posttemporals, the latter two pairs of bones meeting in midline; the ornamentation of the skull roof is of strong, raised tubercules.
Type species. Andrewsolepis lochlani gen. nov.
Etymology. The genus is named in honour of the British palaeontologist Sheila Mahala Andrews (1939–1997), in recognition of her work on sarcopterygian and other fishes.
Andrewsolepis lochlani sp. nov.

Figure 25 (a) Holotype of Andrewsolepis lochlani gen. nov., skull roof, GLAHM 152858. (b) Line drawing of same skull roof showing ornamentation with minor reconstruction of posterolateral lappets. Scale bar = 2.0 mm.
Holotype. GLAHM 152858. Near complete skull roof and counterpart.
Locality and horizon. Carbonaceous shale from the Drumgray coal waste tip at Wester Bracco, North Lanarkshire [NS 8264 6570].
Etymology. The species is named after my grandson, Lochlan James Elliott, born in the same year the holotype of this species was found.
Material. Skull roof, GLAHM 152858, near complete part and incomplete counterpart. A second but incomplete skull roof was also used as a comparison.
Diagnosis. As for genus. Can be distinguished immediately by the Parahaplolepis-type skull roof, with the anterior pit-lines confined to the parietals.
Discussion. Skull roof (Fig. 27c) triangular in form; dermopterotics and parietals not separate; posterolateral lappets present; anterior margin of frontals tapered and pointed, lateral margins of frontals slightly concave; posterior margin of skull roof deeply embayed; transverse suture sinusoid; anterior pit-line does not cross into frontal; pineal foramen faint, located midpoint of frontals; skull roof ornamented with large, round or elongate tubercles. The skull roof of Andrewsolepis lochlani resembles that of Parahaplolepis cf. canadensis (Baird Reference Baird1978; Carpenter et al Reference Carpenter, Falcon-Lang, Benton and Grey2015, p. 679) from Joggins, Nova Scotia, particularly in the tapered anterior margin, the concave lateral margins of the frontals, the ornamentation of the frontals and the chisel-shaped posterolateral lappets. In Andrewsolepis lochlani, however, the anterior margin of the frontals tapers to a point and the ornamentation of the parietals are composed mainly of elongate tubercles, not round ones. In addition, the transverse suture is sinusoid in form, with its path directed at right angles to the mid-sagittal suture (if that suture were clearly marked) and not passing diagonally forward and outward from the mid-sagittal suture as in Parahaplolepis cf. canadensis. There is also a similarity to the skull roof of Parahaplolepis tuberculata (Newberry) Westoll (1944) from Linton, notably in the tapered anterior margin of the frontals and the tuberculate ornamentation over the complete skull roof. However, in P. tuberculata there is a small niche at the apex of the anterior margin of the frontals to receive the nasal bone; also, the form and direction of the transverse suture in P. tuberculata is much the same as that of Parahaplolepis cf. canadensis. The shape of the frontal area of Andrewsolepis lochlani is more-or-less quadrilateral, with the lateral margins having a slight concavity. As the mid-sagittal suture is not visible, the medial margin of the frontals cannot be firmly established; nonetheless, it may be deduced that each frontal is trapezoid in form, with the posterior margin being approximately two-thirds the length of the bone. The transverse suture is well marked and positioned almost axially, is sinusoid in form and curves around the anterior pit-line and terminates at the lateral margin of the bone. The parietals are pentagonal in form, slightly smaller than the frontals with their separation at the mid-sagittal suture clearly visible. The posterolateral lappets are exceptionally long and chisel-shaped. The anterior and middle pit-lines are well-defined and in the usual position, the posterior pit-line being also present but occupying a peculiar position very close to posterior margin of parietals, this pit-line is clearly seen to be encircled by a raised lip in the incomplete counterpart. The pineal foramen is present, but faint, and located mid-point of the frontals. The ornamentation of the skull roof is very similar to that of Parahaplolepis canadensis and Parahaplolepis tuberculate, consisting of strong, round, occasionally elongate tubercles on the frontals. On the parietals, the tubercles combine together to form long rugae aligned more or less axially. The region of the posterolateral lappets is devoid of any kind of ornamentation, except for a few scattered pores.
Approximate length of fish is about 80 mm.
Genus Braccohaplolepis gen. nov.
Diagnosis. A haplolepid with paired skull fenestrations confined to parietals; dermopterotics not present; parietals meeting midline; posterior margin slightly embayed; ornament of skull bones of rugae arranged in concentric patterns remarkably similar to that of Pyritocephalus Fritsch (Reference Fritsch1893).
Type species. Braccohaplolepis fenestratum gen. nov.
Etymology. The genus is named after the Wester Bracco [NS 8264 6570] locality, where several new haplolepid species have been found.
Braccohaplolepis fenestratum sp. nov.

Figure 26 (a) Holotype of Braccohaplolepis fenestratum gen. nov., skull roof, GLAHM 152857. (b) Line drawing of same skull roof showing ornamentation. Scale bar = 2.0 mm.

Figure 27 Diagrams of centre part of skull roof showing pineal foramen, pit-lines and oblique/transverse sutures. Dotted line indicates usual path of the mid-sagittal suture in (a), (b) and (c) and probable extent of fenestrations in (d): (a) Blairolepis wallacei sp. nov.; (b) Parahaplolepis poppaea sp. nov.; (c) Andrewsolepis lochlani gen. nov.; (d) Braccohaplolepis fenestratum gen. nov. Not to scale.
Holotype. GLAHM 152857, skull roof.
Locality and horizon. Found as isolated specimens in or close to the haplolepid yeilding anthracite beds in carbonaceous shale from the Drumgray coal waste tip, Wester Bracco, North Lanarkshire [NS 8264 6570].
Etymology. The species is named from the skull roof fenestrations.
Material. Ten complete skull roofs plus one or two counterparts, of which GLAHM 152857 is erected as the type specimen. No other material is available, as the skull roofs have always been found as isolated specimens.
Diagnosis. Skull roof type (Fig. 26) with paired fenestrations confined to parietals; dermopterotics not present; parietals meeting midline; ornament of skull bones of rugae arranged in intricate concentric patterns. Can be distinguished immediately by the form of the fenestrations on the parietals.
Discussion. Skull roof (Fig. 27d) approximates an isosceles triangle; anterior margin of skull roof tapered; lateral margin convex; posterior margin slightly embayed; frontals approximately rectangular and about the same area as the parietals; transverse suture oblique, curving gently outwards and upwards from the mid-sagittal suture; pineal foramen absent; orbital fenestrae approximately semi-circular in form and confined to the parietals where they occupy almost one-third of the parietal area; skull ornamentation a complex pattern of rugae or tubercles. The general outline of Braccohaplolepis fenestratum is similar to that of Haplolepis corrugata from Linton in that the dermopterotics are absent in both taxa, with only the parietals present. However, the lateral margins of the frontals of Braccohaplolepis fenestratum are decidedly concave, as opposed to convex in Haplolepis corrugata. The general shape of the frontal-parietal margins of Braccohaplolepis fenestratum is quite unhaplolepid and more akin to the form of curvature found in the rhadinichthyids. Furthermore, the curved transverse suture of Braccohaplolepis fenestratum is quite unique, as no other haplolepid from the Lower Coal Measures has this feature which is curved from the mid-sagittal line to the base of the lateral margin of the frontals. It is also the only known haplolepid from the Lower Coal Measures in which the pineal foramen is absent; an observation based on about ten well-preserved skull roofs. The pineal foramen is also absent in the genera Microhaplolepis Lowney (Reference Lowney1980) from Linton; however, in this case the dermopterotics and parietals are separate, whereas in Braccohaplolepis fenestratum the dermopterotics are absent. Compared with Parahaplolepis, which also lacks the dermopterotics, Braccohaplolepis fenestratum does not possess the embayed posterior margin and posterolateral lappets which are an important diagnostic feature of the genera. Nonetheless, the ornamentation of the skull roof is remarkably similar to that of Pyritocephalus, with elongate tubercles running more or less concentrically around each margin of the frontals and parietals with the tubercles occasionally breaking into smaller units, but remaining almost complete down the lateral margins of the frontals and along the anterior margin of the skull roof. On the other hand, the orbital fenestrae in Braccohaplolepis fenestratum occupy much less of the area of the parietals than those in Pyritocephalus and where it falls far short of the frontals; whereas in all known Pyritocephalus genera the fenestrae extend into the posterior margin of the frontals.
Approximate length of fish is 80 mm.
Millerolepis eleionomae Elliott, Reference Elliott2014
Holotype. GLAHM 152372, skull roof.
Paratype. GLAHM 152861, skull roof.
Remarks: A second, though incomplete, skull roof of Millerolepis eleionomae (GLAHM 152861) has been procured and, due to the overlying ornamentation on this specimen being less pronounced than that of the holotype, additional detail of the skull roof morphology can be determined.
Diagnosis (emended). Diagnosis for genus and species. A haplolepid with skull roof in the form of an isosceles triangle; parietals without dermopterotics; parietals extending outwards and backwards; posterolateral lappets absent; anterior margin of frontals tapered, lateral margin planar, posterior margin deeply embayed; extrascapulars and posttemporals meeting at midline: transverse suture near straight and oblique; anterior pit-lines present on parietals and do not cross into frontals; middle pit-lines present and posterior pit-lines absent; pineal foramen strongly marked, positioned close to anterior margin; longitudinal rugae on frontals; skull roof ornamented with rugae and large tubercles.
5. Dermal head bones, morphology, reconstruction and implications
When fossil fish are examined to reconstruct the head bones, it can often be extremely difficult separating and identifying the different dermal elements. This difficulty is evident if specimens have been compressed laterally, as these bones are often positioned one on top of the other and crushed or mangled beyond recognition. This is not usually the case with disarticulated or isolated specimens, since the shape or form of the individual bones, including fine morphological detail, may easily be ascertained. The fact is that dermal bones such as those of the skull roof, and to a lesser extent the maxillae, are the most recognisable remains of different taxa; for this paper these bones are selected as the key elements in the reconstruction of the dermal head bones of specimens. Moreover, the reconstruction of the dermal head bones of several fishes from disarticulated material is not only a demanding undertaking, it is also intrinsically problematical in its modus operandi. Nonetheless, recreating taxa, or at least the arrangement of the head bones and utilising the method discussed earlier in this study, is one means of illustrating how the animal may appear in life. And furthermore, the morphological differences recorded from the different bones of the skull of individual taxa are found to be of significant importance as an aid in the classification of each taxon studied. As mentioned earlier, the utilisation of skull roof morphology can be problematical, especially when it is not known whether a characteristic is the plesiomorphic or the apomorphic state. Besides this, as many characters are indeed synapomorphies, employing parsimony analysis using these morphological features can be of great benefit when combined with other applied methods in the classification of a taxonomic study at the specific, generic and possibly the familial level.
With respect to the use of dermal head bones as a classification technique, Gardiner (Reference Gardiner1967) used the characteristics of these bones to advocate that the primitive conditions of lower actinopterygians can be found in physical conditions such as the dermosphenotics articulating with the nasals, and that the dermopterotics is simply a fusion of the area primitively covered by the intertemporals and supratemporals. Conversely, in a later paper Gardiner & Schaeffer (Reference Gardiner and Schaeffer1989) used the arrangement of the bones of the naso-temporal region to classify the lower actinopterygian fishes according to the position of these bones; however, in this paper the structure of the skull roof bones was used simply to emphasise the physical differences in the dermal bone pattern of the skull. It is those physical differences in skull roof morphology which are used as an aid in the classification of a range of fauna, whether ancient or modern. In fact, Westoll uses this technique to great advantage in his classification of the Haplolepidae.
With regard to this present study there are also, however, implications associated with differences in the physical arrangement of certain dermal bones in actinopterygians. For instance, the angle at which the preoperculum lies across the cheek is known to reflect the angle at which the suspensorium rests relative to the dentary and the skull; an oblique angle is suggested by some authorities (Moy-Thomas & Bradley Dyne Reference Moy-Thomas and Dyne1938; Westoll 1944; Schaeffer & Rosen Reference Schaeffer and Rosen1961; Gardiner & Schaeffer Reference Gardiner and Schaeffer1989; Gardiner Reference Gardiner, Schaeffer and Masserie2005) to be a primitive character of lower actinopterygians in which this angle can be very strongly inclined or lie in a near vertical position. In other words, the evolutionary trend is for a forwardly directed suspensorium. This variation in suspensorium angle is clearly exhibited by several of the fishes described in this study and, depending on the taxon, the preoperculum may or may not possess an expanded anterior arm which is tilted forwards to some degree, or the bone may be reduced to a simple crescentric form. For example, this study shows that Rhadinichthys monensis, Rhadinichthys glabrolepis sp. nov., Rhadinichthys grossarti, and possibly Rhadinichthys? plumosum sp. nov. all possess a suspensorium which is clearly tilted forward; Rhadinichthys? ornatocephalum sp. nov., however, possesses a preoperculum which is only slightly tilted and the suspensorium of Pseudogonatodus aurulentum sp. nov., lies almost vertical. Furthermore, the first four taxa possess a maxilla which has an expanded posterior plate and a long suborbital ramus, with the dentary margin of the maxilla and dentary armed with one or two rows of sharp teeth; Rhadinichthys? ornatocephalum sp. nov., however, possesses only minute teeth and the dentary of Pseudogonatodus aurulentum sp. nov., is short, thickset and edentate. These observations imply that the angle of the suspensorium and, consequently, the angle of the hyomandibular were modified by some groups of fish simply to compensate for a shortening of the jaw as feeding habits altered. Additionally, the rostral notch (first described by Poplin & Lund (Reference Poplin and Lund1995), who stated that this feature could possibly increase buccal cavity suction by entrapping food particles (and with the addition of extralateral gulars (Lund & Poplin Reference Lund and Poplin1997)) is probably characteristic of all rhadinichthyids, including a few other primitive actinopterygians. Nevertheless, with respect to the rhadinichthyids from the Scottish Pennsylvanian mentioned above (Rhadinichthys monensis, Rhadinichthys glabrolepis sp. nov. and most likely Rhadinichthys? ornatocephalum sp. nov.) these are taxa which are all known to exhibit a rostral notch and possibly possess extralateral gulars. Yet both Rhadinichthys monensis and Rhadinichthys glabrolepis sp. nov. posses large conical laniaries on the maxilla and dentary; a feature which usually indicates a predatory hunter. Nevertheless, if the above suggestions concerning a rostral notch are correct, then Rhadinichthys grossarti (possibly including Rhadinichthys? plumosum sp. nov. in this paper) should be reassigned at the generic level, as this taxon possesses a premaxillary, which implies the absence of a rostral notch. Also supporting this assessment is the form of the skull roof pattern of Rhadinichthys grossarti (Fig. 2c): the small parietals; the form of the dermopterotics; the disproportionate size of the scales relative to body size (compare to Fig. 10); and the lack of middle pit lines are all characters which are quite unlike those of other rhadinichthyids under study.
With reference to the new material from North Lanarkshire, especially the well-preserved skull dermal bones and the fine detail of the skull roofs, information obtained from the examination of these specimens was used not only in the reconstruction of the head bones to give an idea of the appearance of some of the fishes, but also in the support of a cladistic analysis of actinopterygian classification, at least in the generic and specific level.
There is no doubt that if, or when, more material is uncovered in the future, we may have a better understanding of these small fishes, nonetheless, there is certainly a strong urgency for an undertaking in the uncompromising task of a reappraisal of the classification of these ancient actinopterygians.
6. Cladistic analysis
Previous work (Elliott Reference Elliott2014) has encompassed cladistic analyses of associations which may have existed between the Haplolepidae and other early actinopterygians. A further analysis was conducted for the present paper, which was again based on the variation in skull roof morphologies to establish possible relationships between certain actinopterygian groups from British fossil locations. This analysis concentrated on genera which constituted a Canobius–Mesopoma–Rhadinichthys complex (see later comments), the Haplolepidae and new material collected from the North Lanarkshire coal shale tips. The information recovered was then used to determine what relationships, if any, there may be between the Haplolepidae and the Canobius-Mesopoma-Rhadinichthys complex and, in addition, to ascertain the relative taxonomic positions for the new taxa, especially that of Pseudohaplolepis argentatum gen. nov. and Braccohaplolepis fenestratum gen. nov.
In the same manner as the previous paper on the Haplolepidae, morphological data were obtained from detailed illustrations in Traquair (Reference Traquair1914), Moy-Thomas & Bradley Dyne (Reference Moy-Thomas and Dyne1938) and Gardiner & Schaeffer (Reference Gardiner and Schaeffer1989) and from personal material, including data from new specimens. A list of 35 taxa and 21 characters with a maximum of seven states was produced (Table 4) and, with the assumption that all characters are valid for all taxa, the information was used to construct a data matrix (Table 5). Parsimony analysis was conducted using PAUP4; all characters being unordered, given equal weight and optimized using ACCTRAN; the tree was set for SPR (subtree pruning and grafting) with polytomies set to hard. Tree values were then calculated by generating a maximum of 1000 most parsimonious trees.
Table 4 List of character and character states for Cladogram A

Table 5 Data matrix for the generation of cladogram A

A 50 % majority consensus tree was then produced, using 35 taxa from fossil localities in Scotland and England, including the data from the new taxa but excluding Rhadinichthys plumosum sp. nov. and Lanarkichthys gardineri gen.nov., due to insufficient skull roof morphological data. The tree was unrooted and the genus Canobius represented the outgroup on the grounds of the possibility of an ancestral lineage with the Haplolepidae (Westoll 1944). The confidence levels of the analysis were tested using bootstrapping and Bremer support. The bootstrapping method gave a greater than 70 % support for all nodes, with the lowest support given to two clades; 71 % to the Protohaplolepis clade which encompasses three taxa (P. traquairi, P. isabellae and P. limnades); and a 90 % support to the Pyritocephalus clade containing two taxa P. rudis and P. youngi. For clade confidence, Bremer support was designated a decay index of 6, as this was the number of steps required in order to collapse all nodes to a single polychotomy. The outcome of these methods does suggest that the confidence levels in the tree values were highly supported. Cladogram A (Fig. 28) illustrates the groups which constitute the paraphyletic Canobius–Mesopoma–Rhadinichthys complex and the Haplolepidae. The figure indicates that the Haplolepidae clade may in fact be paraphyletic and not monophyletic as previously assumed (Elliott Reference Elliott2014).

Figure 28 Cladogram A. Strict consensus based on skull roof morphologies, showing possible relationships of the new taxa from North Lanarkshire with the Canobius–Mesopoma–Rhadinichthys complex and the Haplolepidae. Tree length = 116; consistency index = 0.3879; retention index = 0.6787.
Classification within the aforementioned complex has always been problematical; however, the placement of the different genera shown as a result of the analysis does suggest discrete affinities within the families. For example, Canobius has an obvious close relationship with Mesopoma (Moy-Thomas & Bradley Dyne Reference Moy-Thomas and Dyne1938; Westoll 1944), not only in the almost vertical angle of the suspensorium, but also in the morphological detail of the skull roof; after all, several taxa were reassigned to Mesopoma which were previously placed in Canobius (e.g., C. pulchellus, C. politus, C. macrocephalus and C. crassus (Moy-Thomas & Bradley Dyne Reference Moy-Thomas and Dyne1938)), with C. macrocephalus originally placed with the rhadinichthyids. Morover, the cladogram also suggests that there is a close affinity with Gonatodus, Pseudogonatodus and the rhadinichthyids. This may be due to their physical resemblance, but may also include the similarities in the cheek and skull roof bones of these genera. And furthermore, concerning the association of Pseudohaplolepis argentatum gen. nov. with other groups, the cladogram does suggest that this genus is, in all probability, transitional, as it possesses features which are present in all of these groups. For instance, Pseudohaplolepis has several plesiomorphic characters. These include: the laterally placed nostrils; the separate parietal and dermopterotic, which is present in all early actinopterygians found in North Lanarkshire material except in some haplolepids; a clearly visible supraorbital canal which is not detectable in North Lanarkshire haplolepids; and a posterior pit-line which is located close to the posterior margin of the skull roof, which up until now has been known only in the Haplolepidae. With regard to apomorphies, the most important characteristic associated with Pseudohaplolepis is the pineal foramen, which is normally associated with the remains of the Haplolepidae or Sarcopterygii from Pennsylvanian material. There are, however, other pertinent features which may be included, such as the large lateral gulars; the tall scales; and, in particular, the highly lustrous ganoine ornamentation. The analysis also reinforces the association between the two subfamilies of Haplolepidae, the Haplolepinae and the Parahaplolepinae (Lowney Reference Lowney1983) which, owing to the lateral fenestrations on the skull roof of Braccohaplolepis, may imply that this taxon is a possible transitional taxon with the Pyritocephalus clade in the Parahaplolepinae. Additionally, both Blairolepis loanheadensis and Blairolepis wallacei sp. nov. exhibit separate dermopterotics and parietals, a feature found in the Haplolepinae and other actinopterygians. However, both taxa have been placed in the Parahaplolepinae, as they possess posterolateral lappets and embayed parietals on the skull roof, which suggests that the genus Blairolepis is perhaps transitional from the Parahaplolepinae to the Haplolepinae.
7. Conclusion
Ten new early actinopterygian taxa are described from material collected from five North Lanarkshire coal shale tips. The assemblage encompasses four new haplolepid taxa described from skull roofs (Blairolepis wallacei, Parahaplolepis poppaea, Braccohaplolepis fenestratum and Andrewsolepis lochlani) and includes six non-haplolepid taxa (Rhadinichthys ornatocephalum, Rhadinichthys glabrolepis, Pseudogonatodus aurulentum, Pseudohaplolepis argentatum, Rhadinichthys plumosum and Lanarkichthys gardineri), of which Pseudohaplolepis argentatum and Lanarkichthys gardineri require further study for the purpose of designation at family level. Concerning the Haplolepidae, this study has increased the number of species collected from North Lanarkshire from seven in a previous paper to eleven, the result of which extends the number of haplolepid fossil species in Europe to 20.
In addition to a descriptive analysis of the assemblage, information obtained from an anatomical study of the dermal bones of six non-haplolepid genera was used to reconstruct the dermal heads of three of these taxa. With regard to two named species, Rhadinichthys monensis Egerton and Rhadinichthys grossarti Traquair, an examination of the endocranial material, parasphenoid, palatoquadrate and its associated bones and the hyomandibular provided additional information for these genera; although a further comprehensive study is required for a more complete description.
Furthermore, a phylogenetic analysis for the complete assemblage was undertaken, excluding Rhadinichthys plumosum sp. nov. and Lanarkichthys gardineri gen. nov. due to insufficient skull roof data. The resulting cladogram indicates possible relationships within the Canobius–Mesopoma–Gonatodus–Pseudogonatodus–Rhadinichthys association to be a paraphyletic one, based on plesiomorphies shared with Canobius ramsayi and, in addition, the Canobius–Mesopoma association was found to be monophyletic. However, it was found that a few rhadinichthyid taxa are irregularly placed throughout the groups with regard to these relationships. The analysis also implies that the Haplolepidae are a paraphyletic group and not monophyletic as previously deduced. Nonetheless, two clades are in evidence within the haplolepid complex, one group representing the Haplolepinae and another group representing the Parahaplolepinae, thus strengthening a previous hypothesis. Also presented in the resulting cladogram is the position of Pseudohaplolepis argentatum within the groups, whereby this species exhibits a possible close relationship with the Gonatodus–Pseudogonatodus–Rhadinichthys association; this may be because P. argentatum possesses features which are present in fishes of these groups and may represent a transitional species, possibly indicating a parallel transformation from the rhadinichthyids to another group of fishes.
In the case of the new material from the Pennsylvanian of North Lanarkshire, especially the well-preserved scales, the skull dermal bones and the fine detail of the skull roofs, the information obtained from the examination of these specimens was used as an aid in the reconstruction of some of the fishes and also in the support of a cladistic analysis of actinopterygian classification, at least in the generic and specific level. There is no doubt that if, or when, more material is uncovered in the future, we may have a better understanding of these small fishes. Nonetheless, there is certainly a strong urgency for an undertaking of the uncompromising task of a reappraisal of the classification of these ancient actinopterygians.
8. Acknowledgements
I wish to thank Ramsey and William Kerr, owners of Wester Bracco Farm, for allowing me unlimited access to the shale tips. Special thanks are due to Stanislav Štamberg and Matt Friedman for their invaluable comments and advice which helped to improve this manuscript immensely.
Please note that this study is of a personal nature and is not funded in any way by SUERC.
9. Appendix 1. List of specimens consulted
The following list includes the museum catalogue numbers, specimens consulted with a brief description of the specimens, and if applicable, the designation of the specimens.



































