


Almost 75 years ago, Leo Kanner first described a group of atypically developing children with two distinct symptom profiles: “autistic aloneness” and an “obsessive insistence on the preservation of sameness” (Kanner, Reference Kanner1943). Since that time, various diagnostic algorithms have characterized what has become known as autism spectrum disorder (ASD), the most recent of which is DSM-5 (American Psychiatric Association, 2013; Baker, Reference Baker2013). Paralleling Kanner's original description, DSM-5 describes both social and nonsocial behavioral domains: (a) deficits in social communication and (b) restricted, repetitive patterns of behavior, interests, or activities. Although several theoretical frameworks such as theory of mind and social motivation theory (Baron-Cohen, Leslie, & Frith, Reference Baron-Cohen, Leslie and Frith1985; Chevallier, Kohls, Troiani, Brodkin, & Schultz, Reference Chevallier, Kohls, Troiani, Brodkin and Schultz2012) have been proposed to explain the social deficits seen in ASD, few have adequately accounted for the nonsocial behavioral differences, or captured the bidimensional nature of autism.
In 2009, Baron-Cohen formulated the empathizing–systemizing theory of autism (Baron-Cohen, Reference Baron-Cohen2009), which integrated both social and nonsocial aspects in terms of deficits in empathy (understanding and feeling the mental states of others, or empathizing) and the drive to analyze or construct rule-based systems or patterns (termed systemizing). In individuals with ASD, empathizing traits are inversely correlated with systemizing preferences (Baron-Cohen, Richler, Bisarya, Gurunathan, & Wheelwright, Reference Baron-Cohen, Richler, Bisarya, Gurunathan and Wheelwright2003; Grove, Baillie, Allison, Baron-Cohen, & Hoekstra, Reference Grove, Baillie, Allison, Baron-Cohen and Hoekstra2013); in other words, individuals who score highest for systemizing traits (such as being more detail oriented, tending to follow a structured routine, and being rigidly governed by rules) are more likely to score lower in the empathizing domain (such as valuing relationships, judging the feelings of others, and expressing emotions). Males tend to score higher in the systemizing domain, while females generally tend to score higher in the empathizing domain, which may also help to explain some of the gender differences in ASD (Baron-Cohen, Knickmeyer, & Belmonte, Reference Baron-Cohen, Knickmeyer and Belmonte2005).
Consistent with the concept of systemizing, children with ASD appear to have a preference for predictable, repetitive patterns of sensory stimulation, as observed in the tendency to line up toys, categorize objects, and fixate on familiar patterns and interests. One study using automated eye tracking showed that a sample of young children with ASD had a visual preference for videos of computer-generated geometric patterns over social stimuli (i.e., children moving rhythmically; Pierce, Conant, Hazin, Stoner, & Desmond, Reference Pierce, Conant, Hazin, Stoner and Desmond2011). In the current study, our first aim was to test whether children with ASD prefer to view more organized or structured real-life images with predictable, repetitive patterns. Using a novel eye-tracking paradigm viewing a range of systemized images, we obtained a quantitative assessment of interest or motivation based on gaze preference, independent of verbal or cognitive ability (Boraston & Blakemore, Reference Boraston and Blakemore2007).
In searching for clues regarding the etiology of ASD, few studies have successfully linked both social and nonsocial aspects of the condition within a single neurobiological framework (Strathearn, Reference Strathearn2009). Over recent years, an increasing number of studies, both human and nonhuman mammalian, have suggested that the brain's oxytocinergic system may play a role in the development or manifestation of ASD (Bartz & Hollander, Reference Bartz, Hollander, Neumann and Landgraf2008; Domes et al., Reference Domes, Heinrichs, Kumbier, Grossmann, Hauenstein and Herpertz2013; Domes, Kumbier, Heinrichs, & Herpertz, Reference Domes, Heinrichs, Kumbier, Grossmann, Hauenstein and Herpertz2014; Gordon et al., Reference Gordon, Vander Wyk, Bennett, Cordeaux, Lucas, Eilbott and Pelphrey2013; Modi & Young, Reference Modi and Young2012; Sala et al., Reference Sala, Braida, Lentini, Busnelli, Bulgheroni, Capurro and Chini2011; Stavropoulos & Carver, Reference Stavropoulos and Carver2013). Oxytocin is a neuromodulatory peptide hormone synthesized in the paraventricular and supraoptic nuclei of the hypothalamus, whose function has been linked with social bonding and attachment (Feldman, Reference Feldman2012; Insel, Reference Insel2010; Kim, Fonagy, Koos, Dorsett, & Strathearn, Reference Kim, Fonagy, Koos, Dorsett and Strathearn2014; Strathearn, Fonagy, Amico, & Montague, Reference Strathearn, Fonagy, Amico and Montague2009; Strathearn, Iyengar, Fonagy, & Kim, Reference Strathearn, Iyengar, Fonagy and Kim2012). More nuanced effects have been described recently in the areas of social salience and motivation, reward sensitivity, and stress reactivity (Crespi, Reference Crespi2016). Generally, oxytocin appears to diminish stress responses while increasing the salience and reward value of social cues.
In studies involving healthy individuals, intranasally administered oxytocin enhances feelings of empathy (Hurlemann et al., Reference Hurlemann, Patin, Onur, Cohen, Baumgartner, Metzler and Kendrick2010) and many other characteristics associated with the empathizing trait, such as mentalizing (Domes, Heinrichs, Michel, Berger, & Herpertz, Reference Domes, Heinrichs, Michel, Berger and Herpertz2007), trust (Kosfeld, Heinrichs, Zak, Fischbacher, & Fehr, Reference Kosfeld, Heinrichs, Zak, Fischbacher and Fehr2005), emotion recognition (Domes et al., Reference Domes, Lischke, Berger, Grossmann, Hauenstein, Heinrichs and Herpertz2010) and eye gaze (Guastella, Mitchell, & Dadds, Reference Guastella, Mitchell and Dadds2008). These effects of oxytocin appear to be mediated via functional changes in specific brain regions such as the amygdala and the nucleus accumbens, which are involved in emotion processing and social motivation (Gordon et al., Reference Gordon, Jack, Pretzsch, Vander Wyk, Leckman, Feldman and Pelphrey2016). However, the biological mechanisms by which oxytocin may affect systemizing have not been well studied. One group showed the effect of oxytocin on amygdala functioning to be moderated by individual levels of systemizing (Hirosawa et al., Reference Hirosawa, Kikuchi, Higashida, Okumura, Ueno, Shitamichi and Minabe2012), suggesting that oxytocin may indirectly influence systemizing via its effect on empathizing in the amygdala. As empathizing and systemizing are strongly negatively correlated (Grove et al., Reference Grove, Baillie, Allison, Baron-Cohen and Hoekstra2013), increasing social attunement may diminish the preference for structure and predictability.
Earlier studies in autism using a single dose of intranasal oxytocin revealed promising effects, such as increased emotion recognition when observing the eye region of human faces (Andari et al., Reference Andari, Duhamel, Zalla, Herbrecht, Leboyer and Sirigu2010; Domes et al., Reference Domes, Kumbier, Heinrichs and Herpertz2014; Guastella et al., Reference Guastella, Einfeld, Gray, Rinehart, Tonge, Lambert and Hickie2010), enhanced brain amygdala response to social cues (Domes et al., Reference Domes, Heinrichs, Kumbier, Grossmann, Hauenstein and Herpertz2013, Reference Domes, Kumbier, Heinrichs and Herpertz2014), and increased reciprocated social interaction (Andari et al., Reference Andari, Duhamel, Zalla, Herbrecht, Leboyer and Sirigu2010). Repeated dose studies have been few in number and have shown mixed results. Initial studies showed no significant effects on social interaction skills, repetitive behaviors, or clinical outcomes (Dadds et al., Reference Dadds, MacDonald, Cauchi, Williams, Levy and Brennan2014; Guastella et al., Reference Guastella, Gray, Rinehart, Alvares, Tonge, Hickie and Einfeld2015), but more recent trials have demonstrated modest effects with regard to social behavior and clinical ratings (Watanabe et al., Reference Watanabe, Kuroda, Kuwabara, Aoki, Iwashiro, Tatsunobu and Yamasue2015; Yatawara, Einfeld, Hickie, Davenport, & Guastella, Reference Yatawara, Einfeld, Hickie, Davenport and Guastella2016). Few studies have examined the effect of oxytocin on nonsocial behaviors such as systemizing preference in autism. In one preliminary study, adults with ASD who were given a peripheral infusion of oxytocin showed reduced repetitive behavior (Hollander et al., Reference Hollander, Novotny, Hanratty, Yaffe, DeCaria, Aronowitz and Mosovich2003). Another pilot study testing the chronic administration of intranasal oxytocin in ASD adults showed a marginally significant decrease in lower ordered repetitive behaviors such as stereotyped movements, but no difference in higher order behaviors such as compulsions or insistence on sameness (Anagnostou et al., Reference Anagnostou, Soorya, Chaplin, Bartz, Halpern, Wasserman and Hollander2012). Our second aim was therefore to test whether acutely administered intranasal oxytocin would diminish visual systemizing preferences in ASD.
In summary, we sought (a) to test whether children and adolescents with ASD preferentially view more highly organized/structured images, using a novel preferential looking eye tracking paradigm, and (b) to examine the effect of intranasal oxytocin on these visual systemizing preferences. We hypothesized that participants with ASD would show a visual preference for more highly systemized real-life photographs, compared with matched control participants, and that this preference would be reduced after administration of intranasal oxytocin.
Methods
Participants
Twenty males with ASD (M age = 12.5 years) were recruited through the Autism Treatment Network and Simons Simplex Collection at the Baylor College of Medicine/Texas Children's Hospital's Autism Center. All participants with ASD were assessed by a trained clinician who ensured that they met clinical criteria for autistic disorder or Asperger disorder according to DSM-IV criteria. The Autism Diagnostic Interview—Revised and the Autism Diagnostic Observation Schedule (ADOS; Lord, Rutter, DiLavore, & Risi, Reference Lord, Rutter, DiLavore and Risi1999) were used to confirm that each participant met formal, standardized criteria for a diagnosis of ASD. All of the Simons Simplex Collection children except one received an ADOS Module 3, indicating that they were verbally fluent and spoke in complete, complex sentences. The remaining child received an ADOS Module 2, indicating at least phrase speech and/or simple sentences. These children also received the Differential Abilities Scales—II, which revealed a mean full-scale IQ of 92.3 ± 12.6, a verbal IQ of 92.5 ± 18.4, and a nonverbal IQ of 97.0 ± 12.5. All participants were aged between 8 and 19 years, were English speaking, and were not intellectually impaired, as established by developmentally appropriate psychometric testing. Exclusion criteria included uncorrected visual impairment; neurological disease, such as stroke, brain tumor, or seizure disorder; or a history of meningitis or encephalitis.
The participants with ASD were matched by age and race to a group of typically developing males (M age = 13.2 years), who were recruited from a community database at the Children's Nutrition Research Center, Baylor College of Medicine. Prior to enrollment, control participants were screened by phone to rule out possible learning or behavior problems (i.e., a diagnosed learning disability or need for special education services, as per parent report). Parents subsequently completed the Strengths and Difficulties Questionnaire (Goodman, Reference Goodman1997), which confirmed that all control participants scored at “low risk” for emotional, behavioral, or hyperactivity/concentration problems. To screen for autistic traits, parents completed the Social Communications Questionnaire (Rutter, Bailey, & Lord, Reference Rutter, Bailey and Lord2003), with all control participants scoring well below the study-imposed, stringent cutoff score of 10 (mean score = 2.9, range = 0–7). Parents of participants from both groups completed the Vineland Adaptive Behaviors Scales (2nd ed.; Sparrow, Cicchetti, & Balla, Reference Sparrow, Cicchetti and Balla2005), and as expected, participants with ASD scored significantly lower on overall adaptive functioning (t = 5.2, p < .001), and on each of the individual subscales: social (t = 7.8, p < .001), communication (t = 4.4, p < .001), and daily living (t = 3.3, p < .005; Table 1).
Table 1. Demographic and behavioral characteristics of ASD and control participants

Note: ASD, autism spectrum disorder; ADHD, attention-deficit/hyperactivity disorder.
aBased on medication prescription.
Two participants with ASD did not return for their second visit, and 2 others were excluded from further analyses; 1 had been started on a psychotropic medication between visits, and 1 had poor-quality eye-tracking data. Thus, a total of 16 participants with ASD and 16 corresponding control participants were included in the final analysis. Of those with ASD, 11 (69%) were taking psychotropic medications at the time of the study visits, including stimulant medications for attention-deficit/hyperactivity disorder (n = 6), medication for depression or anxiety (n = 4), and mood stabilizers (n = 4). None of the control participants were on psychotropic medications or had comorbid conditions, as assessed using the Strengths and Difficulties Questionnaire. Additional participant characteristics are described in Table 1.
The Institutional Review Board at the Baylor College of Medicine approved the research protocol, and the participants’ parents provided written informed consent and certified child assent. The Food and Drug Administration approved the use of oxytocin in this study as part of an Investigational New Drug application.
Apparatus and stimuli
Eye tracker
Real-time eye movements and visual fixations were recorded using the Tobii T120 automated eye tracker (Tobii Technology, 2011). The eye tracker consists of two infrared light sources and a camera (640 × 480 frame rate at 30 frames/s) integrated into a 17-in. computer monitor, on which the visual stimuli were displayed. Using corneal reflection techniques to track both bright and dark pupils, x and y coordinates of eye gaze position were recorded at a rate of 60 Hz, with an estimated accuracy between 0.4 and 0.6 degrees (i.e., difference between measured and actual gaze direction; Tobii, 2011). Spatial resolution was estimated to be 0.3 degrees, and drift due to change in lighting was 0.1 degrees.
Visual stimuli
The systemizing picture task. In order to test gaze preferences in ASD, we developed the systemizing picture task, using real-life images of people, animals, scenes, and objects (Figure 1). Each slide consisted of four related images, positioned in each quadrant of the slide, which were equivalent in terms of social relevance but which varied systematically in terms of organization or structure (systemizing).
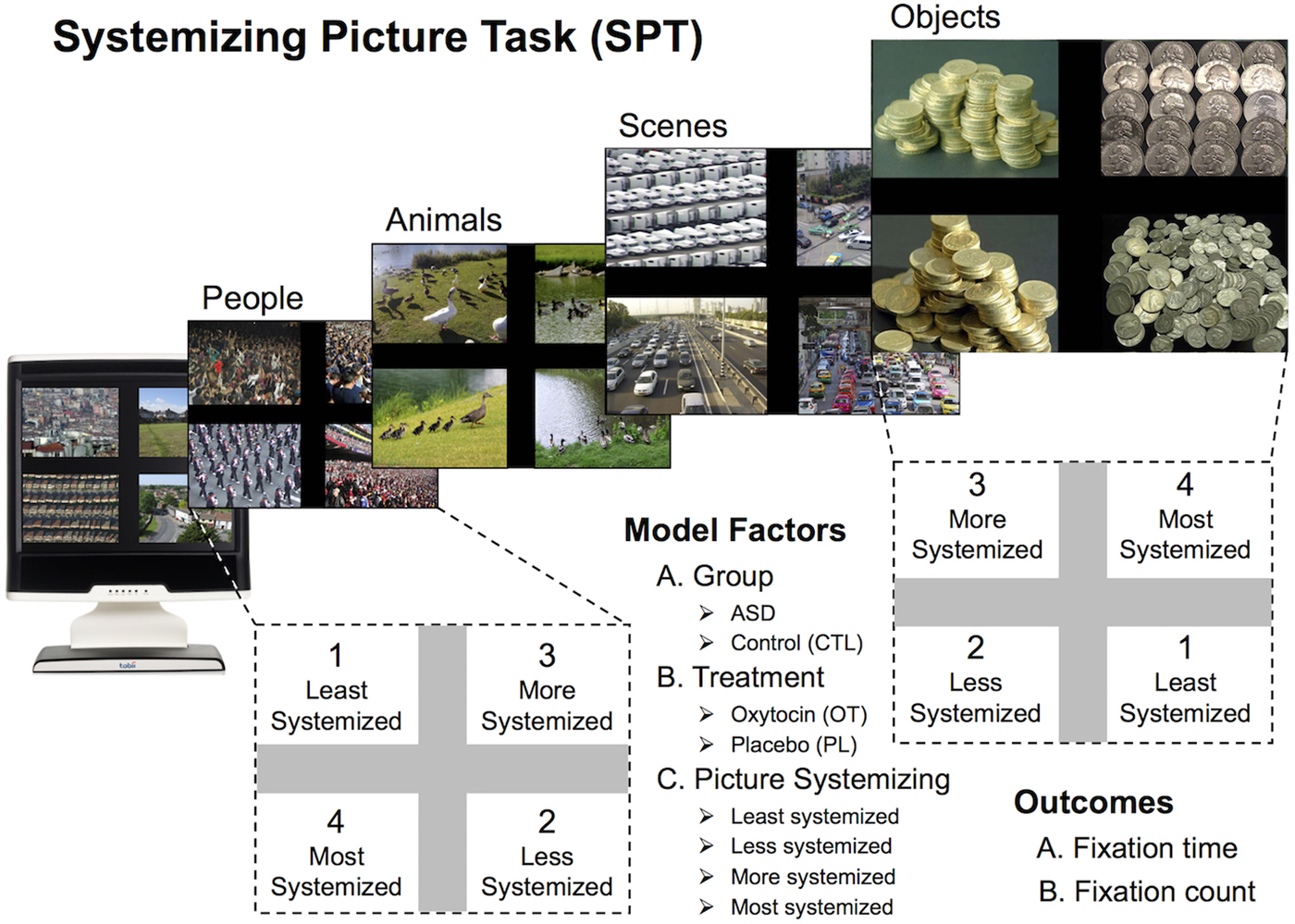
Figure 1. (Color online) Systemizing picture task. Representative slides displayed on the Tobii eye-tracker, showing four levels of picture systemizing. The position of each picture was pseudorandomized across four versions of the task, and the position of the most and least systemized pictures varied between slides. The three primary factors in the model included autism spectrum disorder versus control group, treatment (oxytocin vs. placebo), and level of picture systemizing. Comparable but not actual images used in the task are shown, because of copyright constraints.
Upon initial selection of 22 slides, we sampled 8 individuals to rate each picture on level of systemizing. Systemizing was defined for participants as the level of organization indicated by the picture. Across the participants, this initial rating yielded a median concordance (Kendall W) of .911. We selected 70% agreement on the rank ordering as the minimum criteria (W = .84). On this basis, 7 slides were eliminated. We eliminated an additional slide that appeared to distract participants. The remaining 14 slides were rerated on systemizing by a further group of 39 individuals, which yielded a median Kendall W coefficient of 0.871. These final systemizing ratings were used to assign rank orders to the four pictures within each slide.
Each picture in this final set of slides was also rated for valence (9-point scale), humanness (5-point scale), aliveness (5-point scale), and movement (5-point scale). These ratings yielded high reliability as indicated by Cronbach αs of 0.858, 0.961, 0.837, and 0.828, respectively. The averaged humanness, aliveness, and movement scores were later entered into statistical models to control for potential confounding effects.
Four different versions of the experiment were created, varying the positions of the four pictures within each slide, in order to minimize possible gaze bias based on picture location. The different versions were consistent between matched subjects.
Procedure
Oxytocin administration
As part of a randomized, double-blinded crossover study, participants were randomly assigned one nasal solution on Visit 1 and the alternate solution on Visit 2. The intranasal oxytocin (Syntocinon Spray, Novartis, Switzerland) was purchased from a Swiss pharmacy, and a local compounding pharmacy prepared the placebo sprays using identical base ingredients (for 5 ml of placebo solution: 12.5 mg chlorobutanol hemihydrate, 2 mg methyl-4-hydroxybenzoate, 1 mg propyl-4-hydroxybenzoate, 14 mg citric acid monohydrate, 125 mg glycerine 85%, and 4.85 g purified water; for active drug: 24 IU oxytocin/ml). The oxytocin solution and the placebo (base alone) were stored in identical glass-metered pump spray bottles (0.1 ml/spray) and labeled to ensure that both participants and experimenters were blind to the drug treatment.
Trained research assistants administered either the oxytocin or the placebo spray intranasally. Children 16 years and older received the usual adult dose of 24 IU oxytocin (10 puffs alternating between nostrils with the head in the upright position, each spray containing 2.4 IU), while doses for younger children were titrated downward, similar to Guastella et al. (Reference Guastella, Einfeld, Gray, Rinehart, Tonge, Lambert and Hickie2010). Those aged 12–15 years received 7 puffs (16.8 IU), and children aged 8–11 years received 5 puffs (12 IU total).
After oxytocin administration, participants rested while watching an animated film or reading quietly. A similar activity was provided during the second visit, to standardize the environment across visits. Approximately 30 min after oxytocin administration, the participants commenced the eye-tracking experiment.
Eye-tracking experiment
After performing a 9-point eye-tracking calibration using the Tobii Studio software (Tobii Technology, 2011), an instruction slide appeared that was read aloud to the participant, followed by a practice slide. A crosshair was shown for 1 s between each slide, and participants were instructed to gaze directly at the crosshair when present. During the experiment, the monitor displayed each systemizing picture task slide for 12 s (except for three participants who viewed the slides for 8 s each; sensitivity analysis revealed no difference in the pattern of findings, and therefore these 3 participants were included in all of the analyses). After the entire eye-tracking computer task was completed, participants received compensation, were presented with a certificate of completion, and scheduled to return in approximately 1–2 weeks’ time for the corresponding study visit.
Data analysis
Eye-gaze data were extracted from the four pictures/regions of interest (ROI) on each slide, which depicted the theme at four different levels of organization or structure (i.e., least systemized, less systemized, more systemized, and most systemized; Figure 1). We exported and analyzed, as our outcome measures: (a) fixation time (total duration of fixation) and (b) fixation count (total number of fixations) in each ROI. Fixation was defined as an eye gaze that remained stationary (within 1-degree radius, or approximately 35 pixels) for a minimum of 80 ms. Proportion variables were calculated by dividing the fixation values (time or count) recorded for each quadrant by the respective total value obtained for the entire slide. As the proportions were not distributed normally, a range of transformations was performed. Intercept-adjusted logarithmic transforms offered the closest approximation to normality. The random-effect models controlled for ROI location on the slide and visit order, as we probed for significance of the Group × Treatment × Systemizing interaction. Participants’ performance across four levels of systemizing was represented by random effects parameters. Three additional variables (participant age, picture humanness, and picture aliveness) were added to the model individually, to test for any confounding influence. Analyses were performed in STATA/SE 13.1 and IBM SPSS Statistics (version 21).
Results
Means and standard errors of fixation variables are presented in Table 2. Consistent with our hypothesis, the ASD group showed a visual preference for more highly systemized images in the standard placebo (PL) condition (Figure 2). There was a significant linear trend of increase in the fixation time of ASD participants with increasing levels of picture systemizing (ASDPL βsystemizing = 0.0041 ± 0.002 SE, z = 2.00, p = .045), while no such effect was seen in the control group (CTLPL βsystemizing = 0.0006 ± 0.0018, z = 0.32, p = .748).

Figure 2. (Color online) Visual systemizing preferences in autism spectrum disorder (ASD) versus control (CTL) groups, in the placebo condition. (a) Representative heat maps visualizing differences in relative gaze duration between ASD and control groups for all participants in one version of the systemizing picture task (n = 8) on the coin slide. Participants with ASD showed a gaze preference for the most systemized image. Relative duration was calculated from the sum of individual fixation times relative to the total fixation time for that slide. (b) Participants with ASD showed a linear trend of increase in the fixation time with increasing levels of picture systemizing. This trend was absent in the control group. The interaction effect was not significant. The y-axis represents log-transformed proportion variable. The statistical significance of the linear trend of increase is noted; *p < .05.
Table 2. Visual systemizing preferences based on fixation time and count, in control and ASD groups as a function of oxytocin and level of picture systemizing

Note: The duration and frequency values (estimated marginal means ± SE) are log-transformed proportion values, adjusted for the total duration and number of fixations recorded for each slide. Wald χ2 and p values are those obtained for the best-fitting mixed-effects models for the respective outcome variables, including a subject-level random intercept and a random coefficient for level of picture systemizing. ASD, autism spectrum disorder; df = 10 for all models.
Oxytocin, however, reversed the overall patterns of fixation observed under placebo, as reflected in a significant three-way interaction (βGroup×Treatment×Systemizing = –0.0119 ± 0.0039, z = –3.04, p = .002; Figure 3). For the ASD group, oxytocin eliminated the tendency to fixate on more highly systemized pictures, as seen in a significant two-way interaction (ASD βTreatment× Systemizing = –0.0070 ± 0.0030, z = –2.33, p = .020; Figure 3a). In particular, after receiving intranasal oxytocin compared with placebo, ASD participants showed significantly less fixation on the most systemized pictures (ASD βtreatment = –0.0215 ± 0.0104, z = –2.07, p = .038). The ASD group showed no persisting gaze preference after receiving intranasal oxytocin (OT; ASDOT βsystemizing = –0.0030 ± 0.0022, z = –1.38, p = .169), similar to what was seen in the control group under placebo conditions (CTLPL βsystemizing = 0.0006 ± 0.0018, z = 0.32, p = .748; Figure 3b).

Figure 3. Visual systemizing preferences as a function of drug treatment and level of picture systemizing in autism spectrum disorder (ASD) versus control (CTL) groups. Oxytocin (OT) significantly reversed the overall patterns of fixation time observed under placebo (PL). (a) Oxytocin compared with placebo significantly reduced the tendency for participants with ASD to fixate on the most systemized stimuli, while increasing their fixation on the least systemized stimuli, with marginal significance. After oxytocin, participants with ASD did not demonstrate a significant gaze preference. (b) In the control group, oxytocin produced a significant preference to view more highly systemized pictures. The y-axis represents log-transformed proportion variables. The statistical significance of the linear trend of increase or differences between oxytocin and placebo at specific levels of systemizing is noted; †p < .10, *p < .05, **p < .01, ***p < .001. NS, nonsignificant.
In contrast, the control group became more likely to fixate on more systemized pictures after oxytocin administration (CTLOT βsystemizing = –0.0065 ± 0.0018, z = 3.60, p < .001), with a significant two-way interaction observed (CTL βTreatment×Systemizing = 0.0054 ± 0.0026, z = 2.13, p = .033; Figure 3b). However, the difference in fixation time after oxytocin versus placebo was not statistically significant for either most systemized or least systemized pictures (CTL βtreatment = 0.0164 ± 0.0104, z = 1.57, p = .116; βtreatment = –0.0147 ± 0.0104, z = –1.41, p = .158, respectively).
Overall, oxytocin resulted in a decrease in systemizing preference (βsystemizing) in ASD participants (Figure 3a), but an increase in systemizing preference for control participants (Figure 3b), resulting is a highly significant two-way interaction effect (OT βGroup×Systemizing = –0.0087 ± 0.0028, z = –3.07, p = .002). Figure 4a illustrates the change in slope after oxytocin administration comparing ASD and CTL groups.

Figure 4. (a) Contrasting effect of intranasal oxytocin (OT) on visual systemizing preference, based on the slope of fixation time versus systemizing, in autism spectrum disorder (ASD) versus control (CTL) participants. Oxytocin decreased the systemizing preference in ASD participants, but increased the systemizing preference in control participants. (b) Model of how oxytocin levels may be associated with systemizing and empathizing characteristics in ASD and typically developing individuals. Effects of systemizing are based on results displayed in (a). We propose that intranasal oxytocin may increase oxytocin levels in ASD participants from the low to normal range, while shifting the level in typically developing participants from normal to a high range. An inverted pattern may represent the effect of oxytocin on empathizing abilities, based on other studies of intranasal oxytocin in ASD and typically developing populations (see text). PL, placebo.
Similar gaze patterns were seen after examining the total number of fixations in each region of interest (fixation count; Table 2), except that the interaction effect between drug treatment and level of systemizing in the control group was no longer statistically significant (CTL βTreatment×Systemizing = 0.0041 ± 0.0022, z = 1.86, p = .063).
None of the independent ratings of picture humanness or aliveness were associated with fixation time or count, and these variables did not alter any of the significant main or interaction effects. The age variable was also not associated with either of the outcome variables and did not significantly alter any of the observed effects.
Discussion
This study is the first to examine the effects of intranasal oxytocin, in both ASD and non-ASD groups, on what we have termed visual systemizing preference, a nonsocial manifestation of ASD. First, we confirmed that our novel eye-tracking task was able to distinguish children/adolescents with ASD from non-ASD controls. Second, we showed that when intranasal oxytocin was administered to ASD participants, it effectively eliminated the difference between ASD and control participants. However, it was surprising that oxytocin also increased systemizing preference when administered to control participants. Understanding how oxytocin affects both social and nonsocial aspects of ASD, in affected and unaffected individuals, may provide clues for understanding its potential role in the etiology and treatment of autism.
Oxytocin in ASD
There are no currently approved pharmacotherapies available to treat the core social or nonsocial manifestations of ASD. As noted above, early studies provided preliminary evidence for the beneficial effects of single-dose intranasal oxytocin in adolescents and adults with autism, with regard to both social communication (Andari et al., Reference Andari, Duhamel, Zalla, Herbrecht, Leboyer and Sirigu2010; Guastella et al., Reference Guastella, Einfeld, Gray, Rinehart, Tonge, Lambert and Hickie2010) and repetitive behavior (Anagnostou et al., Reference Anagnostou, Soorya, Chaplin, Bartz, Halpern, Wasserman and Hollander2012; Hollander et al., Reference Hollander, Novotny, Hanratty, Yaffe, DeCaria, Aronowitz and Mosovich2003). Subsequent studies of repeated administrations of oxytocin have shown variable effects on behavioral outcomes, ranging from no significant effects on social skills or repetitive behavior (Dadds et al., Reference Dadds, MacDonald, Cauchi, Williams, Levy and Brennan2014; Guastella et al., Reference Guastella, Gray, Rinehart, Alvares, Tonge, Hickie and Einfeld2015) to improved social reciprocity and clinical ratings of behavior (Watanabe et al., Reference Watanabe, Kuroda, Kuwabara, Aoki, Iwashiro, Tatsunobu and Yamasue2015; Yatawara et al., Reference Yatawara, Einfeld, Hickie, Davenport and Guastella2016). Other human studies in nonclinical populations have shown improvements in a range of responses relevant to autism, including emotional empathy (Hurlemann et al., Reference Hurlemann, Patin, Onur, Cohen, Baumgartner, Metzler and Kendrick2010), eye gaze (Guastella et al., Reference Guastella, Mitchell and Dadds2008), trust (Baumgartner, Heinrichs, Vonlanthen, Fischbacher, & Fehr, Reference Baumgartner, Heinrichs, Vonlanthen, Fischbacher and Fehr2008), and the ability to read the affective state of others (Domes et al., Reference Domes, Heinrichs, Michel, Berger and Herpertz2007).
Despite an abundance of media attention on the promises of oxytocin therapy in conditions such as ASD, concerns remain about anomalous effects or even unanticipated adverse effects (Bales & Perkeybile, Reference Bales and Perkeybile2012; Bales et al., Reference Bales, Perkeybile, Conley, Lee, Guoynes, Downing and Mendoza2013; Bartz et al., Reference Bartz, Simeon, Hamilton, Kim, Crystal, Braun and Hollander2011; De Dreu et al., Reference De Dreu, Greer, Handgraaf, Shalvi, Van Kleef, Baas and Feith2010; Miller, Reference Miller2013). A number of studies have noted variations in oxytocin response depending on individual differences and other contextual factors (Bartz, Zaki, Bolger, & Ochsner, Reference Bartz, Zaki, Bolger and Ochsner2011). Other work has shown, for example, that plasma oxytocin response to social cues (mothers interacting with their infants) varies with adult temperament, being positively associated with sensitivity to moods, emotions, and physical sensations, but negatively associated with compulsive and task-oriented behavior (Strathearn et al., Reference Strathearn, Iyengar, Fonagy and Kim2012), which parallel empathizing and systemizing traits, respectively.
Systemizing and empathizing in ASD
Baron-Cohen first hypothesized that autism may result from an extreme tendency to systemize or seek out regularities or predictable patterns and rules (Baron-Cohen, Reference Baron-Cohen2009). This preference for patterned over unpatterned information may help to explain some of the restricted, repetitive behaviors and interests characteristically observed in individuals with ASD. Consistent with this model, our findings, along with those of Pierce et al. (Reference Pierce, Conant, Hazin, Stoner and Desmond2011), show that individuals with ASD display a preference for more patterned or systemized visual stimuli.
According to Baron-Cohen's theory, hypersystemizing is also often accompanied by deficits in empathizing, or the ability to identify and appropriately respond to another person's thoughts and emotions (Baron-Cohen & Wheelwright, Reference Baron-Cohen and Wheelwright2004), with deficits in social communication characteristically seen in ASD. Individuals with ASD tend to score higher than the general population on measures of systemizing but lower on empathizing measures (Auyeung et al., Reference Auyeung, Wheelwright, Allison, Atkinson, Samarawickrema and Baron-Cohen2009; Baron-Cohen, Reference Baron-Cohen2009).
Effects of oxytocin in non-ASD participants
As noted, we also found that oxytocin increased visual systemizing preference in control participants, resulting in a gaze preference similar to ASD participants under placebo conditions (Figure 3a, b). Although not predicted, this result is consistent with a growing body of literature demonstrating that the effects of oxytocin are significantly modulated by individual differences. Oxytocin receptor-deficient mice, which display social deficits as well as cognitive inflexibility, serve as an animal model for ASD. After intraventricular administration of oxytocin, social deficits improve and cognitive inflexibility decreases (Ring, Reference Ring2011; Sala et al., Reference Sala, Braida, Lentini, Busnelli, Bulgheroni, Capurro and Chini2011). However, when the same dose of oxytocin is administered to unaffected mice, the opposite effect is observed. While Hollander et al. (Reference Hollander, Novotny, Hanratty, Yaffe, DeCaria, Aronowitz and Mosovich2003) reported that intravenous oxytocin resulted in a decrease in repetitive behaviors in adults with autism, higher oxytocin levels are associated with an increase in repetitive behavior in adults with obsessive–compulsive disorder (Leckman et al., Reference Leckman, Goodman, North, Chappell, Price, Pauls and McDougle1994). Although intranasal oxytocin enhances emotion recognition in individuals with ASD (Bartz et al., Reference Bartz, Zaki, Bolger, Hollander, Ludwig, Kolevzon and Ochsner2010; Guastella et al., Reference Guastella, Einfeld, Gray, Rinehart, Tonge, Lambert and Hickie2010), one study showed that oxytocin decreases the accuracy of emotion recognition when administered to a normative sample (Cardoso, Ellenbogen, & Linnen, Reference Cardoso, Ellenbogen and Linnen2014). Oxytocin enhances trust and cooperation in normative populations (Declerck, Boone, & Kiyonari, Reference Declerck, Boone and Kiyonari2010; Kosfeld et al., Reference Kosfeld, Heinrichs, Zak, Fischbacher and Fehr2005), but appears to have the opposite effect in individuals with borderline personality disorder (Bartz, Simeon, et al., Reference Bartz, Simeon, Hamilton, Kim, Crystal, Braun and Hollander2011). Finally, Groppe et al. (Reference Groppe, Gossen, Rademacher, Hahn, Westphal, Grunder and Spreckelmeyer2013) showed that the effect of oxytocin on performance in a social reward and punishment task depended on whether the individuals had high or low sociability ratings. In the low sociability group, oxytocin increased social reward while avoiding punishment, whereas reduced performance was seen in the high sociability group.
Together, these findings suggest that an optimal range may exist for oxytocin functioning, with its behavioral effects dependent on individual differences that may reflect baseline oxytocin levels, oxytocin receptor status, or interactions with other neuroendocrine factors. A similar “optimal range” model for autism was proposed by Zoghbi and Bear (Reference Zoghbi and Bear2012), based on “high” versus “low” levels of gene expression and effects on synaptic plasticity. In our study, intranasal oxytocin appears to move ASD participants toward more typical responses in the systemizing picture task, while moving control participants further away (Figure 4). Studies have suggested that individuals with ASD may have lower peripheral levels of oxytocin compared to matched controls (Feldman, Golan, Hirschler-Guttenberg, Ostfeld-Etzion, & Zagoory-Sharon, Reference Feldman, Golan, Hirschler-Guttenberg, Ostfeld-Etzion and Zagoory-Sharon2014; Green et al., Reference Green, Fein, Modahl, Feinstein, Waterhouse and Morris2001; Modahl et al., Reference Modahl, Green, Fein, Morris, Waterhouse, Feinstein and Levin1998), and that intranasally administered oxytocin increases plasma oxytocin levels (Andari et al., Reference Andari, Duhamel, Zalla, Herbrecht, Leboyer and Sirigu2010). If peripheral oxytocin levels reflect central oxytocin functioning (McGregor, Callaghan, & Hunt, Reference McGregor, Callaghan and Hunt2008; Strathearn et al., Reference Strathearn, Fonagy, Amico and Montague2009; Valstad et al., Reference Valstad, Alvares, Egknud, Matziorinis, Andreassen, Westlye and Quintana2017), then giving intranasal oxytocin may move individuals with ASD from the low to the normal range of functioning (Figure 4b). Likewise, giving oxytocin to neurotypical control participants may move them from the normal to the high oxytocin range, resulting in the observed behavioral differences.
We hypothesize that a similar, but inverted, pattern may also exist for the effect of oxytocin on social responses in ASD, with other studies providing evidence for a nonlinear effect on behaviors more closely associated with empathizing (Bales et al., Reference Bales, van Westerhuyzen, Lewis-Reese, Grotte, Lanter and Carter2007, Reference Bales, Perkeybile, Conley, Lee, Guoynes, Downing and Mendoza2013; Bartz, Simeon, et al., Reference Bartz, Simeon, Hamilton, Kim, Crystal, Braun and Hollander2011; Cardoso et al., Reference Cardoso, Ellenbogen and Linnen2014; Sala et al., Reference Sala, Braida, Lentini, Busnelli, Bulgheroni, Capurro and Chini2011; Figure 4b). Additional studies, which should also collect peripheral measures of oxytocin before and after intranasal administration, are needed to explore this question more fully.
Limitations and future directions
Several limitations of this study need to be considered. Having created a novel eye-tracking task to examine gaze preference for more organized or structured images, the systemizing picture task has not yet been validated against other measures of systemizing. The original systemizing quotient (Baron-Cohen et al., Reference Baron-Cohen, Richler, Bisarya, Gurunathan and Wheelwright2003) and autism spectrum quotient (Baron-Cohen, Wheelwright, Skinner, Martin, & Clubley, Reference Baron-Cohen, Wheelwright, Skinner, Martin and Clubley2001) were not suitable for administration in children and adolescents, and a subsequent children's version that relied solely on parent report (Auyeung et al., Reference Auyeung, Wheelwright, Allison, Atkinson, Samarawickrema and Baron-Cohen2009) was not available at the time that the study commenced. Comparing our task with other validated measures of systemizing will help to expand our understanding of its meaning and clinical significance.
Although we sought to control for variations in picture composition and meaning, such as the humanness and aliveness, other physical aspects of the pictures, such as color, intensity, or background features, may have also influenced gaze preference. Our desire to use real-world images limited our ability to standardize many of these other features. Although age, when added as a covariate, did not significantly alter study findings, the enrolled groups did encompass a broad range of ages, with a relatively small number of participants. Future studies enrolling larger numbers, and including female participants, may be able to look more specifically for age and gender response differences. An additional limitation includes a lack of systematic information on psychiatric comorbidity. Further studies are warranted to investigate possible contributions of psychiatric comorbidity to the findings reported here.
Additional studies will also need to test whether intranasal oxytocin enhances functioning in real-life environments, or influences long-term outcomes in ASD. Only after more intensive studies are conducted evaluating outcome will we know whether a potential opportunity exists to combine pharmacological treatment with therapeutic/behavioral interventions.
Conclusion
Most recent prevalence estimates indicate that up to 1 in 68 children may be affected by ASD (Centers for Disease Cintrol and Prevention, 2016), with affected individuals and families usually requiring intensive and costly therapeutic intervention to optimize long-term developmental and behavioral outcomes. Understanding the neuroendocrine basis for this condition is an urgent imperative. This study provides preliminary evidence for a possible role of oxytocin in the etiology and/or treatment of autism by demonstrating its effect on a nonsocial manifestation of ASD. With oxytocinergic pathways highly sensitive to early developmental experience via epigenetic mechanisms (Champagne, Reference Champagne2008; Champagne, Diorio, Sharma, & Meaney, Reference Champagne, Diorio, Sharma and Meaney2001; Kumsta, Hummel, Chen, & Heinrichs, Reference Kumsta, Hummel, Chen and Heinrichs2013), this understanding may provide an even more important opportunity for early intervention and/or prevention.






