


In this paper, we describe racial discrimination as a critical form of adversity that has the potential to impact stress biology. We note that the key dimensions of adversity associated with racial discrimination (negative social evaluation, social exclusion, and uncontrollability) may have particularly strong impacts on hypothalamic–pituitary–adrenal (HPA) axis activity, as measured through salivary cortisol, and argue that adolescence may serve as a sensitive period for these impacts. We highlight ethnic racial identity (ERI) as a counteracting positive regulatory influence on HPA axis activity, due to its promotion of positive evaluations of the self and feelings of inclusion in one's ethnic or racial group. In past research, our team has found that Black individuals who reported high adolescent racial discrimination had low adult cortisol levels (hypocortisolism), compared to Black and white individuals who reported lower levels of racial discrimination.Footnote 1 Here, we present new analyses showing that ERI, measured prospectively from ages 12 through 32, predicts better regulated cortisol levels in adulthood, particularly for Black participants. We also describe ongoing research that explores whether the promotion of ERI during adolescence can reduce ethnic and racial disparities in stress biology and in emotional health and academic outcomes.
Racial discrimination as an adverse experience
Racial discrimination, defined here as unfair treatment attributed to one's race (Benner & Graham, Reference Benner and Graham2013), is a powerful stressor that is relatively understudied in the developmental literature on early adversity. Racial discrimination takes a variety of forms, including: systemic and institutional racism, such as unfair laws, policies and treatment in housing, work, and academic environments; overt racism, such as verbal, psychological, or physical abuse based on race; racial microaggressions (more subtle and commonplace racial slights and insults), and racial teasing (Benner & Graham, Reference Benner and Graham2013; English et al., Reference English, Lambert, Tynes, Bowleg, Zea and Howard2020; Levy, Heissel, Richeson, & Adam, Reference Levy, Heissel, Richeson and Adam2016; Pager, Bonikowski, & Western, Reference Pager, Bonikowski and Western2009; Pager & Shepherd, Reference Pager and Shepherd2008; Sue et al., Reference Sue, Capodilupo, Torino, Bucceri, Holder, Nadal and Esquilin2007). Racial discrimination events occur both in person and in online environments, and may be experienced either directly or vicariously, through witnessing or hearing about discrimination experiences of others of the same race or ethnicity (English et al., Reference English, Lambert, Tynes, Bowleg, Zea and Howard2020). In between racial discrimination experiences, the memory of past discrimination events or anticipation of future events may have continuing impact. All of these forms of discrimination are contextualized and interpreted within a historical and present structure of hegemonic power and influence in the United States that prioritizes and privileges white Americans.
By adolescence, most members of stigmatized ethnic and racial minority groups are aware of racial discrimination in their environments (e.g., neighborhoods) and in school settings (for review see Brown & Bigler, Reference Spears Brown and Bigler2005). For example, Fisher, Wallace, and Fenton (Reference Fisher, Wallace and Fenton2000) found that Black and Latino adolescents (between ages 13 and 19) were more likely than their Asian and non-Hispanic white peers to report being the victims of institutional discrimination. Discrimination by teachers and peers is also much more commonly reported by Black adolescents when compared to their white peers (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015). In a recent study of Black adolescents living in the United States using a daily diary methodology, an average of five instances of racial discrimination experiences (either direct, or vicarious and both online and offline) were reported per day by Black youth (English et al., Reference English, Lambert, Tynes, Bowleg, Zea and Howard2020), with online racial discrimination experiences being particularly common. Recent police-related killings of unarmed Black teens and adults have spurred national and international protests and civil unrest, bringing widespread societal and media attention to the pervasiveness and severity of racial discrimination experiences for individuals of color, and particularly for Black Americans.
The vast majority of racial discrimination experiences occur for persons of color, who both report and have been objectively determined to experience extensive unfair treatment according to race (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015; Brondolo et al., Reference Brondolo, Brady, Thompson, Tobin, Cassells, Sweeney and Contrada2008; Broudy et al., Reference Broudy, Brondolo, Coakley, Brady, Cassells, Tobin and Sweeney2007; Jackson & Stewart, Reference Jackson and Stewart2003; Pager et al., Reference Pager, Bonikowski and Western2009; Pager & Shepherd, Reference Pager and Shepherd2008). White individuals may perceive themselves to be treated unfairly due to their race/ethnicity; such reports are however less frequent, less severe, and have lesser consequences (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015; Stepanikova & Oates, Reference Stepanikova and Oates2017), occurring in the context of both historic and present-day racial privilege (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015; Stepanikova & Oates, Reference Stepanikova and Oates2017).Footnote 2
Due to pervasive exposure to, and stronger impacts of, racial discrimination for youth of color, we and others have argued that racial discrimination is an important and understudied contributor to ethnic and racial disparities in mental and physical health as well as academic outcomes (Levy et al., Reference Levy, Heissel, Richeson and Adam2016). A recent meta-analysis finds that racial–ethnic discrimination during adolescence has significant detrimental effects on 11 different indicators of well-being, including more depressive and internalizing symptoms, greater psychological distress, lower self-esteem, greater externalizing behaviors, more risky sexual behaviors, greater substance use, more associations with deviant peers, as well as lower academic motivation, lower academic engagement and achievement (Benner et al., Reference Benner, Wang, Shen, Boyle, Polk and Cheng2018). For example, among Latino and Black adolescents, perceived discrimination from teachers and/or expecting future discrimination are related to lower grades, less academic motivation, and lower academic success (Alfaro, Umaña-Taylor, Gonzales-Backen, Bámaca, & Zeiders, Reference Alfaro, Umaña-Taylor, Gonzales-Backen, Bámaca and Zeiders2009; Eccles, Wong, & Peck, Reference Eccles, Wong and Peck2006; Neblett, Phillip, Cogburn, & Sellers, Reference Neblett, Philip, Cogburn and Sellers2006; Stone & Han, Reference Stone and Han2005; Taylor et al., Reference Taylor, Casten, Flickinger, Roberts and Fulmore1994). Given its wide range of negative associations with socioemotional, academic, and economic well-being for youth of color, racial discrimination needs to be investigated more fully as an early adverse experience, identifying the dimensions of adversity associated with it, and the cognitive, emotional, and biological pathways by which it affects this broad array of outcomes.
Dimensions of early adverse experiences
Recent theoretical models have argued for the importance of identifying the specific psychological dimensions associated with particular adverse exposures (Dickerson & Kemeny, Reference Dickerson and Kemeny2004; McLaughlin, Sheridan, & Lambert, Reference McLaughlin, Sheridan and Lambert2014). In doing so, researchers can better isolate the specific neurobiological systems these experiences activate and identify the psychobiological pathways by which various types of adverse experiences impact academic, mental, and physical health outcomes (Dickerson, Gruenewald, & Kemeny, Reference Dickerson, Gruenewald and Kemeny2004; Dickerson & Kemeny, Reference Dickerson and Kemeny2004; McLaughlin et al., Reference McLaughlin, Sheridan and Lambert2014).
In one prominent categorization, McLaughlin et al. (Reference McLaughlin, Sheridan and Lambert2014) proposed two key dimensions along which adverse experiences vary: deprivation (the absence of expected environmental inputs and complexity) and threat (the presence of experiences that represent a threat to one's physical integrity), with each dimension having distinct effects on neurobiology. Dickerson, Kemeny, and colleagues also argued for the importance of identifying the specific psychological dimensions of experiences in understanding how they relate to neurobiological mechanisms (Dickerson et al., Reference Dickerson, Gruenewald and Kemeny2004; Dickerson & Kemeny, Reference Dickerson and Kemeny2004). They highlight the importance of threat to the “social self” as another key dimension of adversity, distinct from physical threat. Dickerson et al. (Reference Dickerson, Gruenewald and Kemeny2004) define threats to the social self as relating to “one's social esteem, status, and acceptance” (Dickerson et al., Reference Dickerson, Gruenewald and Kemeny2004). They argue that negative social evaluation and rejection or social exclusion are key threats to the social self. In our evolutionary past, these social threats were detrimental to both reproduction and survival, thus representing fundamental threats to both emotional and physical well-being. As a result, humans are highly sensitive to perceptions of past, present, or anticipated negative social evaluation and social exclusion. Dickerson et al. (Reference Dickerson, Gruenewald and Kemeny2004) theorize that these threats to the social self can activate negative self-evaluative emotions (especially shame) and promote social withdrawal, as well as activation of biological stress pathways. They argue that shame results when “negative social evaluations are transformed into negative self-evaluations” (p. 1195).
In a meta-analytic study (Dickerson & Kemeny, Reference Dickerson and Kemeny2004), social-evaluative threat (the possibility of being negatively judged by others) and uncontrollability (having limited control over situations and outcomes) were identified as particularly powerful activators of the HPA axis, a key mediator of the body's biological response to stress. Perceived social exclusion and the related construct of loneliness have also been identified as powerful stressors with implications for HPA axis functioning and for immediate and long-term health outcomes (Cacioppo et al., Reference Cacioppo, Hawkley, Crawford, Ernst, Burleson, Kowalewski and Berntson2002; Doane & Adam, Reference Doane and Adam2010; Goosby, Bellatorre, Walsemann, & Cheadle, Reference Goosby, Bellatorre, Walsemann and Cheadle2013; Stroud et al., Reference Stroud, Foster, Papandonatos, Handwerger, Granger, Kivlighan and Niaura2009). Importantly, the flip sides of these dimensions – positive self- and social-evaluations and perceptions of social inclusion, acceptance, and support – are strongly associated with positive developmental and health outcomes (Baumeister & Leary, Reference Baumeister and Leary1995; Blackhart, Eckel, & Tice, Reference Blackhart, Eckel and Tice2007; Uchino, Cacioppo, & Kiecolt-Glaser, Reference Uchino, Cacioppo and Kiecolt-Glaser1996; Wentzel, Jablansky, & Scalise, Reference Wentzel, Jablansky and Scalise2020).
Dimensions of racial discrimination
Examining racial discrimination according to the dimensions of adversity outlined by McLaughlin et al. (Reference McLaughlin, Sheridan and Lambert2014), we argue that for individuals of color, racial discrimination contains moderate to high levels of threat. For example, physical threats associated with racial discrimination for individuals of color involve increased rates of police-related violence resulting in injury or death and, for adolescents, perceived lack of safety both at school and while traveling between home and school (Lacoe, Reference Lacoe2015; Shelley et al., Reference Shelley, Pickett, Mancini, McDougle, Rissler and Cleary2017). Racial discrimination can also result in moderate levels of deprivation, including reduced access to resources such as high-quality neighborhoods, housing, and educational and job opportunities (Pager et al., Reference Pager, Bonikowski and Western2009; Pager & Shepherd, Reference Pager and Shepherd2008) that can contribute to lower socioeconomic conditions for people of color.
Racial discrimination is particularly high on both of the dimensions that Dickerson and Kemeny (Reference Dickerson and Kemeny2004) found to be particularly powerful activators of the HPA axis: social–evaluative threat and social rejection. Racial discrimination carries chronic social–evaluative threat, particularly in the everyday racial microaggressions faced by individuals of color (Sue et al., Reference Sue, Capodilupo, Torino, Bucceri, Holder, Nadal and Esquilin2007). Racial microaggressions are “brief and commonplace daily verbal, behavioral, or environmental indignities, whether intentional or unintentional, that communicate hostile, derogatory, or negative racial slights and insults toward people of color” (Sue et al., Reference Sue, Capodilupo, Torino, Bucceri, Holder, Nadal and Esquilin2007, p. 271). For example, being mistaken for a service worker, being assumed to be less intelligent or accomplished, and being avoided by white people when walking down the street are common microaggressions (Sue et al., Reference Sue, Capodilupo, Torino, Bucceri, Holder, Nadal and Esquilin2007). Implied in its definition and in the provided examples is conscious or unconscious negative social evaluation of the target of the microaggression.
The commonplace nature of microaggressions results in both repeated exposure and ongoing anticipation of their occurrence, such that racial discrimination serves as a chronic form of social–evaluative threat. Pervasive structural and institutional forms of racial discrimination, such as unfair treatment in housing, work, and academic settings (Pager & Shepherd, Reference Pager and Shepherd2008), serve as forms of social exclusion that can have life-long consequences, and thus also contribute to the chronic nature of racial discrimination as a stressor.
Racial discrimination is also high on the dimension of uncontrollability. Racial discrimination is often directed at people on the basis of their skin color or other immutable physical characteristics that aggressors attribute to their race/ethnicity. The power structures and conscious and unconscious biases that give rise to modern instances of racial discrimination are also notoriously difficult to change, and the consequences of generations of egregious mistreatment of persons of color (e.g. slavery) cannot be erased. Thus, individuals of color have relatively little control over whether and when they will be exposed to instances of racial discrimination. Reducing racial discrimination exposures would likely have positive impacts on health and development; but doing so is not easily accomplished, leading scholars to search for factors that can reduce the impact of racial discrimination or reverse some of its negative effects (Neblett, Rivas-Drake, & Umaña-Taylor, Reference Neblett, Rivas-Drake and Umaña-Taylor2012).
Ethnic racial identity as a protective or promotive factor
Given the difficulties in changing exposures to discrimination, researchers have focused on identifying features that diminish or counteract the negative effects of racial discrimination, serving as either a protective factor (diminishing the negative impact of high levels of racial discrimination) or promotive factor (promoting positive outcomes regardless of level of racial discrimination) (Neblett et al., Reference Neblett, Rivas-Drake and Umaña-Taylor2012). The presence of a strong ERI is one key protective and promotive factor that has emerged from the literature on emotional and behavioral development in diverse samples of adolescents (Rivas-Drake et al., Reference Rivas-Drake, Seaton, Markstrom, Quintana, Syed, Lee and Yip2014). Just as racial discrimination is a powerful elicitor of feelings of negative social evaluation and social exclusion, a strong ERI may be an important source of feelings of positive social evaluation and social acceptance and belonging.
A well-developed ERI is a multidimensional and rich composite of connectedness, beliefs, and practices associated with one's ethnic heritage (Umaña-Taylor, Reference Umaña-Taylor, Schwartz, Luyckx and Vignoles2011). Several components of ERI have been found to protect against the negative effects of racial discrimination and promote positive outcomes, particularly for youth of color (Rivas-Drake, Reference Rivas-Drake2011; Sellers, Copeland-Linder, Martin, & Lewis, Reference Sellers, Copeland-Linder, Martin and Lewis2006). In terms of protective effects, feeling good about and proud of one's race/ethnicity (a construct we are calling Individual Regard, also known as affirmation) (Umaña-Taylor et al., Reference Umaña-Taylor, Quintana, Lee, Cross, Rivas-Drake, Schwartz and Seaton2014) has been found to buffer African-American adolescent self-esteem from the effects of racial discrimination (Harris-Britt, Valrie, Kurtz-Costes, & Rowley, Reference Harris-Britt, Valrie, Kurtz-Costes and Rowley2007). Individual Regard contains strong elements of positive self- and social evaluation. In addition, strong positive feelings about and connectedness to one's ethnic–racial community (Community Regard) buffers adolescents against the impact of racial discrimination on academic achievement, emotional well-being, and risky behaviors (Wong, Eccles, & Sameroff, Reference Wong, Eccles and Sameroff2003). Community Regard contains strong elements of perceived social acceptance. In terms of promotive effects, behavioral enactment of traditions connected with one's race/ethnicity (called Ethnic Behavior, after Phinney, Reference Phinney1990) predicts positive coping strategies (Fischer & Shaw, Reference Fischer and Shaw1999; Scott, Reference Scott2003), improved academic performance (Brown, Linver, Evans, & DeGennaro, Reference Brown, Linver, Evans and DeGennaro2009), and better psychological adjustment (Rivas-Drake, Reference Rivas-Drake2011). Ethnic–racial traditions are often enacted in a group setting, contributing to feelings of social belonging.
Adolescence as a sensitive period for the effects of ethnic–racial experiences
Recent research with both animal models and humans has provided support for the idea that adolescence may serve as a sensitive period for the effects of social experience on the brain, on stress biology, and on behavior (Blakemore & Mills, Reference Blakemore and Mills2014; McCormick, Mathews, Thomas, & Waters, Reference McCormick, Mathews, Thomas and Waters2010). Accompanying dramatic physical and cognitive changes (Steinberg & Morris, Reference Steinberg and Morris2001), adolescents also negotiate substantial emotional and social demands as peer and romantic relationships take on an increasingly prominent role in their lives. In addition, exploring and defining one's sense of self, or identity, is an important focus during this stage of adolescence (Umaña-Taylor et al., Reference Umaña-Taylor, Quintana, Lee, Cross, Rivas-Drake, Schwartz and Seaton2014). Establishing positive peer and romantic relationships and the formation of personal and ethnic identity are among the key developmental tasks of adolescence (Umaña-Taylor et al., Reference Umaña-Taylor, Quintana, Lee, Cross, Rivas-Drake, Schwartz and Seaton2014). Neurobiological changes during adolescence reflect and contribute to the prominence of these tasks, including ongoing maturation of regions of the brain serving social cognition and emotion regulation (Blakemore & Mills, Reference Blakemore and Mills2014; Luna, Garver, Urban, Lazar, & Sweeney, Reference Luna, Garver, Urban, Lazar and Sweeney2004). The convergence of the multiple changes of adolescence creates a period of heightened stress that is accompanied by increases in both basal levels and reactivity of glucocorticoid stress hormones and increased sensitivity of glucocorticoid-sensitive brain regions (DePasquale, Donzella, & Gunnar, Reference DePasquale, Donzella and Gunnar2019; Romeo, Reference Romeo2017). The convergence of increased social challenge with increased biological stress reactivity and ongoing brain development and plasticity has led to the suggestion that adolescence may reflect a sensitive period, with the social experiences of adolescence becoming built into adolescents’ developing minds, bodies, and brains in ways that can have a long-term impact on adult health and emotional, cognitive, and behavioral functioning (Blakemore & Mills, Reference Blakemore and Mills2014).
Given that the key psychosocial dimensions of racial discrimination (negative social evaluation and social exclusion) present challenges to one of the key developmental tasks of adolescence (social acceptance and inclusion), we expect that racial discrimination experiences will have particularly potent impacts during adolescence, relative to childhood and adulthood.
Much of the work on adolescence as a sensitive period has focused on the impacts of stressful experiences on stress biology and on regions of the brain that continue to show structural maturation during adolescence, including the amygdala, hippocampal formation, and prefrontal cortex (Romeo, Reference Romeo2017). However, recent research has pivoted to also focus on positive and supportive experiences during adolescence – that is, the presence of positive social and individual assets that may contribute to normative brain development and positive regulation of stress biology (Blakemore & Mills, Reference Blakemore and Mills2014; Doane & Zeiders, Reference Doane and Zeiders2014; Gunnar, DePasquale, Reid, & Donzella, Reference Gunnar, DePasquale, Reid and Donzella2019). We propose that ERI will also have important implications for adolescent social and neurobiological development, particularly for aspects of stress biology that are sensitive to social threats and social acceptance, including HPA axis activity.
Racial discrimination and HPA axis activity
Alterations in HPA axis activity are one way that racial discrimination is thought to affect daily functioning, emotional and physical health, and cognitive performance (Berger & Sarnyai, Reference Berger and Sarnyai2015; Clark, Anderson, Clark, & Williams, Reference Clark, Anderson, Clark and Williams1999; Harrell et al., Reference Harrell, Burford, Cage, Nelson, Shearon, Thompson and Green2011; Levy et al., Reference Levy, Heissel, Richeson and Adam2016; Paradies et al., Reference Paradies, Ben, Denson, Elias, Priest, Pieterse and Gee2015). Levels of cortisol, the primary hormonal product of the HPA axis and most common indicator of HPA axis activity in human research, increase in response to acute stress. Basal (non-stress) cortisol levels also follow a strong diurnal rhythm. Cortisol levels are typically high upon waking, increase further in the 30–40 min after waking, and then decline to low levels by bedtime (Adam & Kumari, Reference Adam and Kumari2009). The postawakening cortisol increase is known as the cortisol awakening response (CAR); the change (typically decline) in cortisol from waking to bedtime is called the diurnal cortisol slope, and the area under the curve (AUC) defined by cortisol levels across the waking day is called total cortisol (Adam & Kumari, Reference Adam and Kumari2009).
In a number of studies, racial discrimination has been linked to alterations in HPA axis activity (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015; Zeiders, Hoyt, & Adam, Reference Zeiders, Hoyt and Adam2014). A recent meta-analysis of 16 studies did not find an overall association between racial discrimination and cortisol when multiple types of basal cortisol measures and cortisol reactivity were examined together as a single outcome (Korous, Causadias, & Casper, Reference Korous, Causadias and Casper2017). Of note, stronger effect sizes were found, however, for experimental studies in which acute cortisol reactivity to racism was examined (Korous et al., Reference Korous, Causadias and Casper2017). By contrast, a recent systematic review concluded that systematic associations between discrimination and both basal cortisol and cortisol reactivity existed, with the size and direction of associations depending on the timing and chronicity of the discrimination and the type of cortisol outcome (Busse, Yim, Campos, & Marshburn, Reference Busse, Yim, Campos and Marshburn2017). The authors observed that studies of chronic exposure to discrimination experienced over multiple decades tended to show evidence of flatter diurnal cortisol slopes, whereas experimentally induced exposures to acute discriminatory events (e.g. racism, sexism) were associated with increased cortisol reactivity (Busse et al., Reference Busse, Yim, Campos and Marshburn2017).
Studies of discrimination in adolescents and emerging adults tend to conform to Busse et al.'s (Reference Busse, Yim, Campos and Marshburn2017) analysis, finding higher adolescent discrimination associated with stronger acute cortisol reactivity (Doane & Zeiders, Reference Doane and Zeiders2014), higher overall cortisol output (Huynh, Guan, Almeida, McCreath, & Fuligni, Reference Huynh, Guan, Almeida, McCreath and Fuligni2016; Zeiders, Doane, & Roosa, Reference Zeiders, Doane and Roosa2012), and a flatter diurnal cortisol slope (Huynh et al., Reference Huynh, Guan, Almeida, McCreath and Fuligni2016; Zeiders et al., Reference Zeiders, Hoyt and Adam2014). Do the effects of discrimination during adolescence having lasting effects on the HPA axis? In our previously published prospective longitudinal study, racial discrimination levels from early adolescence and early adulthood were related to cortisol levels measured in adulthood, at age 32. We found that racial discrimination experienced during adolescence had a stronger effect on adult cortisol than racial discrimination experienced during the early adult years, providing some support for the notion of an adolescent sensitive period effect. With higher histories of racial discrimination during adolescence, diurnal cortisol slopes were flatter for both Black and white youth, whereas total diurnal cortisol across the day was considerably lower only for Black participants (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015).
This low, flat cortisol pattern of cortisol, known as hypocortisolism (Fries, Hesse, Hellhammer, & Hellhammer, Reference Fries, Hesse, Hellhammer and Hellhammer2005; Heim, Ehlert, & Helhammer, Reference Heim, Ehlert and Helhammer2000), is thought to emerge due to wear and tear on the HPA axis from prior repeated or chronic activation of the HPA axis (Miller, Chen, & Zhou, Reference Miller, Chen and Zhou2007). This supports the idea of a transition from an HPA axis that is acutely activated by discrimination to one that becomes under-activated over time with chronic exposure (Busse et al., Reference Busse, Yim, Campos and Marshburn2017). Hypocortisolism predicts worse educational (Blair, Reference Blair2010; Blair, Granger, & Peters Razza, Reference Blair, Granger and Peters Razza2005; Maldonado et al., Reference Maldonado, Fernandez, Trianes, Wesnes, Petrini, Zangara and Ambrosetti2008), cognitive (Vedhara, Hyde, Gilchrist, Tytherleigh, & Plummer, Reference Vedhara, Hyde, Gilchrist, Tytherleigh and Plummer2000), and mental and physical health outcomes (Crofford et al., Reference Crofford, Pillemer, Kalogeras, Cash, Michelson, Kling and Wilder1994; Heim et al., Reference Heim, Ehlert and Helhammer2000). Thus, the chronic effects of discrimination on HPA axis activity in adolescence may have long-term consequences for adult well-being. Consistent with the Dickerson et al. (Reference Dickerson, Gruenewald and Kemeny2004) theory, recent research from our group suggests that racial discrimination affects the HPA axis by way of negative social-evaluative (NSE) emotions. In a daily diary study, we found that racial discrimination was associated with higher NSE. In turn, higher average and day-to-day changes in NSE emotions predicted flatter diurnal cortisol slopes. In formal mediation analyses, NSE emotions mediated the associations between racial discrimination and flatter diurnal cortisol slopes (Hittner & Adam, Reference Hittner and Adam2019).
Ethnic racial identity and HPA axis activity
We argue here that one pathway by which ERI confers positive effects on psychological and behavioral outcomes may be by having positive regulatory effects on HPA axis activity. Very little research, however, on ERI and cortisol has been conducted. One recent study examined associations between ERI and diurnal cortisol activity in a sample of Hispanic Americans, finding that the ERI dimension of affirmation was associated with steeper diurnal cortisol slopes (Zeiders, Causadias, & White, Reference Zeiders, Causadias and White2018).
Research on the development of ERI suggests that from childhood to adolescence, youth of color move from a simple ERI to a more sophisticated and multifaceted ERI (Umaña-Taylor et al., Reference Umaña-Taylor, Quintana, Lee, Cross, Rivas-Drake, Schwartz and Seaton2014). Much less research exists on whether and how ERI changes over the transition to young adulthood (Umaña-Taylor et al., Reference Umaña-Taylor, Quintana, Lee, Cross, Rivas-Drake, Schwartz and Seaton2014). One might expect that as various facets of ERI change (increase, decrease, or differentiate) with development, so too may the influence of those facets on HPA axis activity. Thus, it may be useful to study how various facets of ERI change over the course of adolescence and early adulthood. No prior study has prospectively examined trajectories of ERI development from adolescence through early adulthood, or the long-term impacts of adolescent and young adult ERI on adult cortisol.
Current research
Using data from a 20-year prospective longitudinal sample following participants from adolescence to their early 30s (the same data as used in Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015), we first examine normative developmental changes in ERI through adolescence and young adulthood. We then test whether differences in various ERI components measured prospectively in adolescence and young adulthood are associated with individual differences in adult diurnal cortisol profiles, and whether associations between ERI and cortisol are stronger for Black individuals than white individuals or differ by developmental period.
Given that Black individuals encounter more racial discrimination and, as a result, greater HPA axis dysregulation (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015), we anticipate that ERI, and its dimensions promoting feelings of social acceptance and belonging, will have larger effects on the HPA axis functioning of Black compared to white participants. Given the important developments that occur both in ERI and the HPA axis during adolescence (Umaña-Taylor et al., Reference Umaña-Taylor, Quintana, Lee, Cross, Rivas-Drake, Schwartz and Seaton2014), and the rapidly changing social and biological worlds of adolescents described above, we expect that ERI during adolescence will have stronger effects on adult cortisol than ERI in early adulthood. Conversely, given that ongoing developments in identity and stress-sensitive regions occur through emerging adulthood (Steinberg & Morris, Reference Steinberg and Morris2001; Syed & Azmitia, Reference Syed and Azmitia2008; Taber-Thomas & Pérez-Edgar, Reference Taber-Thomas and Pérez-Edgar2015), ERI may continue to have important influences on HPA axis activity throughout the early adult period.
Method
Study overview
This study draws on subsets of participants from the Maryland Adolescent Development in Context Study (MADICS), a longitudinal study of 1482 adolescents (n = 879 Black, 49% female) designed to examine how adolescent experiences and contexts relate to developmental outcomes in adolescence and young adulthood (Eccles et al., Reference Eccles, Wong and Peck2006; Wong et al., Reference Wong, Eccles and Sameroff2003). MADICS participants were recruited at age 12 and interviewed eight times across 20 years –7th grade (Waves 1 and 2), 8th grade (Wave 3), 11th grade, (Wave 4), a year after high school (Wave 5), 3 years after high school (Wave 6), around age 30 (Wave 7), and around age 32 (Wave 8) (Brodish et al., Reference Brodish, Cogburn, Fuller-Rowell, Peck, Malanchuk and Eccles2011; Fuller-Rowell et al., Reference Fuller-Rowell, Cogburn, Brodish, Peck, Malanchuk and Eccles2012). For more information about the full procedure for each wave, please refer to the MADICS archived data on the Henry A. Murray Research Archive webpage (https://murray.harvard.edu/dataverse), which includes data collection procedures, questionnaires, and codebooks.
At Wave 8, a small subsample (N = 124) was selected to complete a health add-on study which collected biomarkers of stress and health, including salivary cortisol. Add-on participants were selected to include: (a) approximately equal numbers of Black and white participants; (b) approximately equal numbers of low, medium, and high levels of discrimination in prior waves; and (c) willingness to participate in additional measures that included a seven-day diurnal cortisol data collection period and a seven-day daily diary study.
Participants
Our analyses use two subsets of participants from the full MADICS study. For our examination of ERI trajectories over time, our sample included participants from the full study sample with ERI data available at three data points between Wave 3 and Wave 7, including at least one point in adolescence and at least one point in young adulthood, yielding an analytic sample of N = 685 (62% Black and 60% female). Participants came from families with a Wave 3 average income of $50,030 (range $5,000 to $75,000) and an average parent education level of some college (range of less than high school to advanced degree) (see Table 1).
Table 1. Descriptive data on ethnic racial identity (ERI) trajectory sample and cortisol subsample

For our exploration associations between ERI and adult cortisol levels, we used data from participants in the health add-on study for whom both ERI and Wave 8 cortisol data were available. Participants were excluded from our cortisol analysis if they used corticosteroid-based medications, illicit drugs, or had sleeping patterns (such as shift work) that could impact the accuracy of diurnal cortisol patterns. Data for specific days were excluded if the participant was missing wake time or bedtime samples for that day. The resulting cortisol analysis sample included 112 individuals (45% Black and 61% female) from families with a Wave 1 average income of $52,120 (range $5,000 to $75,000) and an average parent education level of some college (from less than high school to advanced degree).
The smaller cortisol analysis subsample differed significantly from the larger ERI development sample on several variables. There are fewer Black participants in the cortisol subsample (t = −4.12, p < .001). Individuals in the cortisol analyses had higher parent education (t = 2.27, p = .024), lower levels of overall, adolescent, and young adult Individual Regard (cumulative: t = −3.56, p < .001; adolescent: t = −2.32, p = .021; young adult: t = −3.52, p < .001); and lower levels of overall, adolescent, and young adult Racial Centrality (cumulative: t = −2.20, p = .028; adolescent: t = −2.12, p = .034; young adult: t = −2.05, p = .041) (see Table 1). No other significant differences between the two subsamples were found.
Racial discrimination
Racial discrimination was assessed in Waves 3 to 7 with questions designed to be developmentally appropriate for each age. Adolescent (vs. adult) racial discrimination was most strongly related to adult hypocortisolism in past research with this dataset (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015), and we were interested in how concurrently measured (adolescent) and subsequent (early adult) ERI levels might modify these adolescent racial discrimination effects. As a result, our analyses focused on an adolescent racial discrimination scale (α = .87) created from eight items assessed at each of Wave 3 and Wave 4. Items (e.g. “How often have you felt that teachers/counselors discourage you from taking certain classes because of your race?”) were standardized and averaged within and across waves (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015; Wong et al., Reference Wong, Eccles and Sameroff2003).
Ethnic–racial identity
ERI was assessed using youth self-reports on ten items available from Waves 3–7. Questions were adapted from contemporaneous instruments or developed by the MADICS team; they have been used in previous publications (Peck, Brodish, Malanchuk, Banerjee, & Eccles, Reference Peck, Brodish, Malanchuk, Banerjee and Eccles2014; Wong et al., Reference Wong, Eccles and Sameroff2003). Exploratory factor analysis indicated the presence of four ERI scales within the 10 items: Individual Regard (single item: How proud are you of your racial background?), Community Regard (e.g. People of my race/ethnicity are very supportive of each other; people of my race/ethnicity have a culturally rich heritage, α = .90), Racial Centrality (e.g. How important is it for you to know about your racial background?; α= .83), and Ethnic Behavior (e.g. How often do you celebrate any special days connected to your racial background?; α = .81). ERI items were averaged within each wave to create wave-specific ERI scale scores. Wave-specific scores were then averaged together across all years (Waves 3–7) to create cumulative ERI measures and within the two developmental periods to create adolescent ERI (Waves 3 and 4) and young adult ERI (Waves 5–7) scores. Racial Centrality items were not available in Wave 7; we therefore use Waves 3–6 for the cumulative version and Waves 5 and 6 for the young adult version of this specific ERI subscale). The four ERI scales of Individual Regard, Community Regard, Centrality, and Ethnic Behavior were used in separate analyses, given limited degrees of freedom and their moderate to high intercorrelations (ranging from r = .35 to r = .72). Note that we consider both Individual and Community Regard to be subsets of the larger construct of Private Regard, which was introduced by Sellers, Chavous, and Cooke (Reference Sellers, Chavous and Cooke1998).
Salivary cortisol
Saliva samples were gathered upon waking (“as soon as possible after opening your eyes”), 30 min after waking (signaled by a preset timer), and immediately before bedtime for seven consecutive days. Participants provided saliva by passive drool through a small plastic straw into a 2 mL polypropylene vial. Participants were told to: (a) keep sampling materials by their bed; (b) not eat, drink, or brush their teeth during the 30 min prior to sampling; (c) record their exact times of collection on each vial; (d) store samples in their refrigerators after collection; and (e) return samples by regular postal mail (Clements & Parker, Reference Clements and Parker1998).
Cortisol values were natural log-transformed for use in analyses, and cortisol outcome variables were standardized for ease of interpretation. Data from the second morning cortisol sample were removed if it was taken less than 20 min or greater than 40 min after the first sample. Our primary two outcome variables were the AUC, defined by the cortisol data points across the day (Pruessner, Kirschbaum, Meinlschmid, & Hellhammer, Reference Pruessner, Kirschbaum, Meinlschmid and Hellhammer2003), and the diurnal cortisol slope from waking to bedtime, which was calculated by subtracting bedtime from waking cortisol values and dividing by total time awake each day (Adam & Kumari, Reference Adam and Kumari2009). Indices were calculated for each day of data and averaged across days. We do not include the CAR as an outcome, as our measurement protocol did not conform to current guidelines for CAR measurement (Stalder et al., Reference Stalder, Kirschbaum, Kudielka, Adam, Pruessner, Wüst and Hellhammer2016).
Demographic and health behavior covariates and moderators
Demographic information for participants was reported by parents at Wave 3. We incorporated key demographic covariates known to predict cortisol levels: gender (0 = male; 1 = female), race/ethnicity (0 =white, 1 = Black), parent education (in years), and family income (in thousands of dollars) (Desantis, Kuzawa, & Adam, Reference Desantis, Kuzawa and Adam2015). Information about adolescent ethnic–racial background was self-reported at Wave 1 by youth. Time of waking, reported on the days of cortisol sampling and averaged across days, was also included as a covariate (Adam & Kumari, Reference Adam and Kumari2009). In order to ensure that effects were specific to positive self-regard with respect to race, rather than to general levels of positive self-regard, self-esteem was measured and added as a covariate. Self-esteem measures from the same time period as each ERI variable (i.e. cumulative, adolescent, and young adult) were utilized, and self-esteem by Race×Ethnicity interactions were tested. All continuous variables were standardized; indicator variables were entered uncentered.
Missing data
To address missing data and maintain as large a sample size as possible in our cortisol analyses, we used multiple imputation (MI) techniques (Graham, Reference Graham2009; Jeličić, Phelps, & Lerner, Reference Jeličić, Phelps and Lerner2009; Schafer & Graham, Reference Schafer and Graham2002). Our imputation model was run using SPSS and included all of the relevant variables in our analyses including the dependent variable and interaction terms (Manly & Wells, Reference Manly and Wells2015). Our multiple imputation used 20 imputations and 200 iterations. Means and standard deviations of the dataset including imputed values were similar to those observed in the original nonreplaced sample.
Analysis
Our central analyses explored: (a) how ERI changes from adolescence through young adulthood; (b) the associations between cumulative, adolescent, and young adult ERI and cortisol levels at age 32; and (c) whether these findings vary by race–ethnicity.
Changes in ERI from adolescence through young adulthood
Two-level hierarchical linear modeling (HLM) growth curve analyses were employed using HLM 7 (Raudenbush, Bryk, Cheong, & Congdon, Reference Raudenbush, Bryk, Cheong and Congdon2011) to estimate (a) linear change in aspects of ERI across adolescence and young adulthood (at Level 1) and (b) whether ERI levels and trajectories vary by race/ethnicity (at Level 2) (Bryk & Raudenbush, Reference Bryk and Raudenbush1992). At Level 1, each ERI scale was predicted from age, with age measured in years and centered at each person's age at Wave 5 (the approximate midpoint of data collection; an average of 18.8 years old). In these models, the intercept (π0) represents levels of ERI during Wave 5 (an approximation of average ERI levels), and the growth term (π1) represents the change in ERI per year across all waves of measurement. At Level 2, race/ethnicity (white = 0; Black = 1) was added as a predictor of these coefficients, testing whether ERI levels at Wave 5 (midpoint levels) and change in ERI from adolescence through young adulthood vary by race–ethnicity. Growth trajectory values and significance levels (simple slopes) for Black participants were obtained by rerunning analyses with race/ethnicity re-centered (Black = 0; white = 1). All reported fixed effects used robust standard errors.
Regression models predicting cortisol from ERI levels
In a series of regression models, cortisol outcomes (average cortisol, diurnal cortisol slope) were predicted from adolescent racial discrimination levels and covariates, including race–ethnicity, gender, family income, parent education, time of waking, and levels of self-esteem. Next, levels on each ERI scale were added in separate analyses. Racial discrimination by ERI interactions were also added; because these were not significant in predicting cortisol, they were not included in our final models. Finally, race/ethnicity by racial discrimination and race/ethnicity by ERI interactions were added.
Significant race/ethnicity by ERI interactions were probed using simple effects analysis (Dearing & Hamilton, Reference Dearing and Hamilton2006; Hayes & Matthes, Reference Hayes and Matthes2009; Preacher, Curran, & Bauer, Reference Preacher, Curran and Bauer2006). That is, simple or main effects are reported for both white and Black youth, along with the interaction effects, which reflect the significance of the differences between simple effects. Main effects (simple slope) results for Black participants come from models using a white indicator for race/ethnicity (Black = 0; white = 1). In addition to examining average levels of ERI in each developmental period, we also examined the impact of ERI trajectories (increases or decreases in each ERI variable over time), by extracting the growth coefficients from our HLM models and including them as predictors of cortisol. ERI levels on average and in each developmental period were more predictive of adult cortisol than trajectories. As a result, we focus on ERI levels, rather than ERI growth trajectories as predictors of cortisol in our current analyses.
Results
ERI development over time and individual differences by race/ethnicity
Trajectories of ERI subscales
As seen in Figure 1, Black individuals had higher levels of ERI (as indicated by significant differences at the intercept) than whites across all ERI scales including Individual Regard (b = .88, t = 12.93, p < .001), Community Regard (b = .70, t = 13.03, p < .001), Centrality (b = 1.51, t = 21.17, p < .001), and Ethnic Behavior (b = .60, t = 11.20, p < .001). Individual Regard showed significant declines with age from adolescence through young adulthood (b = −.04, t = −12.79, p < .001) for Black and white participants combined. When trajectories were compared by race/ethnicity, Black and white participants showed significant decreases in Individual Regard over time (b = −.031, t = −8.30, p < .001; b = −.044, t = −7.09, p < .001 respectively), with decreases for white participants being marginally greater than for Black participants (b = −.013, t = −1.75, p = .08).
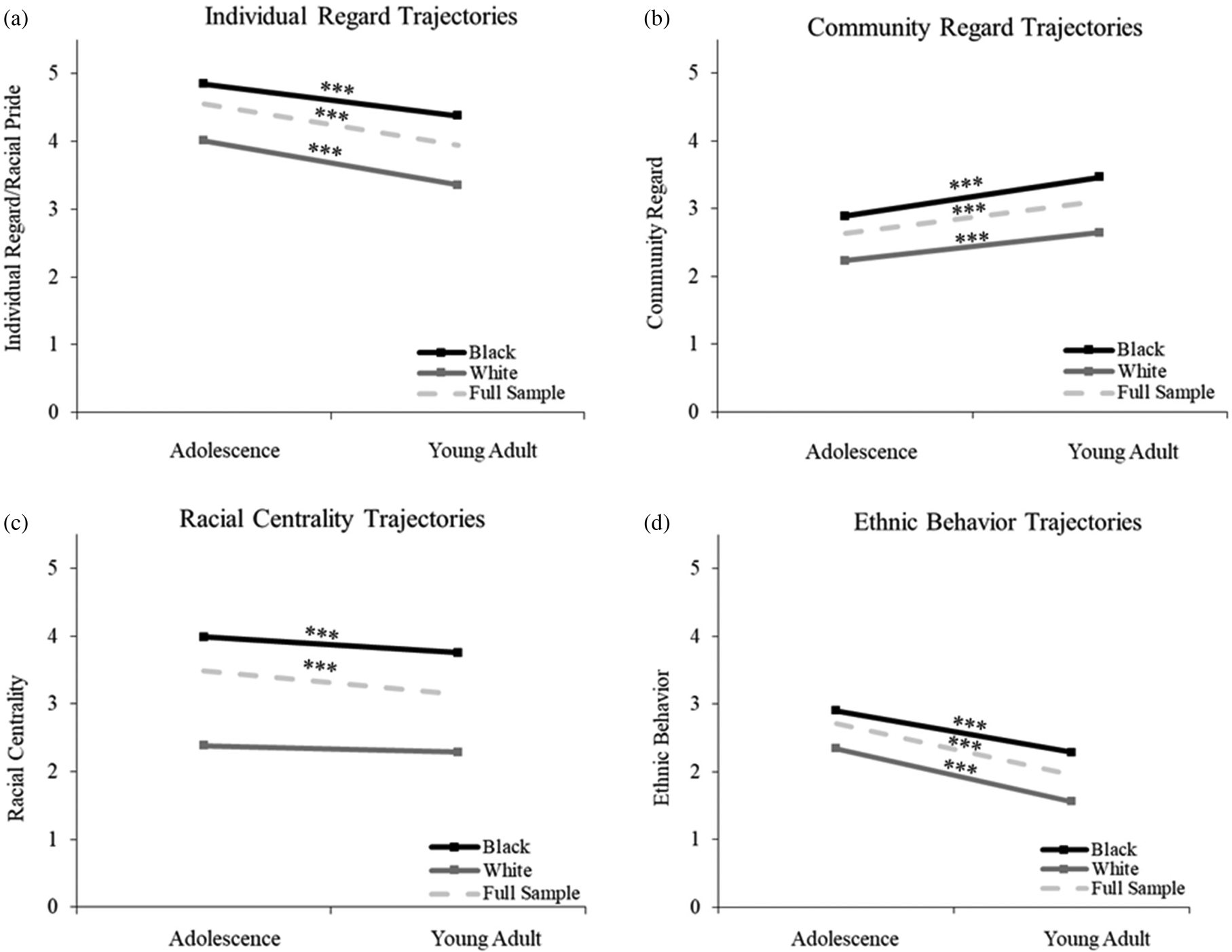
Figure 1. Linear growth trajectories for aspects of ethnic racial identity (ERI) across adolescence and young adulthood for the combined sample (dotted lines) and by race (sold lines). Significant simple slopes are indicated along lines; *<=0.5, **<=.01,***<=.001.
Community Regard increased over time from adolescence through young adulthood (b = .032, t = 9.96, p < .001). Both Black and white participants showed significant increases (Blacks: b = .038, t = 8.74, p < .001; whites: b = .027, t = 5.87, p < .001), with Black participants showing marginally greater increases than white participants (b = .011, t = 1.70, p = .09). Centrality declined significantly with age (b = −.065, t = −.05, p < .001). There was a significant decrease in Centrality for Black participants (b = −.043, t = −4.14, p < .001) but not for white participants (b = −.018, t = −.81, p = .417), but the linear slopes were not significantly different (b = −.025, t = −1.03, p = .305). Ethnic Behavior declined significantly with age (b = −.05, t = −16.15, p < .001) for both Black and white individuals (Blacks: b = −.041, t = −9.67, p < .001; whites: b = −.052, t = −11.23, p < .001), and white participants showed marginally greater decreases than Blacks (b = −.011, t = −1.80, p = .07).
Regression results of ERI levels predicting adult cortisol
Our next models examined associations between ERI variables and both total cortisol levels and diurnal cortisol slopes in adulthood. These multivariate models included racial discrimination, ERI, and racial discrimination and ERI by Race/ethnicity interactions, along with covariates (see Table 2 for ERI and total cortisol; results for cortisol slopes are presented in text). Several of the ERI scales showed significant associations with adult total cortisol, as seen in Table 2 and Figure 2.

Figure 2. Interactions between overall Individual Regard and Community Regard and race/ethnicity (a and b); individual and Community Regard for adolescence (c and d) and young adulthood (e and f). Significance interactions between ethnic racial identity (ERI) and race/ethnicity are indicated by a triangle. Significant simple slopes are indicated along lines; *<=.05, **<=.01, ***<=.001.
Table 2. Regression results of ethnic racial identity (ERI) subscales on total cortisol

Note: RD = racial discrimination. ERI = ethnic and racial identity. Model 1 includes covariates plus adolescent RD and ERI. Model 2 includes covariates, RD, ERI, and the interaction terms for race/ethnicity, RD, ERI, and Self-Esteem. Models include covariates Black, Female, Family Income, Parent Education, and Wake Time. Betas for covariates vary slightly across ERI subscales.
Individual Regard
Individual Regard averaged across both developmental periods was associated with lower total cortisol among whites (b = −.352, t = −2.97, p = 0.003) and higher total cortisol among Black individuals (b = .519, t = 2.19, p = .028), with large and significant differences by ethnicity/race (b = .854, t = 3.03, p = 0.002) (see Figure 2a). Significant race/ethnicity by Individual Regard interactions were also found in each separate developmental period (see Table 2 and Figure 2c and 2e), with patterns suggesting a convergence in cortisol levels between Black and white participants at higher levels of Individual Regard.
Community Regard
Higher Community Regard across the full developmental time span was associated with higher adult total cortisol for Black and white participants combined (b = .243, t = 2.15, p = 0.031); the Community Regard by Race/ethnicity interaction was not statistically significant (b = .218, t = 1.00, p = .317; see Figure 2b). There were no significant associations between Community Regard in adolescence and adult cortisol (see Figure 2d). In young adulthood, however, Community Regard predicted higher adult cortisol across Black and white participants combined (b = .305, t = 2.68, p = .007) (see Figure 2f).
Racial Centrality and Ethnic Behavior
Racial Centrality and Ethnic Behavior did not predict adult total cortisol cumulatively or in any developmental period for either Black and white individuals or Black and white participants combined (see Table 2).
ERI and cortisol slopes
There were very few significant findings for the associations between ERI and wake-to-bedtime cortisol slopes. There were no significant results for Individual Regard or Racial Centrality. For Community Regard, there was a trend for higher young adult Community Regard to predict steeper diurnal cortisol slopes across both Black and white individuals (b = −.232, t = −1.88, p = .06); this effect did not vary significantly by race-ethnicity. Higher young adult Ethnic Behavior associated with flatter slopes for white participants (b = .404, t = −2.24, p = .03 but not for Black participants (b = −.134, t = −.86, p = .391), with a significant difference between the effects of Ethnic Behavior on cortisol slopes for Black vs. white participants (b = −.522, t = −2.12, p = .03).
Discussion
Our study demonstrates that significant changes in ERI occur during adolescence and early adulthood and provides evidence that certain aspects of ERI (particularly Individual Regard and Community Regard) have significant associations with adult cortisol regulation, contributing primarily to a normalizing of (i.e., an increase in) low cortisol levels for Black individuals. Here we discuss these findings in more detail, highlighting the potential impact of these ERI components for cortisol regulation, for disparities in health and education, and for the design of interventions aimed at reducing these disparities.
Developmental changes in ERI
ERI levels are not static phenomena; rather, our data revealed trends in ERI from adolescence through the transition to adulthood. Decreases were seen over the transition to adulthood for both Black and white participants in Individual Regard and Ethnic Behavior and for Black participants only in Racial Centrality. These were offset, however, by relatively strong increases in Community Regard for both Black and white participants. That is, although individual pride in one's race-ethnicity, personal enactment of ethnic–racial traditions, and the importance placed on race-ethnicity (for Black participants only) declined from adolescence through young adulthood, positive regard for one's ethnic–racial community and perceptions of its supportiveness increased over this same time period.
The decrease in Individual Regard and shift toward an appreciation of one's ethnic–racial community (Community Regard) over the transition to early adulthood may reflect an emergence from the developmental phase of adolescent egocentrism (Elkind, Reference Elkind1999; Schwartz, Maynard, & Uzelac, Reference Schwartz, Maynard and Uzelac2008). In addition, the observed decrease in Individual Regard may emerge from increased encounters with negative attitudes about and behaviors toward members of one's ethnic–racial group as youth leave the relative security of the home and typical neighborhood and encounter a broader range of individuals and opinions. For Black participants, that may take the form of increased encounters with ethnic–racial prejudice, overt discrimination, and microaggressions. For whites, this may take the form of increased awareness of the existence of negative opinions about white Americans and white privilege in light of white America's role in historic and present-day wrongs against people of color in US society.
Changes in social context likely play a role in developmental changes (decreases) in Ethnic Behavior as well; over the transition to adulthood, many individuals are moving out of the family home (in which ethnic traditions may be encouraged or enacted), to living on their own, in a college or work setting, or beginning to form families of their own. These contextual trends may account for the observed decrease in Ethnic Behaviors (as traditions may be less likely to be enacted outside of the family home) and increase in Community Regard, as youth come to have greater interactions with and focus on one's friends and ethnic–racial community as a source of engagement and support (Azmitia, Syed, & Radmacher, Reference Azmitia, Syed and Radmacher2013). Additional research is needed describing developmental trends in ERI development, replicating the current findings and testing whether they generalize to other contexts. Research should also aim to specifically test potential explanations underlying developmental changes in ERI. As noted later, the described developmental changes from adolescence through early adulthood may help us to interpret the nature and timing of our findings of ERI effects on adult cortisol.
Ethnic–racial differences in ERI
Even more distinct than developmental trends in ERI are the profound ethnic–racial differences we found in ERI levels. Black Americans had significantly higher levels of Individual Regard (ethnic–racial pride), Community Regard, Racial Centrality, and Ethnic Behavior than whites, with effect sizes on the order of 1 to 2 SDs higher for most ERI variables (see Figure 1). Thus, a strong ERI is a characteristic and notable cultural strength of the Black Americans in this sample; our data showed this strength to be important for regulating stress biology.
ERI and total (AUC) cortisol levels
Higher levels of Individual and Community Regard predicted higher total cortisol levels primarily among a group (Black Americans) that had previously evidenced a low or “hypocortisolemic” profile (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015), after controlling for the effects of racial discrimination and covariates. Our interpretation of these results presumes that low total daily cortisol is problematic, based on prior theory and evidence linking low cortisol (hypocortisolism) to a wide range of negative mental and physical health outcomes (Fries et al., Reference Fries, Hesse, Hellhammer and Hellhammer2005; Heim et al., Reference Heim, Ehlert and Helhammer2000). The higher cortisol associated with Individual and Community Regard for Black participants does not result in high levels of cortisol for Black participants, but rather an increase from low levels to levels that are moderate and more similar to those seen in the white participants in the sample (and to levels seen in black individuals with low levels of discrimination; see Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015). We anticipate that these more typical cortisol levels, in turn, will later be associated with more positive (and more equitable) health and human capital outcomes; these hypotheses remain to be tested in future research. Of note, the effect size of overall ERI associations with total cortisol levels are moderate to large, including .64 SD units across adolescence and young adulthood and .71 SD units for ERI in young adulthood.
The two ERI components found to be significantly related to total cortisol levels (Individual and Community Regard) include two of the elements described by Dickerson and Kemeny (Reference Dickerson and Kemeny2004) to be important for regulating HPA axis activity – they include positive social evaluations of one's ethnic–racial group (particularly Individual Regard), and feelings of belonging to and support from one's ethnic–racial community (particularly Community Regard). Our analyses controlled for levels of self-esteem, providing evidence that ERI was significantly associated with cortisol over and above the effects of general levels of positive self-regard – positive regard for one's ethnic identity and one's ethnic group were key to our results. The importance of positive regard for one's ERI (Individual Regard) and one's ethnic–racial community (Community Regard) in relation to cortisol in the current study aligns with reviews of the literature on the impact of ERI on other outcomes, in which ethnic–racial affirmation is most consistently found to be associated with positive outcomes for minority youth (Neblett et al., Reference Neblett, Rivas-Drake and Umaña-Taylor2012).
One function of a well-developed ERI may be to interrupt the cycle by which racial discrimination impacts negative thoughts and feelings about the self. As noted previously, perceptions of negative social evaluation can be translated into negative self-evaluations, which in turn can be experienced as feelings of shame which are potent influences on the HPA axis (Hittner & Adam, Reference Hittner and Adam2019). Pride in one's ethnic identity and perceived support from one's ethnic group may allow individuals to deflect negative social evaluations such as those associated with racial discrimination, attributing flaws to the actors who perpetrates the racial discrimination, rather than to the self. Thus, a positive ERI may help to reduce experiences of shame, replacing them with feelings of pride, and potentially of disdain or anger towards racial discrimination perpetrators. Further research should explore the emotional consequences of ERI, and the subsequent consequences of altered emotions for developmental and health outcomes. Thus, our results support the idea that positive self- and social evaluations and feelings of social belonging are particularly potent regulators of HPA axis activity.
ERI and diurnal cortisol slopes
Few significant results were found for another key indicator of diurnal cortisol rhythms, the diurnal cortisol slope, which has been previously associated with negative socio-emotional experience (Adam, Reference Adam, Dennis, Buss and Hastings2012; Adam & Kumari, Reference Adam and Kumari2009) and negative health outcomes (Adam et al., Reference Adam, Quinn, Tavernier, McQuillan, Dahlke and Gilbert2017). There was a trend for Community Regard to predict steeper slopes for both Black and white individuals, a pattern typically associated with lower perceived stress and improved health (Adam et al., Reference Adam, Quinn, Tavernier, McQuillan, Dahlke and Gilbert2017). By contrast, increased Ethnic Behavior predicted flatter cortisol slopes for whites; it is unclear what aspect of cultural practice for whites is driving this effect or whether particular ethnic subgroups within white Americans may show differing effects.
Prior research has found acute/recent discrimination experiences among individuals of color to predict flatter cortisol slopes (Zeiders et al., Reference Zeiders, Doane and Roosa2012), and concurrent ERI to predict steeper diurnal cortisol slopes (Zeiders et al., Reference Zeiders, Causadias and White2018). It is unclear why we did not find associations between ERI and cortisol slopes for young adults of color in our current analyses. It may be that ERI has initial acute effects on diurnal rhythms, through acute increases in morning cortisol levels and lower bedtime cortisol, but over longer time scales (of years and decades), only effects on average cortisol are seen. The latter conclusion would suggest that hair cortisol may be a useful indicator of the long-term impacts of racial discrimination and ERI on HPA axis functioning (Stalder et al., Reference Stalder, Steudte-Schmiedgen, Alexander, Klucken, Vater, Wichmann and Miller2017)
Protective versus promotive effects of ERI
It is worth noting that we found main effects of ERI, rather than significant interactions between ERI and racial discrimination in predicting cortisol, suggesting promotive rather than protective effects of ERI. That is, higher ERI was associated with high cortisol levels for individuals both high and low in perceived discrimination. Rather than only buffering individuals from the negative effects of discrimination, ERI appears to have positive effects regardless of racial discrimination level. This suggests that efforts to support or promote ERI would be helpful for all individuals (and particularly persons of color), not just those with high racial discrimination exposure, a finding which has implications for intervention, as discussed in more detail below.
Developmental timing of ERI effects on cortisol
Across our results, effects for ERI measured cumulatively across adolescence and adulthood, and for ERI measured just in young adulthood were the most robust; few results were found for adolescent ERI on its own. This runs counter to our prediction that adolescence would serve as a sensitive period for the impact of ERI on HPA axis activity, although given that the development of ERI, and of the brain extend into early adulthood, the issue here may not be with the theory but rather with our definition of what ages qualify as “adolescent”. Our findings regarding ongoing ethnic identity development during the 20s and the importance of young adult ERI for adult cortisol may add to the case of scholars arguing for a reclassification of adolescence as extending into the 20s, or at least recognizing that third decade of life is a dynamic developmental period that may itself continue to be a sensitive period for the effects of identity-related processes on adult biological, psychological well-being and health.
The developmental timing effects of our ERI subscales in relation to adult cortisol may be interpreted in light of our normative ERI trajectory analyses. The only significant effect we found in adolescence was for Individual Regard, which already had high levels in adolescence; Individual Regard peaked in adolescence and declined slightly over young adulthood. By contrast, Community Regard, which was associated with cortisol in young adulthood but not adolescence, was at low to moderate levels in adolescence and increased significantly for both Black and white individuals over the transition to young adulthood. Our Community Regard scale included items regarding social connectedness, which as noted above has been found to be a powerful regulator of stress biology; perhaps increasing contact with one's broader ethnic–racial community over the transition to adulthood plays a role in the stronger ERI effects on cortisol in young adulthood. In addition, ERI levels in young adulthood are measured more proximally in time to the cortisol measurement; it is possible that the closer temporal proximity of these measures contributed to more robust associations between young adult ERI and cortisol. Of note, however, in prior research, adolescent racial discrimination predicted adult cortisol more strongly than more proximal young adult racial discrimination measures (Adam et al., Reference Adam, Heissel, Zeiders, Richeson, Ross, Ehrlich and Eccles2015). The fact that cumulative and young adult ERI have strong promotive effects on adult cortisol is somewhat reassuring, suggesting that ERI developments through adolescence and beyond may help to reverse some of the prior negative effects of adolescent racial discrimination experiences.
Limitations and strengths
This study has a number of limitations. First, although the trajectory analyses involve a large sample of 685 youth, the sample is not representative of the general population, and reflect individuals from a particular geographic place and historical time period. The saliva sampling was conducted on a smaller subsample of 112 youth, who were different from the original larger sample in a number of ways, including having a smaller number of Black participants, higher parent education, and lower levels on some of the ERI variables. Perhaps a larger concern is that objective measurement of compliance with the timing of saliva sampling was lacking. Such monitoring is strongly recommended, as noncompliance may introduce both error and bias into cortisol measurement (Stalder et al., Reference Stalder, Kirschbaum, Kudielka, Adam, Pruessner, Wüst and Hellhammer2016). In contrast, compliance monitoring is particularly important for measures of the cortisol awakening response; our key analyses and results focused on total cortisol levels across the full waking day and diurnal cortisol slopes, which are slightly less subject to bias introduced by improper sample timing. Finally, although longitudinal in design, our results are correlational in nature, and we did not have baseline measures of cortisol available to include in our models.
This study also has a number of strengths. We gathered seven days of cortisol data, contributing to more reliable measurement of cortisol (Adam & Kumari, Reference Adam and Kumari2009). Most importantly, we used 20 years of longitudinal data. This ensured that both racial discrimination and ERI were measured prospectively, reducing recall biases. It allowed us to examine developmental trends in ERI from adolescence through young adulthood, a time period that is understudied in ERI research (Umaña-Taylor et al., Reference Umaña-Taylor, Quintana, Lee, Cross, Rivas-Drake, Schwartz and Seaton2014). It also afforded us the opportunity to compare the relative efficacy of ERI during several developmental periods, revealing early adulthood to be a particularly important time period for the promotive effects of ERI.
Future research directions
Our study suggests many priorities for future research. As noted above, more research is needed on developmental changes in ERI, and more importantly, the determinants of developmental changes in ERI across different developmental periods. Additional research is also needed on how broader macrostructural factors influence ERI and ERI development. Recent secular and historical shifts such as increases in openly racist and anti-immigrant discourse in social media and at the highest levels of politics in the United States; increased awareness of racial disparities revealed by the COVID-19 pandemic; police misconduct and brutality against Black individuals and the subsequent Black Lives Matter movement and protests; and recent attempts to combat anti-blackness are all likely to have impacted levels and forms of racial discrimination, ERI, and perceived and biological stress.
Although our analyses included 20 years of prospective longitudinal data on ERI from adolescence through young adulthood, it is also likely that developmental histories of race-related stress and ERI prior to adolescence may affect the regulation of adult stress biology. More research is also needed on how childhood and adolescent histories of racial discrimination and ERI affect later acute responses to race-related stress. Do individuals with high past levels of race-related stress respond more dramatically to current race-related stressors? Or, do past histories of racial discrimination and ERI modify stress biology in ways that buffer stress biology from the impact of current race-related stress, making the individual better-suited to navigate a stressful ethnic–racial environment? Evolutionary models such as the adaptive calibration model would hypothesize that past histories of racial discrimination and ERI become built into and modify individual stress biology in ways that can be helpful for negotiating current environments (Del Giudice, Ellis, & Shirtcliff, Reference Del Giudice, Ellis and Shirtcliff2011).
Our primary outcome variables for the current study were levels (and diurnal patterns) of cortisol across the day, with our findings suggesting more and stronger effects on total cortisol level than diurnal cortisol slopes. The associations between total cortisol levels and various aspects of health and developmental well-being should be explored in future analyses of these data. In addition, associations between ERI and other stress biomarkers would be of interest.
In addition, the emotional pathways by which ERI affects cortisol levels and other outcomes deserves attention. As noted above, we hypothesize that ERI, and particularly ERI affirmation, will reduce feelings of shame in response to racial discrimination experiences. More generally, however, higher individual and Community Regard are likely to promote increased feelings of pride and belonging and decreased feelings of sadness and loneliness.
Finally, a key priority for future research is to take a prospective longitudinal approach to the measurement of, and examining relations between, both ERI and cortisol. The ability to observe changes in cortisol emerging over time in relation to changes in ERI would provide a more compelling case for the impacts of ERI on cortisol. Even more convincing evidence, however, would be provided by the implementation of interventions to promote ERI, and observation of the impacts of those interventions on stress biology.
Implications for interventions
If research continues to support the promotive effects of ERI for the regulation of stress biology, an important next step would involve the development and testing of interventions designed to promote positive ERI development in adolescents or young adults. Previous evidence suggests that it is possible to promote at least some aspects of ERI exploration and attainment during adolescence (Umaña-Taylor, Douglass, Updegraff, & Marsiglia, Reference Umaña-Taylor, Douglass, Updegraff and Marsiglia2018a). Whether promotion of Individual and Community Regard are possible and in turn have positive effects on cortisol regulation remains to be seen, and the most efficacious timing of such an intervention also remains to be determined. Our study suggests the possibility that support of feelings of positive regard and belonging to one's ethnic group and community in adolescence and early adulthood may be important contributors to the positive regulation of stress biology.
Our finding that ERI appears to be promotive rather than protective against the impact of racial discrimination on cortisol also has implications for intervention design, suggesting that a universal ERI-promotion intervention (serving individuals at all levels of racial discrimination) may be most appropriate, rather than a targeted approach in which individuals are selected based on high levels of racial discrimination. Conversely, given that effects are significantly stronger for Black participants, ERI intervention efforts may have the largest effect for Black students. The extent to which other students of color would also benefit from an ERI promotion intervention remains to be seen. Pilot results from a universal ERI promotion intervention called the Identity Project suggest promising effects for a multi-ethnic sample of adolescents (Umaña-Taylor, Kornienko, Bayless, & Updegraff, Reference Umaña-Taylor, Kornienko, Bayless and Updegraff2018b; Umaña-Taylor et al., Reference Umaña-Taylor, Douglass, Updegraff and Marsiglia2018a). Adolescents in intervention condition showed significant increases in ERI exploration, as compared to the control condition (Umaña-Taylor et al., Reference Umaña-Taylor, Douglass, Updegraff and Marsiglia2018a). Youth with increased ERI exploration also showed greater subsequent ERI resolution, higher self-esteem, lower depression, and increased grades (Umaña-Taylor et al., Reference Umaña-Taylor, Kornienko, Bayless and Updegraff2018b).
Informed by these findings, our research team is currently implementing a randomized control trial of the Identity Project Intervention with high school freshman, called the BIO study (Biology, Identity and Opportunity study; funded by the Spencer Foundation). In the BIO study, we are examining whether random assignment to and participation in the Identity Project Intervention results in increases in multiple aspects of ERI, relative to participation in an attentional control condition. We will also examine whether experimentally manipulated increases in ERI are associated with better-regulated cortisol profiles, improved sleep, better executive functioning, and improvements in self-esteem, depression, and academic outcomes.
Conclusion
Overall, our research suggests that the presence of a strong ERI from adolescence through young adulthood is associated with better regulation of stress biology in adulthood, particularly in Black youth. In ongoing research, we are employing a randomized design of an ERI-promotive intervention to test the causal impact of ERI development on cortisol regulation as well as other aspects of development related to HPA axis functioning, such as academic, mental health, and physical health outcomes (Adam et al., Reference Adam, Quinn, Tavernier, McQuillan, Dahlke and Gilbert2017; Levy et al., Reference Levy, Heissel, Richeson and Adam2016). Ultimately, well-being will be best promoted by societal and intervention efforts to reduce youth exposure to the adverse social experience of racial discrimination, in addition to bolstering ERI. In the meantime, efforts aimed at supporting and promoting the development of a positive ERI may be a promising approach to improving cortisol regulation, particularly for youth of color. More broadly, this research provides evidence of the ongoing sensitivity of the HPA axis to both stressful and supportive social experience, in adolescence and into early adulthood, and the importance of studying racial discrimination as an adverse early experience, and ERI as a racial and ethnic strength promoting the positive regulation of stress biology and related outcomes.
Acknowledgement
The authors would like to thank Jacquelynne S. Eccles, Stephen C. Peck, Amanda B. Brodish, Oksana Malanchuk, Thomas E. Fuller-Rowell, Jennifer A. Richeson; Katherine B. Ehrlich, Margaret Kemeny, and Wendy Berry Mendes for their roles in conceptualization, design and data collection of the MADICS data utilized in this paper. Data from this study are archived at the Henry A. Murray Research Archive at Harvard University.
Financial Statement
This research was supported by multiple grants over the years. The first three waves of the study were funded by the MacArthur Network on Successful Adolescent Development in High Risk Settings (Chair: R. Jessor). Waves 4, 5 were funded by NICHD Grant #R01 HD33437 to Jacquelynne S. Eccles and Arnold J. Sameroff. Wave 6 was funded by Spencer Foundation Grant MG #200000275 to Tabbye Chavous and JSE. Waves 7, 8 were funded by NICHD Grant #R01 HD048970 to Jacquelynne S. Eccles and Stephen C. Peck and the Wave 8 biomarker data were funded by NIA Grant # RC2AG03678001 to JSE, SCP, EKA, Jennifer Richeson, Margaret Kemeny, and Wendy Berry-Mendes. Support was also provided from the Institute for Policy Research at Northwestern University to E.K.A., E.H., S.T. and S.C.V.
Conflicts of Interest
None.




