Aggressive–disruptive behavior and attention problems in the elementary school years are well-established predictors of a wide range of untoward outcomes in adolescence (Kellam et al., Reference Kellam, Brown, Poduska, Ialongo, Wang and Toyinbo2008; Petras et al., Reference Petras, Kellam, Brown, Muthén, Ialongo and Poduska2008; Schaeffer, Petas, Ialongo, Poduska, & Kellam, Reference Schaeffer, Petras, Ialongo, Poduska and Kellam2003), including the early onset and transition to heavy use of substances (Reboussin, Hubbard, & Ialongo, Reference Reboussin, Hubbard and Ialongo2007; Reboussin & Ialongo, Reference Reboussin and Ialongo2010). Within the context of two generations of randomized controlled trials of elementary school-based, universal preventive interventions, we have established the malleability of aggressive–disruptive behavior and attention problems in response to the interventions (Dolan et al., Reference Dolan, Kellam, Brown, Werthamer-Larsson, Rebok and Mayer1993; Ialongo et al., Reference Ialongo, Werthamer, Kellam, Brown, Wang and Lin1999). Moreover, follow-up through adolescence and early adulthood has yielded evidence of intervention impact on the survival to first use of substances (Kellam & Anthony, Reference Kellam and Anthony1998; Wang at al., Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) and the transition to heavy use, abuse, and dependence (Kellam et al., Reference Kellam, Brown, Poduska, Ialongo, Wang and Toyinbo2008). In the first-generation trial, the focus of the interventions was exclusively on improving teacher instructional and behavior management practices as the mechanism by which aggressive–disruptive behavior and attention problems would be reduced. Whereas in the second-generation trial, the focus was expanded to include parent as well as teacher instructional and behavior management skills as intervention targets. Thus, the second-generation trial featured a classroom-centered intervention (CC), wherein the focus was on improving the classroom teacher's instructional and behavior management practices, and a family–school partnership intervention (FSP), which centered on improving parent behavior management and instructional skills, along with fostering a strong parent–teacher partnership (Ialongo et al., Reference Ialongo, Werthamer, Kellam, Brown, Wang and Lin1999).
Here we examine whether effects of one of these two interventions on first cigarette smoked varied as a function of the genetic makeup of the participants. Rather than examining a small number of candidates genes, we rely on a polygenic composite, drawn from genome-wide association study results pertaining to tobacco use, composed of 12,000 + single nucleotide polymorphisms (SNPs; Rose et al., Reference Rose, Behm, Drgon, Johnson and Uhl2010; Uhl, Drgon, Johnson, Ramoni, et al., Reference Uhl, Drgon, Johnson, Ramoni, Behm and Rose2010).
The Theoretical Basis for the Interventions
Our conceptualization of normal and pathologic development and, in turn, the choice of our preventive interventions and their proximal and distal targets have been guided by the life course/social field framework (Ialongo et al., Reference Ialongo, Rogosch, Cicchetti, Toth, Buckley, Petras and Cicchetti2006; Kellam & Rebok, Reference Kellam, Rebok, McCord and Tremblay1992). This framework focuses on the measurement within epidemiologically defined populations of early maladaptive responses to social task demands that increase the risk of mental and substance abuse disorders over the life course. Central to life course/social field theory is the concept that individuals face specific social task demands in various social fields across the major periods of the life span (Ialongo et al., Reference Ialongo, Rogosch, Cicchetti, Toth, Buckley, Petras and Cicchetti2006). The social task demands the individual confronts are defined by individuals in each social field whom we have termed “natural raters.” The natural rater not only defines the tasks but also rates the individual's performance in that social field. Parents function as natural raters in the family, peers in the peer group, teachers in the classroom, and supervisors in the workplace (Ialongo et al., Reference Ialongo, Rogosch, Cicchetti, Toth, Buckley, Petras and Cicchetti2006; Kellam & Rebok, Reference Kellam, Rebok, McCord and Tremblay1992). This interactive process of demand and response is termed “social adaptation,” and the judgments of the individual's performance by the natural raters are termed “social adaptational status” (Ialongo et al., Reference Ialongo, Rogosch, Cicchetti, Toth, Buckley, Petras and Cicchetti2006; Kellam & Rebok, Reference Kellam, Rebok, McCord and Tremblay1992).
In line with the organizational approach to development (Cicchetti & Schneider-Rosen, Reference Cicchetti, Schneider-Rosen, Cicchetti and Schneider-Rosen1984), normal development is viewed within the life course/social fields framework as marked by the integration of earlier competencies into later modes of functioning, with the earlier competencies remaining accessible, ready to be activated and utilized during times of stress, crisis, novelty, and creativity. It follows then that early successful social adaptational status in the face of prominent developmental challenges tends to promote later adaptation as the individual traverses the life course and encounters new and different social task demands across the main social fields (Cicchetti & Schneider-Rosen, Reference Cicchetti, Schneider-Rosen, Cicchetti and Schneider-Rosen1984). This key developmental principle, along with a growing empirical literature, forms the basis for our focus on successful adaptation to the developmental challenges of early elementary school as a means of improving social adaptational status over the life course.
The integration of our life course/social fields perspective and Patterson and colleagues' (Granic & Patterson, Reference Granic and Patterson2006; Patterson, Reid, & Dishion, Reference Patterson, Reid and Dishion1992) model of the development of antisocial behavior and substance use provides the theoretical basis for the impact of the intervention components targeting the early antecedent risk behaviors of aggressive–disruptive behavior and attention problems and their distal correlates. According to Patterson et al., a major pathway to substance use, antisocial behavior, depression, poor academic achievement, and high risk sexual behavior in adolescence, begins in the toddler years, when parental success in teaching their child to interact within a normal range of compliance and aversive behavior is a prerequisite for the child's development of social survival skills (Granic & Patterson, Reference Granic and Patterson2006; Patterson et al., Reference Patterson, Reid and Dishion1992). Alternatively, the parents' failure to effectively punish coercive behavior during these formative years and to teach reasonable levels of compliance comprises the first step in a process that serves to “train” the child to become progressively more coercive and antisocial. In the classroom setting, such children prove difficult for teachers or peers to “teach” appropriate forms of social interaction and problem solving. Moreover, their coercive style may be further reinforced in the presence of inconsistent and coercive teacher disciplinary practices. Ultimately, the coercive child is rejected by parents, teachers, and well-adjusted peers, which results in the child's failure to develop the necessary academic, social, and occupational “survival” skills that presage successful adaptation in adolescence and beyond. That is, the opportunities to learn these social survival skills thorough interaction with teachers, parents, and mainstream peers are greatly reduced due to the rejection.
Patterson and colleagues (Granic & Patterson, Reference Granic and Patterson2006; Patterson et al., Reference Patterson, Reid and Dishion1992) further argue that the lack of adequate monitoring by parents in early adolescence, and rejection by teachers and mainstream peers, precipitates “drift” into a deviant peer group, wherein a wide array of antisocial and delinquent behavior, including alcohol and drug use, may be reinforced, along with rejection of main stream norms and mores (Brook, Brook, Zhang, & Cohen, Reference Brook, Brook, Zhang and Cohen2009; Doherty, Green, Reisinger, & Ensminger, Reference Doherty, Green, Reisinger and Ensminger2008). The deviant peer group may also serve as a further training ground for coercive behavior, resulting in not only its maintenance but also its escalation. Concomitant with the drift into a deviant peer group, the opportunities for obtaining positive reinforcement from mainstream natural raters, such as parents, teachers, and well-adjusted peers, are significantly reduced. In turn, the coercive youth will be more likely to use substances as a means of obtaining reinforcement and negating the reductions in reinforcement dispensed by mainstream natural raters. In addition, the lack of positive reinforcement received from mainstream natural raters may lead to decrements in psychological well-being (La Greca & Moore Harrison, 2007; Kim, Capaldi, Pears, Kerr, & Owen, Reference Kim, Capaldi, Pears, Kerr and Owen2009), which the youth seeks to alleviate through substance use (Shivola et al., Reference Shivola, Rose, Dick, Pulkkinen, Martunnen and Kaprio2008; Chen, Anthony, & Crum, Reference Chen, Anthony and Crum1999).
In keeping with our life course/social fields perspective and its integration with Patterson and colleagues (Granic & Patterson, Reference Granic and Patterson2006; Patterson et al., Reference Patterson, Reid and Dishion1992), we hypothesized that our universal preventive interventions would reduce the aggressive–disruptive behavior and attention problems and their distal correlates via improved teachers' and parents' behavior management practices, respectively, which should then result in a reduction of the early antecedent risk behaviors of aggressive–disruptive behavior and attention problems. As a result of the reduction in aggressive–disruptive behavior and attention problems, we reasoned there should be fewer opportunities for the youth to learn inappropriate behavior through modeling of their classmates' aggressive behavior. The youth should then be at decreased risk of being rejected by parents/caregivers, teachers, and peers, and in turn, be less likely to drift into a deviant peer group, where substance use and antisocial behavior may be reinforced.
Variation in Developmental Course and Intervention Response
Variation in intervention response is often the rule rather than the exception in evaluations of universal preventive interventions. Consistent with the elaboration of the prevention research cycle in the Institute of Medicine's 1994 (Mrzazek & Heagarty, 1994) report on prevention, understanding the source of such variation is critical to improving upon extant preventive interventions and informing the next generation of interventions. A focus on understanding variation and malleability in developmental paths is also in line with the concepts of resilience and turning points as elaborated in the organizational approach to development (Cicchetti & Schneider-Rosen, Reference Cicchetti, Schneider-Rosen, Cicchetti and Schneider-Rosen1984).
Over the last 25 years, we have found substantial evidence of variation in intervention impact and developmental outcomes in our first- and second-generation universal preventive intervention trials. The sources of variation have included the characteristics of the participant and their family, school, peer group, and neighborhood environments (e.g., Ialongo, Poduska, Werthamer, & Kelam, Reference Ialongo, Poduska, Werthamer and Kellam2001; Muthen & Asparouv, Reference Muthen, Asparouhov, Fitzmaurice, Davidian, Verbeke and Molenberghs2008; Petras, Masyn, Buckley, Ialongo, & Kellam, Reference Petras, Masyn, Buckley, Ialongo and Kellam2011; Schaeffer et al., Reference Schaeffer, Petras, Ialongo, Poduska and Kellam2003). In general, consistent with Belsky and colleagues' (Belsky et al., Reference Belsky, Bakermans-Kranenburg and van IJzendoorn2007; Belsky & Pleuss, Reference Belsky and Pluess2009, Reference Belsky and Pluess2013; Ellis et al., Reference Ellis, Boyce, Belsky, Bakermans-Kranenburg and van IJzendoorn2011) elaboration of the differential susceptibility hypothesis, the greatest impact of the interventions has been found for those children who entered first grade with mild to moderate elevations in the constellation of antecedent risk behaviors targeted by our interventions. The differential susceptibility hypothesis/framework emphasizes plasticity of response to environmental variation, both positive and negative, as opposed to the diathesis–stress framework and its exclusive focus on risk in terms of the characteristics of the individual and the environment.
Given that quantitative genetic studies point to the heritability of the proximal (poor academic performance, aggressive–coercive, and inattentive/impulsive/hyperactive behaviors) and distal targets of our interventions (e.g., substance use initiation, heavy use, and abuse/dependence; e.g., Kendler & Prescott, Reference Kendler and Prescott2008) and their putative moderators and mediators (e.g., Horowitz & Neiderhiser, Reference Horwitz and Neiderhiser2011; Jaffee & Price, Reference Jaffee and Price2007; Knafo & Jaffee, Reference Knafo and Jaffee2013), we also have begun to explore the role of genetics as a source of variation in our intervention and developmental outcomes.
Returning to Patterson and colleagues' model of the development of antisocial behavior and substance abuse/dependence, one mechanism by which genetic factors may contribute to variation in intervention outcomes is through their influence on child characteristics, such as oppositional (Stringaris et al., Reference Stringaris, Zavos, Leibenluft, Maughan and Eley2012), inattentive, impulsive, and hyperactive behaviors (Beach, Brody, Lei, & Philbert, Reference Beach, Brody, Lei and Philbert2010; Kan et al., Reference Kan, Dolan, Nivard, Middeldorp, van Beijsterveldt and Willemsen2013). These behaviors may serve to increase the risk of inconsistent and coercive discipline on the part of parents and teachers and in turn accelerate the coercive cycle described by Patterson and colleagues, resulting in drift into a deviant peer group where antisocial behavior and substance use may be reinforced. Such a mechanism would be in keeping with an evocative gene–environment correlation as described in Narusyte et al. (Reference Narusyte, Neiderhiser, Andershed, D'Onofrio, Reiss and Spotts2011) and the concept of general/common genetic influences (e.g., Haberstick et al., Reference Haberstick, Zeiger, Corley, Hopfer, Stallings and Rhee2011; Kendler, Prescott, Myers, & Neale, Reference Kendler, Prescott, Myers and Neale2003). Regarding the latter, genetic influences on substance use/abuse may be mediated through child evocative behaviors. It may also be the case, as Beach et al. (Reference Beach, Brody, Lei and Philbert2010) point out, that the child evocative behaviors implicated in the Patterson et al.'s coercive cycle share the same genetic architecture underlying the drug craving seen in adolescence and adulthood.
Besides general/common genetic influences, there is also evidence of unique genetic contributions to substance use as reflected in Kendler et al. (Reference Kendler, Prescott, Myers and Neale2003) and more recently in Haberstick et al. (Reference Haberstick, Zeiger, Corley, Hopfer, Stallings and Rhee2011). In the latter, variation in perceived positive and negative effects of tobacco, alcohol, and marijuana use was found to be heritable in adolescences and young adults. Such unique genetic influences may also contribute to variation in prevention intervention response. For example, whether one transitions from experimentation with tobacco to heavy use may be a function of genetically influenced positive and negative responses to its use.
The Current Study
Preventive intervention trials featuring randomized designs provide an optimal context to study Gene × Intervention interactions, given the inherent balance across conditions or environments. The literature on such interactions is in its infancy, with Bakermans-Kranenburg, van IJzendoorn, Pijlman, Mesman, and Juffer (Reference Bakermans-Kranenburg, van IJzendoorn, Pijlman, Mesman and Juffer2008) the first to publish on a Gene × Intervention effect, followed by Brody and colleagues (e.g., Beach et al., Reference Beach, Brody, Lei and Philbert2010; Brody, Beach, Philibert, Chen, & Murry, Reference Brody, Beach, Philibert, Chen and Murray2009) and Kegel, Bus, and van IJzendoorn (Reference Kegel, Bus and van IJzendoorn2011).
In Bakermans-Kranenburg et al. (Reference Bakermans-Kranenburg, van IJzendoorn, Pijlman, Mesman and Juffer2008), Beach et al. (Reference Beach, Brody, Lei and Philbert2010), and Kegel et al. (Reference Kegel, Bus and van IJzendoorn2011), the seven-repeat allele of the dopamine D4 receptor gene was associated with greater intervention response. The focus in the Bakermans-Kranenburg et al. study was externalizing behavior in childhood, whereas the outcomes in Beach et al. and Kegel et al. were substance use in adolescence and early literacy, respectively. Brody and colleagues (Beach et al., Reference Beach, Brody, Lei and Philbert2010; Brody et al., Reference Brody, Beach, Philibert, Chen and Murray2009) have demonstrated Gene × Intervention interactions for a number of other outcomes and genetic variants within the context of their randomized trial of the Strong African American Families program (Brody et al., Reference Brody, Murry, Gerrard, Gibbons, Molgaard and McNair2004). The Strong African American Families program focuses on rural African American adolescents and their parents living in economically distressed areas. Like Bakersman-Kranenburg et al. (2008) and Kegel et al. (Reference Kegel, Bus and van IJzendoorn2011), Brody and colleagues' findings have been generally consistent with the differential susceptibility hypothesis (Belsky et al., Reference Belsky, Bakermans-Kranenburg and van IJzendoorn2007; Belsky & Pleuss, Reference Belsky and Pluess2009, Reference Belsky and Pluess2013; Brody et al., Reference Brody, Chen, Yu, Beach, Kogan and Simons2012; Ellis et al., Reference Ellis, Boyce, Belsky, Bakermans-Kranenburg and van IJzendoorn2011), such that the greatest intervention impact has been found for those participants with genetic variants that have been previously associated with a number of adverse outcomes in the absence of intervention or a naturally enriched environment.
In the current study, we seek to add this nascent literature. More specifically, we extend the findings of Wang et al. (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) by examining genetics as a source of variation in our CC intervention's effect on age of first cigarette smoked through late adolescence into early adulthood. There is substantial evidence of genetic influences on smoking initiation, the transition to daily and heavy smoking, tobacco dependence, and quit success. This evidence stems from both the quantitative (Horimoto et al., Reference Horimoto, Oliveira, Giolo, Soler, Andrade and Krieger2012; Lessov et al., Reference Lessov, Martin, Statham, Todorov, Slutske and Bucholz2004; Sullivan & Kendler, Reference Sullivan and Kendler1999) and the molecular genetic literatures (Belsky et al., Reference Belsky, Moffitt, Baker, Biddle, Evans and Harrington2013; Liu et al., Reference Liu, Tozzi, Waterworth, Pillai, Muglia and Midleton2010; MacQueen et al., Reference MacQueen, Heckman, Blank, Van Rensburg, Park and Drobes2014; Uhl et al., Reference Uhl, Walther, Musci, Fisher, Anthony and Storr2014; Wilkinson et al., Reference Wilkinson, Bondy, Wu, Wang, Dong and D'Amelio2012). As noted, there is also evidence of both common and unique genetic influences. The former are particularly prominent in terms of experimentation across common substances, such as tobacco, alcohol, and marijuana (Ducci et al., Reference Ducci, Kaakinen, Pouta, Hartikainen, Veijola and Isohanni2011; Wilkinson et al., Reference Wilkinson, Bondy, Wu, Wang, Dong and D'Amelio2012), and appear to be in part mediated via personality characteristics such as novelty seeking and risk taking. Ducci et al. (Reference Ducci, Kaakinen, Pouta, Hartikainen, Veijola and Isohanni2011) found that the TTC12-ANKK1-DRD2s predicted smoking behavior in adolescence but was not associated with the transition to heavy smoking and persistence in adulthood. The effect of the TTC12-ANKK1-DRD2s on smoking in adolescence was in part mediated via “personality characteristics promoting drug-seeking behavior” (Ducci et al., Reference Ducci, Kaakinen, Pouta, Hartikainen, Veijola and Isohanni2011, p. 50). In a sample of Mexican youth, Wilkinson et al. (Reference Wilkinson, Bondy, Wu, Wang, Dong and D'Amelio2012) reported the OPRM1, SNAP25, and HTR1B genes were associated with experimentation among “committed never smokers,” whereas the HTR2A, DRD2, and SLC6A genes were predictive of experimentation in what they termed “susceptible” youth.
Evidence from the molecular genetic literature highlights the salience of nicotinic acetylcholine receptor (nAChRs) genes in regular smoking, the transition from regular to heavy smoking, nicotine dependence, and quit success (Beirut, Reference Bierut2009; Belsky et al., Reference Belsky, Moffitt, Baker, Biddle, Evans and Harrington2013; MacQueen et al., Reference MacQueen, Heckman, Blank, Van Rensburg, Park and Drobes2014; Uhl, Drgon, Johnson, Walther, et al., Reference Uhl, Drgon, Johnson, Ramoni, Behm and Rose2010; Wilkinson et al., Reference Wilkinson, Bondy, Wu, Wang, Dong and D'Amelio2012). Nicotine serves a surrogate for acetylcholine in neuronal transmission. The addictive properties of nicotine are thought to be a function of the connections between nAChRs and the mesocorticolimbic dopamine pathways. The latter system has been postulated as the “reward” center in the brain, although this is likely an oversimplification. Current thinking suggests that the addictive properties of nicotine are likely a product of the complex transaction between the glutamate, dopamine, and GABA systems (Corrigal, Coen, & Adamson, Reference Corrigall, Coen and Adamson1994; Laviolette & Van de Kooy, Reference Laviolette and Van de Kooy2004; Tapper, Nashmi, & Lester, Reference Tapper, Nashmi, Lester, Madras, Colvis, Pollock, Rutter, Shurtleff and Zastrow2006). This view seems to be consistent with evidence from genome wide-association studies that have implicated multiple loci across the genome in terms of smoking behavior, particularly loci on chromosomes 15 (e.g., Gabrielsen, Romundstad, Langhammer, Krokan, & Skorpen, Reference Gabrielsen, Romundstad, Langhammer, Krokan and Skorpen2013; Liu et al., Reference Liu, Tozzi, Waterworth, Pillai, Muglia and Midleton2010), 19 (e.g., Bloom et al., Reference Bloom, Baker, Chen, Breslau, Hatsukamu and Bierut2014), 11 (e.g., Tobacco and Genetics Consortium, 2010), and 9 (e.g., Tobacco and Genetics Consortium, 2010). Moreover, studies of smoking behavior utilizing polygenic scores, as opposed to single candidate genes or SNPS, have yielded substantially larger effect sizes in predicting smoking behaviors (e.g., Belsky et al., Reference Belsky, Moffitt, Baker, Biddle, Evans and Harrington2013; Uhl, Drgon, Johnson, Ramoni, et al., Reference Uhl, Drgon, Johnson, Ramoni, Behm and Rose2010). Along these lines, Uhl, Walther, Bem, and Rose (Reference Uhl, Walther, Bem and Rose2011) provides evidence of the role of TRPA1 (transient receptor potential, TRP) variants on chromosome 8 with respect to preference for mentholated cigarettes. The TRPA1 gene encodes for the TRPA1 ion channel that is one of a number TRP channels that facilitates neuronal transmission among the nociceptive primary afferent nerve fibers in the lung (Uhl et al., Reference Uhl, Walther, Bem and Rose2011). Menthol is postulated to mitigate the experience of the noxious stimuli associated with smoking via its activation of the TRPA1 channel and in turn the innervation of the nociceptive nerves. Uhl et al. (Reference Uhl, Walther, Bem and Rose2011) reason that the less one experiences the noxious effects of smoking, the more they are likely to smoke and the less likely they are to quit.
Returning to the current study, we use a polygenic score rather than a candidate gene approach. The advantage of the former is that it reflects the considerable evidence of the multiple loci involved in smoking as detailed above and the prevailing belief in the molecular genetics literature that human behavior is influenced by multiple genes as opposed to a single gene (see Duncan, Pollastri, & Smoller, Reference Duncan, Pollastri and Smoller2014). The polygenic score we employed was found by Uhl, Drgon, Johnson, Ramoni, et al. (Reference Uhl, Drgon, Johnson, Ramoni, Behm and Rose2010) to predict smoking cessation in adults and the frequency of tobacco, marijuana, and alcohol use in the aggregate from early adolescence into young adulthood (Uhl et al., Reference Uhl, Walther, Musci, Fisher, Anthony and Storr2014). A higher score was associated with a greater likelihood of quit success (Uhl, Drgon, Johnson, Walther, et al., Reference Uhl, Drgon, Johnson, Ramoni, Behm and Rose2010) and lower use of tobacco, marijuana, and alcohol, collectively, in adolescence and young adulthood (Uhl et al., Reference Uhl, Walther, Musci, Fisher, Anthony and Storr2014). As noted, the basis for the polygenic score was genome-wide association data from participants in three adult smoking cessation randomized trials (Uhl, Drgon, Johnson, Ramoni, et al., Reference Uhl, Drgon, Johnson, Ramoni, Behm and Rose2010). Twelve thousands SNPs were included in the score based on their being associated at a nominal level of significance (p < .01) with quit success in the three trials.
As to the rationale for using Uhl's polygenic quit success score in the context of an analysis focused on first use, Wang et al. (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) reported that greater the number of one's friends who smoked in the middle school years, the earlier the age that one reported being offered a cigarette and accepting the offer. In light of Okoli, Kelly, and Hahn (Reference Okoli, Kelly and Hahn2007)'s review of the literature on secondhand smoke and nicotine exposure, it seems reasonable then to postulate that frequent and prolonged exposure to cigarette smoke may increase the likelihood of inhaling secondary smoke and experiencing the reinforcing effects of nicotine. Assuming that the quit success score is at least in part an index of one's sensitivity to the addictive properties of nicotine, the greater the sensitivity to nicotine, the more likely one would be to accept the offer to smoke after extended exposure. Our focus on age of first use is supported by the data suggesting that the earlier one starts smoking, the less likely one is to successfully quit smoking over the life course (Breslau, Fenn, & Peterson, Reference Breslau, Fenn and Peterson1993; Breslau & Peterson, Reference Breslau and Peterson1996; D'Avanzo, Vecchia, & Negri, Reference D'Avanzo, La Vecchia and Negri1994).
Consistent with Bakersman-Kranenburg et al. (2008), Kegel et al. (Reference Kegel, Bus and van IJzendoorn2011), and Brody and colleagues' findings (Beach et al., Reference Beach, Brody, Lei and Philbert2010; Brody et al., Reference Brody, Beach, Philibert, Chen and Murray2009), one might expect that the greatest intervention impact will be seen in those participants with a lower polygenic score, which would putatively reflect a higher sensitivity to the reinforcing characteristics of nicotine. However, consistent with Belsky and colleagues' concept of “for better and worse” (Belsky et al., Reference Belsky, Bakermans-Kranenburg and van IJzendoorn2007; Belsky & Pluess, Reference Belsky and Pluess2009, Reference Belsky and Pluess2013; Ellis et al., Reference Ellis, Boyce, Belsky, Bakermans-Kranenburg and van IJzendoorn2011), it may be just as likely that potentially advantageous genetic variants are optimally realized in the context of an enriched environment, such as one associated with a preventive intervention. Thus, in our case, an alternative hypothesis is that we will see greater intervention impact among those with higher polygenic scores, or lower sensitivity to the reinforcing aspects of nicotine exposure.
As noted, we only focus here on our CC intervention, because Wang et al. (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) did not find a significant effect for the FSP intervention on age of first cigarette smoked through late adolescence, although the FSP effect was in the expected direction. Of note, Storr et al. (Reference Storr, Ialongo, Kellam and Anthony2002) and Furr-Holden, Ialongo, Anthony, Petras, and Kellam (Reference Furr-Holden, Ialongo, Anthony, Petras and Kellam2004) did find that the FSP had a beneficial effect on the age of first use and ever use in early adolescence, respectively. We reasoned in the Wang et al. (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) studies that the lack of significant effects for the FSP intervention on age of first use through late adolescence may reflect that overall the CC intervention had a greater impact on the proximal targets of poor academic performance and the constellation of aggressive–coercive behavior and attention/impulsivity/hyperactivity problems. We provide a more elaborate treatment of this issue in the Discussion Section.
Method
Participants
The primary data from this study come from a longitudinal randomized controlled trial testing the impact of the CC and FSP interventions relative to a control condition. A detailed description of the participants and design is provided elsewhere (Ialongo et al., Reference Ialongo, Werthamer, Kellam, Brown, Wang and Lin1999). Data collection began in 1993 with 678 first graders and their caregivers. The evaluation battery consisted of structured teacher, parent, and child interviews. A randomized block design was employed with schools serving as the blocking factor. Children and teachers were randomly assigned to classroom and then classrooms were randomly assigned to intervention condition, with each of the three conditions being represented in each of the nine participating schools. The interventions were provided over the first-grade year only, following a pretest in the early fall. Data for the present study only included those individuals in the CC intervention and the control group.
Of the original 678 participants, 53.2% were male, 86.8% were African American, and 13.2% were Caucasian. In addition, 63.4% of the participants qualified for free or reduced lunch, a proxy for low socioeconomic status (Ensminger et al., Reference Ensminger, Forrest, Riley, Kang, Green and Starfield2000). As for the racial breakdown by design, 188 African Americans were in the control condition, while 201 African Americans were in the CC intervention condition, and 196 African Americans were in the FSP intervention condition. In addition, 31 Caucasians were in the control condition, and 28 Caucasian and 33 Caucasian participants were in the CC intervention and FSP intervention conditions, respectively. The participants' ages at the start of first grade ranged from 5.3 years to 7.7 years (mean = 6.2, SD = 0.34). Assessments were carried out in the fall of Grade 1, with annual follow-up assessments in the spring of Grades 6 through 12. Genetic samples were collected shortly after high school. Written informed consent was obtained from each participant, and the institutional review board approved the study. For additional information on the design of the trial, see Bradshaw, Zmuda, Kellam, and Ialongo (2009) and Ialongo et al. (Reference Ialongo, Werthamer, Kellam, Brown, Wang and Lin1999, Reference Ialongo, Poduska, Werthamer and Kellam2001).
Chi-square tests revealed no significant differences in terms of ethnicity (χ2 = 4.974, p = .083, free or reduced lunch status (χ2 = 2.126, p = .163), and design status (χ2 = 1.145, p = .766) between the 556 who provided both phenotypic and genetic data and those who did not; and t tests revealed no differences between these groups in terms of age at entrance to first grade. However, those included in this study were more likely to be female compared to those not included in this study (χ2 = 7.473, p = .007).
Intervention
The CC intervention was designed to reduce the early risk behaviors of poor achievement and aggressive behavior through enhancements to the curriculum, improvements in teacher instructional and classroom behavior management practices, and specific strategies for children not performing adequately (Ialongo et al., Reference Ialongo, Werthamer, Kellam, Brown, Wang and Lin1999). Each intervention classroom was divided into three heterogeneous groups, which provided the underlying structure for the curricular and the behavioral components of the intervention. In addition, the intervention program enhanced the Baltimore City Public School curriculum in language arts and mathematics by adding material to increase critical thinking, composition, and comprehension skills (Petras, Masyn, & Ialongo, Reference Petras, Masyn and Ialongo2011). The primary behavior management component was a behaviorally focused classroom management program called the Good Behavior Game, which in previous trials demonstrated a beneficial impact on student behavior (Barrish, Saunders, & Wolf, Reference Barrish, Saunders and Wolf1969; Kellam et al., Reference Kellam, Brown, Poduska, Ialongo, Wang and Toyinbo2008). The Good Behavior Game is a whole-class strategy that aims to decrease disruptive behaviors by assigning children to teams and only allowing the teams that do not exceed a specified criterion of precisely defined off-task, disruptive, and aggressive behaviors to “win.”
Measures
Genotype scores
Using Affymetrix 6.0 genotype data that passed overall quality control metrics for each participant, we assessed alleles at the 12,058 SNPs that comprised the previously described v1.0 quit success score. The SNP list, including their rs SNP designator, chromosome, base pair, allele associated with abstinence, and weight used in contributing to the v1.0 score are available from the authors upon request or can be seen in the supplemental information from Uhl et al. (Reference Uhl, Walther, Musci, Fisher, Anthony and Storr2014). Briefly, these SNPs were selected from at least one of three initial smoking cessation success clinical trial samples had identified nominally high-significant (p < .01) differences between successful and unsuccessful quitters, weighted based on strength and replicability of the associations in these studies as described, and included in the analysis as a continuous variable with a higher score indicating greater success in smoking cessation. Scores were standardized for ease of interpretation.
First tobacco use
Participants were asked yearly “Have you ever smoked tobacco, even just a puff?” The possible responses were “yes” or “no.” The earliest age when the participant answered “yes” to this question was used to indicate the age when tobacco use was initiated, varying from age 6 to age 16. As reported in previous studies (e.g., Audrain-McGovern, Lerman, Wileyto, Rodriguez, & Shields, Reference Audrain-McGovern, Lerman, Wileyto, Rodriguez and Shields2004; Johnston, O'Malley, & Bachman, Reference Johnston, O'Malley and Bachman1995), a relatively large percentage of adolescent smokers are only beginner smokers and have never smoked a “full” cigarette. Therefore, use of this question allowed us to examine the very earliest stage of nicotine involvement, which commonly happens during youth or adolescence.
Population stratification
When exploring genetic associations, population stratification, or genetic differences between subpopulations, it is important to identify and control for so that any significant associations found is not due to ancestry. Previous studies have emphasized the importance of this control, particularly in admixed populations (e.g., African American; Montana & Pritchard, Reference Montana and Pritchard2004; Sankararaman, Sridhar, & Halperin, Reference Sankararaman, Sridhar and Halperin2008). The process through which we created the variables to control for population stratification was multidimensional scaling (MDS), completed in PLINK. This process extracts, from genome-wide SNP data, clusters of individuals based on their estimated identity by descent. Subjects are assigned a score on each of these clusters representing their membership in a given population cluster. To reduce the computational intensity, we selected a set of one million SNPs randomly across the genome to test for the presence of stratification using the MDS approach. Although these were not a priori identified ancestry information markers, it has been shown that “randomly” selected SNPs perform equally well (Pritchard & Rosenberg, Reference Pritchard and Rosenberg1999). The results of the MDS allowed for the use of one accounted for a majority of the variance in population stratification.
Measure of preintervention aggression
The Teacher Observation of Classroom Adaptation—Revised was used to assess the participants' aggressive behaviors at baseline (i.e., fall of Grade 1 prior to randomization; Werthamer-Larsson, Kellam, & Wheeler, Reference Werthamer-Larsson, Kellam and Wheeler1991). This measure included items such as, “harms or hurts others physically” and “starts fights with classmates to assess aggression.” Teachers rated student behavior on a 6-point Likert scale from almost never to always. Previous research on the revised Teacher Observation of Classroom Adaptation has demonstrated a high level of predictive validity (Petras, Chilcoat, Leaf, Ialongo, & Kellam, Reference Petras, Chilcoat, Leaf, Ialongo and Kellam2004; Petras, Masyn, & Ialongo, Reference Petras, Masyn, Buckley, Ialongo and Kellam2011). See Petras, Masyn, and Ialongo (Reference Petras, Masyn, Buckley, Ialongo and Kellam2011) for additional information on the reliability and validity of these measures.
Analysis
A discrete-time survival analysis (DTSA) using Mplus version 7.1 (Muthén & Muthén, Reference Muthén and Muthén1998–2013) was performed to explore longitudinal risk of tobacco initial use (Muthén & Masyn, Reference Muthén and Masyn2005). DTSA is a specific type of survival analysis that models the timing of events, specifically when events are measured in discrete-time or grouped-time intervals (Masyn, Reference Masyn, Sloboda and Petras2014). This model specification allows for the inclusion of time-varying and time-invariant predictors. The event of interest for this particular analysis is defined for each participant as tobacco use initiation, and the survival time is defined as the time elapsed from age to the first cigarette smoked. The time scale was recorded in discrete-time intervals (age) so although the time-to-event process may actually be more continuous in nature, the data limitations required the process be modeled using DTSA. Fall of first-grade aggression and the population stratification variable were grand mean centered to ease in the interpretation of the interaction. In order to explore the moderation of one covariate effect by another, an interaction term is included as a predictor in the model. The interaction effect can be then decomposed by displaying hazard curves for differing levels of the covariates included in the interaction (Masyn, Reference Masyn, Sloboda and Petras2014). School/grade/section was included to account for cluster of students in classrooms. This technique, using the clustering command in Mplus, uses a sandwich procedure to calculate robust errors (Muthén & Muthén, Reference Muthén and Muthén1998–2013).
Missing data
Using full information maximum likelihood estimation, Mplus (Muthén & Muthén, Reference Muthén and Muthén1998–2013) assumes that the data were missing at random. This technique adjusts the estimates of the parameters for attrition. Full information maximum likelihood is considered the appropriate method for handing data missing at random (Little & Rubin, Reference Little and Rubin2002; Muthén & Shedden, Reference Muthén and Shedden1999; Shafer & Graham, Reference Schafer and Graham2002).
Results
Descriptive and univariate statistics
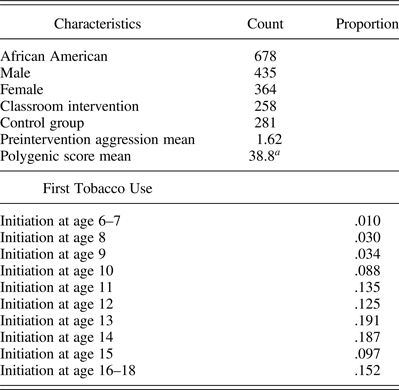
Proportions of first use at each age can be seen in Table 1. The proportion of individuals initiating began at only 1% of the population at the first time point. Subsequently, there was an increase in first-use rates across the time period, peaking at age 13 and age 14, with around 20% of the remaining sample initiating at each age, respectively. At the last time point, 15% of the remaining sample reported their first use. A majority of the participants were African American, with about half being in the intervention group and half in the control group. Mean scores for fall teacher-rated aggression and the polygenic score can also be seen in Table 1.
Table 1. Participant characteristics

aIndicates prestandardization mean.
Survival analysis for tobacco first use
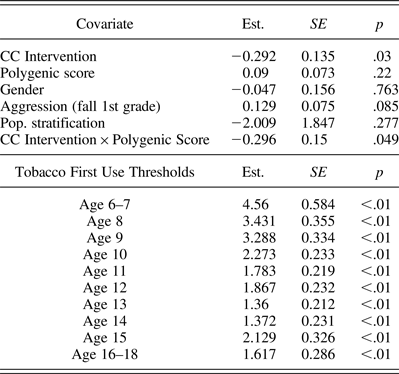
A DTSA model was ran using Mplus version 7.1 (Muthén and Muthén, Reference Muthén and Muthén1998–2013). The proportional hazard assumption was not violated for the polygenic score (–2 log likelihood [–2LL] = 7.42, df = 9, p = .59), which suggest that the relationship between the polygenic score and first tobacco use remained constant across grades. Similarly, the proportional hazard assumption was not violated for the population stratification variable (–2LL = 8.46, df = 9, p = .49). Intervention was a significant predictor of survival to first tobacco use (est. = –0.292, SE = 0.135, OR = 0.75, p = .03), suggesting that those who received the intervention had a decreased risk of first tobacco use. The polygenic score was not a significant predictor of risk of first tobacco use (est. = –0.090, SE = 0.073, p = .220).
With respect to the covariates included in the model, the relationship between preintervention aggression levels and age of first use approached significance (est. = 0.129, SE = 0.075, p = .085). However, none of the remaining covariates approached significance, including the population stratification variable (est. = –2.009, SE = 1.847, p = .277) or gender (est. = –0.047, SE = 0.156, p = .763). For regression estimates of all covariates, see Table 2.
Table 2. Model results for the final survival analysis model
Note: Log likelihood = –624.066, number of parameters = 16.
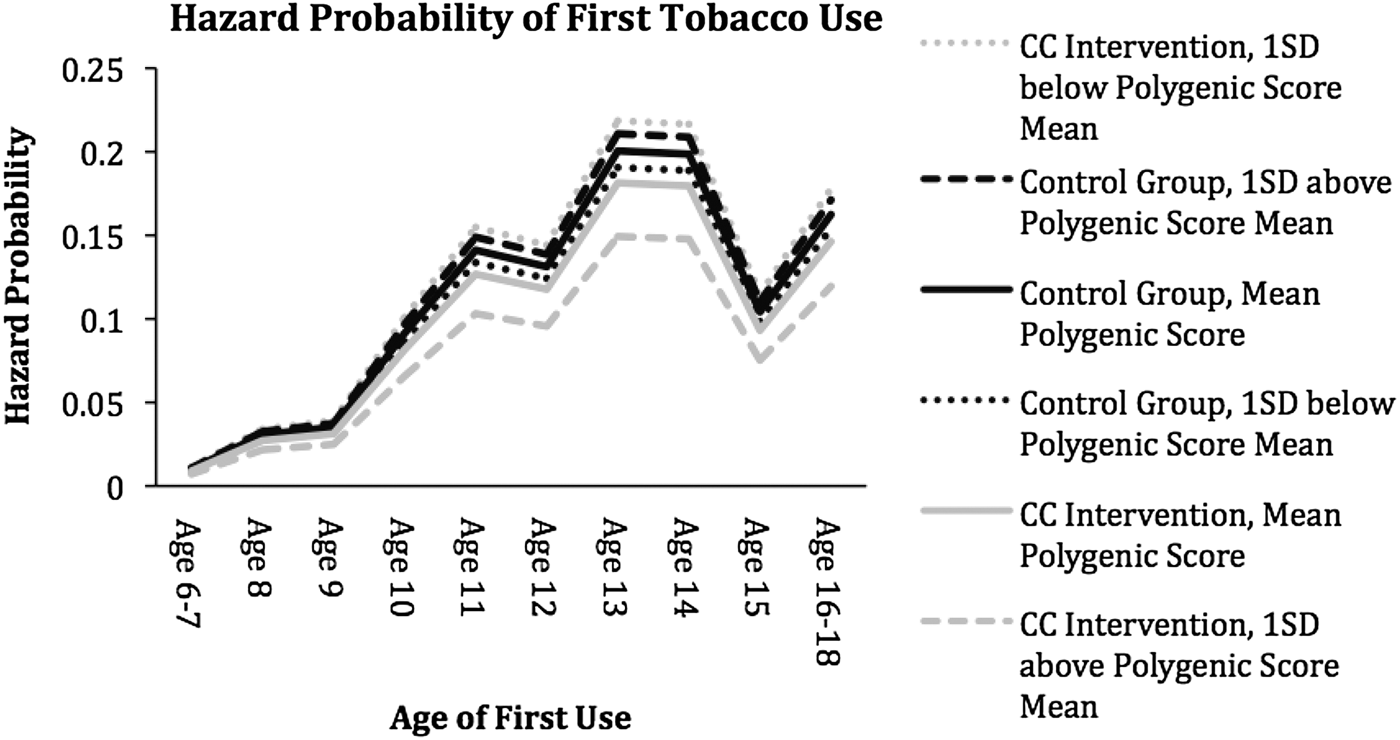
In the moderation model, an interaction term was created to measure the interaction between the polygenic score and intervention. This interaction term was a significant predictor of risk of first tobacco use (est. = –0.296, SE = 0.150, p = .049), suggesting that those individuals who received the CC intervention and who had a high polygenic score had the lowest risk of tobacco use from age 6 to age 16 (see Figure 1). In the presence of an interaction, we took steps to decompose the interaction to further facilitate its interpretation. Hazard probabilities were plotted for the control and intervention groups at the mean level of the polygenic score, 1 SD above the mean of the polygenic score and 1 SD below the mean of the polygenic score. As can be seen in Figure 1, hazard probabilities were lowest across waves for individuals in the intervention condition and 1 SD above the mean for the polygenic score. This suggests that individuals in the CC who had a high polygenic score (1 SD above the mean) had the lowest risk for first use of tobacco. The second lowest hazard probability was found in individuals in the control group and the mean level of the polygenic score.
Figure 1. Hazard probability of tobacco initiation. Covariates not involved in the moderation are presented at the mean level. Black lines correspond to hazard curves of the control groups, while gray lines correspond to hazard curves to the classroom-centered intervention groups. Solid lines are associated with the hazard curves of the mean polygenic scores, while dotted lines are associated with curves 1 SD on below the mean of the polygenic score. Dashed lines are associated with hazard curves 1 SD above the mean of the polygenic score.
In addition, adjusted hazard odds were calculated for the CC intervention at the mean of the polygenic score, 1 SD above and 1 SD below the mean of the polygenic score. The adjusted hazard odds ratio for the effect of the CC intervention on first tobacco use suggests that the classroom intervention had a protective, but not significant, effect at the mean of the polygenic score (hazard odds ratio [hOR] = –0.124, p = .321). The adjusted hazard odds for the effect of the CC intervention on first tobacco use suggests that the intervention had a protective and significant effect at 1 SD above the mean of the polygenic score (hOR = –0.420, p = .034).
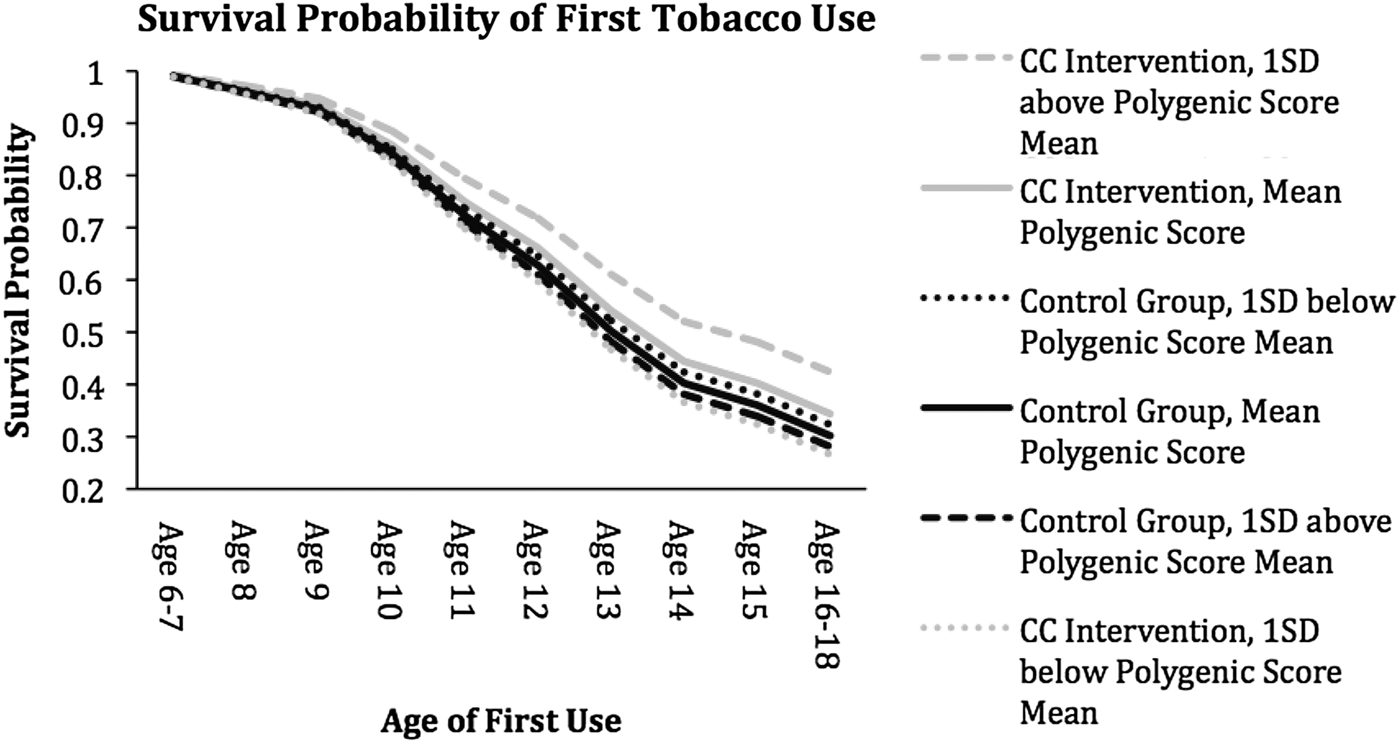
The cumulative survival plot (Figure 2) shows that individuals in the intervention group and with polygenic scores one standard deviation above the mean had the highest probability of survival across waves. From this plot, we can see the median survival times (survival probability = 0.5) for each subgroup, suggesting that individuals in the CC intervention with a polygenic score 1 SD above the mean had a median survival time of around 14.5 years whereas individuals in the CC intervention with a polygenic score 1 SD below the mean had a median survival time of around 13 years.
Figure 2. Survival probability of tobacco initiation. Covariates not involved in the moderation are presented at the mean level. Black lines correspond to survival curves of the control groups, while gray lines correspond to survival curves to the classroom-centered intervention groups. Solid lines are associated with the survival curves of the mean polygenic scores, while dotted lines are associated with curves 1 SD below the mean of the polygenic score. Dashed lines are associated with survival curves 1 SD above the mean of the polygenic score.
Discussion
As noted earlier, Wang et al. (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) found a main effect of the CC intervention on age of first cigarette smoked. Highlighting the significance of this finding is evidence that the earlier the onset of smoking, the less likely one is to successfully quit smoking over the life course (Breslau et al., Reference Breslau, Fenn and Peterson1993; Breslau & Peterson, Reference Breslau and Peterson1996; D'Avanzo et al., Reference D'Avanzo, La Vecchia and Negri1994). In the current study we sought to extend the findings of Wang et al. (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) to include the role of genetics as a source of variation in the CC intervention's effect on the age of first cigarette smoked. The search for such interactions within the context of randomized intervention trials is in accord with Belsky and colleagues' (Belsky et al., Reference Belsky, Bakermans-Kranenburg and van IJzendoorn2007; Belsky & Pleuss, Reference Belsky and Pluess2009, Reference Belsky and Pluess2013; Ellis et al., Reference Ellis, Boyce, Belsky, Bakermans-Kranenburg and van IJzendoorn2011) differential susceptibility hypothesis and the considerable evidence of genetic influences on the frequent targets of such interventions, including the initiation and course of substance use in adolescence and young adulthood. It is also in line with the substantial evidence of genetic influence on the early risk behaviors and processes associated with these distal targets. Our findings add to the small but growing body of literature on Gene × Intervention interactions in the context of randomized trials of preventive interventions in childhood and adolescence.
Consistent with Uhl, Drgon, Johnson, Walther, et al. (Reference Uhl, Drgon, Johnson, Ramoni, Behm and Rose2010), we found that the greatest intervention impact was among those participants with higher polygenic scores. As we speculated in the introductory section, it may be that the polygenic score we utilized represents an index of an individual's sensitivity to the reinforcing effects of nicotine. In line with Wang et al.'s (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) finding that friends' tobacco use was the best predictor of survival to first cigarette smoked and the evidence of increased plasma levels of nicotine as a function of exposure to secondhand smoke (Okoli et al., Reference Okoli, Kelly and Hahn2007), participants with such an increased sensitivity may have been less likely to refuse offers to smoke from peers or older siblings. Thus, the effects of the CC intervention may have been magnified in this subgroup relative to their control group counterparts. These results appear to be consistent with Belsky and colleagues' concept of “for better or worse” (Belsky et al., Reference Belsky, Bakermans-Kranenburg and van IJzendoorn2007; Belsky & Pleuss, Reference Belsky and Pluess2009, Reference Belsky and Pluess2013; Ellis et al., Reference Ellis, Boyce, Belsky, Bakermans-Kranenburg and van IJzendoorn2011), wherein the expression of potentially advantageous genetic variants are realized in the context of a preventive intervention. That is, the effect of the intervention in the current study appeared to be maximized at the upper end of the polygenic score.
We acknowledge that our explanation of mechanisms underlying the increased intervention impact seen for those with higher polygenic scores is purely speculation. To that end, we plan to undertake a decomposition of the variants making up the polygenic score in an effort to determine their biological function. We also plan to include measures of subjective responses to smoking tobacco and the use of the other substances in our future assessments of the study sample, along with a history of childhood respiratory diseases. The incidence of asthma is extremely high in urban, economically distressed, African Americans and may serve as a protective factor when it comes to initiating smoking. Our next steps also include carrying out candidate gene analyses using the candidate genes utilized by Bakersman-Kranenberg et al. (2008), Kegel et al. (Reference Kegel, Bus and van IJzendoorn2011), and Brody and colleagues (Beach et al., Reference Beach, Brody, Lei and Philbert2010; Brody et al., Reference Brody, Beach, Philibert, Chen and Murray2009).
With regard to the practical implications of our findings, as we pointed out in Ialongo et al. (Reference Ialongo, Rogosch, Cicchetti, Toth, Buckley, Petras and Cicchetti2006), the use of genetic information to tailor preventive or treatment interventions is fraught with a myriad of difficulties. First, classification accuracy is still relatively poor. Simply because a variable predicts an outcome does not mean it can accurately classify an individual with respect to an outcome as Kraemer et al. (Reference Kraemer, Kazdin, Offord, Kessler, Jensen and Kupfer1999) have pointed out. Second, the cost and logistics associated with obtaining DNA samples from millions of school children and then assaying the samples and translating the results of them into individualized intervention plans are simply untenable at this point in time. More reasonable is the use of phenotypic indicators of common genetic influences, such as measures of temperament and personality.
It is important to note that though statistically significant, the magnitude of effects found for the CC intervention found were relatively modest, even among those with higher polygenic scores. As we indicated in Wang et al. (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012), the ideal design would feature universal interventions throughout elementary, middle, and high school. Selective and indicated interventions would then be nested within these universal interventions to address needs of students at elevated risk of untoward outcomes. It is also important to point out that although Wang et al. (Reference Wang, Browne, Petras, Stuart, Wagner and Lambert2009, Reference Wang, Storr, Green, Zhu, Stuart and Lynne-Landsman2012) did not find a main effect for the FSP intervention, the effect was in the expected direction. It may simply have been due to the practical limitations of providing enough of a “dose” of the intervention to parents. More specifically, the FSP intervention parents received only a fraction of the training and mentoring compared to the CC interventions, who were run by teachers.
In terms of study limitations, we relied on retrospective self-reports of smoking. Thus, our findings may be subject to both social desirability and recall biases. That the substance use questions were self-administered lessens our concern with the former, although biological measures of the metabolites of smoking tobacco would have strengthened the study. With regard to recall biases, our smoking assessments began prior to the age that even most early initiators begin smoking. Serving to mitigate the recall bias concern is that we asked the participants to report on age of first use on a yearly basis through age 18. Nevertheless, recall bias may be greatest among those participants with attention/concentration problems, who, in turn, may be more likely to initiate substance use at a younger age, further exacerbating recall bias. Another limitation is that like most community-based randomized intervention trials, there was variation in the degree that the intervention was implemented with fidelity by teachers. In addition, as is the case in many urban, socioeconomically distressed communities, school absence can be high; thus, not all students may have received an equivalent level of exposure to the intervention. It is important that the majority of teachers were observed to implement with a high degree of fidelity, and no teacher failed to implement at least some aspect of the CC intervention. Moreover, there was no evidence of differential absenteeism rates across conditions. A final limitation centers on generalizability of our findings to other ethnic groups living in more geographically and socioeconomically diverse settings. Replication in other ethnic groups and settings is necessary, particularly in light of the potential for population stratification in admixed samples.
In contrast to these limitations, our study features a number of strengths. First, randomized universal prevention trials offer one of the most methodologically rigorous means of testing Gene × Environment interactions. Second, the sample is representative of the entire cohort of first graders entering the nine participating elementary schools at the onset of the study. Thus, the selection biases associated with samples of convenience were avoided. That we did not find differential attrition over the course of the study further bolsters our confidence that our results were not influenced by sampling bias. Third, the annual follow-up from entrance into middle school and through high school is relatively rare in the field. Such annual follow-ups allowed a more precise timing of the intervention effects. Fourth, our use of a polygenic score reflects the current thinking in the molecular genetics literature that complex human behavior is likely a product of multiple genes working in concert and whose collective influence is greater than any one of its parts.