


Through its normative daily diurnal rhythms (Gunnar & Quevedo, Reference Gunnar and Quevedo2007; Sapolsky, Romero, & Munck, Reference Sapolsky, Romero and Munck2000), and acute, real-time responses to psychological stress (Kirschbaum & Hellhammer, Reference Kirschbaum and Hellhammer1989), the hypothalamus–pituitary–adrenal (HPA) axis, a core physiological stress system, supports our abilities to adapt to environmental demands (McEwen, Reference McEwen2000; Sapolsky et al., Reference Sapolsky, Romero and Munck2000). However, prolonged and extensive activation of the HPA axis can lead to “wear and tear,” or allostatic load, that can have long-term detrimental effects on health, cognition, and neural structure and function (McEwen, Reference McEwen2000; McEwen & Wingfield, Reference McEwen and Wingfield2003). As such, increasing attention is being paid to the ways through which young children's early experiences may calibrate (Del Giudice, Ellis, & Shirtcliff, Reference Del Giudice, Ellis and Shirtcliff2011) or tune their physiological stress systems over time: physiological adaptation to context.
Nuanced aspects of these contemporary theories of physiological adaptation (e.g., Blair & Raver, Reference Blair and Raver2012; Boyce & Ellis Reference Boyce and Ellis2005; Del Giudice et al., Reference Del Giudice, Ellis and Shirtcliff2011; Parker & Maestripieri, Reference Parker and Maestripieri2011), remain largely untested. The aim of the present study is to leverage our prospective longitudinal design to begin to test these complexities empirically.
HPA Axis and Development
HPA axis activity is evident early in development. Unbound, biologically active levels of cortisol, the end-product hormone of the HPA axis, are observed at birth, with newborns able to mount a stress response to aversive environmental stimuli (e.g., heal stick; Davis et al., Reference Davis, Townsend, Gunnar, Georgieff, Guiang and Ciffuentes2004; see Gunnar & Quevedo, Reference Gunnar and Quevedo2007; Hostinar, Sullivan, & Gunnar, Reference Hostinar, Sullivan and Gunnar2014). The normative circadian rhythms of the HPA axis tend to be detectable in a nascent form by 4 to 6 months (Larson, White, Cochran, Donzella, & Gunnar, Reference Larson, White, Cochran, Donzella and Gunnar1998) and become increasingly adultlike as children give up their afternoon naps (Watamura, Donzella, Kertes, & Gunnar, Reference Watamura, Donzella, Kertes and Gunnar2004). The HPA axis also undergoes substantial developmental change across infancy and early childhood. In particular, theory and a growing body of empirical evidence suggest that young children's developing adrenocortical systems are highly regulated by their early experiences (Blair & Raver, Reference Blair and Raver2012; Boyce & Ellis, Reference Boyce and Ellis2005; Del Giudice et al., Reference Del Giudice, Ellis and Shirtcliff2011; Gunnar & Vasquez, 2006; Hostinar et al., Reference Hostinar, Sullivan and Gunnar2014).
HPA Axis and Social Interactions in Infancy
As reviewed by Gunnar and Herrera (Reference Gunnar, Herrera and Zelazo2013) and Hostinar et al. (Reference Hostinar, Sullivan and Gunnar2014), there is ample evidence from well-controlled studies of rodents and nonhuman primates that early caregiving environments play a powerful role in the organization and function of the developing HPA axis. Meaney and Szyf's (Reference Meaney and Szyf2005) comprehensive research program with rodents has shown that normative differences in maternal behavior, such as licking and grooming by rat dams, can have long-lasting effects on the infant rats’ developing physiological stress systems (Caldji, Diorio, & Meaney, Reference Caldji, Diorio and Meaney2000; Caldji et al., Reference Caldji, Tannenbaum, Sharma, Francis, Plotsky and Meaney1998; Weaver et al., Reference Weaver, Cervoni, Champagne, D'Alessio, Sharma and Seckl2004) and in turn behavior (Weaver, Menaey, & Szyf, Reference Weaver, Meaney and Szyf2006; Toki et al., Reference Toki, Morinobu, Imanaka, Yamamoto, Yamawaki and Honma2007). Similar findings are evident with nonhuman primates. For instance, young monkeys that experience repeated, unpredictable separations from their mothers (i.e., not complete removal; e.g., Sanchez et al., Reference Sánchez, Noble, Lyon, Plotsky, Davis and Nemeroff2005) tend to show atypical diurnal cortisol rhythms. Some findings indicate similar effects with regard to HPA axis reactivity to acute experiential stressors (Wiener, Bayart, Faull, & Levine, Reference Wiener, Bayart, Faull and Levine1990).
Growing evidence considering normative variation in young children's environments suggests that experience plays a similar role in the development of the HPA axis in early childhood. A well-developed literature indicates that stress physiology in young children is regulated by their social experiences with meaningful adults (see Gunnar & Donzella, Reference Gunnar and Donzella2002). For example, from an attachment perspective, children with secure relationships with their caregivers, thought to reflect sensitive caregiving, tend to show lower levels of stress reactivity to typical stressors, such as separation and physical examinations (Gunnar, Reference Gunnar1992; Gunnar, Broderson, Nachmias, Buss, & Rigatuso, Reference Gunnar, Brodersen, Nachmias, Buss and Rigatuso1996). Flinn's intensive longitudinal work with children and families in Dominica has shown that heightened levels of cortisol tend to occur after meaningful family events, such as a conflict at home (Flinn, Reference Flinn2006; Flinn & England, Reference Flinn and England1995). Similarly, in prior work with the present sample, our group has noted links between lower quality parenting and heightened cortisol levels in infancy (Blair et al., Reference Blair, Granger, Willoughby, Mills-Koonce, Cox and Greenberg2011).
Physiological Adaptation to Context
Building on Boyce and Ellis’ (2005) seminal work, recent evolutionarily informed theoretical models highlight the notion that this social regulation of children's developing physiological stress systems (including, but not limited to, the HPA axis) plays an important adaptive role ontogenetically and phylogenetically. Over time, children's normative rearing experiences are proposed to calibrate the organization and “threshold activity” of children's physiological stress systems, tuning these systems to local environmental demands (Blair & Raver, Reference Blair and Raver2012; Del Giudice et al., Reference Del Giudice, Ellis and Shirtcliff2011). In turn, this conditional adaptation, or adaptive calibration, to context is theorized to regulate physiology and behavior in fitness-relevant ways, ultimately impacting phenotypes ranging from physical growth and competitive risk taking to learning and reproduction (see Del Giudice at al., 2011).
Central to these models is the idea of physiological “openness” or “sensitivity” to experience. Children's typical rearing conditions are theorized to tune their developing physiological stress systems to become comparatively more or less sensitive to their subsequent experiences. Del Giudice et al.'s (2011) well-cited adaptive calibration model (ACM) makes specific predictions about the functional form of this relation. Invoking the concept of life history (LH) strategies from evolutionary ecology, they propose that the effects of early experience on the emerging organization on children's physiological stress systems likely culminates in these systems functioning in a U-shaped manner.
Early experiences marked by characteristically unsupportive or challenging social experiences are theorized to cause repeated, chronic activation of the autonomic and adrenocortical systems in ways that psychologically (e.g., cognitive biases) and physiologically (e.g., limbic fear circuits and weakened cortical downregulation) sensitize the organization of these nascent systems toward heightened vigilance and responsiveness to environmental change (e.g., perceived threat). This may be represented physiologically by amplification of acute response of these systems, as well as changes in basal setpoint of these systems (Feeney, Gaffney, & O'Mara, Reference Feeney, Gaffney and O'Mara2012; Del Giudice et al., Reference Del Giudice, Ellis and Shirtcliff2011; Montoya, Terberg, Bos, & van Honk, Reference Montoya, Terburg, Bos and van Honk2012; van Honk et al., Reference van Honk, Tuiten, van den Hout, Koppeschaar, Thijssen and de Haan1998). Such “vigilant” physiological profiles are posited to reflect so-called fast LH strategies that confer short-term adaptive advantages by supporting the organism to cope effectively with unpredictable physical and psychological challenges, as well as longer term physiological (e.g., pubertal timing) and psychological traits (e.g., risk taking and earlier sexual debut), that would have maximized fitness advantages in the context of unpredictable and/or dangerous environments in our evolutionary past.
Moderate levels of support and/or challenge, such as those experienced by children navigating more normative experiential “ups and downs” of early childhood (e.g., temporary parental separation or insensitivity, sibling/peer relations, and child-care entry), are thought to initiate more intermittent and ultimately surmountable physiological and psychological stress responses. Sometimes referred to as “steeling” (Rutter, Reference Rutter2012) or “stress inoculation” (Lyons & Parker, Reference Lyons and Parker2007), the moderate (yet reasonably common) activation and recovery of these systems is thought to support the development of comparatively less labile physiological profiles that help to buffer the organism against the effects of subsequent environmental stressors. Such buffered profiles are proposed to reflect slower LH strategies that confer fitness-relevant traits that are phylogenetically suited to such environments (e.g., slower pubertal timing, low anxiety, and resilience to changes in the caregiving environment).
In contrast, very supportive contexts are proposed to lead to minimal activation of the stress-response systems early in life. As such, the organization of these children's physiological systems are, according to theory, neither desensitized to the ebb and flow of moderate levels of stress, nor sensitized for physiological and psychological vigilance to threat. Rather, these “sensitive” physiological profiles are thought to maintain a physiological plasticity to their (prototypically positive) experiences. Specifically, Del Giudice et al. (Reference Del Giudice, Ellis and Shirtcliff2011) posit that, despite our general tendency to align physiological stress responses with affectively negative experiences, the autonomic nervous system and HPA axis often show similar activation patterns with respect to the anticipation of and response to affectively salient positive experiences.
This “positive end” of the physiological arousal spectrum has been studied far less empirically. However, some work supports the idea. For instance, in the same long-term study of children of Dominica mentioned above, Flinn (Reference Flinn2006) showed that, on Christmas Eve, children's anticipation of Christmas Day was predictive of cortisol levels that were approximately 1 SD more elevated that their typical levels. These positive relations were particularly pronounced for children from lower risk homes, who had high positive expectations about Christmas. Work emerging from the temperament literature suggests that temperamentally extroverted children, who typically enjoy social interactions, tend to show elevated cortisol levels in the social context of their peers. For example, in multiple samples, temperamentally exuberant children with high levels of activity and positive emotion have been found to show cortisol increases over the course of the school day, with the magnitude of the effect growing stronger across the first week of school (Bruce, Davis, & Gunnar, Reference Bruce, Davis and Gunnar2002; Davis, Donzella, Krueger, & Gunnar, Reference Davis, Donzella, Krueger and Gunnar1999). Similarly, when they experience positive peer relations (e.g., friendships and popularity), even socially reticent preschoolers have been found to evince rather substantial increases in their basal cortisol level over the course of the school year (Tarullo, Mliner, & Gunnar, Reference Tarullo, Mliner and Gunnar2011). In addition, studies of adults have shown that athletes often show anticipatory and/or postmatch increases in their cortisol levels after competing. These increases typically emerge for both the winners and the losers, with some evidence suggesting larger cortisol increases for the winners (see Salvador & Costa, Reference Salvador and Costa2009).
The idea that positive experiences may support heightened levels of physiological arousal is also consistent with theoretical and empirical work suggesting that moderate increases in catecholamines (e.g., norepinephrine and dopamine; Vijayraghavan, Wang, Birnbaum, Williams, & Arnsten, Reference Vijayraghavan, Wang, Birnbaum, Williams and Arnsten2007) and glucocorticoids (e.g., cortisol; Blair, Granger, & Peters Razza, Reference Blair, Granger and Peters Razza2005; Lupien, Maheu, Tu, Fiocco, & Schramek, Reference Lupien, Maheu, Tu, Fiocco and Schramek2007) may support optimal higher order cognitive functioning and regulatory control, broader phenotypes predicted by positive caregiving (Blair, Raver, & Berry, Reference Blair, Raver and Berry2014). The proposal that these vigilant physiological profiles translate into both heighted physiological responsiveness to social experiences and optimal self-regulatory control plays a central role in the “slow” LH strategies that this profile is thought to reflect. Specifically, when experiential threats to early survival and eventual reproductive success are minimal, maturation slows and reproductive success depends increasingly on (a) the creation and maintenance of social relationships, and (b) investing in the quality (rather than quantity) of one's offspring. Social responsiveness and effective self-regulation are vital toward these ends.
Although empirical tests of the ACM are only beginning to emerge in studies of children (e.g., Del Giudice, Hinnant, Ellis, & El-Sheik, Reference Del Giudice, Hinnant, Ellis and El-Sheikh2012), these predictions are seemingly consistent with findings from “stress inoculation” studies with nonhuman primates (see Parker & Maestripieri, Reference Parker and Maestripieri2011). For instance, squirrel monkeys exposed to moderate, surmountable levels of stress shortly after weaning (~17 weeks), induced by short but somewhat frequent maternal separations, have been found to show reductions in their autonomic and adrenocortical response to subsequent stressors, compared with their peers who are not separated from their mothers. Although it remains untested, such differences in the acute response may be partially explained by more systematic shifts in the basal setpoints of these systems. For instance, these authors found that monkeys experiencing these modest separations showed more efficient cortisol-induced downregulation of the acute HPA axis response (Lyons, Lopez, Yang, & Schatzberg, Reference Lyons, Lopez, Yang and Schatzberg2000; Parker, Buckmaster, Schatzberg, & Lyons, Reference Parker, Buckmaster, Schatzberg and Lyons2004). Across the board, the developmental timing here is noteworthy, because these separations took place during a span in which squirrel monkeys typically show nascent independence, rather than in, say, early infancy when such separations could be experienced as being more stressful. Nonetheless, these findings suggest that moderate experiences of stress during a period of very early independence may “buffer” or “steel” these developing physiological systems to the impacts of subsequent stress exposures.
In contrast, consistent with a vigilant physiological profile, when stressors are more substantial, they tend to predict more pronounced autonomic and adrenocortical responses. For instance, in their studies of bonnet macaques, Coplan et al. (Reference Coplan, Andrews, Rosenblum, Owens, Friedman and Gorman1996, Reference Coplan, Trost, Owens, Cooper, Gorman and Nemeroff1998) experimentally induced maternal stress and low-quality parenting by forcing some mothers to face unpredictable foraging demands when their offspring were infants. They found that, as adolescents, infants raised under these conditions tended to show considerably elevated levels of several stress-induced hormones/neurochemicals (e.g., cortisol, corticotropin-releasing factor, serotonin, dopamine, and norepinephrine), compared with those raised by mothers with predictable foraging demands (Coplan et al., Reference Coplan, Andrews, Rosenblum, Owens, Friedman and Gorman1996, Reference Coplan, Trost, Owens, Cooper, Gorman and Nemeroff1998).
Taken together, there is clear evidence that early experiences impact adrenocortical functioning. Social supports afforded by primary caregivers, such as their sensitivity and responsiveness to the child's emotional needs, are presumed to be especially important early in life, given the substantial external regulatory support required during this developmental span (Feldman, Reference Feldman2007; Fox & Calkins, Reference Fox and Calkins2003). Contemporary models of physiological adaptation to context have led to increasingly specific developmental hypotheses about the nature of these relations. In particular, they suggest that children's typical experiences with caregivers over time may “calibrate” the HPA axis to be more or less vigilant/sensitive to changes in environmental cues.
Prospective longitudinal designs can be leveraged to begin to address these hypotheses. Specifically, with longitudinal data, one can partition between-person effects due to between-child variation in children's typical experiences over extended periods of time from within-person effects due to within-person, time-specific changes from his/her typical levels over time. Similar to the idea of “states” and “traits,” between-person relations may reflect effects due to longer term systematic components (i.e., traits) of the rearing environment that are theorized to tune the sensitivity of the HPA axis to experience. In contrast, within-person effects, representing changes in physiology that occur in the context of the changing environment (i.e., states), may reflect physiological sensitivity to experience.
The ACM implies rather specific predictions about the respective between- and within-person effects of early caregiving experiences, such as caregiver sensitivity, on children's adrenocortical functioning. Between children, one might expect that young children who experience characteristically highly sensitive and supportive caregiving over time will tend to have lower averageFootnote 1 basal cortisol levels, relative to their peers who experience characteristically less sensitive caregiving.
Within-person relations, of course, are interpreted quite differently. They address the extent to which deviations (i.e., increases or decreases) from one's own “typical” caregiving experience are associated with shifts in cortisol from one's own “typical” cortisol level. More simply, they address the question: are changes in caregiver sensitivity associated with contemporaneous changes in children's cortisol levels? With respect to such within-person effects, the ACM implies a cross-level interaction. Specifically, the model suggests that the magnitude (and direction) of the within-person relation between sensitive caregiving and children's adrenocortical functioning should differ for children who typically experience low versus moderate versus high levels of caregiver sensitivity over time, because these typical traitlike aspects of the caregiving environment are the theorized mechanism underlying the development of particularly vigilant, buffered, and sensitive physiological profiles, respectively.
That is, if characteristically very low and very high levels of caregiver sensitivity are predictive of physiologically more malleable profiles, as posited by the vigilant and sensitive profiles of the ACM, respectively, then one would expect that children who typically experience very low or very high levels of caregiver sensitivity over time should have adrenocortical systems that are particularly responsive to changes in these caregiving environments (i.e., stronger within-person maternal-sensitivity effects). In contrast, consistent with the model's proposed “buffered” physiological profile, changes in the maternal sensitivity would be expected to be largely unrelated with adrenocortical functioning for children who tend to experience characteristically moderate levels of maternal sensitivity over time.
In addition, the ACM raises the possibility that the direction of the respective within-person effects may be different for children with typically low versus typically high levels of caregiver sensitivity over time. On average, children who typically experience low levels of caregiver sensitivity over time are theorized to evince cortisol levels that are chronically more elevated than their peers who typically experience highly sensitive care (i.e., negative between-child relation). For these former children, increases in caregiver sensitivity may serve as an important self-regulatory support to help downregulate their prototypically elevated cortisol levels. In contrast, increases in maternal sensitivity may mean something quite different for children who have experienced characteristically highly sensitive caregiving and maintained characteristically low levels of physiological arousal. Similar to the way socially motivated children respond physiologically to peer interactions (Bruce et al., Reference Bruce, Davis and Gunnar2002; Davis et al., Reference Davis, Donzella, Krueger and Gunnar1999), for children who have historically experienced highly sensitive care, increases in sensitivity may serve as a cue to prepare physiologically and psychologically for the “good things to come.” That is, for these children, increases in maternal sensitivity may stimulate cortisol increases from their prototypically low cortisol baselines.
In summary, the ACM suggests that (a) young children's typical experiences with their caregivers may impact their physiological reactions to deviations from these experiential norms, and (b) contemporary theory (e.g., Blair & Raver, Reference Blair and Raver2012; Boyce & Ellis, Reference Boyce and Ellis2005; Del Giudice et al., Reference Del Giudice, Ellis and Shirtcliff2011; Lyons & Parker, Reference Lyons and Parker2007) suggests that this relation will be curvilinear (possibly U-shaped), such that those typically experiencing very low or very high levels of caregiver sensitivity will tend have physiological profiles that are comparatively more labile to experience than those with histories of moderate levels of caregiver sensitivity.
The Present Study
The aim of the present study was to leverage our longitudinal design to test the extent to which within-person relations between time-varying maternal sensitivity and children's cortisol levels were particularly pronounced for those at the low and high ends of maternal-sensitivity distribution. Specifically, we addressed the following research questions:
-
1. Between children, do those who experience higher levels of maternal sensitivity on average across infancy and toddlerhood tend to have lower basal cortisol levels than children who experience lower average levels of maternal sensitivity over this period?
-
2. Are within-person changes in maternal sensitivity predictive of contemporaneous changes in children's cortisol levels?
-
3. If so, is this relation moderated by children's typical levels of maternal sensitivity over time? Specifically, are the within-person effects particularly pronounced for children who typically experience either very low or very high levels of maternal sensitivity (compared with those with more moderate average maternal-sensitivity levels)?
Informed by the ACM (Del Giudice et al., Reference Del Giudice, Ellis and Shirtcliff2011) between children, we hypothesized a negative relation, such that children experiencing higher levels of maternal sensitivity, on average, between 7 and 24 months of age, will tend to show cortisol levels that are lower than those of their peers who experience lower average levels of maternal sensitivity over this period. Based on prior between-person findings (e.g., Blair et al., Reference Blair, Granger, Willoughby, Mills-Koonce, Cox and Greenberg2011), we expected this relation to be linear, though we allowed for the possibility of curvilinear function. We expected this relation to be robust in propensity-score-weighted models that adjust for a large number of potential covariates.
Within children, we hypothesized that (in absolute terms) the magnitudes of the within-child effects would be particularly pronounced for children who typically experienced either very low or very high levels of maternal sensitivity on average across infancy and toddlerhood. Specifically, perhaps reflecting a vigilant physiological profile, we hypothesized a negative within-person effect, such that increases in maternal sensitivity would be associated with contemporaneous cortisol decreases from these children's prototypically high cortisol levels (and the inverse for maternal sensitivity decreases). In contrast, perhaps reflecting a sensitive profile, we hypothesized a positive within-person relation, such that increases in maternal sensitivity would be associated with contemporaneous cortisol increases from these children's prototypically low cortisol levels (and the inverse for maternal sensitivity decreases). Finally, potentially reflecting a buffered physiological profile, we hypothesized that changes in maternal sensitivity would not be associated with changes in children's cortisol levels.
Method
Participants
The Family Life Project (FLP) was designed to study young children and families in two of the four major geographical areas of the United States with high poverty rates (Dill, Reference Dill1999). Specifically, three counties in Eastern North Carolina and three counties in Central Pennsylvania (PA) were selected to be indicative of African American families in the South and Caucasian families in Appalachia, respectively. The FLP adopted a developmental epidemiological design in which sampling procedures were employed to recruit a representative sample of 1,292 children whose families resided in one of the six counties at the time of the child's birth. Low-income families in both states and African American families in North Carolina were oversampled (African American families were not oversampled in Pennsylvania because the African American populations of these counties was <5%). A comprehensive description of the sampling procedure is provided by Vernon-Feagans, Cox, and the Key FLP Investigators (Reference Vernon-Feagans and Cox2013). Sampling weights were used in all analyses (Asparouhov, Reference Asparouhov2008).
Procedure
The current study focused on data collected across infancy and toddlerhood. At the ages of 7, 15, and 24 months, saliva samples were collected from children during home visits to assess basal or “non(intentionally) stimulated” levels. Saliva samples were collected after the data collectors had been in the home for at least 1 hr interviewing the primary caregiver and prior to conducting a number of assessments with children. The modal time of collection was approximately 10:00 a.m. at each wave of collection; however, there was some variability (Table 1). Time varying and mean time of day were adjusted in all models to account for these timing differences. We use the term basal given that no active, purposeful stimulation was present, and children were given ample time (~1 hr) to return to baseline after the arrival of the research assistants (RAs). There is little evidence that exposure to novel adults serves as an acute stressor with children of this age (even when intended to do so; Gunnar, Talge, & Herrara, 2009). We cannot rule out, however, that our basal cortisol levels partly capture HPA axis stimulation due to the visit. Saliva samples were also collected during/after acute stress paradigms that were conducted at each visit. Because the stress paradigms varied somewhat over time, measurement changes are intrinsically conflated with cortisol changes. This unfortunately renders measures of cortisol stress responses inappropriate for our questions, because temporal changes in cortisol serve as our core dependent variable.
Table 1. Zero-order correlations and descriptive statistics for the main substantive variables and select covariates

Note: Cort, Basal cortisol; M. Sens, maternal sensitivity.
†p < .10. *p < .05. **p < .01. ***p <.001.
Unstimulated whole saliva was collected using either cotton or hydrocellulose absorbent material and expressing sample into 2-ml cryogenic storage vials using a needleless syringe (cotton) or by centrifugation (hydrocellulose). Prior studies have indicated no differences in cortisol concentrations associated with the two collection techniques (Granger et al., Reference Granger, Kivlighan, Fortunato, Harmon, Hibel and Schwartz2007; Harmon, Hibel, Rumyantseva, & Granger, Reference Harmon, Hibel, Rumyantseva and Granger2007). Children's saliva samples were immediately frozen at –20 °C and subsequently stored at –80 °C. Intra- and interassay coefficients of variability for cortisol (μg/dl) were an average of less than 10% and 15%. All samples were assayed in duplicate. The criterion for repeat testing was variation between duplicates greater than 20%; the average of the duplicates was used in all analyses. During these same home visits, data collectors conducted interviews with the primary caregiver and made observations of the household and neighborhood.
Measures
Salivary cortisol
All samples from the 7-, 15-, and 24-month assessments were assayed for salivary cortisol using a highly sensitive enzyme immunoassay US FDA 510k cleared for use as an in vitro diagnostic measure of adrenal function (Salimetrics, State College, PA). The cortisol distributions were subject to log transformation to correct positive skew. We examined child temperature and use of medications (e.g., acetaminophen) as influences on child cortisol and consider each, in addition to time of day in which the saliva collection occurred, as time-varying and time-invariant (i.e., mean levels over time) control covariates. At 7 months of age, the effects of time since eating and waking were also assessed. They were unrelated to cortisol or maternal sensitivity; however, these data were not collected at the 15- and 24-month visits.
Maternal sensitivity
Primary-caregiver sensitivity was based on observer ratings of positive parenting during a semistructured play task at 7, 15, and 24 months of age (see Cox, Paley, Burchinal, & Payne, Reference Cox, Paley, Burchinal and Payne1999). In this task, the primary caregiver and infant were presented with a set of developmentally appropriate toys, and caregivers were asked to play with their infants as they would normally. Primary caregiver behavior was recorded and subsequently coded for levels of sensitivity, stimulation, detachment, intrusiveness, positive regard, negative regard, and animation during their interactions with the child. Each was rated on a 5-point Likert-type scale (1 = not at all characteristic, 5 = highly characteristic) at the 7- and 15-month assessments and on a 7-point Likert-type scale at the 24-month assessment (this was rescaled to a 5-point scale for the current analyses). Informed by prior work with these data (blinded and peer reviewed), we created a positive parenting composite at each time point by taking the average of the primary caregiver's ratings across the sensitivity, animation, stimulation, positive regard, and detachment (reversed) levels. Because 99.6% of primary caregivers in our sample were the target child's biological mother and because our parenting measure is virtually identical to composites called maternal sensitivity (e.g., NICHD Early Child Care Research Network, 1997), we use that term presently. Two independent raters rated approximately 30% of the caregiver–child interactions; the cross-rater intraclass correlations for all subscales were above 0.80 at each time point and the positive parenting composite ranged between 0.87 and 0.91. Intraclass correlations of these longitudinal measures over time showed notable within-person variability in positive parenting over time; approximately 40% of the total variation was within persons. That is, as expected, mothers show some consistency in their sensitivity levels over time. However, they also show a substantial within-person variation in their sensitivity levels over time.
Control covariates
In addition to the weighting procedure discussed below, we included a number of time-varying and time-invariant demographic, maternal, and child control covariates in all models. We discuss these measures in detail in the online supplementary material. In brief, our time-varying control covariates included family income to needs ratio, time of day in which the saliva sample was collected, whether the child was taking prescription or over the counter medication, and age. Our time-invariant control covariates included research site; mean family income; mean time of day; mean medication taken; maternal education, reading ability, depression, and anxiety; and child sex, race, temperament, and birth weight. Other than temperament, which was measured at 7 months, all control covariates were measured at 2 months of age (i.e., pretreatment).
Missing data
Missing data were modest. For the cortisol measures, missingness ranged between 13% and 26%, the latter representing children's cortisol levels at 24 months. There was no clear pattern to the missingness; missingnesss was not associated with maternal education, income, or partnered status. Missingness in the predictors ranged between 7% and 18%. To adjust for biases emerging due to missing data, we fitted all models using full information likelihood estimation. This estimation helps to reduce biases to the extent to which missingness is missing at random, that is, conditionally random after adjusting for the observed variables included in the likelihood function (Enders, Reference Enders2010).
Data analytic plan
We adopted multiple strategies to leverage the longitudinal nature of the data and reduce potential unobserved variables bias. In the context of a multilevel modeling framework (Singer & Willett, Reference Singer and Willett2003), we tested a two-level model (Equation 1a–e) in which our time-varying Level 1 predictors, π1i and π3i were group-mean (i.e., person) centered. As such, each carries only within-person variation and is orthogonal to between-child variation in Level 2 predictors. Let π1i represent the within-child effect of maternal sensitivity, π2i represent the linear rate of change in cortisol between 7 and 24 months, and π3i represent the respective effects of a vector of several time-varying control covariates. Sometimes called a “hybrid model” (Allison, Reference Allison2009), this multilevel specification is more efficient than typical “child-fixed effect” models common to the econometrics literature, while still retaining the advantages of child-fixed effect models. Specifically, the estimates for Level 1 predictors are identical to those obtained via child-fixed effects and, thus, account for all potential observed and unobserved, time-invariant confounds. The hybrid approach also affords one the ability to simultaneously model between-person relations at Level 2. It is important that, for our purposes, they also allow one to estimate cross-level interactions. Specifically, as shown by parameter γ11, we address our question regarding the extent to which the within-child effect of maternal sensitivity varies as a function of mean level of maternal sensitivity. As displayed, the model for the residual variances includes only within-person, time-specific residual (ε ij ) and a random intercept (ζ0i ), implicitly constraining the within-person effects (including linear growth) to be identical across children. This parsimonious specification is tested and relaxed, as appropriate.
Level 1:
 $$\eqalign{{\rm Cor}{{\rm t}_{ij}} & = {\rm \;} {{\rm \pi} _{0i}} + {{\rm \pi} _{1i}}{\rm \;}\!\! \left( {{\rm Sen}{{\rm s}_{ij}} - \overline {{\rm Sen}{{\rm s}_i}}} \right) + {{\rm \pi} _{2i}}{\rm \;} \!\!\left( {{\rm Ag}{{\rm e}_{ij}} - 7\,{\rm m}{{\rm onths}_i}} \right) \cr & \quad\ \ { + _\;}{{\rm \pi} _{3i}}{\rm \;} \!\! \left( {{\rm cova}{{\rm r}_{ij}} - \overline {{\rm cova}{{\rm r}_i}}} \right) + {{\rm \varepsilon} _{ij}}.}$$
$$\eqalign{{\rm Cor}{{\rm t}_{ij}} & = {\rm \;} {{\rm \pi} _{0i}} + {{\rm \pi} _{1i}}{\rm \;}\!\! \left( {{\rm Sen}{{\rm s}_{ij}} - \overline {{\rm Sen}{{\rm s}_i}}} \right) + {{\rm \pi} _{2i}}{\rm \;} \!\!\left( {{\rm Ag}{{\rm e}_{ij}} - 7\,{\rm m}{{\rm onths}_i}} \right) \cr & \quad\ \ { + _\;}{{\rm \pi} _{3i}}{\rm \;} \!\! \left( {{\rm cova}{{\rm r}_{ij}} - \overline {{\rm cova}{{\rm r}_i}}} \right) + {{\rm \varepsilon} _{ij}}.}$$
Level 2:
 $${{\rm \pi} _{0i}} = {{\rm \gamma} _{00}} + {{\rm \gamma} _{01}}\overline {{\rm Sen}{{\rm s}_i}} + {{\rm \gamma} _{03}}\overline {{\rm cova}{{\rm r}_i}} + {{\rm \zeta} _{0i}},$$
$${{\rm \pi} _{0i}} = {{\rm \gamma} _{00}} + {{\rm \gamma} _{01}}\overline {{\rm Sen}{{\rm s}_i}} + {{\rm \gamma} _{03}}\overline {{\rm cova}{{\rm r}_i}} + {{\rm \zeta} _{0i}},$$
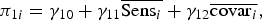 $${{\rm \pi} _{1i}} = {{\rm \gamma} _{10}} + {{\rm \gamma} _{11}}\overline {{\rm Sen}{{\rm s}_i}} + {{\rm \gamma} _{12}}\overline {{\rm cova}{{\rm r}_i}},$$
$${{\rm \pi} _{1i}} = {{\rm \gamma} _{10}} + {{\rm \gamma} _{11}}\overline {{\rm Sen}{{\rm s}_i}} + {{\rm \gamma} _{12}}\overline {{\rm cova}{{\rm r}_i}},$$
 $${{\rm \pi} _{2i}} = {{\rm \gamma} _{20}},$$
$${{\rm \pi} _{2i}} = {{\rm \gamma} _{20}},$$
 $${{\rm \pi} _{3i}} = {{\rm \gamma} _{30}}.$$
$${{\rm \pi} _{3i}} = {{\rm \gamma} _{30}}.$$
Although our hybrid model affords the advantage of explicitly partitioning within- and between-person effects (which can differ in magnitude and direction), a notable disadvantage is that our between-person (Level 2) estimates could be at comparatively higher risk for unobserved variables bias, that is, biases due to confounded “third variables” not included in the model. In an attempt to limit these threats, we weighted the between-persons component of our model (see Asparouhov, Reference Asparouhov2008) using inverse probability weights, based on a generalized propensity score (GPS) approach (Hirano & Imbens, Reference Hirano, Imbens, Gelman and Meng2004; Imai & van Dyk, Reference Imai and van Dyk2004). Specifically, GPSs were estimated to create inverse probability weights (IPWTs) for each child's average maternal sensitivity level across 7, 15, and 24 months (i.e., between-child variable).
We provide a detailed description of our GPS methods in the online-only supplementary materials; however, the basic logic is as follows: IPWTs are analogous to sampling weights. Sampling weights are based on the inverse probability of being sampled. When these weights are applied they create a pseudopopulation that is representative of the sampled population when the actual observed sample may be quite nonrepresentative (e.g., oversampling for certain characteristics). Presuming that one can validly estimate the probability of receiving a given level of maternal sensitivity (i.e., a continuously scaled treatment) as a function of all meaningful confounds, pretreatment, “third variables” that are the true drivers of an otherwise spurious association between mean maternal sensitivity and children's mean cortisol levels, the inverse of these probabilities can serve a similar weighting function. Specifically, they create a pseudopopulation in which maternal sensitivity is unrelated to the observed confounds used to estimate the weights, even when this may not be the case in the actual unweighted sample. In other words, the weights balance the confounds across each unit of maternal sensitivity, as one might expect if maternal sensitivity were randomly assigned. We detail our estimation methods, balance checks, and assumptions in the online-only supplementary Figure S.1.
In short, we created an overall weight based on the product of (a) the inversed probability of level of maternal sensitivity, and (b) inversed sampling probability (see Vernon-Feagans et al., 2013). These overall IPWTs were subsequently included as sampling weights in taxonomy of multilevel models fitted using a robust maximum likelihood estimator (Mplus, 5.21; Muthén & Muthén, Reference Muthén and Muthén2009). We regressed children's longitudinal cortisol levels on time-varying and time-invariant (i.e., mean) maternal sensitivity, age (months), and several time-varying and time-invariant control covariates. We subsequently tested whether the within-person effect of maternal sensitivity varied as a function of average-level maternal sensitivity by adding this cross-level interaction to the model.
Model constraints were fitted to test the simple slopes for statistically significant cross-level interactions. Specifically, within-person effects were tested at high (grand mean + 1 SD) and low (grand mean – 1 SD) mean levels of the same variable, testing, for example, whether the negative within-person effect of maternal sensitivity was particularly pronounced for those who experience low levels of maternal sensitivity, on average. All models were fitted using continuous variables: the simple slopes are merely conditional relations estimated from these models at field-typical high and low values in the (average) maternal sensitivity distribution.
To test specifically whether the direction of the within-person sensitivity effect differed for children with prototypically high versus prototypically low levels of maternal sensitivity, we fitted an additional model constraint in which the within-person sensitivity simple slope for those with high levels of mean maternal sensitivity (grand mean + 1 SD) was constrained to equal the inverse of within-person sensitivity simple slope for those with low levels of mean maternal sensitivity (grand mean – 1 SD). More simply, this addressed the question: in absolute terms, are the within-person sensitivity effects at either end of the mean maternal-sensitivity distribution statistically identical?
As specification checks, we also fitted nested models in which the respective effects of time-varying maternal sensitivity and time were allowed to vary randomly across children. All models comparisons were based on Satorra–Bentler (Satorra & Bentler, Reference Satorra and Bentler2001) adjusted likelihood-ratio tests. In addition, we tested Level 1 interactions between maternal sensitivity and age and income, and cross-level interactions between mean maternal sensitivity and time and income. Given that none of these alternative specifications improved model fit beyond levels expected by chance, we do not discuss these analyses further.
Results
Preliminary results
As shown in Table 1, on average, children's cortisol levels tended to decline between 7 and 24 months of age. Rank-order stability in children's cortisol levels was typically statistically significant, yet modest, ranging from 0.08 to 0.15. Maternal sensitivity showed moderate to strong rank-order stability; however, intraclass correlations also indicated a substantial amount of within-person variability (40%). Maternal sensitivity was correlated with children's cortisol levels in the hypothesized direction, such that lower levels of sensitivity were associated with higher cortisol levels, albeit typically in the modest to moderate range (e.g., r ~ .05 to .15).
Maternal sensitivity and HPA axis functioning across infancy and toddlerhood
Preliminary models suggested a statistically significant between-person relation, such that children with higher mean levels of maternal sensitivity tended to have lower mean cortisol levels, on average. No within-person maternal-sensitivity main effects were evident in these preliminary models (M1). As hypothesized, there was evidence of a cross-level interaction, such that, in an absolute sense, the within-person effect of maternal sensitivity was particularly pronounced for children who tended to experience either low levels or high levels of maternal sensitivity, on average. The effect was robust after adjusting for time-varying and time-invariant control covariates, as well as after weighting the model using IPWTs (M2; B = 0.20, p = .015).
As shown in the lower section of Table 2 (M2), there was a descriptive trend such that (a) the simple slope for the within-person relation between maternal sensitivity and children's cortisol levels was statistically zero for children whose average levels of maternal sensitivity over time were at the grand mean (B meansens = –0.04, p = .388), and (b) the absolute magnitudes of the within-person effects grew stronger as the distance from the sensitivity mean increased (i.e., positively or negatively). For children whose average levels of maternal sensitivity over time were low (grand mean – 1 SD; B lowsens = –0.17, p = .015) or very low (grand mean – 2 SD; B _Vlowsens = –0.31, p = .009), there was evidence of a negative within-person relation. In contrast, for children whose average levels of maternal sensitivity over time were high, there was evidence of positive within-person relation. This positive conditional effect only approached statistical significance for children with very high average levels of maternal sensitivity (grand mean + 2 SD; B Vhisens = –0.24, p = .056).Footnote 2
Table 2. A taxonomy of multilevel models testing the respective within- and between-person relations between maternal sensitivity with children's cortisol levels between 7 and 24 months of age (N = 1,292)

Note: The within-person maternal sensitivity effect was also adjusted for all the control covariates. However, they were excluded from the table for visual clarity; none were statistically significant. WP, Within person; BP, between person; Sens, maternal sensitivity; Inc, income to needs ratio; Daytime, time of day (hr) in which the saliva sample was taken; Med, child took medication; Year, year indicator, centered on 7 months; Married, married dummy; Ed, mother highest level of education; B.weight, child birth weight; AAmer, dummy for African American; NC, site dummy; Male, 1 = boy; M.Lit, mother functional literacy; Anx, mother anxiety symptoms; Dep, mother depression symptoms; Tmprmnt, child 7-month temperament; –2LL, –2 log likelihood.
†p < .10. *p < .05. **p < .01. ***p < .001.
To test the whether the magnitude of the within-person maternal sensitivity effect for children with typically high levels of maternal sensitivity (mean + 1 SD) differed in absolute terms from the within-person maternal sensitivity effect for children with typically low levels of maternal sensitivity (mean + 1 SD), we fitted a model constraint in which the former was constrained to be equal to the inverse of the latter (i.e., B hisens = [B lowsens x – 1]). Tests of nested models indicated that that the absolute magnitude of these slopes were statistically identical (Satorra–Bentler –2 log likelihood = 0.78, Δdf = 1, p = .38). As such, the simple slopes on either side of the maternal sensitivity grand mean were essentially mirror images of one another. We provide the fitted estimates from this more parsimonious constrained model in Table 2 (M3) and display the between-person and conditional within-person slopes in Figure 1 (time is held constant at 7 months, and all other variables are held at their respective grand means).

Figure 1. (Color online) Fitted between- and within-person relations between maternal sensitivity and basal cortisol between 7 and 24 months of age from the lowest to the highest absolute levels of maternal sensitivity. Y axis = 2 SD.
As shown by the dashed line representing the between-child relation, on average, children with higher mean levels of maternal sensitivity between the ages of 7 and 24 months of age tended to have lower cortisol levels across this period, relative to children with lower mean levels of maternal sensitivity (B = –0.07, p = .03). Scaling on the between-person variation in maternal sensitivity and children's cortisol levels, this corresponds to a modest standardized relation of approximately –0.19.
As displayed by the solid slopes in Figure 1, the magnitude and direction of the within-person maternal sensitivity effects differed for children who experienced higher versus lower levels of maternal sensitivity, on average, over this period. For children who typically experienced low (B lowsens = –0.14, p = .015) or very low (B Vlowsens = –0.28, p = .015) levels of maternal sensitivity, within-child increases in maternal sensitivity were associated with contemporaneous decreases in their characteristically high cortisol levels. Using the within-person standard deviations of maternal sensitivity and cortisol levels to calculate the standardized effects, these relations correspond to standardized simple slopes of approximately –0.09 and –0.16, respectively. Albeit modest in an absolute sense, they may be nontrivial substantively, because they suggest that within-person gains in maternal sensitivity may ameliorate longer term effects of historically low levels of maternal sensitivity during this period.
In contrast, the within-person maternal sensitivity simple slopes for children experiencing typically high (B Vhisens = 0.14, p = .015) or very high (B Vhisens = 0.28, p = .015) levels of maternal sensitivity were statistical mirror images of their low-maternal sensitivity counterparts On average, increases in maternal sensitivity from these children's already high levels of maternal sensitivity were associated with contemporaneous increases in their prototypically low cortisol levels. As above, these conditional relations reflect standardized simple slopes of approximately 0.09 and 0.16, respectively.
There was no evidence that the independent between-person or conditional within-person relations were curvilinear. Visual inspection of conditional scatterplots did not reveal any nonlinear trends (e.g., Figures A.1 and A.2 in Appendix A), and all tested quadratic effects were statistically nonsignificant. However, as displayed by the dotted curve in Figure 1, there was a descriptive indication of a curvilinear function when between- and within-child variation were considered collectively (i.e., total maternal sensitivity effect). Specifically, the dotted curvilinear line (in green online) represents an interpolated function that connects the lowest level of maternal sensitivity (i.e., Point A; those with low means levels, on occasions in which they are 1.5 SD lower than their low norm), moderate levels of maternal sensitivity (i.e., Point B; moderate levels of mean maternal sensitivity, in which the within-person effect is statistically 0), and the highest levels of maternal sensitivity (i.e., Point C; high mean levels, on occasions in which they are 1.5 SD higher than their high norm). That is, the function presents the estimated cortisol level from the lowest low level of (total) maternal sensitivity to the highest high levels of (total) maternal sensitivity. We adopted a 1.5 SD criterion for within-person sensitivity, because higher values would lead to extrapolation.
Collectively, there was a descriptive indication that, in absolute terms, the relation between maternal sensitivity and children's cortisol levels took the shape of a (reversed) J function. Statistically, a series of model constraints indicated that, although the estimated values at Points A and C were jointly greater than estimated value at Point B (Wald χ2 = 10.336, df = 2, p = .006), this joint difference was driven largely by the difference between Points A and B. Thus, although curvilinear, the relation only approximated a (reversed) J-shaped function.
Discussion
Theory and an emergent empirical literature suggest that young children's early experiences, most notably, their interactions with meaningful adults, shape their developing physiological stress systems (Gunnar & Donzella, Reference Gunnar and Donzella2002; Gunnar & Quevedo, Reference Gunnar and Quevedo2007; Hostinar, 2014). This may be particularly the case in the first years of life, when children require high levels of regulatory support (Feldman, Reference Feldman2007). Multiple theoretical models have recently highlighted the possibility that young children's early experiences may “tune” or “calibrate” the organization of the autonomic nervous system and HPA axis in fitness-relevant ways that lead some children to be more physiologically vigilant or sensitive to their experiences than others (Blair & Raver, Reference Blair and Raver2012; Boyce & Ellis, Reference Boyce and Ellis2005; Del Giudice et al., Reference Del Giudice, Ellis and Shirtcliff2011; Parker & Maestripieri, Reference Parker and Maestripieri2011). Specifically, the adaptive calibration model (Del Giudice et al., Reference Del Giudice, Ellis and Shirtcliff2011), informed by Boyce and Ellis (Reference Boyce and Ellis2005), posits that the relation between experience and physiological sensitivity is likely best represented as a U-shaped function. Highly unsupportive/uncaring (though not abusive and neglectful) and extremely supportive/caring environments are both theorized to calibrate the autonomic nervous system and HPA axis to be particularly vigilant and sensitive to subsequent experiential stimuli. In contrast, consistent with the idea of “steeling” or “stress inoculation,” those who experience normative environments marked by regular, but minor and surmountable stressors are theorized to show a less labile, buffered physiological profile. Our aim was to begin to test these hypotheses.
Between-child differences in maternal sensitivity across infancy and toddlerhood
Our findings indicated that, on average, children who experienced more sensitive caregiving between an average of 7 and 24 months tended to have lower mean cortisol levels over this period compared to those who experienced less sensitive care. In an absolute sense, the effect size was somewhat modest: a 1 SD difference in average maternal sensitivity was associated with an approximate 0.19 SD in children's cortisol levels. Note that in a relative sense, this relation is approximately one-third of the size of the standardized effect, one of the most well-known and biologically grounded causes of nonstimulated cortisol differences: time of day (i.e., β = 0.53). As such, this between-child association may well be substantively meaningful, despite its modest absolute size (McCartney & Rosenthal, Reference McCartney and Rosenthal2000).
This association between maternal sensitivity and children's basal cortisol levels is largely consistent with prior work considering between-person relations between maternal sensitivity and children's cortisol at different time points in the present sample (blinded, peer reviewed), as well as relations established in studies of infant attachment (see Gunnar & Donzella, Reference Gunnar and Donzella2002) and children's home contexts (Flinn & England, Reference Flinn and England1995). They also align with randomized control trials studying the effects of parenting interventions with infants and toddlers. Although the findings from this growing literature are somewhat mixed (see Slopen, McLaughlin, & Shonkoff, Reference Slopen, McLaughlin and Shonkoff2014), emerging work with infants and toddlers from high-risk contexts has shown that high-quality parenting may downregulate children's normative cortisol levels. For instance, Dozier and colleagues (Reference Dozier, Peloso, Lewis, Laurenceau and Levine2008) found that the infants and toddlers of foster parents assigned to an attachment-based parenting intervention had basal cortisol levels that were substantially lower than those assigned to a cognition/language-based intervention and statistically identical to a nonrisk comparison group. A parenting intervention aimed at the parents of infants with pre- or perinatal medical complications has shown quite similar impacts (e.g., Bugental, Schwartz, & Lynch, Reference Bugental, Schwartz and Lynch2010).
Unlike such randomized control trials, our study of natural variation does not support causal inferences. However, the reported between-child effects were robust after adjusting for IPWTs, which balanced a sizable number of potential confounding variables across levels of mean maternal sensitivity. If the assumptions underlying the IPWT approach are plausible (see online-only supplemental material), this balance functions much as if mean maternal sensitivity were randomly assigned. As such, although we make no causal claims, our methodological approach nonetheless likely strengthens the internal validity of our inferences.
Within-child differences in maternal sensitivity across infancy and toddlerhood
Beyond relations between traitlike aspects of children's maternal sensitivity levels across infancy and toddlerhood, we also found evidence that within-person deviations from these normative parenting levels were associated with contemporaneous changes in children's cortisol levels. Note that, largely consistent with our hypotheses, the magnitude (and direction) of these within-child effects varied as a function of children's mean levels of maternal sensitivity over time. Specifically, for children experiencing low levels of maternal sensitivity, on average, between 7 and 24 months, time-specific decreases from this already low level of maternal sensitivity were associated with contemporaneous cortisol increases. This largely aligns with the possibility that extended exposure to nonsupportive caregiving may calibrate the HPA axis to be more vigilant to changes in the psychological and physical environment. In addition, although it is the case that children with normatively low levels of maternal sensitivity fair the worst in the context of within-child decreases in maternal sensitivity, they also show the largest cortisol decreases in the context of within-child increases in maternal sensitivity. As such, this theorized “vigilance” seemingly functions in a “for better or for worse” manner, consistent with theory (Belsky & Pluess, Reference Belsky and Pluess2009; Boyce & Ellis, Reference Boyce and Ellis2005). On average, on occasions in which low-sensitivity children experience a 1 SD increase in their (typically low) sensitivity levels, their cortisol levels are statistically identical to those for children with typically high level of maternal sensitivity. That is, just as decreases in maternal sensitivity may exacerbate these children's already high cortisol levels, increases in sensitivity may also ameliorate them.
Consistent with the theorized buffered physiological profile, those with moderate mean levels of maternal sensitivity between 7 and 24 months tended to show moderate cortisol levels. However, within-child changes in maternal sensitivity had no impact on these children's cortisol levels. On average, these children tended to maintain their moderate cortisol levels, regardless of increases or decreases in maternal sensitivity over this period. Although the extant literature with respect to the effects of cortisol on cognition and behavior have been somewhat mixed (Alink et al., Reference Alink, van IJzendoorn, Bakermans-Kranenburg, Mesman, Juffer and Koot2008; Het, Ramlow, & Wolf, Reference Het, Ramlow and Wolf2005), these children's moderate cortisol levels may be somewhat optimal. Mounting theory and empirical evidence suggests that the relations between several hormones (e.g., cortisol, dopamine, and norepinephrine) and optimal cognitive functioning may be U-shaped (de Kloet, Oitzl, & Joels, Reference de Kloet, Oitzl and Joëls1999; Lupien et al., Reference Lupien, Maheu, Tu, Fiocco and Schramek2007; Vijayraghavan et al., Reference Vijayraghavan, Wang, Birnbaum, Williams and Arnsten2007).
These findings are largely consistent with predictions made by the ACM. Specifically, these authors conjectured that the anticipation and/or experience of affectively salient positive experiences may evoke physiological activation patterns that approximate those stimulated by negatively valenced stimuli, particularly for children with sensitive profiles for whom such positive experiences are the norm and negative experiences are rare. Although this idea has not been tested extensively, there is some indication that salient positive events, such as the anticipation of Christmas day (Flinn, Reference Flinn2006) or peer interactions for highly socially motivated children (Bruce et al., Reference Bruce, Davis and Gunnar2002; Davis et al., Reference Davis, Donzella, Krueger and Gunnar1999), can stimulate HPA axis activity. For young children who have experienced predominantly highly sensitive caregiving over time, positive changes in the rearing environment may function quite similarly. Specifically, like Christmas or peer engagement for highly socially motivated children, for young children who have historically experienced highly sensitive care, increases in sensitivity may serve as a cue for the child to prepare physiological and psychologically engage in the “good things to come.”
Of course, these conjectures remain questions to be tested directly. However, collectively, our findings are consistent with our more general hypotheses that the magnitude of within-person relations between maternal sensitivity and children's cortisol levels would be stronger for children experiencing levels of maternal sensitivity that are typically quite low or quite high, relative to those who typically experience more moderate levels.
There was no evidence of nonlinear relations between maternal sensitivity and adrenocortical functioning for the respective between- or within person relations when considered independently. However, there was at least a descriptive indication of nonlinear relation between total levels of maternal sensitivity and children's cortisol levels. For instance, when mean and time-specific maternal sensitivity level are combined, prototypical children with the lowest sensitivity levels (those with low mean levels, on occasions in which they are lower than even normal) tended to have comparatively higher cortisol levels than their peers with moderate levels of mean sensitivity. Consistent with a (reverse) J-shaped function, there was a descriptive indication that the same was true for those with the highest levels of maternal sensitivity. That is, on occasions in which children who typically experienced high levels of maternal sensitivity had an additional “bump” in maternal sensitivity above their already high levels, they tended to show cortisol levels that were somewhat higher than those of children typically experiencing moderate maternal sensitivity. This specific comparison failed to reach traditional levels of statistical significance. As such, the exact functional form of the curvilinear relation between total maternal sensitivity and children's cortisol levels remains somewhat unclear.
This highlights a difficulty inherent in testing theoretical models that require the full range of a given experiential distribution: characteristics of the sample may truncate the tails of these distributions, making it impossible to model the entirety of the theorized effect. Heterogeneity in the populations sampled may partially explain the notable variability in the magnitudes and even directions of the effects across studies (Boyce & Ellis, Reference Boyce and Ellis2005). This may be the case presently, given our predominantly low-income sample. The cumulative stressors of economic adversity may trickle down to affect the quality of parent–child interactions, on average. As such, this might shift the entire maternal sensitivity distribution downward compared to low-risk families, truncating the highest levels of maternal sensitivity in our sample. This, of course, remains an open question.
However, the ACM actually includes a fourth unemotional physiological profile reflecting either hypo- or hyperphysiological reactivity to context. This group was intentionally omitted from our discussion because our sample, although low income, was not representative of the more extreme negative environments thought to underlie this physiological profile.
Limitations and conclusions
Despite our aim to strengthen internal validity, neither the reported propensity-score weighted between-person effects nor the within-person child-fixed effects can be interpreted causally. However, it is noteworthy that, consistent with causal logic, within-person changes in maternal sensitivity were associated with changes in children's cortisol levels. In addition, the within-person effects (by design) adjust for all potential time-invariant confounds. Nonetheless, time-varying confounds may have biased our estimates.
Given the scope of this in-depth, longitudinal study, we were unable to collect multiple saliva samples over the course of a day or across several consecutive days, which would both strengthen the reliability of our cortisol measures and be more directly informative with regard to children's diurnal cortisol patterns (Hellhammer et al., Reference Hellhammer, Fries, Schweisthal, Schlotz, Stone and Hagemann2007). Rather, our cortisol measures were based on samples collected modally around 10:00 a.m., when the child was at rest, after he or she had been given time to acclimate to the RAs in the home and prior to engaging the child in a broader battery of tasks. There was little behavioral evidence that the children were particularly stressed by the visitors’ presence. This was supported by maternal interviews suggesting that the RA visits were unassociated with children's moods. Maternal ratings of their children's general mood during the visit were also unassociated with children's cortisol levels. These observations align with the extant literature, which indicates that exposure to novel adults characteristically fails to elicit a cortisol response in infancy and toddlerhood (even when that is the intended aim; Gunnar et al., Reference Gunnar, Talge and Herrera2009). However, we cannot rule out the possibility that our cortisol measures partially tapped reactivity to the RA's presence.
Another limitation concerning our cortisol measure is that our within-person design limited our analyses to considering only children's basal levels, rather than their acute HPA axis reactivity. The ACM makes predictions about acute HPA reactivity that are quite similar to those for children's basal or “resting” levels. Establishing consistent findings across these two aspects of HPA axis functioning would bolster support for the plausibility of the model and remains a pressing question for future work.
It is also worth noting that terms like ameliorate with respect to the negative relation between time-varying maternal sensitivity and cortisol levels (for children with typically low levels of maternal sensitivity) implicitly imbue high cortisol levels with a substantively negative meaning. Similar implicit interpretations are embedded in terms like buffer. To be clear, our interpretations are based on a relative scale in which lower, perhaps especially moderate, cortisol levels have been found to be associated with more effective cognitive functioning (Blair et al., Reference Blair, Granger, Willoughby, Mills-Koonce, Cox and Greenberg2011) and comparatively more socially desirable behavior in early childhood (Alink et al., Reference Alink, van IJzendoorn, Bakermans-Kranenburg, Mesman, Juffer and Koot2008). They do not reflect absolute values, for instance, with respect to successful adaptation to context (which may well contradict general social norms). Given that very low or “blunted” cortisol levels have also been linked to less optimal cognitive and social outcomes in older children (Alink et al., Reference Alink, van IJzendoorn, Bakermans-Kranenburg, Mesman, Juffer and Koot2008; Lupien et al., Reference Lupien, Maheu, Tu, Fiocco and Schramek2007), an alternative interpretation of the present findings could be that lower cortisol levels are “worse” than higher levels. Although possible, we are hesitant to adopt this latter hypocortisolism interpretation. In particular, it seems fairly inconsistent with the present and prior findings. For instance, in the present study, this would suggest that moderate levels of maternal sensitivity are more detrimental to broader developmental outcomes than are low levels, an interpretation contrary to theory and a well-developed empirical literature. Nevertheless, the broader implications of the present findings, with respect to their potential secondary effects on cognition and behavior, remain open questions that we intend to address in subsequent work.
Despite these limitations and areas of future study, the present findings add to a growing literature suggesting that children's social experiences play a meaningful role in adrenocortical-system functioning very early in life. Specifically, long-term aspects of children's experiences of maternal sensitivity were associated with meaningful differences in the extent to which within-child changes in maternal sensitivity are linked to changes in HPA axis functioning. Consistent with contemporary theoretical models of physiological adaptation, in absolute terms, children who experienced either very low or very high levels of maternal sensitivity on average between 7 and 24 months showed notable within-person effects of maternal sensitivity: changes in maternal sensitivity were associated with contemporaneous changes in children's cortisol levels. In contrast, no within-person relations were evident for those who experienced moderate levels of maternal sensitivity, on average, across this period. Collectively, these findings are largely consistent with the idea that children's typical social experiences with their primary caregivers may lead to meaningful individual differences in their physiological sensitivity to changes in these experience over time.
Supplementary Material
To view the supplementary material for this article, please visit http://dx.doi.org/10.1017/S0954579416000158.
Appendix A

Figure A.1. Conditional scatterplot displaying the within-person relation between maternal sensitivity and basal children's cortisol levels for those with very low levels, mean – (≤2 SD), of maternal sensitivity, on average, between 7 and 24 months of age.

Figure A.2. Conditional scatterplot displaying the within-person relation between maternal sensitivity and children's basal cortisol levels for those with very high levels, mean + (≥2 SD), of maternal sensitivity, on average, between 7 and 24 months of age.







