


“Developmental psychopathology is an evolving scientific discipline whose predominant focus is elucidating the interplay among the biological, psychological, and social–contextual aspects of normal and abnormal development across the life span” (Cicchetti, Reference Cicchetti, Cicchetti and Cohen2006). There are few situations during the life course where the interplay between biology and environment (i.e., gene–environment interactions) are more striking than during prenatal development. During this important period, “critical windows” are narrow and certain disturbances may alter fetal growth and development, leading to health and behavioral consequences that manifest and possibly persist across the life course (Maccani & Marsit, Reference Maccani and Marsit2009). This has been suggested by a variety of studies that are summarized below.
Intrauterine Development: An Important Critical Window
A number of epidemiological studies have explored the links between adverse prenatal conditions and increased risk for diseases, health problems, and psychological outcomes later in life. Such studies have examined historical examples, such as the Dutch Famine Birth Cohort, which consists of men and women born as term singletons in Amsterdam, The Netherlands (de Rooij, Wouters, Yonker, Painter, & Roseboom, Reference de Rooij, Wouters, Yonker, Painter and Roseboom2010; Stein, Susser, Saenger, & Marolla, Reference Stein, Susser, Saenger and Marolla1972). The formation of the Dutch Famine Birth Cohort, resulting from the Dutch Famine of 1944–1945, provided researchers with an opportunity to study the particular adverse exposure of starvation during pregnancy and a number of health and developmental outcomes potentially associated with this challenging adverse exposure. Associations have been reported between adverse intrauterine environment as influenced by famine and a number of diseases and conditions, such as increased risk for Type 2 diabetes mellitus, cardiovascular disease, and other metabolic disorders and decreased cognitive function later in life (Argente, Mehls, & Barrios, Reference Argente, Mehls and Barrios2010; Barker & Clark, Reference Barker and Clark1997; de Rooij et al., Reference de Rooij, Wouters, Yonker, Painter and Roseboom2010). In addition, researchers found that maternal weight loss or moderate to low weight gain was significantly associated with infant birth weight, length, and other measures of fetal growth status, as well as with trimester of exposure to famine (Stein, Ravelli, & Lumey, Reference Stein, Ravelli and Lumey1995). In terms of psychological outcomes, increased risk of affective disorders has been found in males exposed to famine during their second trimester (Brown, Susser, Lin, Neugebauer, & Gorman, Reference Brown, Susser, Lin, Neugebauer and Gorman1995). Findings such as these underscore the importance of considering timing of prenatal exposure to adverse conditions, such as famine, as well as potential confounding elements, such as the gender of the infant, when looking at outcomes across development. Birth cohorts such as the Dutch Famine Birth Cohort have provided researchers with many of the earliest tools necessary to investigate epidemiological associations between adverse intrauterine conditions and postnatal health and disease.
Maternal Cigarette Smoking During Pregnancy
We now move beyond the example of famine to consider one of the most common, potentially hazardous environmental exposures during pregnancy: maternal cigarette smoking. Studies have shown that there are more than 4,000 chemicals in cigarette smoke, including benzo(a)pyrene, nicotine, and carbon monoxide, and more than 40 of these chemicals are known carcinogens (Thielen, Klus, & Muller, Reference Thielen, Klus and Muller2008; US Department of Health and Human Services, 2010). Nicotine crosses the placenta, and fetal concentrations of nicotine can be 15% higher than maternal concentrations (Lambers & Clark, Reference Lambers and Clark1996). Despite a number of studies showing a decrease in the overall prevalence of smoking in women in the past 20 years, key studies have suggested that the prevalence of smoking in young pregnant women under 20 years of age has increased, with prevalence rates of 30%–40% (Jaakkola, Jaakkola, Gissler, & Jaakkola, Reference Jaakkola, Jaakkola, Gissler and Jaakkola2001; Mohsin & Bauman, Reference Mohsin and Bauman2005; US Department of Health and Human Services, 2004). Others have reported that 12%–15% of all women of childbearing age smoke while pregnant (Cnattingius, Reference Cnattingius2004; Goodwin, Keyes, & Simuro, Reference Goodwin, Keyes and Simuro2007). Considering that women who smoke during pregnancy are more likely to be nicotine dependent, less likely to quit, and have a partner who smokes (Agrawal et al., Reference Agrawal, Knopik, Pergadia, Waldron, Bucholz and Martin2008; Knopik et al., Reference Knopik, Sparrow, Madden, Bucholz, Hudziak and Reich2005), it is important to also keep the effects of second-hand smoke exposure in mind, which are difficult to tease apart from direct prenatal primary smoke exposure. Women who are pregnant may also be exposed to second-hand smoke in homes, vehicles, the workplace, or public areas. More than 126 million nonsmoking adults continue to be exposed to second-hand smoke and current estimates suggest that almost 60% of children, aged 3–11, are exposed to second-hand smoke (US Department of Health and Human Services, 2006). Thus, exposure to cigarette smoke remains a common and hazardous in utero exposure.
Previous work has suggested that maternal cigarette smoking during pregnancy is associated with increased risk for spontaneous abortion (Castles, Adams, Melvin, Kelsch, & Boulton, Reference Castles, Adams, Melvin, Kelsch and Boulton1999), preterm delivery (Castles et al., Reference Castles, Adams, Melvin, Kelsch and Boulton1999; Kaddar et al., Reference Kaddar, Rouault, Chien, Chebel, Gadoux and Salles2009; Shah & Bracken, Reference Shah and Bracken2000), respiratory disease (Cook & Strachan, Reference Cook and Strachan1999), immune system difficulties such as asthma and allergies (Prescott & Clifton, 2009), and cancer later in life (Doherty, Grabowski, Hoffman, Ng, & Zelikoff, Reference Doherty, Grabowski, Hoffman, Ng and Zelikoff2009). The results also suggest that there are a variety of placental complications linked to prenatal exposure to cigarette smoke, including alterations to the development and function of the placenta (Einarson & Riordan, Reference Einarson and Riordan2009).
A number of studies have suggested that prenatal tobacco exposure is associated with a number of serious neurodevelopmental and behavioral consequences in infants, children, and adolescents. For example, studies in infants have shown that maternal smoking during pregnancy is associated with delayed psychomotor and mental developmental scores (as measured by the Bayley Scales of Infant Development; Kiechl-Kohlendorfer et al., Reference Kiechl-Kohlendorfer, Ralser, Pupp Peglow, Reiter, Griesmaier and Trawoger2010). Kable, Coles, Lynch, and Carroll (Reference Kable, Coles, Lynch and Carroll2009) analyzed the impact of maternal smoking during pregnancy on auditory brainstem responses and determined that, after controlling for a number of potential confounders, maternal smoking during pregnancy was negatively related to auditory brainstem response latency in infants. It should be noted that because alterations in infants’ auditory processing have been previously found to be highly predictive of future reading and language difficulties, these findings suggest that maternal cigarette smoking during pregnancy may negatively affect a child's speech and language development (Benasich & Tallal, Reference Benasich and Tallal2002; Kable et al., Reference Kable, Coles, Lynch and Carroll2009; Molfese, Reference Molfese2000).
There are many examples of prenatal smoking exposure research in childhood (for a review, see Knopik, Reference Knopik, Jacob, Haber, Swenson and Howell2009), perhaps because of more sensitive assessments for this age group, relative to infants. For instance, Olds, Henderson, and Tatelbaum (Reference Olds, Henderson and Tatelbaum1994) conducted a study investigating associations between maternal cigarette smoking during pregnancy and offspring intelligence at age 4 and found that children whose mothers smoked 10 or more cigarettes per day during their pregnancy exhibited intellectual impairment relative to children whose mothers did not smoke cigarettes during pregnancy. Huijbregts, Seguin, Zoccolillo, Boivin, and Tremblay (Reference Huijbregts, Seguin, Zoccolillo, Boivin and Tremblay2007, Reference Huijbregts, Seguin, Zoccolillo, Boivin and Tremblay2008) found associations between maternal cigarette smoking during pregnancy and physical aggression during early childhood. This is consistent with multiple studies suggesting an association of prenatal smoke exposure and increased externalizing disorders, such as conduct disorder and attention-deficit/hyperactivity disorder (Abbott & Winzer-Serhan, Reference Abbott and Winzer-Serhan2012; Cornelius & Day, Reference Cornelius and Day2009; Knopik, Reference Knopik2009). Cognitive function has also been shown to be negatively affected by maternal smoking during pregnancy, with deficits in sustained attention (Fried, O'Connell, & Watkinson, Reference Fried, O'Connell and Watkinson1992; Fried, Watkinson, & Gray, Reference Fried, Watkinson and Gray1992), response inhibition, memory, and impulsivity, overall cognitive function, receptive language (Fried, O'Connell et al., 1992), verbal learning and design memory (Cornelius, Ryan, Day, Goldschmidt, & Willford, Reference Cornelius, Ryan, Day, Goldschmidt and Willford2001), problem solving (Cornelius et al., Reference Cornelius, Ryan, Day, Goldschmidt and Willford2001), speech and language (Makin, Fried, & Watkinson, Reference Makin, Fried and Watkinson1991), school performance (Lambe, Hultman, Torrang, Maccabe, & Cnattingius, Reference Lambe, Hultman, Torrang, Maccabe and Cnattingius2006), and auditory processing (McCartney, Fried, & Watkinson, Reference McCartney, Fried and Watkinson1994). Dose–response relationships, in which the smoking-related relative risk increases with the amount smoked, have also been reported for general cognitive ability (Sexton, Fox, & Hebel, Reference Sexton, Fox and Hebel1990), arithmetic, and spelling (Batstra, Hadders-Algra, & Neeleman, Reference Batstra, Hadders-Algra and Neeleman2003), suggesting the presence of vulnerable periods during fetal development (Ernst, Moolchan, & Robinson, Reference Ernst, Moolchan and Robinson2001).
Recent research has also suggested that second-hand smoke exposure during pregnancy is associated with a decrease in infant mental development index score and an increase in the risk of developmental delay in the infant (Lee et al., Reference Lee, Hong, Park, Ha, Kim and Chang2011), thereby extending the risk profile related to smoking exposure beyond just direct, primary smoke exposure during pregnancy. Overall, these observations suggest that smoke exposure during pregnancy remains a common exposure that can have major ramifications on the in utero growth and development of the fetus. Moreover, due to the plethora of scientific data suggesting negative consequences associated with smoking during pregnancy, pregnant women are cautioned against smoking while pregnant (Cornelius & Day, Reference Cornelius and Day2009; Shea & Steiner, Reference Shea and Steiner2008). However, despite the numerous research articles suggesting undesirable outcomes in children exposed to prenatal smoke exposure and warnings encouraging women to stop smoking while pregnant, the underlying biological processes in humans are not well understood. Thus, we turn our attention now to describing one potential piece of this puzzle, namely, fetal programming and how this may play an important role in future child development.
Fetal Programming
What is meant by fetal programming? The theory of fetal programming has been described as a model of gene–environment interaction that explains the influence of the in utero environment on the molecular character of development (Barker & Clark, Reference Barker and Clark1997; Hales & Barker, Reference Hales and Barker1992). One of the forefathers of this theory is David Barker. Starting in the 1980s and 1990s, he proposed and tested the hypothesis that an adverse fetal nutritional environment but plentiful food in adulthood might be a major factor associated with a number of adult diseases (Schulz, Reference Schulz2010). The Barker hypothesis further hypothesized that adverse intrauterine conditions may result in a negative, or poor, maternal forecast, commonly manifested in small for gestational age status or reduced infant birth weight. This poor maternal forecast “predicts” that the child will be born into a postnatal environment in which resources are scarce; thus, the child has been forecasted or programmed to thrive in such a poor environment. Poor maternal forecasts can prove incorrect if a child is born into what is or soon becomes a nutrient-rich environment.
We emphasize that current research expands the notion of an adverse intrauterine environment beyond the traditional, nutrient-poor environment first elucidated by Barker and colleagues in their analysis of infants from the Dutch Famine Birth Cohort (Barker & Clark, Reference Barker and Clark1997; Hales & Barker, Reference Hales and Barker1992). Adverse intrauterine conditions may also be the result of exposure to viruses (e.g., influenza); increased levels of stress during pregnancy; and maternal cigarette smoking and second-hand (passive) smoke exposure during pregnancy, which is of the most importance for this article. In this expanded view of adverse intrauterine conditions and the resulting effects on the fetus, an exposure such as prenatal exposure to cigarette smoke may lead to a negative maternal forecast for the fetus. Although prenatal smoke exposure certainly contributes to this adverse environment, evidence that maternal cigarette smoking during pregnancy is correlated with other potential contributors to adverse in utero environmental conditions can make causal attribution difficult (for a review, see Knopik et al., Reference Knopik, Jacob, Haber, Swenson and Howell2009). This is supported by recent work from our own group and others suggesting that maternal smoking during pregnancy is correlated with many risk factors, such as lower levels of maternal education (D'Onofrio, Singh, Iliadou, Lambe, Hultman, Neiderhiser, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Grann2010), spousal/significant other substance dependence (Knopik et al., Reference Knopik, Sparrow, Madden, Bucholz, Hudziak and Reich2005, Reference Knopik, Heath, Jacob, Slutske, Bucholz and Madden2006), nicotine dependence (Agrawal et al., Reference Agrawal, Knopik, Pergadia, Waldron, Bucholz and Martin2008), as well as maternal attention-deficit/hyperactivity disorder (ADHD) and other psychopathology (D'Onofrio, Singh, Iliadou, Lambe, Hultman, Neiderhiser, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Neiderhiser2010; Huizink & Mulder, Reference Huizink and Mulder2006; Knopik, Reference Knopik2009; Knopik et al., Reference Knopik, Jacob, Haber, Swenson and Howell2009), which may come with a host of additional influences on intrauterine environment that also predict poor offspring performance.
Figure 1 summarizes the concept of fetal programming and the contributions of the variety of stimuli that make up the intrauterine environment on maternal forecasts. As shown in the figure, in utero stimuli include many exposures that may influence the intrauterine environment, such as exposure to nutrients, stress, drugs, trauma, and smoking. A largely positive and healthy intrauterine environment may result in the mother imparting a rich maternal forecast on her developing fetus, predicting a rich postbirth environment in which resources are predicted to be plentiful and negative exposures are predicted to be at a minimum. A relatively negative or adverse intrauterine environment may result in the mother imparting a poor maternal forecast on her developing fetus, sometimes characterized by a “thrifty phenotype” (Hales & Barker, Reference Hales and Barker1992) or small for gestational age status, thereby preparing her child to survive in a poor postbirth environment, where resources are predicted to be scarce and/or negative postnatal exposures to be frequent and abundant. Maternal forecasts that do not accurately predict the postbirth environment have been hypothesized to lead to negative consequences for the health of the child over its life course, such as increased risk for metabolic diseases and decreased cognitive functioning in offspring that had been given a poor maternal forecast but were born into a rich environment (Argente et al., Reference Argente, Mehls and Barrios2010; Barker & Clark, Reference Barker and Clark1997; de Rooij et al., Reference de Rooij, Wouters, Yonker, Painter and Roseboom2010; Hochberg et al., Reference Hochberg, Feil, Constancia, Fraga, Junien and Carel2010).
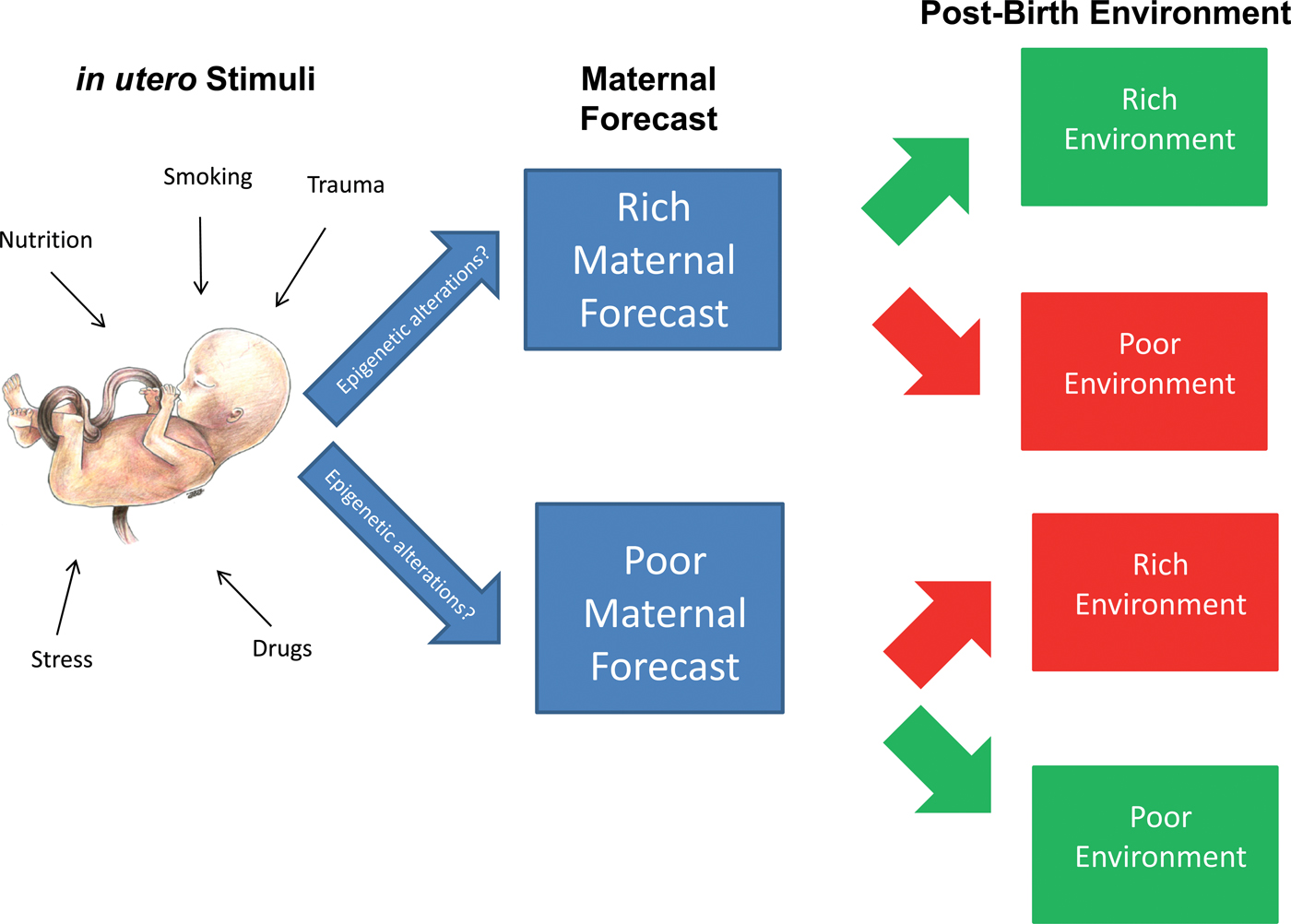
Figure 1. In utero stimuli-associated epigenetic alterations and maternal forecasts. In utero stimuli comprise the many exposures that may characterize the intrauterine environment, including exposure to nutrients, stress, and drugs. A largely positive and healthy intrauterine environment may result in the mother imparting a rich maternal forecast on her developing fetus, predicting a rich postbirth environment in which resources are predicted to be plentiful and negative exposures at a minimum level. A relatively negative or adverse intrauterine environment may result in the mother imparting a poor maternal forecast, often characterized by a “thrifty phenotype,” on her developing fetus, thereby preparing her child to survive in a poor postbirth environment in which resources are predicted to be scarce and/or negative exposures (e.g., second-hand smoke exposure) to be frequent and abundant. Maternal forecasts that do not accurately predict the postbirth environment have been hypothesized to lead to negative consequences for the health of the child over its life course, such as increased risk for metabolic diseases in offspring born into a rich environment with a poor maternal forecast. In utero stimuli may influence maternal forecasting through epigenetic mechanisms, both directly and indirectly. These epigenetic-mediated maternal forecasts may be accessible to measurement through techniques established for measuring changes to epigenetic modes of regulation. In addition, these epigenetic marks, such as DNA methylation profiles or miRNA expression, may have utility as diagnostic biomarkers capable of predicting increased risk for diseases or disease progression and as therapeutic targets. Scientific artwork by Jennifer Z. Joukhadar. [A color version of this figure can be viewed online at http://journals.cambridge.org/dpp]
Despite the large literature suggesting undesirable outcomes in children exposed to maternal smoking during pregnancy, the underlying biological processes in humans are not well understood. Work is under way to better understand the underlying mechanisms of fetal programming, and many researchers have focused on how epigenetic mechanisms may play a role in mediating the effects of environmental exposures on future outcomes, such as the physical and behavioral health of the individual in infancy, childhood, and adulthood (Maccani & Marsit, Reference Maccani and Marsit2009). Understanding how fetal programming may lead to future health consequences have caused some to adopt an even more expanded theory of fetal programming, namely the “developmental origins of health and disease” (Barker, Reference Barker2004; Gillman, Reference Gillman2005).
Epigenetics: Mechanisms by Which Fetal Programming May Occur
The study of epigenetics, or the study of changes in gene expression that are not caused by changes in the sequence of DNA (Bird, Reference Bird2007), has intrigued scores of scientists over the past few decades. It is these epigenetic mechanisms that can influence whether one's genes are switched on or off (i.e., gene expression). It may be through epigenetic mechanisms that environmental factors like diet, stress, prenatal nutrition, or prenatal drug exposure can lead to changes in gene expression from one cell to its daughter cells and, in some cases, from one generation to the next. Work in this area has focused on examining four main modes of epigenetic gene regulation: DNA methylation, imprinting, histone modification, and noncoding RNA-mediated gene regulation, especially by microRNA (miRNA). (For a more comprehensive review on these four modes of epigenetic regulation, as well as on the technological advances that have made it possible to measure such changes, see Maccani & Marsit, Reference Maccani and Marsit2009.)
As hinted above, new subfields have emerged to explore epigenetic effects in a variety of settings. The field of “environmental epigenetics” studies how environmental exposures affect epigenetic mechanisms, which was described by Reamon-Buettner, Mutschler, and Borlak (Reference Reamon-Buettner, Mutschler and Borlak2008). Many researchers are interested in uncovering how environmental exposures at sensitive periods of development, such as maternal cigarette smoking during pregnancy, might influence epigenetics and thus affect the developing fetus and fetal programming. Research in both human cohorts and model systems (e.g., mice) continues to characterize the influence of environmental exposures on epigenetics. Figure 1 describes how certain prenatal exposures may, both directly and indirectly, influence maternal forecasting through epigenetic mechanisms. In addition, these epigenetic mechanisms, such as DNA methylation profiles or miRNA expression, may have utility not only as diagnostic biomarkers capable of predicting increased risk for behavioral deficits, diseases, or disease progression, but also as therapeutic targets. This is similar to recent reports using gene-expression differences, which were identified via a blood test, to predict youth at risk for early onset depression (Pajer et al., Reference Pajer, Andrus, Gardner, Lourie, Strange and Campo2012).
As we highlight in this article, the research community has placed particular focus on investigating the influence of maternal cigarette smoking during pregnancy on DNA methylation patterns, whereas work on the effects of maternal cigarette smoking during pregnancy on miRNA has been much less comprehensive. Investigations of specific associations between prenatal smoke exposure and histone modifications or imprinting have yet to be described in the literature, but others have cited these two modes of epigenetic regulation as important future considerations for research in the context of in utero smoke exposure (Swanson, Entringer, Buss, & Wadhwa, Reference Swanson, Entringer, Buss and Wadhwa2009).
Maternal Cigarette Smoking During Pregnancy Is Associated With Atypical DNA Methylation Patterns
DNA methylation is one of the more widely studied and well characterized of the main modes of epigenetic regulation. DNA methylation of cytosine residues is performed by one of a number of DNA methyltransferases that add a methyl group to a specific cytosine residue. These cytosine residues often reside in cytosine- and guanine-rich stretches of DNA called cytosine nucleotide–phosphate–guanine nucleotide (CpG) islands. Sometimes DNA methylation is also referred to as CpG methylation. In general, a DNA methylation-regulated gene whose promoter region shows a great degree of methylation will be effectively silenced. When the same gene's promoter region is not methylated (i.e., the promoter is hypomethylated), the gene will likely not be silenced, and thus the gene will be expressed. Research has determined that the blocking of transcription in a methylated gene is not due to the methylation of DNA alone, but rather due to the irregular binding of a variety of proteins. In the presence of DNA methylation, proteins that normally bind DNA and enable transcription to proceed are unable to bind as well or at all, which effectively limits or stops transcription. Research is continuing to develop a better understanding of how DNA methylation in a gene's promoter region controls the complex regulatory environment necessary for transcription and, thus, gene expression.
During the period of embryonic development, methylation patterns of the germline and somatic cell lineages are established (Maccani & Marsit, Reference Maccani and Marsit2009). During the cleavage phase (i.e., the early cell divisions that occur as a fertilized egg begins to develop into an embryo), methylation in the zygote's genome is almost completely removed. After implantation, as the cells produced during the cleavage phase begin to organize themselves (i.e., gastrulation), the organism's methylation patterns are reestablished by de novo methylation (Jaenisch, Reference Jaenisch1997; Kafri et al., Reference Kafri, Ariel, Brandeis, Shemer, Urven and McCarrey1992; Monk, Boubelik, & Lehnert, Reference Monk, Boubelik and Lehnert1987; Razin & Shemer, Reference Razin and Shemer1995). Such patterning and repatterning of methylation marks also occurs in trophoblast lineages (i.e., the various specialized cells comprising the placenta; Jaenisch, Reference Jaenisch1997). Proper setting and resetting of methylation marks throughout development is crucial for the proper health and development of the embryo.
Recent studies have even reported initial findings suggesting that an additional mode of epigenetic regulation, namely, conversion of 5-methylcytosine to 5-hydroxymethylcytosine, may play a role in the reprogramming of the zygote (Wossidlo et al., Reference Wossidlo, Nakamura, Lepikhov, Marques, Zakhartchenko and Boiani2011). Further studies will be necessary to understand more completely the implications of these findings on the patterning of methylation marks during development, as well as the effect of exposures on the underlying mechanisms.
When considering epigenetic research in general, DNA methylation is the most heavily studied mode of epigenetic regulation (Bird, Reference Bird2007). Thus, it is not surprising that the majority of published articles on the epigenetics of maternal cigarette smoking during pregnancy describe associations with DNA methylation. A number of studies have characterized associations between prenatal smoke exposure and aberrant DNA methylation patterns in a variety of tissues. The following sections describe many of these efforts.
DNA methylation and the placenta
One of the most important functional organs critical to the survival and in utero development of the fetus is the placenta. The placenta provides nutrients, assists in the transfer of waste for ultimate excretion by the mother, and plays an important role in protecting the fetus from maternal immune system attack (Maccani & Marsit, Reference Maccani and Marsit2009). Another key function of the placenta is to secrete hormones that regulate pregnancy stages and protect the fetus, when possible, from harmful xenobiotic (i.e., foreign) exposures, such as maternal drug use during pregnancy (Sood, Zehnder, Druzin, & Brown, Reference Sood, Zehnder, Druzin and Brown2006). All of these functions, as well as placental gene expression, which is modulated by epigenetic regulation, can be affected by environmental insults (Guo et al., Reference Guo, Choufani, Ferreira, Smith, Chitayat and Shuman2008; Sood et al., Reference Sood, Zehnder, Druzin and Brown2006). Therefore, many consider the placenta an important and relatively accessible record or history of in utero exposure and pathology (Maccani & Marsit, Reference Maccani and Marsit2009).
Important findings using the human placenta have revealed associations between maternal cigarette smoking during pregnancy and DNA methylation in a gene specific and even global fashion. Suter et al. (Reference Suter, Ma, Harris, Patterson, Brown and Shope2011) observed that maternal tobacco use is associated with aberrant placental epigenome-wide DNA methylation and gene expression. Their work also suggested that maternal smoking during pregnancy is associated with altered site-specific CpG methylation, which further correlates with important changes in gene expression in pathways crucial for ensuring proper growth and development. In a separate study, Suter et al. (Reference Suter, Abramovici, Showalter, Hu, Shope and Varner2010) also noted that maternal tobacco smoking may modify placental cytochrome P450, family 1, subfamily A, polypeptide 1 (CYP1A1) expression by altering methylation at CpG sites proximal to the 5′-xenobiotic response element transcription factor binding site. CYP1A1 is involved in Phase I of the metabolism of carcinogenic compounds such as polycyclic aromatic hydrocarbons, which are present in cigarette smoke. Specifically, CYP1A1 converts such compounds in a way that can eventually be excreted in Phase II metabolism, a process modulated in part by other genes (Suter et al., Reference Suter, Abramovici, Showalter, Hu, Shope and Varner2010). Suter et al. found that methylation at these sites was significantly lower in the placentas of babies born to mothers who smoked during pregnancy versus nonsmoking controls. This downregulation of methylation was also significantly correlated with increased placental CYP1A1 expression, a finding that may have substantial potential implications for future behavior. Wilhelm-Benartzi et al. (Reference Wilhelm-Benartzi, Houseman, Maccani, Poage, Koestler and Langevin2011) showed that differential methylation of repetitive elements (i.e., stretches of DNA exhibiting a large number of repeated bases) in the placenta is associated with birth weight percentile and maternal smoking during pregnancy. Specifically, they found that mean methylation levels of the repetitive element AluYb8 significantly differed by maternal tobacco use during pregnancy.
Overall, these reports suggest that cigarette smoke may elicit some of its downstream consequences on the placental epigenome in both a global- and site-specific fashion (Suter et al., Reference Suter, Abramovici, Showalter, Hu, Shope and Varner2010, Reference Suter, Ma, Harris, Patterson, Brown and Shope2011). Moreover, repetitive elements may have specific methylation patterns influenced by environmental exposures, underscoring the need to carefully consider how “globally representative” a particular repetitive element's methylation pattern may be without utilizing a representative genome-wide methylation assessment for comparison (Wilhelm-Benartzi et al., Reference Wilhelm-Benartzi, Houseman, Maccani, Poage, Koestler and Langevin2011). These findings in the human placenta build on work described by Breton et al. (Reference Breton, Byun, Wenten, Pan, Yang and Gilliland2009) in human buccal cells whereby they described that in utero exposure to tobacco smoke is associated with alterations to global and gene-specific methylation.
Umbilical cord blood and maternal blood
Others have investigated changes in DNA methylation associated with maternal cigarette smoking during pregnancy in umbilical cord serum. Guerrero-Preston et al. (Reference Guerrero-Preston, Goldman, Brebi-Mieville, Ili-Gangas, Lebron and Witter2010) tested their hypothesis that global DNA hypomethylation in serum from the umbilical cord is associated with prenatal exposure to maternal smoking and perflouroalkyl compounds, which are used in a wide-range of consumer and industrial products, such as stain-resistant coatings. They determined that global DNA methylation was most reduced in cord blood from newborns with mothers who smoked during pregnancy. Future studies using larger sample sets, as well as utilizing other surrogates of “global” DNA methylation, such as the repetitive elements long interspersed nuclear element 1 or AluYb8, or DNA methylation microarrays, are needed in order to assess the possibility of utilizing DNA methylation status in cord blood as a biomarker for prenatal smoke exposure.
Associations between nutrient levels during pregnancy and DNA methylation have also been assessed using blood. A number of previous groups have described importance of folate as a key nutrient involved in the process of establishing and maintaining DNA methylation (Chmurzynska, Reference Chmurzynska2010). Ba et al. (Reference Ba, Yu, Liu, Geng, Zhu and Zhu2011) used both maternal blood and umbilical cord blood to measure specific gene promoter methylation in insulin-like growth factor 2, which is believed to be a major growth factor in the developing fetus. They found that whereas promoter 2 and promoter 3 methylation in cord blood was not significantly associated with folate levels in either cord blood or maternal blood, promoter 3 methylation was significantly associated with vitamin B12 serum levels in maternal blood. Furthermore, promoter 2 methylation in mother's blood was associated with vitamin B12 maternal blood serum levels, passive smoke exposure, and maternal weight gain during pregnancy. Although not focused on smoking during pregnancy per se, the investigators did consider passive smoke exposure, which is an indication of second-hand smoke exposure during pregnancy. Moreover, their study underscores the importance of environmental exposures and maternal dietary factors in potentially mediating methylation patterns in specific biological pathways.
These studies collectively suggest the future utility of using DNA methylation patterns in cord blood or maternal blood as biomarkers of prenatal exposures. Future studies using more agnostic approaches (e.g., DNA methylation microarrays) may be important in developing comparatively less biased, hypothesis-generating methods capable of further exploring the often complex pathways affected by prenatal smoke exposure. In addition, such technological approaches may discover DNA methylation biomarkers with greater sensitivity and specificity for assessing maternal cigarette smoking during pregnancy.
Leukocyte DNA
Terry and colleagues (Reference Terry, Ferris, Pilsner, Flom, Tehranifar and Santella2008) analyzed DNA methylation profiles in leukocyte (white blood cell) DNA in a multiethnic birth cohort from New York City. Multivariable models indicated that overall levels of DNA methylation were significantly associated with maternal smoking during pregnancy and a number of other covariates. Terry et al. point out that their data, if replicated in a larger samples, suggest that exposures experienced throughout the course of life (i.e., from fertilization onward) may be associated with DNA methylation in adulthood. Longitudinal studies capable of measuring within-individual changes in DNA methylation in a variety of tissues over time will yield important data informative of the intragenerational plasticity of DNA methylation.
Brain and neurodevelopmental outcomes
There is great interest in better understanding the influences of maternal cigarette smoking during pregnancy on the brain and infant neurodevelopment; however, investigations considering epigenetic pathways are only beginning to emerge. Toledo-Rodriguez et al. (Reference Toledo-Rodriguez, Lotfipour, Leonard, Perron, Richer and Veillette2010) investigated whether prenatal exposure to maternal cigarette smoking is associated with promoter methylation of brain-derived neurotrophic factor (BDNF) in blood samples from adolescents whose mothers smoked during pregnancy. BDNF, which is important for long-term memory, acts on certain neurons of the central nervous system and the peripheral nervous system, helping to support the survival of existing neurons, and encourage the growth and differentiation of new neurons and synapses (Huang & Reichardt, Reference Huang and Reichardt2001). Toledo-Rodriguez et al. (Reference Toledo-Rodriguez, Lotfipour, Leonard, Perron, Richer and Veillette2010) found that exposure to maternal cigarette smoking in utero is associated with a higher degree of DNA methylation in the BDNF exon 6 in adolescents whose mothers smoked during pregnancy, suggesting that exposure to cigarette smoke while in utero may have long-term consequences still measurable into adolescence. The BDNF gene contains several exons, the expression of which is crucial for the proper functioning of BDNF protein and that have been shown to be differentially regulated in an exon-specific manner. Another preliminary study noted that measurement of methylation upstream of BDNF exon I may have promise as a diagnostic blood biomarker for major depression (Fuchikami et al., Reference Fuchikami, Morinobu, Segawa, Okamoto, Yamawaki and Ozaki2011). It is interesting that exon 6 of the BDNF gene appears to be particularly sensitive to epigenetic modification from environmental factors related to depression. Specifically, chronic stress results in epigenetic changes in histones related to hippocampal BDNF that reduce expression whereas treatment with the antidepressant imipramine results in epigenetic changes of histones associated with the same exon that increase expression rates (Calabrese, Molteni, Racagni, & Riva, Reference Calabrese, Molteni, Racagni and Riva2009). This example highlights the importance the various types of epigenetic modification that may increase (or decrease) the risk of psychopathology by modifying the same gene in different ways and how such mechanisms may overlap with prenatal smoke exposure. Abnormal methylation upstream of one BDNF exon may change the ultimate expression of BDNF and thus play a role in altering downstream pathways important for proper brain development and plasticity (Toledo-Rodriguez et al., Reference Toledo-Rodriguez, Lotfipour, Leonard, Perron, Richer and Veillette2010). Future studies using primary brain tissue may prove challenging but essential in determining the role of BDNF and related biological pathways in brain development and later behavior.
Future analyses focused on assessing epigenetic mechanisms mediating the effects of maternal cigarette smoking during pregnancy on brain, developmental, and behavioral outcomes using both animal models and human cohorts will be essential. Such studies will aid in further defining how in utero exposures may influence neurodevelopmental outcomes and risk for behavioral disorders and diseases.
Summary: DNA methylation
Although studies to date have been limited in number and scope, it is important to consider the role of DNA methylation in the pathway from maternal cigarette smoking during pregnancy to later child and adolescent outcomes. However, it is clear that we have much to learn. Specifically, a better understanding of the mechanisms involved or what these associations imply for later offspring outcomes, such as birth weight, cognitive performance, executive function, externalizing behavior, and other health-related phenotypes, including mental health, will be key. In order to accomplish this, we need to elucidate potential etiological pathways linking prenatal exposure-altered DNA methylation and later behavioral deficits or problems in childhood and adolescence. Finally, there is a need to continue to develop new techniques for assessing the utility of DNA methylation profiles in accessible tissues, such as the placenta and the umbilical cord blood, further exploring the potential for DNA methylation as a biomarker for prenatal smoke exposure and other adverse prenatal exposures.
Maternal Cigarette Smoking During Pregnancy Is Associated With Differential miRNA Expression
Compared to the growing body of literature characterizing associations between DNA methylation patterns and maternal cigarette smoking during pregnancy, much less is known about associations between maternal cigarette smoking during pregnancy and miRNA expression. miRNA are small, ~22 nucleotide-long noncoding RNA molecules, which were previously shown to be highly ubiquitous and conserved across many species (Lee, Feinbaum, & Ambros, Reference Lee, Feinbaum and Ambros1993). miRNA control gene expression by base pairing to the 3′-untranslated region of a target messenger RNA (mRNA). In some cases, there will be an imperfect match, or imperfect complementarity, between the miRNA and its mRNA target, which results in problems translating the mRNA into a protein molecule (i.e., translational repression). In other cases of perfect complementarity between the miRNA and the target mRNA, the result is degradation of the target mRNA. Research has found that because partial or imperfect complementarity of a miRNA to a target mRNA can still result in translational repression, a single miRNA has the capability of regulating a number of genes (Du & Zamore, Reference Du and Zamore2007). Through the mechanism of negative regulation, miRNA have been shown to be involved in regulating a number of key biological processes, including cell proliferation, differentiation, and apoptosis (i.e., cell death; Maccani & Knopik, Reference Maccani and Knopik2012; Maccani & Marsit, Reference Maccani and Marsit2009; Miska, Reference Miska2005).
One of the few studies investigating maternal cigarette smoking during pregnancy and miRNA expression was conducted by Maccani et al. (Reference Maccani, Avissar-Whiting, Banister, McGonnigal, Padbury and Marsit2010). They used a candidate miRNA approach to investigate the changes in four human placental miRNA associated with maternal cigarette smoking during pregnancy. These four miRNA were selected for investigation because they had been reported to be expressed in the placenta and had been shown to be involved in the regulation of key cell processes. The work identified the association of maternal cigarette smoking during pregnancy with the downregulation of three specific miRNA: miR-16, miR-21, and miR-146a. Based on these findings, the research was expanded to consider three placental cell lines from different stages of placental development. The aim was to further investigate the effects of nicotine and benzo(a)pyrene, two components of cigarette smoke previously suggested to have negative effects on both the placenta and fetus, on miRNA expression in placental cells. Downregulation of miR-146a in T cell leukemia/lymphoma 1 cell treated with nicotine and benzo(a)pyrene suggested that miR-146a may be especially sensitive to agents of cellular stress, such as prenatal smoke exposure. Moreover, this result suggests that two specific components of cigarette smoke that affect the expression of miR-146a in term placentas may be nicotine and benzo(a)pyrene (Maccani et al., Reference Maccani, Avissar-Whiting, Banister, McGonnigal, Padbury and Marsit2010).
However, Maccani et al.'s (2010) observations were limited by a relatively small set of samples (n = 25), as well as a lack of data regarding the duration of cigarette smoking during pregnancy, daily cigarette usage, or more extensive environmental exposure information (e.g., environmental pollutant exposure or second-hand/passive cigarette smoke exposure). Despite these limitations, their data comprise an important first step in determining associations between maternal cigarette smoking during pregnancy and aberrant miRNA expression in the placenta. More work is needed to investigate associations between maternal cigarette smoking during pregnancy and abnormal miRNA expression in a larger sample set with more complete environmental exposure information, as well as potentially using a hypothesis-generating approach (e.g., using miRNA microarrays; Maccani et al., Reference Maccani, Avissar-Whiting, Banister, McGonnigal, Padbury and Marsit2010; Maccani & Marsit, Reference Maccani and Marsit2011). Future work assessing associations between exposure-influenced placental miRNA profiles and downstream neurobehavioral and developmental outcomes is also needed. For instance, longitudinal analyses across the life span will be critical for elucidating the impact of miRNA-mediated fetal programming on later health, behavior, and disease.
As reviewed by Maccani and Marsit (Reference Maccani and Marsit2011), future experiments using cigarette smoke condensate (Crane-Godreau et al., Reference Crane-Godreau, Maccani, Eszterhas, Warner, Jukosky and Fiering2009) may enable researchers to better understand effects of complex mixtures of components of cigarette smoke on placental cells and cells derived from other tissues. Of course, such experiments will also present challenges: determining the specific composition of the complex mixture as well as the physiological relevance of exposure to such a mixture on a given cell type. Other more extensive experiments aimed at better understanding the more complex effects of organism-wide cigarette smoke exposure might be designed using mouse (or other animal) models of chronic environmental tobacco smoke exposure as described previously (Maccani & Marsit, Reference Maccani and Marsit2011; Xiong, Leme, Ray, Shapiro, & Lee, Reference Xiong, Leme, Ray, Shapiro and Lee2011). In such an experimental design, pregnant mice might be exposed to environmental tobacco smoke throughout their entire gestation and euthanized at key time points in gestation, with placentas and other key organs harvested for measurement of miRNA expression and for determination of the effects of dysregulated miRNA expression on target gene protein levels and function. The findings from such experiments would elaborate on the work done to characterize the association of maternal cigarette smoking during pregnancy with changes in placental miRNA and expand investigations into new tissues. In addition, such findings using an in vivo model system may provide crucial mechanistic data that may further strengthen data from previous epidemiologic association studies with findings from a model system experimental paradigm.
Integration of Epigenetic Characterization Into Existing Human Research Paradigms
There is a growing literature on maternal cigarette smoking during pregnancy and epigenetic phenomena, such as DNA methylation and miRNA expression. How do we align these recent advances with the overall literature on smoking during pregnancy, particularly the reported associations between smoking during pregnancy and later child/adolescent behavior? There is a large body of literature suggesting undesirable outcomes in children exposed to maternal smoking during pregnancy. This evidence (in human cohorts) has been limited until recently by researchers’ frequent inability to separate prenatal exposure effects from other confounding environmental and genetic factors. The majority of studies have provided limited control for prenatal exposures possibly being correlated with parental behaviors that could act as more proximal risk factors that are in turn transmitted to their offspring (Knopik, Reference Knopik2009). In other words, mothers who smoke during their pregnancies share other risk factors with their children and it may be these other risk factors that are associated with the observed adverse outcomes rather than only maternal cigarette smoking during pregnancy per se (Kuja-Halkola, D'Onofrio, Iliadou, Langstrom, & Lichtenstein, Reference Kuja-Halkola, D'Onofrio, Iliadou, Langstrom and Lichtenstein2010). These factors can include unmeasured familial environmental influences, such as second-hand smoke, socioeconomic status, and parental education. Additional factors might be genetic influences, such as genetic variation influencing (a) neurocognitive phenotypes through nonexposure contingent mechanisms, (b) pharmacokinetic and/or pharmacodynamic aspects of nicotinic effects, or (c) metabolism of the 4,000+ other xenobiotics (e.g., foreign substances) found in cigarette smoke. Epigenetic modification has long been considered a possible influence in this regard; however, this article indicates that it largely remains to be empirically tested when it comes to behavioral or neurodevelopmental phenotypes.
Because of ethical and practical concerns about research considering prenatal smoke exposure, maternal cigarette smoking during pregnancy in humans cannot be randomized. The consequence of this is that prenatal smoke exposure effects on human offspring outcomes are quasiexperimental in nature (Shadish, Cook, & Campbell, Reference Shadish, Cook and Campbell2002). However, innovative designs such as full sibling pairs discordant for prenatal exposure (an extension of the case-crossover design; D'Onofrio, Singh, Iliadou, Lambe, Hultman, Grann, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Grann2010; D'Onofrio, Singh, Iliadou, Lambe, Hultman, Neiderhiser, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Neiderhiser2010; D'Onofrio et al., Reference D'Onofrio, Van Hulle, Waldman, Rodgers, Harden and Rathouz2008; Knopik, Reference Knopik2009; Knopik, McGeary, Nugent, Francazio, & Heath, Reference Knopik, McGeary, Nugent, Francazio and Heath2010; Kuja-Halkola et al., Reference Kuja-Halkola, D'Onofrio, Iliadou, Langstrom and Lichtenstein2010; Lumley & Levy, Reference Lumley and Levy2000; Meyer, Williams, Hernandez-Diaz, & Cnattingius, Reference Meyer, Williams, Hernandez-Diaz and Cnattingius2004) and in vitro fertilization cross-fostering approaches (Rice et al., Reference Rice, Harold, Boivin, Hay, van den Bree and Thapar2009, Reference Rice, Harold, Boivin, van den Bree, Hay and Thapar2010; Thapar et al., Reference Thapar, Harold, Rice, Ge, Boivin and Hay2007, Reference Thapar, Rice, Hay, Boivin, Langley and van den Bree2009) have enabled better controls for confounding factors. Results from these studies suggest that some associations suggested between maternal cigarette smoking during pregnancy and offspring behavioral outcomes, a common finding in the wider literature, may indeed be influenced by design limitations, specifically, an inability to adequately control for shared familial influences, including genetic factors. This has been shown for offspring conduct problems (D'Onofrio et al., Reference D'Onofrio, Van Hulle, Waldman, Rodgers, Harden and Rathouz2008), criminality (D'Onofrio, Singh, Iliadou, Lambe, Hultman, Grann, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Grann2010), academic achievement (D'Onofrio, Singh, Iliadou, Lambe, Hultman, Neiderhiser, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Neiderhiser2010; Lambe et al., Reference Lambe, Hultman, Torrang, Maccabe and Cnattingius2006), intellectual performance (Lundberg et al., Reference Lundberg, Cnattingius, D'Onofrio, Altman, Lambe and Hultman2009) and ADHD (Knopik et al., Reference Knopik, McGeary, Nugent, Francazio and Heath2010; Thapar et al., Reference Thapar, Rice, Hay, Boivin, Langley and van den Bree2009). Thus, the suggestion that maternal cigarette smoking during pregnancy is exclusively responsible for a broad range of later offspring behavioral outcomes must be tempered and reconsidered in light of these recent findings across multiple research groups.
Moreover, preliminary efforts to study the fetal programming hypothesis, as described above, from a phenotypic standpoint (i.e., in the absence of epigenetic data), using low birth weight as a proxy for a poor gestational environment, suggest that this mechanism does not wholly explain potential prenatal smoke exposure influences on later psychological functioning (Kuja-Halkola et al., Reference Kuja-Halkola, D'Onofrio, Iliadou, Langstrom and Lichtenstein2010). Kuja-Halkola et al. (Reference Kuja-Halkola, D'Onofrio, Iliadou, Langstrom and Lichtenstein2010) also suggest that possible effects of low birth weight on later offspring behavior may also be due to familial confounding. Thus, further biological characterization, such as specific genetic influences, as well as epigenetic modification resulting from prepregnancy smoking, which has an influence regardless of the decision to smoke during pregnancy, may further highlight influences beyond maternal cigarette smoking during pregnancy and add clarity to this complicated story.
The conclusions of this body of quasiexperimental approaches to this research question should not be overinterpreted to suggest that maternal cigarette smoking during pregnancy is a benign influence. Substantial evidence exists to suggest prenatal cigarette smoke exposure is associated with numerous adverse outcomes related to birth and infancy (Cnattingius, Reference Cnattingius2004; D'Onofrio, Singh, Iliadou, Lambe, Hultman, Grann, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Grann2010; D'Onofrio, Singh, Iliadou, Lambe, Hultman, Neiderhiser, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Neiderhiser2010; D'Onofrio et al., Reference D'Onofrio, Turkheimer, Eaves, Corey, Berg and Solaas2003; Gilman, Gardener, & Buka, Reference Gilman, Gardener and Buka2008; Johansson, Dickman, Kramer, & Cnattingius, Reference Johansson, Dickman, Kramer and Cnattingius2009; Rice et al., Reference Rice, Harold, Boivin, Hay, van den Bree and Thapar2009; Sexton & Hebel, Reference Sexton and Hebel1984). Thus, there is a need for continued smoking cessation efforts in pregnant mothers. However, given that smoking during pregnancy is correlated with a host of other behaviors (Agrawal et al., Reference Agrawal, Knopik, Pergadia, Waldron, Bucholz and Martin2008; D'Onofrio, Singh, Iliadou, Lambe, Hultman, Neiderhiser, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Grann2010; Knopik, Reference Knopik2009; Knopik et al., Reference Knopik, Sparrow, Madden, Bucholz, Hudziak and Reich2005, Reference Knopik, Heath, Jacob, Slutske, Bucholz and Madden2006), treatment and prevention efforts might need to also consider other factors, such as maternal alcohol dependence, maternal nicotine dependence, and maternal ADHD.
Given the above considerations, the potential influence of maternal cigarette smoking during pregnancy on epigenetic mechanisms, which may have consequences for downstream behavior, is considerable. First, there is the possibility of intergenerational transmission of smoking-related epigenetic consequences (i.e., grandmaternal smoking influences may affect germline cells or may escape epigenetic reprogramming during development). Second, we can begin to understand the extent to which epigenetic modification may alter critical neurobehavioral circuitry in the developing brain. Third, questions regarding the influence of smoke exposure induced methylation on the xenobiotic metabolism of cigarette smoke by-products can begin to be answered. These are just a few of the currently unanswered questions for which epigenetic research might begin to provide answers. Fortunately, at least for DNA methylation, existing data sets with collected DNA samples may provide some opportunities to investigate such possibilities. For example, methylomic characterization of samples collected in sibling pairs discordant for prenatal exposure might provide initial evidence of longer lasting affects of prepregnancy smoking, which may make maternal smoking during pregnancy per se less important than the overall maternal smoking history. Use of animal models in mutually informative translational research (especially in inbred lines where the genetic background is held constant) may further strengthen our ability as a field to investigate these tantalizing hypotheses.
Preliminary evidence already points to important genetic differences in xenobiotic metabolism with downstream effects on intrauterine growth (Price, Grosser, Plomin, & Jaffee, Reference Price, Grosser, Plomin and Jaffee2010) and behavior (Knopik, McGeary, Nugent, Francazio, & Heath, in press). Further characterization of these samples from an epigenetic standpoint could explain additional variance in behavioral outcomes while simultaneously identifying potentially targetable mechanisms to reduce such effects.
Conclusion and Future Directions
Important advances in understanding DNA methylation patterns and altered miRNA expression associated with maternal cigarette smoking during pregnancy have laid the groundwork for exciting research in this domain. Research is ongoing to understand more completely the reported associations reviewed in this article and the mechanisms underlying the effects of maternal cigarette smoking during pregnancy. It will also be important to expand the current literature pertaining to the epigenetic mechanisms mediating the effects of maternal cigarette smoking during pregnancy on neuropsychiatric and neurobehavioral outcomes across development. In addition, more work needs to be done to explore the role of maternal cigarette smoking during pregnancy on histone modification and imprinting, two crucial epigenetic mechanisms that play key roles in development and health and that have been vastly understudied in the context of maternal cigarette smoking during pregnancy.
One area of research yet to be fully explored is determining which epigenetic mechanisms underlie psychological disorders associated with maternal cigarette smoking during pregnancy. This could perhaps be done using mechanisms described in the theory of behavioral teratogenicity, such as fetal brain exposure to nicotine and the pathological activation of acetylcholine nicotinic receptors during early stages of brain development (Paz, Barsness, Martenson, Tanner, & Allan, Reference Paz, Barsness, Martenson, Tanner and Allan2007). Given that there is a high degree of comorbidity between nicotine dependence and neuropsychiatric conditions (Maughan, Taylor, Caspi, & Moffitt, Reference Maughan, Taylor, Caspi and Moffitt2004), researchers have struggled to determine how influential exposure to cigarette smoke is on the risk of developing psychological disorders.
A variety of previous studies suggest that prenatal nicotine exposure causes an increased risk for developing ADHD, major depressive disorder, and substance abuse in adolescents and children (Fergusson, Woodward, & Horwood, Reference Fergusson, Woodward and Horwood1998; Linnet et al., Reference Linnet, Wisborg, Obel, Secher, Thomsen and Agerbo2005; Maughan et al., Reference Maughan, Taylor, Caspi and Moffitt2004; Mick, Biederman, Faraone, Sayer, & Kleinman, Reference Mick, Biederman, Faraone, Sayer and Kleinman2002; Milberger, Biederman, Faraone, Chen, & Jones, Reference Milberger, Biederman, Faraone, Chen and Jones1996; Paz et al., Reference Paz, Barsness, Martenson, Tanner and Allan2007; Thapar et al., Reference Thapar, Fowler, Rice, Scourfield, van den Bree and Thomas2003; Weissman, Warner, Wickramaratne, & Kandel, Reference Weissman, Warner, Wickramaratne and Kandel1999). However, others suggest that these associations are due less to prenatal nicotine and more to shared familial influences, including genetics (D'Onofrio, Singh, Iliadou, Lambe, Hultman, Grann, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Grann2010; D'Onofrio, Singh, Iliadou, Lambe, Hultman, Neiderhiser, et al., Reference D'Onofrio, Singh, Iliadou, Lambe, Hultman and Neiderhiser2010; D'Onofrio et al., Reference D'Onofrio, Van Hulle, Waldman, Rodgers, Harden and Rathouz2008; Knopik, Reference Knopik2009; Knopik et al., Reference Knopik, McGeary, Nugent, Francazio and Heath2010; Kuja-Halkola et al., Reference Kuja-Halkola, D'Onofrio, Iliadou, Langstrom and Lichtenstein2010; Lumley & Levy, Reference Lumley and Levy2000; Rice et al., Reference Rice, Harold, Boivin, Hay, van den Bree and Thapar2009, Reference Rice, Harold, Boivin, van den Bree, Hay and Thapar2010; Thapar et al., Reference Thapar, Harold, Rice, Ge, Boivin and Hay2007, Reference Thapar, Rice, Hay, Boivin, Langley and van den Bree2009). Paz et al. (Reference Paz, Barsness, Martenson, Tanner and Allan2007) used a self-administered maternal nicotine consumption model to assess how prenatal nicotine exposure might influence a variety of neuropsychiatric disorders in mice. They found that prenatal nicotine exposure had a significant effect on increasing latency to escape in a learned helplessness paradigm, the degree of spontaneous locomotion, and an increase in addictive behavior as assessed by a preference for a place previously associated with cocaine. Paz et al.'s findings suggest that prenatal nicotine exposure may significantly influence the increased prevalence of major depressive disorder, ADHD, and substance abuse found in children and adolescents whose mothers smoked cigarettes during pregnancy, a finding that underscores the behavioral teratogenic effects of maternal cigarette smoking during pregnancy. Moreover, they speculate that this may be due to a direct interaction between nicotine and acetylcholine nicotine receptors in the fetal brain. Variations in the genes coding for such receptors may affect the entire biological pathway from prenatal exposure to later development.
Important work still needs to be done to determine the mechanisms affected by exposures that may lead to neurodevelopmental and neurobehavioral outcomes. DNA methylation patterns in both candidate gene promoters as well as on genome-wide levels should be assessed using both animal models of maternal cigarette smoking during pregnancy and human cohorts. Additional studies that may be of special importance might include an investigation of associations of maternal cigarette smoking during pregnancy on DNA methylation of the promoter of the glucocorticoid receptor (GR). This receptor is known to be a mediator of glucocorticoid signaling (a major component of the stress-response system, which is closely linked to depression and anxiety psychopathology), and it was previously shown to be partly regulated by DNA methylation (Filiberto et al., Reference Filiberto, Maccani, Koestler, Wilhelm-Benartzi, Avissar-Whiting and Banister2011; Herbeck, Gulevich, Amelkina, Plyusnina, & Oskina, Reference Herbeck, Gulevich, Amelkina, Plyusnina and Oskina2010; Ke et al., Reference Ke, Schober, McKnight, O'Grady, Caprau and Yu2010; Oberlander et al., Reference Oberlander, Weinberg, Papsdorf, Grunau, Misri and Devlin2008; Weaver et al., Reference Weaver, Cervoni, Champagne, D'Alessio, Sharma and Seckl2004, Reference Weaver, La Plante, Weaver, Parent, Sharma and Diorio2001). Although previous studies have controlled for maternal cigarette smoking during pregnancy and did not find significant associations between maternal smoking and aberrant methylation at the 13 CpG sites in exon 1F of the GR gene (Filiberto et al., Reference Filiberto, Maccani, Koestler, Wilhelm-Benartzi, Avissar-Whiting and Banister2011), a more comprehensive analysis of the promoter region controlling expression of GR may reveal CpG loci more responsive to and influenced by maternal cigarette smoking during pregnancy. Future work should be conducted using animal models and human cohorts and should utilize a variety of tissues for analysis, including placenta, specific brain regions, saliva, and blood. A more comprehensive investigation of altered DNA methylation of the GR gene promoter associated with maternal cigarette smoking during pregnancy might better describe effects of environmental toxicant exposure during pregnancy on mediators of the hypothalamus–pituitary–adrenal axis, which may contribute to an increased risk for developing psychological disorders in childhood or later in life. Investigation of the epigenetic mechanisms that may underlie a suite of psychological disorders, including but not limited to posttraumatic stress disorder, depression, schizophrenia, and ADHD, may provide researchers with a greater understanding of the potential epigenetic nature of these disorders while also providing new therapeutic and pharmacokinetic targets for treatment. Both candidate and genome-wide association studies should be employed to investigate how maternal cigarette smoking during pregnancy might influence risk for neuropsychiatric disease through epigenetic mechanisms.
Even though brain researchers have limited access to the tissue of interest compared to other areas of medicine, progress in the areas of brain-based neurological illness provides evidence of the importance of this research. The literature related to the epigenetics of Alzheimer disease and Parkinson disease, more specifically miRNA, which may be involved in regulating genes described to be critical for disease progression, has grown substantially over the past decade. Meza-Sosa, Valle-Garcia, Pedraza-Alva, and Perez-Martinez (Reference Meza-Sosa, Valle-Garcia, Pedraza-Alva and Perez-Martinez2011) have extensively reviewed recent findings demonstrating that miRNA may play important roles in the development of the central nervous system as well as in a variety of neuropathologies, including Alzheimer and Parkinson diseases. Future studies in both human cohorts and animal models investigating epigenetic mechanisms altered by maternal cigarette smoking during pregnancy should also consider exploring exposure-associated epigenetic links to Alzheimer and Parkinson diseases. Such links may both deepen our understanding of these complex etiologies and may reveal new therapeutic targets for future treatment options.
In summary, the utilization of a variety of approaches will allow researchers to more effectively investigate changes in DNA methylation, miRNA expression, histone modification, and imprinting, which may be associated with in utero exposures, behavior, and even disease development. Profiles of epigenetic regulation in a variety of tissues associated with maternal cigarette smoking during pregnancy will inform researchers of the impact of maternal cigarette smoking during pregnancy on the epigenetic mechanisms in a particular tissue and may also prove to be powerful biomarkers of exposure, disease susceptibility or progression, and even pharmacokinetic efficacy. In such ways, translational epigenetics may be brought from benchtop-to-bedside and may prove advantageous in aiding clinicians in the diagnosis of exposure, disease, and risk for mental health and behavioral problems, as well as treatment.
Finally, a consistent body of research stresses the need for a more careful examination of maternal cigarette smoking during pregnancy within the context of genetic background. Although prenatal smoke exposure-induced epigenetic modification has been consistently found to affect physical health phenotypes, the examination of epigenetic modification as a moderator of later neurobehavioral effects is in its infancy. The same biological systems investigated in studies of genetic variation (e.g., nicotinic and dopaminergic systems, xenobiotic metabolism pathways) remain viable candidates for epigenetic influences on neurobehavioral outcomes in exposed offspring. The comodeling of genetic and epigenetic variation as they relate to these outcomes has the potential to bring new clarity to the field. Although epigenetic patterns change over time and with environmental exposures (and, therefore, are not as static as genetic variation), DNA samples in existing data sets may be used to provide an informative cross-sectional view of this phenomenon, provided that careful attention is given to the time of sample collection. Promising findings can be followed up with longitudinal studies with sequential sampling designed to assess the relative impact of environmental exposures. For example, in the context of prenatal smoke exposure research, sampling at the beginning and the end of pregnancy in smokers who did and did not decide to quit smoking while pregnant would allow for an examination of the extant methylation resulting from prepregnancy smoking. This would also allow for investigation of whether or not smoking cessation during pregnancy has an impact on epigenetic mechanisms.
In sum, we are on the verge of a new and more complete understanding of the biological impact of maternal cigarette smoking during pregnancy that, when coupled with advances in the field of prenatal exposure research, holds great promise of increasing our understanding of the consequences of such behavior for offspring. Such measures will be essential in understanding and addressing the effects of maternal cigarette smoking during pregnancy on fetal programming and child development.

