


Perinatal and early life experiences can shape behavior and physiology. The development of some traits may be triggered in pregnancy or early life and persist into adulthood (Morgan & Bale, Reference Morgan and Bale2011; Weaver et al., Reference Weaver, Cervoni, Champagne, D'Alessio, Sharma, Seckl and Meaney2004). Clinical studies have shown that maternal prenatal stress factors are significantly associated with poor pregnancy outcomes, such as low infant birth weight and decreased gestational age at birth (Wadhwa, Sandman, Porto, Dunkel-Schetter, & Garite, Reference Wadhwa, Sandman, Porto, Dunkel-Schetter and Garite1993). In particular, psychological distress during pregnancy (i.e., due to low socioeconomic status, or as a result of psychiatric disorders such as depression) is known to be related to increased obstetric complications with adverse birth outcomes and impaired infant mental and physical health and development (Alder, Fink, Bitzer, Hösli, & Holzgreve, Reference Alder, Fink, Bitzer, Hösli and Holzgreve2007; Gavin, Hille, Hawkins, & Maas, Reference Gavin, Hill, Hawkins and Maas2011; Lawlor, Relton, Sattar, & Nelson, Reference Lawlor, Relton, Sattar and Nelson2012).
As the potential first point of interaction between genes and the environment, pregnancy is considered a critical window for epigenetic modifications or epimutations that are regarded as biological substrates for the programming and inheritance of experience-induced behavior (Babenko, Kovalchuk, & Metz, Reference Babenko, Kovalchuk and Metz2015; Bohacek & Mansuy, Reference Bohacek and Mansuy2015).
In addition, while it has been reported that maternal early life stress (ELS) has effects on the offspring's psychopathology in childhood (Plant, Jones, Pariante, & Pawlby, in press) the effects of maternal ELS or trauma on pregnancy outcomes and pathophysiological links between maternal trauma and child outcomes are less clear. It has been proposed that early childhood abuse in the mother is related to the offspring's birth weight, but the biological mechanisms have not yet been clarified (Gavin et al., Reference Gavin, Hill, Hawkins and Maas2011). Thus, it is important to investigate how preconceptional parental stress (Yuan et al., Reference Yuan, Li, Sun, Ouyang, Campos, Rocha and So2016), as well as postconceptional maternal stress in pregnancy may affect the child's outcome.
Consistent with these findings, in rodents, reports show that embryos exposed to chronic maternal stress in utero show depressive-like behavior in adulthood which remains after cross-fostering (Del Cerro et al., Reference Del Cerro, Pérez-Laso, Ortega, Martín, Gómez, Pérez-Izquierdo and Segovia2010). Moreover, stress during adolescence has been shown to increase anxiety across two generations (Saavedra-Rodriguez & Feig. Reference Saavedra-Rodriguez and Feig2013). Thus, rodent research also suggests that prenatal experiences shape the behavior of the offspring and that stress-induced behavior may be transmitted over several generations.
It has been shown that the activation of the maternal hypothalamus–pituitary–adrenal (HPA) system, cortisol passing the placenta (Baibazarova et al., Reference Baibazarova, van de Beek, Cohen-Kettenis, Buitelaar, Shelton and van Goozen2013), and glucocorticoid exposure of the fetus may lead to low birth weight and its subsequent disease liability (Cottrell, Seckl, Holmes, & Wyrwoll, Reference Cottrell, Seckl, Holmes and Wyrwoll2014). While, generally, cortisol in amniotic fluid is closely related to maternal HPA system activity (Sarkar, Bergman, O'Connor, & Glover, Reference Sarkar, Bergman, O'Connor and Glover2008), the placental enzyme 11ß-hydroxysteroid dehydrogenase-2 (HSD2) may function as an enzymatic buffer against the deleterious effects of exposure to maternal glucocorticoids. However, this enzyme can be downregulated by adverse prenatal experiences, which is thought to lower the capacity to protect the developing fetus (O'Donnell et al., Reference O'Donnell, Jensen, Freeman, Khalife, O'Connor and Glover2012). In rats, chronic stress during gestation may lead to lowered HSD2 mRNA, both in the placenta and the brain, probably by epigenetic mechanisms (Peña, Monk, & Champagne, Reference Peña, Monk and Champagne2012). Specifically, placental HSD2 downregulation has been related with maternal anxiety (O'Donnell et al., Reference O'Donnell, Jensen, Freeman, Khalife, O'Connor and Glover2012) rather than depression (Conradt et al., Reference Conradt, Fei, Lagasse, Tronick, Guerin, Gorman and Lester2015). Maternal depression is related with increased mRNA levels of placental glucocorticoid receptors (GR; Reynolds et al., Reference Reynolds, Pesonen, O'Reilly, Tuovinen, Lahti, Kajantie and Räikkönen2015) implying altered placental GR sensitivity in depressed mothers. In contrast to HSD2, placental HSD1 regenerates cortisol from cortisone and may increase fetus's glucocorticoid exposure. Rodent and human (Monk et al., Reference Monk, Feng, Lee, Krupska, Champagne and Tycko2016) studies have shown that the expression of HSD1 and HSD2 is differentially regulated by maternal stress (Seckl & Holmes, Reference Seckl and Holmes2007). Recently, Reynolds's group reported maternal stress-induced reduction of placental HSD2 mRNA levels in a sex-specific manner in female, but not male offspring (Mina, Räikkönen, Riley, Norman, & Reynolds, Reference Mina, Räikkönen, Riley, Norman and Reynolds2015). In contrast to HSD2, maternal stress induced an increase of HSD1 in obese women (Mina et al., Reference Mina, Räikkönen, Riley, Norman and Reynolds2015). Thus, maternal stress may lead to an increase of GR-mediated placental effects and high fetal cortisol exposure, both by lowered HSD2 and increased HSD1 in a sex- and weight-specific manner.
The human brain undergoes complex morphological and functional changes during intrauterine development. This is especially true in the first and second trimesters of pregnancy, when environmental challenges may interfere with the processes of neuronal proliferation, migration, and organization that peak in this time period (Fumagalli, Molteni, Racagni, & Riva, Reference Fumagalli, Molteni, Racagni and Riva2007). Neurotrophins play an important role in the brain's development, and it has been known for decades that brain-derived neurotrophic factor (BDNF) is essential for the development and maturation of the brain (Maisonpierre et al., Reference Maisonpierre, Belluscio, Friedman, Alderson, Wiegard, Furth and Yancopoulos1990).
BDNF is essential for the outgrowth and activity-dependent remodeling of axonal arbors in vivo (Jeanneteau, Deinhardt, Miyoshi, Bennett, & Chao, Reference Jeanneteau, Deinhardt, Miyoshi, Bennett and Chao2010). Specifically, it has been shown that BDNF is involved in embryonal hippocampal neurogenesis (Glenn et al., Reference Glenn, Gibson, Kirby, Mellott, Blusztajn and Williams2007) and plays a role in the embryonal neurodevelopment of dopaminergic (Nakakura, Suzuki, Watanabe, & Tanaka, Reference Nakakura, Suzuki, Watanabe and Tanaka2007), noradrenergic (Guo, Hellard, Huang, & Katz, Reference Guo, Hellard, Huang and Katz2005), and serotonergic neurons (Rumajogee et al., Reference Rumajogee, Vergé, Hanoun, Brisorgueil, Hen, Lesch and Miquel2004).
Prenatal and early life adversity may be related to lasting effects on the methylation (Kertes et al., Reference Kertes, Bhatt, Kamin, Hughes, Rodney and Mulligan2017) and regulation of the BDNF gene (Buchmann et al., Reference Buchmann, Hellweg, Rietschel, Treutlein, Witt, Zimmermann and Deuschle2013), and in rats it has been shown that in pregnancy, environmental challenges may lead to differential BDNF gene methylation both in blood and in the brain (Kundakovic et al., Reference Kundakovic, Gudsnuk, Herbstman, Tang, Perera, Champagne and Pfaff2015). The effect of prenatal maternal stress on BDNF expression is region, stressor, and time specific (Braithwaite, Kundakovic, Ramchandani, Murphy, & Champagne, Reference Braithwaite, Kundakovic, Ramchandani, Murphy and Champagne2015; Gilmore, Jarskog, & Vadlamudi, Reference Gilmore, Jarskog and Vadlamudi2003; Wang & Ye, Reference Wang and Ye2008). Increased BDNF DNA methylation and respectively decreased BDNF expression has been found in the offspring of prenatally stressed rats (in the amygdala and hippocampus; Boersma et al., Reference Boersma, Lee, Cordner, Ewald, Purcell, Moghadam and Tamashiro2013) or depressed mothers during pregnancy (in buccal cells; Braithwaite et al., Reference Braithwaite, Kundakovic, Ramchandani, Murphy and Champagne2015). In contrast, increased BDNF has been found in the fetal cortex in pregnant rats exposed to infection (Gilmore, Jarskog, & Vadlamudi, Reference Gilmore, Jarskog and Vadlamudi2003). In addition, increased BDNF expression has been reported in the fetal spinal cord of prenatally stressed rats (Winston, Li, & Sarna, Reference Winston, Li and Sarna2014). Moreover, the effect of prenatal stress on BDNF expression has mostly been studied at birth or adulthood of the offspring, while there is little information on stress-induced BDNF regulation during pregnancy.
Glucocorticoids have repeatedly been shown to lower BDNF synthesis, which is also true in pregnancy (Yu, Lee, Lee, & Son, Reference Yu, Lee, Lee and Son2004). Therefore, prenatal maternal stress-induced HPA activation and fetal glucocorticoid exposure is a candidate mechanism to modulate BDNF synthesis in pregnancy. Thus, prenatal maternal stress may lead to epigenetic modulation of fetal BDNF regulation, and maternal HPA system activation may play a role in this association between maternal qualities and fetal BDNF.
Because free diffusion occurs bidirectionally between the amniotic fluid and the fetus across fetal skin, placenta, and umbilical cord from 10 to 20 weeks of gestation, amniotic fluid composition becomes similar to that of fetal plasma during this period (Cho, Shan, Winsor, & Diamandis, Reference Cho, Shan, Winsor and Diamandis2007), and it is hypothesized that the concentrations of neurotrophins in amniotic fluid reflect fetal neurotrophin levels. As the blood–brain barrier is not fully matured at that gestational age, the passage of many proteins between the brain and the circulation might be possible (Engelhardt Reference Engelhardt2003). In rats, it has been shown that during development, blood BDNF levels and brain BDNF expression are correlated (Karege, Schwald, & Cisse, Reference Karege, Schwald and Cisse2002). Thus, potential environmental-induced alterations of the fetus's BDNF regulation in the brain could be reflected in fetal plasma and amniotic fluid. Based on the idea that BDNF in amniotic fluid reflects fetal BDNF, it has previously been shown that the fetal BDNF Val66Met genotype relates to BDNF concentrations in amniotic fluid (Cattaneo et al., Reference Cattaneo, Bocchio-Chiavetto, Zanardini, Marchina, Bellotti, Milanesi and Gennarelli2010).
Given this background, we were interested in the association of maternal stress with both the glucocorticoid concentrations in amniotic fluid, due to its association with pregnancy outcome, and BDNF, due to its major role in fetal neurodevelopment. We hypothesized that ELS (measured with the Childhood Trauma Questionnaire; CTQ) and current stress of the mother-to-be, as well as ELS, are positively related to glucocorticoid concentrations in amniotic fluid. In addition, we hypothesized that high current stress and ELS are related with BDNF concentrations in amniotic fluid. We defined current stress as a psychiatric disorder (either major depression or anxiety disorder), state anxiety as measured by Spielberger State–Trait Anxiety Inventory S-anxiety scale (STAI-S), and subjective perceived stress or impaired social security by proxy of net family income as these variables have been shown before to be related to untoward pregnancy outcomes (Dowd, Reference Dowd2007; Staneva, Bogossian, Pritchard, & Wittkowski, Reference Staneva, Bogossian, Pritchard and Wittkowski2015).
Method
Participants
We studied women undergoing their first amniocentesis for karyotyping the early second trimester of pregnancy in the outpatient department of a community clinic or a private practice (2013–2015). All parents were of European origin. Only samples from singleton pregnancies with a structurally normal fetus and appropriate fetal growth were included. Exclusion criteria were fetal structural abnormalities and amniotic fluid samples contaminated with blood. Samples with aneuploidy were excluded. None of the subjects had an underlying medical disorder or was on any significant medication (e.g., steroid treatment). No mother-to-be was on any medication, except thyroid hormones and vitamin supplementation. In addition, no subject had to be excluded due to a medical disorder that would have interfered with the dependent variables. The obstetricians asked a total of 125 women to participate. Of these, 26 mothers-to-be declined contact with the study team, 4 women had a twin pregnancy, in 6 subjects karyotyping showed a significant finding, and in 2 cases not enough biomaterial was available. Thus, we included 87 women in the BDNF analysis. After additional exclusion of 8 subjects (incomplete psychological data [n = 7]; extreme outlier of BDNF [>5 SD; n = 1]), we analyzed data of a total of 79 subjects with a gestational age of 15.9±0.86 (range: 14.3–18.9) weeks (see description of study participants in Table 1).
Table 1. Demographic, psychological, and endocrine data

Note: BMI, body mass index. EDPS, Edinburgh Prenatal Depression Scale. STAI-S, State–Trait Anxiety Inventory S-anxiety scale. STAI-T, State–Trait Anxiety Inventory T-anxiety scale.
All patients received written study information in the hour immediately after amniocentesis. If they were interested in the study, the women contacted the study team. Additional oral explanation was then given and the participants gave written consent. The study was approved by the ethics committee of the Medical Faculty Mannheim. The study included an interview and questionnaires in the week after amniocentesis. All interview and questionnaire information was assessed before the women received any results of their amniocentesis.
Assessments of stress, psychiatric symptoms, and disorders
We used a Mini-International Neuropsychiatric Interview (M.I.N.I.), as well as a clinical interview to assess current and lifetime psychiatric diagnoses. The presence of a current depressive state was defined as major depression (unipolar disorder, n = 8), while anxiety disorder was defined as diagnoses of panic disorder or generalized anxiety disorder (n = 9). With regard to comorbidities, one patient with a current anxiety disorder had a lifetime diagnosis of major depressive disorder and two patients with current depression had a lifetime diagnosis of an anxiety disorder. Depression severity was assessed using the Edinburgh Postnatal Depression Scale (EDPS; self-reported). For the assessment of anxiety, we used the STAI-S (self-reported). We used the Perceived Stress Scale (PSS; self-reported) as a measure for the subjective experience of stress. A Likert scale (0 = no stress; 10 = max. stress) was used to assess the perceived stress of amniocentesis (self-reported). The CTQ (28-items; self-reported) was used to assess early life adversities of the pregnant participants. The CTQ, 28-item version, is a standard self-rating instrument asking about aversive experiences in childhood and adolescence. The 28-question assessment covers multiple types of maltreatment. Subscales with 5 items (plus 3 validity items) allow differentiation between emotional abuse, physical abuse, sexual abuse, physical neglect, and emotional neglect (Bernstein et al., Reference Bernstein, Stein, Newcomb, Walker, Pogge, Ahluvalia and Zule2003). We used the monthly family net income as a proxy for socioeconomic stress. We grouped the family income in 21 categories (1: <150 € to 21: >10.000 €; self-reported). All ratings and interviews were done by the same interviewer (F.H.), who is a doctoral candidate and received M.I.N.I. training.
Physical health
We asked for the parents’ smoking habits before and during pregnancy (smoked in early pregnancy, n = 17), alcohol consumption, and body mass index (BMI). In addition, we asked for relevant physical disorders of the participants and used the information of the expectant mother's record of prenatal and natal care for BMI and other data. Nineteen subjects suffered from goiter, 4 subjects had hypertension, 1 subject had a congenital heart disease, and 1 subject suffered from a coagulation disorder. Four subjects developed gestational diabetes. None of the mothers had a known physical disorder that would possibly affect HPA regulation at the time of amniocentesis.
Biosamples
Amniocentesis was performed for maternal age-related risks (n = 61), findings in first trimester screening (n = 5), or ultrasound (n = 3) to exclude cytogenetic alterations. In 10 subjects, amniocentesis was done without indication of a specific maternal risk, mainly following the mothers’ concern and wish for amniocentesis. Two obstetricians provided amniotic fluid to the laboratory, and the time of the transport at room temperature was significantly different (obstetrician 1: 20 samples, 19–23 hr; obstetrician 2: 59 samples, 30–75 min transport). However, BDNF (145 ± 80 vs. 172 ± 117 pg/ml), cortisol (5.74 ± 5.46 vs. 4.68 ± 1.41 ng/ml), and cortisone concentrations in amniotic fluid (11.01 ± 3.05 vs. 11.49 ± 2.99 ng/ml) did not differ between patients from these two obstetricians. In addition, there was no significant difference in the independent variables between patients of these obstetricians. After completion of regular cytogenetic analysis, residual amniotic fluid was frozen at –20° C and subsequently stored at –80 °C within 24 hr. BDNF (Buchmann et al., Reference Buchmann, Hellweg, Rietschel, Treutlein, Witt, Zimmermann and Deuschle2013), free cortisol (F), and free cortisone (E) in amniotic fluid were analyzed as described earlier (Sánchez-Guijo, Hartmann, Shi, Remer, & Wudy, Reference Sánchez-Guijo, Hartmann, Shi, Remer and Wudy2014). However, due to the low concentration of endogenous BDNF in amniotic fluid, all samples were diluted only with the same amount of “block & sample buffer” for BDNF-ELISA.
Statistical analysis
In order to avoid multiple testing and to control for the interrelation of the independent variables, we used multivariate analysis of variance (MANOVA) to analyze the effect of (a) current psychiatric disorder (major depression or anxiety disorder), (b) socioeconomic status (proxy: family net income), (c) state anxiety (STAI-S), (d) perceived stress (PSS), (e) ELS (proxy: CTQ), and (f) amniocentesis-induced stress on BDNF and cortisol (F), cortisone (E), total glucocorticoid (E + F) concentration in amniotic fluid as well as HSD2 activity (E / [E + F]). CTQ, EDPS, STAI-S, PSS, and amniocentesis-induced stress ratings were all included as continous variables. As dependent variables, we considered BDNF, cortisol, and cortisone concentrations. In addition, we used the sum of cortisol and cortisone (E + F) as a proxy of glucocorticoid concentration and the ratio of cortisone and cortisol plus cortisone (E / [E + F]) as a proxy of placental HSD2 activity (Ghaemmaghami, Dainese, La Marca, & Zimmermann, Reference Ghaemmaghami, Dainese, La Marca, Zimmermann and Ehlert2014). As the enzymatic function of HSD2 consists in the conversion of cortisol to cortisone, the ratio between the two compounds reflects its functioning. We controlled for the fetuses’ sex, maternal age, gestational age at the time of amniocentesis, obstetrician 1 versus 2, and prepregnancy BMI. The MANOVA model was fully controlled and considered all potential interaction effects. We repeated the MANOVA with high versus low CTQ values (median split: high: >36; low: ≤36) to test the interaction between ELS (CTQ) and current life stress (major depression or anxiety disorder). CTQ total and subscores were not normally distributed. Therefore, we used natural log (ln)-transformed CTQ values in all analyses. Next to the MANOVA results, we report only the fully controlled an of analysis of variance (ANOVA) findings that were significant in the MANOVA. In addition, we analyzed the intercorrelation of the independent variables. Group differences (subjects with vs. without psychiatric disorder) were analyzed by analyses of covariance or Student t tests as appropriate. For the analyses of associations, we used Pearson correlation.
As this is one of the first reports on BDNF in human amniotic fluid, we regard our approach as hypotheses generating and therefore did not control for multiple testing. Data were analyzed using SPSS Statistics. Values are presented as mean ± SD. P values <.05 were considered to be significant.
Results
Interrelation of independent variables
There were significant associations between the psychological independent variables (see Table 2), while family income was not related to any of the psychological variables. In particular, the ln-transformed CTQ values (raw data: 41.5 ± 13.1 vs. 35.0 ± 12.9, p = .036), perceived stress (PSS; 25.7 ± 7.0 vs. 16.6 ± 7.1; p = .001), and anxiety (STAI-S; 44.8 ± 8.7 vs. 34.8 ± 8.9; p = .001) values were higher in the 17 participants with versus the 62 subjects without major depression or anxiety disorder. Anxiety (STAI-S) was significantly correlated with perceived stress (PSS, r = .79) and CTQ values (r = .29; see Table 2).
Table 2. Pearson correlation of CTQ, PSS, STAI-S, and EDPS

Note: CTQ, Childhood Trauma Questionnaire. PSS, Perceived Stress Scale. STAI-S, State–Trait Anxiety Inventory, S-anxiety scale. EDPS, Edinburgh Postnatal Depression Scale.
Effect of stress on HSD2 activity, cortisol, cortisone, cortisol + cortisone, and BDNF concentration in amniotic fluid
We found significant associations of CTQ, Wilks λ = 0.815, F (4, 60) = 3.41, p = .014, partial η2 = 0.185, power = 0.82, and family net income, Wilks λ = 0.840, F (4, 60) = 2.85, p = .031; partial η2 = 0.160, power = 0.74, on the dependent variables. There was no association of major depression or anxiety disorder, F (4, 60) = 1.67, perceived stress, F (4, 60) = 0.54, or state anxiety, F (4, 60) = 0.31, but the power for these variables was less than 0.70. There was no interaction effect of Psychiatric Disorder (major depression or anxiety disorder) × CTQ (high vs. low), Wilks λ = 0.915; F (4, 57) = 1.27, ns.
Specifically, fully controlled ANOVA showed CTQ to be positively related to BDNF concentration in amniotic fluid, F (1, 63) = 10.9, p < .002; partial η2 = 0.15, but not cortisol, F (1, 63) = 0.46, cortisone, F (1, 63) = 1.38, or glucocorticoid (F + E) concentration, F (1, 63) = 2.16, ns, and HSD2 activity, F (1, 63) = 0.002. In addition, we found that net income was associated with HSD2 activity, F (1, 63) = 4.00, p = .05; partial η2 = 0.060, cortisol, F (1, 63) = 5.83, p = .019; partial η2 = 0.085, and total glucocorticoid concentration in amniotic fluid, F (1, 63) = 6.24, p = .015; partial η2 = 0.090. There was a trend association between psychiatric disorders (major depression or anxiety disorders) with increased HSD2 activity, F (1, 63) = 3.19, p = .79; partial η2 = 0.048, but not with BDNF concentrations, cortisol, cortisone, or glucocorticoid (E + F) concentrations; all F (1, 63) < 2.1.
ELS (CTQ) and BDNF concentration in amniotic fluid
Total CTQ score was positively correlated with BDNF concentrations (r = .435, p = .0001; see Figure 2). The ln-transformed values of all CTQ subscales correlated positively with BDNF concentrations in amniotic fluid (emotional neglect: r = .292, p = .011; sexual abuse: r = .335, p = .003; physical abuse: r = .356, p = .002; emotional abuse: r = .424, p = .001), and physical neglect (r = .287, p = .012). Although 21 of our participants had relatively high CTQ total scores (≥40; > mean + 1 SD of normative data; (Scher, Stein, Asmundson, McCreary, & Forde, Reference Scher, Stein, Asmundson, McCreary and Forde2001), none of our subjects suffered from posttraumatic stress disorder as assessed by the M.I.N.I. interview.
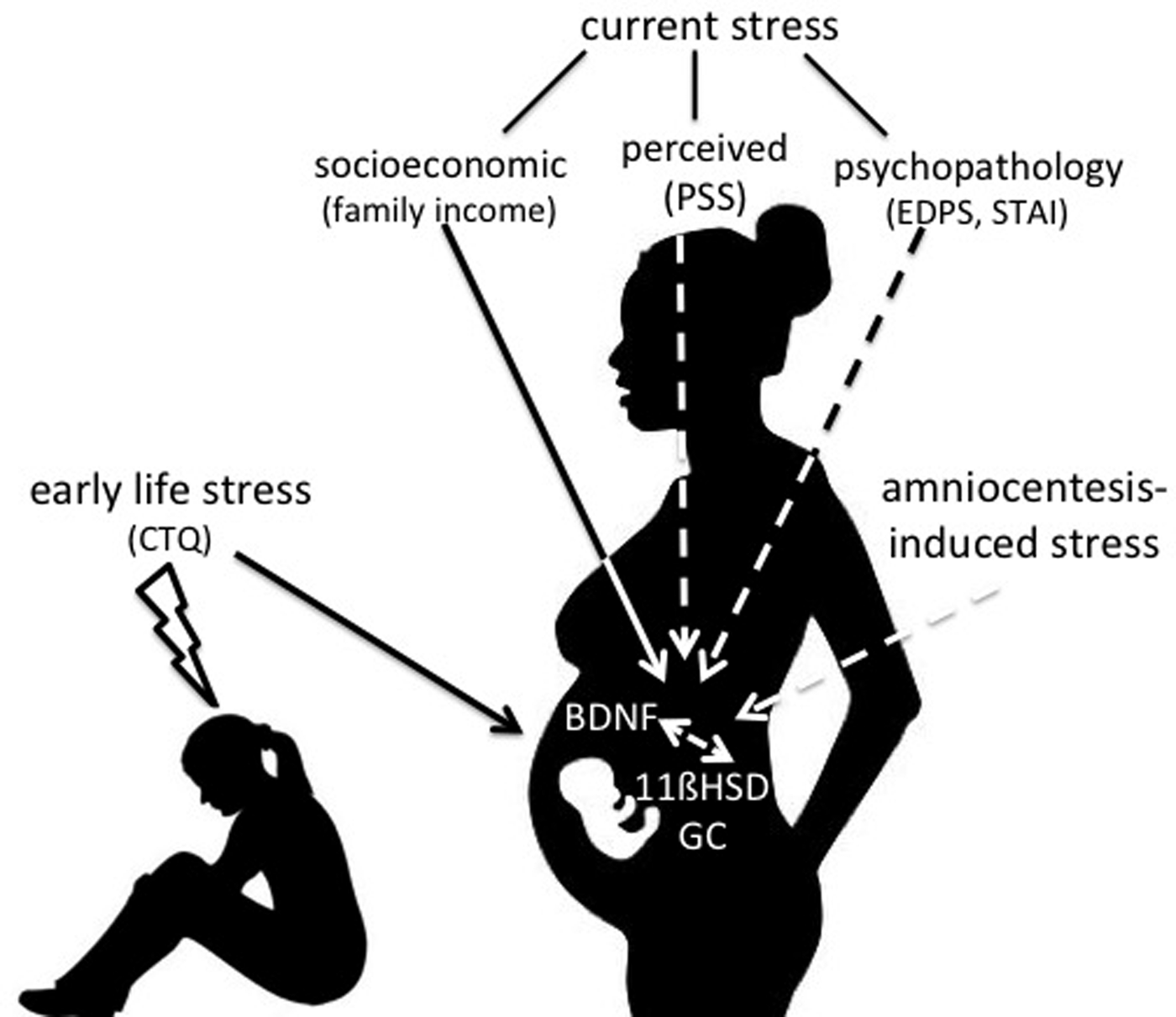
Figure 1. Study design. We tested the hypotheses that early and current maternal stress is related to BDNF and glucocorticoid concentrations in amniotic fluid.

Figure 2. CTQ ratings and BDNF in amniotic fluid (r = .435, p = .0001).
Socioeconomic status (family income) and glucocorticoid concentrations
Net family income as a proxy of socioeconomic status correlated negatively with glucocorticoid concentrations (r = –.279, p = .013) and tended to correlate negatively with BDNF (r = –.197, p = .081) and tended to be positively associated with HSD2 activity (r = .199, p = .079).
Psychiatric disorder (major depression or anxiety disorder) and HSD2 activity
Comparing the 17 participants with major depression or anxiety disorder with 62 participants without a psychiatric disorder, we found evidence for increased HSD2 activity in subjects with psychiatric diagnoses (0.746 ± 0.076 vs. 0.687 ± 0.102, Student t test, p = .030), while glucocorticoid and BDNF concentrations did not significantly differ. Specifically, we compared 62 subjects without depression with 8 subjects with major depression (n = 4: recurrent major depressive disorder; n = 4: first episode) and 9 subjects with anxiety disorders (panic disorder or generalized anxiety disorder). These subjects differed significantly with regard to EDPS (5.3 ± 4.7 vs. 14.0 ± 4.6 vs. 9.8 ± 4.0); F (2, 78) = 14.9, p < .0001; state (35.3 ± 9.0 vs. 48.3 ± 6.7 vs. 42.9 ± 9.4); F (2, 78) = 9.3, p < .0001; and trait anxiety (34.0 ± 9.0 vs. 49.4 ± 8.0 vs. 40.8 ± 9.3); F (2, 78) = 13.3, p < .001; univariate ANOVA. We found a trend for increased HSD2 activity in depressed subjects (0.687 ± 0.102 vs. 0.741 ± 0.087 vs. 0.750 ± 0.0687; F (2, 78) = 2.43, p = .095.
Amniocentesis-induced stress and glucocorticoids in amniotic fluid
Amniocentesis-induced stress was negatively related to cortisol (r = –2.79, p = .013), but not cortisone (r = –.051, ns) and HSD2 activity (r = .13, ns) in amniotic fluid.
Cortisol, cortisone, and BDNF in amniotic fluid
We used univariate ANOVA and controlled for the confounders BMI and net income as they showed an association with the dependent variables (see above), but did not find any association between either HSD2 activity, F (1, 78) = 0.68, ns, or glucocorticoid concentration, F (1, 78) = 0.03, ns, with BDNF in amniotic fluid.
Discussion
The present study analyzed the associations between maternal state anxiety, current major depression or anxiety disorder, maternal early life stressors, socioeconomic status by proxy of income, perceived stress, and the concentration of glucocorticoids, a proxy of placental HSD2 activity, and BDNF in amniotic fluid of a sample of 79 pregnant women in the early second trimester. The concentration of cortisol tended to be lower compared to a recent publication measuring steroids in amniotic fluid at a similar stage of gestation with a similar method (Fahlbusch et al., Reference Fahlbusch, Heussner, Schmid, Schild, Ruebner, Huebner and Rauh2015), indicating that the fetal glucocorticoid exposition in our cohort was not increased. However, there was no evidence for hypocortisolemia when comparing our and Fahlbusch's data. This suggests that the developing fetuses in our study were not exposed to excessive levels of glucocorticoids. Although these fetuses might have been exposed to “normative levels” of glucocorticoids, there was considerable variance in the concentrations of cortisol and cortisone in amniotic fluid, which might be due to current or past life experiences. We found robust positive associations between maternal ELS and BDNF concentrations in amniotic fluid. There was some evidence for socioeconomic status (family income) to be negatively associated with glucocorticoid concentrations in amniotic fluid. There was no significant association between current maternal psychiatric disorder (major depressive or anxiety disorder) and any of the glucocorticoid measures. Perceived stress, state anxiety, as well as amniocentesis-induced stress had no significant effects on cortisol regulation or BDNF concentrations.
Neurotrophic factors play a critical role in neurodevelopment as they promote brain development and ultimately lead to an increase in the number of functional synapses. Accordingly, animal models of intrauterine growth restriction have shown reduced expression of BDNF and other growth factors in the brain of the fetus (Fukami et al., Reference Fukami, Nakayama, Sasaki, Mimura, Mori and Watanabe2000) and low BDNF levels in amniotic fluid or cord blood has been suggested to be a marker of central nervous system disorder in utero (Chouthai, Sampers, Desai, & Smith, Reference Chouthai, Sampers, Desai and Smith2003), suggesting that BDNF in amniotic fluid may be related to the infant's central nervous system neurodevelopment.
We found BDNF to be increased in amniotic fluid of participants with evidence of adverse early life environments. Of note, the effect of ELS was independent from current psychiatric disorder (major depression or anxiety disorder). However, the CTQ scores of our cohort showed considerable variance with the mean value not being high, suggesting that our finding may not be generalized to clinical populations, especially those with posttraumatic stress disorder. There is evidence that maternal trauma is related to fetal autonomic system regulation (Gustafsson, Doyle, Gilchrist, Wernr, & Monk, Reference Gustafsson, Doyle, Gilchrist, Werner and Monk2016). In line with this observation, our report shows that early rather than current stress of the mother is related to BDNF in amniotic fluid. While the effect of maternal ELS on fetal BDNF has never been studied in humans, it has repeatedly been shown that stress during pregnancy lowers BDNF in young (Berry et al., Reference Berry, Panetta, Luoni, Bellisario, Capoccia, Riva and Cirulli2015) and adult rodent offsprings’ brains (Fumagalli, Bedogni, Perez, Racagni, & Riva, Reference Fumagalli, Bedogni, Perez, Racagni and Riva2004). In accordance with this finding, Braithwaite et al. (Reference Braithwaite, Kundakovic, Ramchandani, Murphy and Champagne2015) also described increased BDNF DNA methylation in buccal cells of depressed mothers’ newborns. However, in some states of acute environmental stress for the fetus, such as maternal infection, an increase in BDNF in the fetal brain has been reported and discussed as an adaptation to stress (Gilmore et al., Reference Gilmore, Jarskog and Vadlamudi2003). With regard to humans, there is one report of low amniotic BDNF levels in small-for-date newborns, which is a typical result of chronic intrauterine stress (Wang & Ye, Reference Wang and Ye2008). Thus, the level of BDNF in amniotic fluid may be a function of the time (e.g., early life vs. current) and duration of exposure (acute vs. chronic stress) and birth outcome of maternal stress (low birth weight; impaired neurodevelopment).
Our finding of maternal ELS to be positively related with amniotic BDNF contrasts with some findings of studies on stress during pregnancy. As with stress induced by infection (Gilmore et al., Reference Gilmore, Jarskog and Vadlamudi2003), increased amniotic BDNF in mothers with ELS may be a marker of stress or of adaptation to stress.
The mechanism of how maternal ELS may transgenerationally impact on infant BDNF is not clear. However, it may be possible that, similar to other transgenerational effects of famine or stress (Heijmans et al., Reference Heijmans, Tobi, Stein, Putter, Blauw, Susser and Lumey2008; Radtke et al., Reference Radtke, Ruf, Gunter, Dohrmann, Schauer, Meyer and Elbert2011), epigenetic mechanisms may play a role. Given the role of neurotrophic factors during pregnancy (Chouthai et al., Reference Chouthai, Sampers, Desai and Smith2003; Fukami et al., Reference Fukami, Nakayama, Sasaki, Mimura, Mori and Watanabe2000; Wang & Ye, Reference Wang and Ye2008), it may be hypothesized that maternal ELS is related to fetal neurodevelopment.
Exposure to high concentrations of glucocorticoids in utero can be considered deleterious for the fetus. Physiological levels of glucocorticoids are necessary for healthy fetal development, particularly of the lungs. However, the cortisol increase experienced by pregnant women occurs mostly in the third trimester of pregnancy (Jung et al., Reference Jung, Ho, Torpy, Rogers, Doogue, Lewis and Inder2011). The present samples come from amniocentesis performed during the second trimester, a known ontogenic window of brain development in which cortisol overexposure is thought to be neurotoxic. Glucocorticoids are considered to lower BDNF synthesis, especially during pregnancy (Yu et al., Reference Yu, Lee, Lee and Son2004). However, we did not find any relation between glucocorticoid concentration, HSD activity, and BDNF in amniotic fluid. We hypothesized that depression and cortisol in pregnancy interfere with the regulation of BDNF expression. Yet we only found a trend association between current psychiatric disorder and HSD2 activity. Moreover, Braithewaite's (Reference Braithwaite, Kundakovic, Ramchandani, Murphy and Champagne2015) study showed the association between prenatal depression and the fetal epigenetic BDNF gene regulation to be independent of cortisol. Thus, a maternal prenatal psychological condition might have lasting effects on BDNF regulation in the offspring, regardless of current cortisol levels. Mechanisms beyond glucocorticoids that may translate maternal stress in fetal development have broadly been discussed elsewhere (Beijers, Buitelaar, & de Weerth, Reference Beijers, Buitelaar and de Weerth2014).
It is believed that maternal depression during pregnancy comes along with increased HPA system activity (Davis et al., Reference Davis, Glynn, Schetter, Hobel, Chicz-Demet and Sandman2007; Giesbrecht, Campbell, Letourneau, Kooistra, & Kaplan, Reference Giesbrecht, Campbell, Letourneau, Kooistra and Kaplan2012), although several studies have failed to show this association (Braig et al., Reference Braig, Grabher, Ntomchukwu, Reister, Stalder, Kirschbaum and Genuneit2016; Shea et al., Reference Shea, Streiner, Fleming, Kamath, Broad and Steiner2007). The placental HSD2 is considered to be a barrier for maternal glucocorticoids, but is regulated by environmental factors and maternal stress (Togher et al., Reference Togher, O'Keeffe, Khashan, Gutierrez, Kenny and O'Keeffe2014). In rodents, acute cortisol infusion into pregnant mice led to an initial upregulation of placental HSD2 mRNA; this potentially adaptive mechanism faded, resulting in low mRNA levels indicating increased glucocorticoid exposure in the long run (Cuffe, O'Sullivan, Anderson, & Moritz, Reference Cuffe, O'Sullivan, Simmons, Anderson and Moritz2012). It has also been reported that acute stress leads to increased placental HSD2 expression (Welberg, Thrivikraman, & Plotsky, Reference Welberg, Thrivikraman and Plotsky2005), while chronic stress instead may cause a reduction of placental HSD2 mRNA and protein levels (Mairesse et al., Reference Mairesse, Lesage, Breton, Bréant, Hahn, Darnaudéry and Viltart2007). In humans, maternal state anxiety (as measured by STAI) and depressive symptoms (as measured by EDPS) at birth have been found to be associated with reduced activity and expression of placental HSD2 (O'Donnell et al., Reference O'Donnell, Jensen, Freeman, Khalife, O'Connor and Glover2012). The Mercy Pregnancy and Emotional Wellbeing Study showed maternal depression and anxiety to downregulate placental HSD2 (Seth, Lewis, Saffery, Lappas, & Galbally, Reference Seth, Lewis, Saffery, Lappas and Galbally2015). In contrast to these reports, our findings did not show a significant association between current psychiatric disorder and any of the glucocorticoid measures. We only found a trend for increased placental HSD2 activity in subjects with depression or anxiety disorder. One possible explanation is the moderate severity of depression (EDPS ≤ 18) in our participants. It may be that depression is related to downregulation of HSD2 in severely depressed, hypercortisolemic subjects, but not in moderately depressed women without HPA system activation. Thus, it is possible that the time of pregnancy (i.e., middle vs. late), HPA system activation, chronicity, severity of depression, or some combination of these factors may determine whether depression has an effect on fetal glucocorticoid exposure or HSD2 activity in the placenta.
Previously, it has been reported that women with low socioeconomic status in pregnancy had significantly higher evening cortisol, consistent with a pattern of chronic stress (Thayer & Kuzawa, Reference Thayer and Kuzawa2014). Moreover, infants whose mothers experienced the greatest level of socioeconomic adversity during pregnancy had the lowest placental HSD2 methylation (Appleton et al., Reference Appleton, Armstrong, Lesseur, Lee, Padbury, Lester and Marsit2013). These reports show that low socioeconomic status may be a strong stressor in pregnancy, leading to activation of the maternal HPA system and to differential methylation of placental HSD. In accordance with these reports, we found increased cortisol and total glucocorticoid concentration in amniotic fluid of women with low family income. Thus, our data suggest lowered activity of HSD2. Due to the limited power of our analysis, these findings await replication.
Contrary to our hypothesis, we did not find an association between maternal perceived stress and HPA system regulation, a finding which contrasts with earlier reports, relating perceived stress in middle pregnancy to maternal HPA system activity as measured by hair cortisol (Hoffman, Mazzoni, Wagner, Laudenslager, & Ross, Reference Hoffman, Mazzoni, Wagner, Laudenslager and Ross2016). This inconsistency may be due to differences in method of HPA measures. In contrast to hair, which reflects cortisol levels over time, measuring glucocorticoids in amniotic fluid is indirect and reflects glucocorticoids at a specific time point. Thus, we might have missed an effect of chronic maternal HPA activation that would better be reflected in hair cortisol. Maternal HPA system activity is of relevance for the outcome of pregnancy (as measured by preterm birth or low birth weight), but this was not the focus of our research.
Similarly, Ehlert's group elegantly measured the maternal HPA response to the stress of amniocentesis (Ghaemmaghami et al., Reference Ghaemmaghami, Dainese, La Marca, Zimmermann and Ehlert2014). The nature of our study design did not allow measuring the immediate effect of amniocentesis-induced stress on maternal HPA system as our study participants were informed and gave consent after amniocentesis. We found only a moderate effect of self-rated amniocentesis-induced stress on cortisol in amniotic fluid. The direction of the association rather suggested high subjective amniocentesis-induced stress to lead to low cortisol in amniotic fluid.
Several limitations of our study should be noted. We did not directly measure maternal HPA system activity, which is a clear limitation of our study. Similarily, we did not directly measure 11ßHSD activity, but used the cortisol:cortisone ratio in amniotic fluid as a proxy, which might not be completely independent from current maternal HPA activity. Cortisol and cortisone in amniotic fluid have been used to estimate placental 11ßHSD acitivity (Ghaemmaghami et al., Reference Ghaemmaghami, Dainese, La Marca, Zimmermann and Ehlert2014). To the best of our knowledge, however, there is no direct proof in humans that amniotic fluid glucocorticoid concentrations mirror placental 11ßHSD activity. We found interesting associations of maternal ELS and socioeconomic status with BDNF and steroids in amniotic fluid, but it is unclear whether or not these endocrine changes reflect developmental changes in the fetus. In addition, most stressors such as severity of depression in our study participants were moderate and different or stronger effects on glucocorticoids or BDNF could be expected in severely stressed pregnant women, who could not be recruited in this study. We were able to compare stressed and nonstressed women. However, women electing amniocentesis are a self-selected population that may not be comparable to the general population. Next, it should be noted that as we consider this first trial on BDNF, glucocorticoids, and stress in a difficult to recruit mid-pregnancy group of women a hypothesis-generating study, we do not correct for multiple testing. However, testing 30 hypotheses (6 independent and 5 dependent variables), the association between maternal ELS (CTQ) and BDNF concentration in amniotic fluid would have survived even an adjusted α error of 0.002. Finally, our independent variables were partly interrelated. While we control for interaction effects in the MANOVA model, the interpretation of associations of single independent parameters with the dependent variables is limited due to interrelatedness of the independent variables. Therefore, we focussed our analyses on the independent variables showing significant effects in the fully controlled MANOVA model.
Taken together, we found evidence that adverse events early in maternal life are reflected in increased BDNF concentration in amniotic fluid. Further exploration into possible epigenetic mechanisms of these findings is needed. Low socioeconomic status is directly related to increased glucocorticoid exposure of the fetus. While net family income is related to glucocorticoid exposure of the fetus, psychological dimensions of current stress showed no significant associations with glucocorticoids in amniotic fluid. Our findings support the notion that environmental conditions during pregnancy and the mother's early life may have an impact on physiology and development of the fetus. While the association of prenatal stress with the offspring's health and behavior is front and center in current neuroscience research, our findings suggest that the role of maternal stress and the environment in early life deserves more attention.




