The parent–child relationship is a key source of children's development. Research suggests that parenting is associated with a wide range of psychopathological outcomes in youth, including substance abuse problems (Lynch et al., Reference Lynch, Turkheimer, D'Onofrio, Mendle, Emery and Slutske2006), conduct problems (Burt, Krueger, McGue, & Iacono, Reference Burt, Krueger, McGue and Iacono2003; Burt, McGue, Krueger, & Iacono, Reference Burt, McGue, Krueger and Iacono2005; Klahr, McGue, Iacono, & Burt, Reference Klahr, McGue, Iacono and Burt2011; Klahr, Rueter, McGue, Iacono, & Burt, Reference Klahr, Rueter, McGue, Iacono and Burt2011), and anxiety problems (McLeod, Wood, & Weisz, Reference McLeod, Wood and Weisz2007). Although most research focuses on the influences of parents on children, the process of parenting a child is also a transformative experience in adult development. The effects of parents on children may be rivaled in importance only by the effects of children on parents. Raising children is associated with changes in parental personality and attitudes (e.g., increased conservatism; Antonucci & Mikus, Reference Antonucci, Mikus, Michaels and Goldberg1988), higher well-being (Huijts, Kraaykamp, & Subramanian, Reference Huijts, Kraaykamp and Subramanian2011; Menaghan, Reference Menaghan1989), lower marital relationship quality (Twenge, Campbell, & Foster, Reference Twenge, Campbell and Foster2003), and even measurable brain activity in response to child-related stimuli (e.g., stronger amygdala responses to crying in parents compared to nonparents; Seifritz et al., Reference Seifritz, Esposito, Neuhoff, Lüthi, Mustovic and Dammann2003). Given the importance of the parent–child relationship in the lives of both parents and children, research identifying factors that impact the nature and quality of the parent–child relationship constitutes an important contribution to the literature.
Extant research has indicated that parenting behaviors with their children are multidetermined—a function of the broader family context (e.g., parental marital quality and neighborhood characteristics), the parent's own rearing environment, and broader societal and cultural factors (Belsky & Jaffee, Reference Belsky and Jaffee2006; Kendler, Sham, & MacLean, Reference Kendler, Sham and MacLean1997; Maestripieri, Reference Maestripieri1999). Characteristics of the parent are also key factors in shaping parenting behavior. These characteristics include biological factors (e.g., neuroendocrine and genetic influences; Maestripieri, Reference Maestripieri1999), parent personality (Prinzie, Stams, Deković, Reijntjes, & Belsky, Reference Prinzie, Stams, Deković, Reijntjes and Belsky2009), and psychopathology (Dix & Meunier, Reference Dix and Meunier2009; Jaffee, Belsky, Harrington, Caspi, & Moffitt, Reference Jaffee, Belsky, Harrington, Caspi and Moffitt2006). Although these sociocultural and parent-driven factors have a clear role in shaping parenting behaviors, children also influence the parenting they receive. Supportive evidence for this “child effects model” comes from a seminal study by Anderson, Lytton, and Romney (Reference Anderson, Lytton and Romney1986) in which conduct- disordered and nonproblem boys, and their respective mothers, interacted in unrelated pairs. When interacting with nonproblem children, the mothers of conduct-disordered children did not differ from the mothers of nonproblem children in commands, positive behaviors, or negative behaviors. However, all mothers gave more commands to children with conduct disorder than to nonproblem children, suggesting that the maladaptive interactions between conduct-disordered boys and their mothers were driven at least partially by the child. Cunningham and Barkley (Reference Cunningham and Barkley1979) similarly found that when children's nonattentive and noncompliant behavior is improved by the administration of stimulant drugs, their mothers become less controlling and mother–child interactions are nearly normalized. Children thus do not appear to be passive recipients of the parenting provided by their caregivers; rather, they play an active role in evoking these parenting experiences. Put simply, children's behavior serves to shape the behaviors of their caregivers in a fluid, lifelong exchange.
Given that child behavior and temperament are at least partially genetically influenced (Burt, Reference Burt2009; Goldsmith, Lemery, Buss, & Campos, Reference Goldsmith, Lemery, Buss and Campos1999), it further follows that children's genetic predispositions may also shape the parenting they receive. In particular, it may be the case that genetically influenced characteristics of the child elicit specific behaviors from the parent, a process referred to as an evocative gene–environment correlation (rGE; Scarr & McCartney, Reference Scarr and McCartney1983). Child-based twin studies of parenting are particularly well suited for identifying these evocative rGE effects (Klahr & Burt, Reference Klahr and Burt2012). Because the twins in this design are the children (rather than the parents), the interpretation of the genetic and environmental effect estimates must also occur at the level of the children (who are experiencing, rather than providing, the parenting behavior in question). Genetic influences on parenting obtained from child-based twin designs thus do not index the impact of parental genes on the parenting behavior they provide but instead capture the influence of the child's genetic makeup on the parenting behavior he or she receives (via evocative rGE). Estimates of the shared environment in child-based designs index factors that increase similarity in the parent's behavior across children regardless of the twins' degree of genetic relatedness. Such factors would include a number of potential predictors of parenting behavior, including parental genes, parental personality characteristics, broader familial practices, and sociocultural expectations regarding child rearing. As an example, the genetic similarity between parents and their biological children is always 50%, regardless of the level of genetic similarity between the twins. Parental genes (and passive rGEs, as described in Scarr & McCartney, Reference Scarr and McCartney1983) should thus load onto the shared environment in child-based twin designs (Neiderhiser et al., Reference Neiderhiser, Reiss, Pedersen, Lichtenstein, Spotts and Hansson2004). Estimates of the nonshared environment would, by contrast, capture differences in parental responses to their children (e.g., if one twin suffers an accident or illness and subsequently receives different parenting than the co-twin).
Using this interpretative framework, numerous child-based twin studies have uncovered significant genetic influences (and thus significant evocative rGE) on parenting and the parent–child relationship. These findings persist across child informant reports (Harlaar et al., Reference Harlaar, Santtila, Björklund, Alanko, Jern and Varjonen2008; Herndon, McGue, Krueger, & Iacono, Reference Herndon, McGue, Krueger and Iacono2005; Kendler, Reference Kendler1996; Lichtenstein et al., Reference Lichtenstein, Ganiban, Neiderhiser, Pedersen, Hansson and Cederblad2003; Neiderhiser, Reiss, Lichtenstein, Spotts, & Ganiban, Reference Neiderhiser, Reiss, Lichtenstein, Spotts and Ganiban2007; Neiderhiser et al., Reference Neiderhiser, Reiss, Pedersen, Lichtenstein, Spotts and Hansson2004; Plomin, McClearn, Pedersen, Nesselroade, & Bergeman, Reference Plomin, McClearn, Pedersen, Nesselroade and Bergeman1988; Plomin, Reiss, Hetherington, & Howe, Reference Plomin, Reiss, Hetherington and Howe1994; Wade & Kendler, Reference Wade and Kendler2000), parent informant-reports (Kendler, Reference Kendler1996; Neiderhiser et al., Reference Neiderhiser, Reiss, Pedersen, Lichtenstein, Spotts and Hansson2004, Reference Neiderhiser, Reiss, Lichtenstein, Spotts and Ganiban2007; Plomin et al., Reference Plomin, Reiss, Hetherington and Howe1994; Wade & Kendler, Reference Wade and Kendler2000), and observer reports (Deater-Deckard & O'Connor, Reference Deater-Deckard and O'Connor2000; Eley, Napolitano, Lau, & Gregory, Reference Eley, Napolitano, Lau and Gregory2010; Rende, Slomkowski, Stocker, Fulker, & Plomin, Reference Rende, Slomkowski, Stocker, Fulker and Plomin1992), suggesting that the findings of evocative rGE influences on parenting are robust to sampling and methodological considerations.
Additional evidence for evocative rGE effects on parenting has also been uncovered within child-based adoption study designs, which similarly rely on the interpretive framework for child-based designs described above. One such examination found that children at genetic risk for antisocial behavior (as indexed by birth mother self-reported antisocial behavior) were more likely to receive negative parenting from their adoptive parents (O'Connor, Deater-Deckard, Fulker, Rutter, & Plomin, Reference O'Connor, Deater-Deckard, Fulker, Rutter and Plomin1998). Similar results have also been found among adopted adolescents and their parents (Ge et al., Reference Ge, Conger, Cadoret, Neiderhiser, Yates and Troughton1996). In sum, a growing body of research thus supports the notion that child-driven influences, including genetic influences and particularly evocative rGE processes, partially underlie the parent–child relationship.
Despite the substantial evidence supporting the importance of child-driven genetic influences, the specific processes by which children's genetic characteristics evoke parenting behaviors remain largely unknown. Research has only just begun to identify the specific genetically influenced characteristics of children that are most important for predicting parenting. Moreover, no study to date has explored whether and how these genetically driven influences play out in day-to-day interactions between parents and children. A recent study by Eley and colleagues (Reference Eley, Napolitano, Lau and Gregory2010), for example, found that observed maternal control overlapped significantly with child self-reported anxiety and that this overlap was primarily genetic in origin. These findings suggest that genetic influences on anxious behaviors in children may elicit particular parental behaviors. Nevertheless, because child anxiety was measured globally using a child-report measure (rather than during a specific interaction), they were not able to identify whether or how child anxiety served to elicit parental control in the moment. This is also the case for the adoption studies noted above (O'Connor et al., Reference O'Connor, Deater-Deckard, Fulker, Rutter and Plomin1998; Ge et al., Reference Ge, Conger, Cadoret, Neiderhiser, Yates and Troughton1996), which indicate that genetic risk for antisocial behavior in adolescents may influence parental behavior but do not clarify how this process unfolds over time.
One framework from which to examine this unfolding of interpersonal behavior in the parent–child relationship is provided by contemporary interpersonal theory (Kiesler, Reference Kiesler1996; Pincus, Reference Pincus2005). Interpersonal theory classifies interpersonal behavior along the two orthogonal dimensions of control (dominance to submission) and affiliation (coldness to warmth) using the interpersonal circumplex (IPC; see Figure 1) as a measurement model (Leary, Reference Leary1957; Wiggins, Reference Wiggins1996). The IPC has been found to reliably and validly measure interpersonal behavior across a range of dyads, including adult peers (Sadler et al., Reference Sadler, Ethier, Gunn, Duong and Woody2009), romantic partners (Markey, Markey, & Tinsley, Reference Markey, Markey and Tinsley2005), and therapist–patient relationships (Anchin & Pincus, Reference Anchin and Pincus2010).

Figure 1. The interpersonal circumplex.
The constructs examined in interpersonal theory are variations of those typically examined in the parenting literature. The orthogonal dimensions of warmth and control in the IPC are analogous to the dimensions of demandingness and responsiveness in parenting first suggested by Baumrind (Reference Baumrind1971) and later clarified by Maccoby and Martin (Reference Maccoby, Martin, Mussen and Hetherington1983). Parental behaviors can thus be divided into four quadrants or parenting styles: authoritative, authoritarian, neglectful, or indulgent (see Figure 2), which are equivalent to the corresponding quadrants in the IPC. Moreover, the IPC provides added value in that both child and parent behavior can be captured simultaneously within the same framework. In this way, the relationship between parent and child behaviors can be examined as they unfold. An IPC model of the mother–child relationship thus places emphasis on both parent and child-driven behaviors, which is a significant improvement over simpler models of parenting that account only for the behavior of the parent. The prediction that a person's behavior in one moment influences the behavior of the other person in a subsequent moment is a core component of interpersonal theory (Leary, Reference Leary1957). Interpersonal theory and the IPC are therefore well suited not only for identifying specific genetically influenced child behaviors that evoke parental responses but also for evaluating how evocative rGE processes might unfold during mother–child interactions.

Figure 2. Two dimensions of parenting and four parenting styles. Baumrind (Reference Baumrind and Pick1973) and Maccoby and Martin (Reference Maccoby, Martin, Mussen and Hetherington1983) theorized that there are two factors in parental behavior (responsiveness and demandingness) and four subsequent parenting styles. [A color version of this figure can be viewed online at http://journals.cambridge.org/dpp]
We thus sought to examine the genetic and environmental etiology of mother and child interpersonal behaviors as assessed on a moment-by-moment basis. To do so, we employed a recently developed video coding technique that utilizes a computer joystick to capture ratings of interpersonal behavior twice per second, allowing for a relatively continuous measure of interpersonal behaviors (Sadler et al., Reference Sadler, Ethier, Gunn, Duong and Woody2009). These ratings can then be averaged across each minute of the interaction to provide a minute-by-minute index of each individual's interpersonal behavior. We would argue that these real-time ratings are better suited for uncovering the etiology of parenting behavior than are the global ratings typically obtained in observer-rating studies (i.e., when coders provide one overall assessment of the entire interaction; Neiderhiser et al., Reference Neiderhiser, Reiss, Pedersen, Lichtenstein, Spotts and Hansson2004; O'Connor, Hetherington, Reiss, & Plomin, Reference O'Connor, Hetherington, Reiss and Plomin1995). Global ratings may be heavily influenced by recency or primacy effects, such that late or early events are recalled more readily than events that transpired during the middle portion of the video (Stone & Shiffman, Reference Stone and Shiffman1994). Assessing process data on a momentary basis, by contrast, allows each moment to be weighted equally. Global ratings are also likely to be more heavily influenced by moments that stood out to the rater (e.g., because they were emotional, unexpected, or significant; Thomas, Hopwood, Ethier, & Sadler, Reference Thomas, Hopwood, Ethier and Sadler2012). In short, moment-by-moment ratings, even when they are collapsed across each minute of the interaction, have significant advantages over global ratings for our understanding of the etiology of interpersonal behavior. As such, we sought to determine whether evocative rGE influences on the mother–child relationship can be observed in real-time interactions. We also sought to examine the genetic and environmental overlap between child and maternal interpersonal behaviors in order to begin to elucidate the processes by which child-driven genetic influences impact parents. Should there be evidence of genetic influences on maternal behavior within moment-by-moment mother–child interactions, it would provide the clearest confirmation for the role of evocative rGE in the mother–child relationship to date. Moreover, information on the extent to which genetic influences on maternal behavior overlap with genetic influences on child behavior over the course of the interaction would begin to explicate how these evocative rGE unfold over time.
Method
Participants
Participants from this study were a part of the population-based Michigan State University Twin Registry (MSUTR), which includes several independent twin projects (Klump & Burt, Reference Klump and Burt2006). The current study included 546 twin families who were assessed as part of the ongoing Twin Study of Behavioral and Emotional Development in Children (TBED-C) within the MSUTR. The TBED-C includes both a completed population-based sample (current N = 500 families; video data were available for 485 families with 970 mother–child dyad pairs) and an ongoing at-risk sample for which inclusion criteria also specified that participating twin families lived in moderately to severely disadvantaged neighborhoods (current N = 61 families; 122 mother–child dyad pairs). Results were identical with and without the at-risk sample, and thus these families were retained for analysis. Assessments took place either in our laboratory at Michigan State University or in participant's homes (in the event that families were unable to travel to the university, current n = 38 familiesFootnote 1). Children provided informed assent, and parents provided informed consent for themselves and their children. The twins ranged in age from 6 to 10 years (mean = 8.23, SD = 1.44; although a few twins had turned 11 by the time they participated) and were 47.4% female. To be eligible for participation in the TBED-C, neither twin could have a cognitive or physical handicap (as assessed via parental screen) that would preclude completion of the assessment.
Families for both samples were recruited via State of Michigan birth records in collaboration with the Michigan Department of Community Health (MDCH). The MDCH manages birth records in Michigan, which are confidential, and is therefore able to identify all twin births in the state. MDCH identified twins living within 120 miles of East Lansing, Michigan, who were between the ages of 6 and 10 years. Twins were identified either directly from birth records or via the Michigan Twins Project, a large-scale population-based registry of twins in lower Michigan that were themselves recruited via birth records. MDCH then utilized the Michigan Bureau of Integration, Information, and Planning Services database to locate current addresses through parent driver's license information. Using these addresses, MDCH mailed premade recruitment packets to parents of twins. A reply postcard was included for parents to indicate their interest in participating, which was addressed to the MSUTR researchers. Interested families were then contacted directly by project staff. Parents who did not respond to the first mailing were sent additional mailings approximately 1 month apart until either a reply was received or up to four letters had been mailed.
This recruitment strategy yielded an overall response rate of 62%, which is similar to or better than those of other twin registries that use anonymous recruitment mailings (Baker, Barton, & Raine, Reference Baker, Barton and Raine2002; Hay, McStephen, Levy, & Pearsall-Jones, Reference Hay, McStephen, Levy and Pearsall-Jones2002). The final population-based sample was broadly representative of the area population and of recruited families more specifically (as assessed via a brief questionnaire screen administered to 70% of nonparticipating families). Fourteen percent of families in the sample lived at or below federal poverty guidelines, as compared to 14.8% across the state of Michigan. Participating families endorsed ethnic group memberships at rates comparable to area inhabitants (e.g., 86.4% and 85.5% Caucasian and 5.4% and 6.3% African American for the participating families and the local census, respectively). Participating twins did not differ from nonparticipating twins in their average levels of conduct problems, emotional symptoms, and hyperactivity (as assessed via the Strength and Difficulties Questionnaire; Goodman & Scott, 1999; Cohen d standardized effect sizes = –0.047, 0.010, and –0.076, respectively; all ps ≥ .29). Participating families also did not differ from nonparticipating families (all ps ≥ .16) in paternal felony convictions (d = –0.08), paternal years of education (d = 0.00), proportion of Caucasian twins (d = 0.01), rate of single parent homes (d = –0.09), twin zygosity (d = –0.08), maternal or paternal age at assessment (both d ≤ 0.10), use of fertility medications to conceive the twins (d = –0.05), number of children in the home (d = –0.10), or maternal and paternal alcohol problems (d = 0.08 and 0.04, respectively). However, participating mothers reported slightly more years of education (d = 0.17, p = .02) and a slightly lower rate of felony convictions (d = –0.19, p = .03) than did nonparticipating mothers. A trend was also observed for family income, although income was slightly lower in participating as compared to nonparticipating families (d = –0.13, p = .06).
Zygosity was established using physical similarity questionnaires administered to the twins' primary caregiver (Peeters, Van Gestel, Vlietinck, Derom, & Derom, Reference Peeters, Van Gestel, Vlietinck, Derom and Derom1998). There were a total of 259 monozygotic (MZ) twin pairs and 287 dizygotic (DZ) twin pairs, including 26 opposite-sex DZ twin pairs (the at-risk sample includes opposite-sex twin pairs as well). On average, the physical similarity questionnaires used by the MSUTR have accuracy rates of 95% or better.
Procedures
Observer ratings of parent and child interpersonal behavior were obtained using 80-min videotaped interactions of mother–child dyads. Mothers interacted separately with each of their twins. The on-campus interactions took place in laboratory offices that were set up to resemble living rooms, with cameras inconspicuously installed in the ceiling. For those assessments occurring in participants' homes, interactions took place in a family living space with a video camera placed on a tripod in the room. Each mother–child dyad was asked to complete a mildly to moderately frustrating task (i.e., use an Etch-a- Sketch to draw specific pictures, but parent and child may only use one dial each, thereby requiring cooperation; Deater-Deckard, Pylas, & Petrill, Reference Deater-Deckard, Pylas and Petrill1997). The task was originally designed for use in child twin families and has been found to be a reliable and valid tool for assessing the parent–child relationship with school-age children.
Joystick coding method
Trained observers viewed the interactions and coded interpersonal behavior using the joystick method of video coding (Sadler et al., Reference Sadler, Ethier, Gunn, Duong and Woody2009). For this method, a computer joystick apparatus (the Microsoft Sidewinder Force Feedback 2) is utilized to provide momentary ratings of an individual's interpersonal behavior throughout the course of an interaction. The joystick-monitoring software program displays a Cartesian plane that is approximately 6.8 cm wide by 6.6 cm tall on the computer screen and depicts the axes of interpersonal theory. A dot is shown in the Cartesian plane and moves in accordance with the current position of the joystick. The x axis represented warmth, and the y axis represented control. The scale on both axes ranged from –1000 to 1000, with 1000 on the x axis indicating extreme warmth and 1000 on the y axis indicating extreme control (see Figure 1).
Observers were trained to use movement along the horizontal axis to indicate changes in warmth-related behaviors and movement along the vertical axis to indicate changes in control-related behaviors. Horizontal and vertical movements may occur simultaneously and to varying degrees in order to indicate behaviors that are relevant along both dimensions (as was typically the case). The software program was set to write the joystick position within the Cartesian plane (i.e., the x and y coordinates) to a text file twice per second. Trained observers were able to view the videotaped interactions and the joystick monitor on the screen simultaneously, in order to continuously watch the interaction and move the joystick position accordingly. Raters coded the interpersonal behaviors of one person in the interaction at a time. Thus, each interaction was viewed and rated twice, once for the mother and once for the child. Ratings of the same interaction were never obtained sequentially.
Training
Each observer underwent approximately 10 hr of training with the joystick apparatus, following the procedures outlined in Sadler et al. (Reference Sadler, Ethier, Gunn, Duong and Woody2009). Training included familiarization with the computer joystick apparatus and software, practice using the joystick to rate interpersonal descriptors, and the viewing and coding of mother–child interactions with live observation and feedback from the first two authors of the study. Because of the time-dependent nature of the data, the critical importance of starting the joystick software at the correct time was emphasized. Coders were instructed to begin coding exactly 5 s into the beginning of a video clip.
Trainees were further instructed to pay close attention to both verbal and nonverbal interpersonal behaviors and to code moment-to-moment changes in interpersonal behavior rather than any general sense of the interaction overall. Examples of warm behaviors include social smiling or leaning toward the other person, verbal praise, eye contact, and warm physical contact (such as a hug or pat on the back), whereas behaviors such as looking away, unresponsiveness, and rude or sarcastic comments are indicative of coldness. Examples of controlling behaviors include giving instructions and grabbing the Etch-a-Sketch toy, whereas submissive behaviors include following the other person's lead and asking for permission. During a lull in behavior, coders were instructed to allow the joystick to remain stationary (unless the lull became interpersonally meaningful; e.g., if a child ignored a question asked by the mother).
Warmth and control
A total of 12 research assistants worked as coders on this project. In order to minimize idiosyncratic interpretations/random error by individual coders, coders were divided into teams of 3 to 4, all of whom coded the interactions assigned to that team. Ratings were then averaged across coders to obtain a composite rating for each moment of the interaction, as suggested by Sadler et al. (Reference Sadler, Ethier, Gunn, Duong and Woody2009). The composite ratings of each moment were then averaged across all moments, separately for each min of the interaction, creating mean scores of warmth and control for each member of the dyad for each minute (i.e., mothers and children each received an overall score for both control and warmth during Minutes 0–1,1–2, 2–3, 3–4, 4–5, 5–6, and 6–7Footnote 2). These mean scores were thus the average of moments within a given minute rather than global summary scores of the interaction. In order to capture overall levels of warmth and control over the entire interaction, we also computed averages of warmth and control across all 8 min. Different teams coded the two mother–twin dyads within each family. Twins in a given family were thus always coded by different coders, thereby eliminating any inflation of within-family correlations due to shared “informant” effects. The same teams of coders coded both the mother and child within each dyad, although the two were never coded consecutively.
Reliability was calculated in two ways. We calculated moment-by-moment reliability by examining correlations between raters over a time series. Moment-by-moment reliability from the training sessions was calculated by comparing the trainee's ratings to the ratings of the study's first two authors (r = .68 for warmth, r = .86 for control) to ensure rater's aptitude in the coding procedures. Following the coding of the data, interrater reliability of coding teams was also calculated for minute average levels of warmth and control across the entire sample using the Cronbach alpha. For the team coding older twins and their mothers, reliability was 0.87 for child control (range across the 7-min segments = 0.86–0.91), 0.59 for child warmth (range = 0.24–0.67), 0.89 for maternal control (range = 0.87–0.96), and 0.73 for maternal warmth (range = 0.56–0.75). For the teams coding the younger twins and their mothers, reliability was 0.93 for child control (range = 0.90–0.95), 0.69 for child warmth (range = 0.53–0.78), 0.89 for maternal control (range = 0.89–0.96), and 0.68 for maternal warmth (range = 0.48–0.89).
Statistical analyses
Twin methodology uses the difference in the proportion of genetic variance shared between MZ and DZ twin pairs to estimate genetic and environmental contributions to variance in observed behaviors or characteristics (phenotypes). MZ, or identical, twins result from a single fertilized zygote splitting and hence share 100% of their segregating genes. DZ, or fraternal, twins are the result of two independent conceptions and so, like all full siblings, share an average of 50% of their segregating genes. Utilizing these differences, the variance within observed behaviors is partitioned into three components: additive genetic effects (a 2), shared environmental effects (c 2), and nonshared environmental effects plus measurement error (e 2). The additive genetic component (a 2) is the effect of individual genes summed over loci. Additive genetics effects, if acting alone, would create MZ correlations that are double those of DZ correlations. The shared environment (c 2) is that part of the environment that is common to both members of a twin pair and acts to make the twins within a pair similar to each other regardless of the proportion of genes shared. Nonshared environment (e 2) encompasses environmental factors unique to each twin within a pair. Nonshared environment, which also does not differ by zygosity, differentiates each twin within a pair, making them less similar. Measurement error, which similarly acts to reduce both MZ and DZ similarity, is also contained within e 2.
Crucial to this methodology is the equal environments assumption, which assumes that MZ twin pairs are no more likely to share the environmental factors that are etiologically relevant to the phenotype under study than are DZ twin pairs. Under this assumption, any differences in the MZ and DZ correlations are due to differences in their genetic similarity. The equal environments assumption has been repeatedly tested and found to be valid for numerous phenotypes (Hettema, Neale, & Kendler, Reference Hettema, Neale and Kendler1995; Kendler, Neale, Kessler, Heath, & Eaves, Reference Kendler, Neale, Kessler, Heath and Eaves1993; Morris-Yates, Andrews, Howie, & Henderson, Reference Morris-Yates, Andrews, Howie and Henderson1990; Scarr & Carter-Saltzman, Reference Scarr and Carter-Saltzman1979), but it remains an assumption for any particular phenotype until subjected to empirical testing. (For a more detailed explanation of twin study methodology, interested readers are referred to Plomin, DeFries, McClearn, & McGuffin, Reference Plomin, DeFries, McClearn and McGuffin2008.)
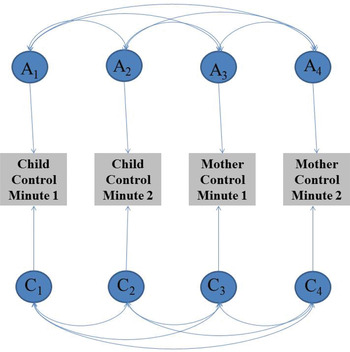
For our primary analyses, we first made use of a standard bivariate correlated factors model in which the variance within and the covariance between overall (i.e., across the entire interaction) child and maternal warmth were decomposed into their genetic and environmental components. This enabled us to obtain estimates of etiological influences on overall child and mother interpersonal behaviors, as well as on their overlap. In order to further assess the nature of etiological overlap over time, however, we made use of a series of multivariate correlated factors models in which the variance within and the covariance between child and maternal warmth across 2 min were decomposed into their genetic and environmental components (e.g., child warmth in Minutes 0–1 and 1–2 along with maternal warmth in Minutes 0–1 and 1–2; see Figure 3). Analyses were repeated for each pair of consecutive minutes (1–2, 2–3, 3–4, etc.) for both control and warmth.

Figure 3. The multivariate correlated factors model. Path diagram of a full genetic (A), shared environmental (C), and nonshared environmental (E) correlated factors model for child and maternal control over Minutes 1 and 2 of the interaction. The variance in each phenotype is parsed into additive genetic effects (A1, A2, A3, and A4), shared environmental effects (C1, C2, C3, and C4), and nonshared environmental effects (E1, E2, E3, E4, not pictured). Genetic and environmental correlations are indicated via the double-headed arrows. [A color version of this figure can be viewed online at http://journals.cambridge.org/dpp]
Although the etiology of child interpersonal behavior is quite straightforward to interpret (e.g., they represent genetic and environmental influences on the children's behaviors), the etiology of maternal behavior requires additional inference within a child-based design. As noted previously, genetic influences on maternal interpersonal behavior in this design do not reflect the mother's genetic influences but rather her response to the genetically influenced characteristics in her children. In other words, the presence of genetic influences on maternal interpersonal behavior reflects that mothers are responding more similarly to their MZ twins than to their DZ twins.
We also examined genetic and environmental correlations to more fully specify the proportion of genetic and environmental influences on child interpersonal behavior that persist to maternal interpersonal behavior (e.g., the correlation between genetic influences on child control during Minute 1 and genetic influences on maternal control during Minute 2). A genetic correlation of 1.0 would indicate that the genetic influences on maternal control, for example, overlap entirely with those on child control. A correlation of 0.0 would indicate no genetic overlap across mother and child behavior. These models thus enabled us to explicitly estimate the extent to which genetic and environmental influences on child interpersonal behavior contribute to maternal interpersonal behavior overall (in the bivariate model) and during a 2-min slice of the interaction (in the multivariate models).
Because there was a small amount of missing data (2.8% of videos were not able to be coded; there was no sound, the file was corrupted, etc.), we made use of full information maximum likelihood raw data techniques, which produce less biased and more efficient and consistent estimates than techniques like pairwise or listwise deletion in the face of missing data (Little & Rubin, Reference Little and Rubin1987). Mx (Neale, Boker, Xie, & Maes, Reference Neale, Boker, Xie and Maes2003) was used to fit models to the raw data. In order to control for potential differences due to child age, sex, or ethnicity, these variables were regressed out of the data prior to analysis, a commonly employed procedure in the field of behavior genetics (McGue & Bouchard, Reference McGue and Bouchard1984). When fitting models to raw data, variances, covariances, and means of those data are first freely estimated by minimizing minus twice the log likelihood (–2LL). The minimized value of –2LL in the baseline model was then compared with the –2LL obtained in the biometric models to yield a likelihood ratio chi-square test. The χ2 was then converted to the Akaike information criterion (AIC; Akaike, Reference Akaike1987), AIC = χ2 (2 × df) in order to measure model fit relative to parsimony. Lower AIC values reflect better fit.
Results
Descriptive statistics
As seen in Table 1, mothers exhibited significantly higher mean levels of control (Cohen d = 0.76; p < .01) as compared to their children. Mothers similarly exhibited higher mean levels of warmth, although this difference was less pronounced (Cohen d = 0.27; p < .01). Furthermore, standard deviations for mother and child control were greater than for mother and child warmth (F = 250.09, p < .01; F = 83.52, p < .01, for control and warmth, respectively), suggesting that there was greater variability in controlling behaviors during the interaction than in warmth behaviors. Phenotypic correlations are presented in Table 2. It is worth noting that maternal control and maternal warmth were not entirely orthogonal (as would be expected based on interpersonal theory). Instead, lower maternal warmth was modestly associated with higher maternal control.
Table 1. Mean levels of overall warmth and control for twins and mothers

Note: Average control and warmth scores could conceivably range from –1000 to 1000. A zero thus reflects neither dominant nor submissive (or neither warm nor cold) interpersonal behavior.
**p < .01. Means are significantly different between mothers and children.
Table 2. Phenotypic and intraclass correlations for twin and maternal overall levels of warmth and control

Note: Intraclass correlations are underscored. Cross-twin, cross-trait correlations are bold. Monozygotic correlations are below the diagonal, and dizygotic correlations are above the diagonal.
*p < .05. **p < .01.
Intraclass correlations
Prior to model fitting analyses, intraclass and cross-twin, cross-trait correlations were computed for MZ and DZ twin pairs using the overall mean scores of warmth and control in order to obtain a preliminary indication of the extent to which genetic and environmental factors influence twin and mother interpersonal behaviors. MZ correlations, which are double those of DZ correlations, are indicative of genetic effects, whereas MZ correlations that are less than double but still greater than DZ correlations suggest the importance of genetic and shared environmental effects. As seen in Table 2, intraclass correlations for twin and maternal control suggest the presence of genetic and shared environmental effects (twin control MZ r = .60, DZ r = .39; maternal control MZ r = .55, DZ r = .44). The intraclass correlations for twin warmth also suggest genetic and shared environmental effects, whereas the intraclass correlations for maternal warmth suggest an absence of genetic influence. Notable are the cross-twin, cross-trait correlations between, for example, the warmth of Twin 1 and the warmth of the mother with Twin 2 (also in Table 2). These correlations suggest that maternal control with Twin 1 is significantly correlated with the level of control exhibited by Twin 2 and that this association is stronger for MZ twins than DZ twins. Such findings are suggestive of genetic influences on the overlap between child and maternal control. However, this pattern of results was not observed for warmth.
Bivariate model-fitting analyses
Bivariate model-fitting results are presented in Table 3. For control (baseline model: –2LL = 4440.25 on 2055 df; ACE model: –2LL = 4480.10 on 2072 df, AIC = –28.15), parameter estimates indicated significant genetic (A), shared environmental (C), and nonshared environmental (E) influences on child control (A = 0.35, C = 0.23, E = 0.41) and maternal control (A = 0.25, C = 0.31, E = 0.45). As noted, the presence of genetic influences on the latter indicates that maternal control behaviors are more similar if she has MZ twins than if she has DZ twins, suggesting that mothers are responding to the genetically influenced behaviors of their children. The genetic and environmental correlations between child and maternal control were also examined. The genetic and shared environmental correlations were estimated at –.94 and –1.0, respectively (negatively signed, since high levels of control by one member of the dyad elicit submissive behaviors in the other). Such results indicate that the genetic and shared environmental influences on child control overlap almost completely (in the case of genetic influences) and entirely (in the case of shared environmental influences) with those on maternal control. The nonshared environmental correlation was estimated at –.68, suggesting that most, but not all, nonshared environmental effects on child control also influenced maternal control.
Table 3. Genetic and environmental etiology of twin and maternal control and warmth and their overlap

Note: A, genetic variance; C, shared environmental variance; E, nonshared environmental variance. The 95% confidence intervals are presented in parentheses. Correlations whose confidence intervals overlap with zero are not statistically different from zero.
*p < .05 indicates that the correlation is statistically significant.
Parameter estimates for warmth were quite different (baseline model: –2LL = 5264.25 on 2056 df; ACE model: –2LL = 5473.39 on 2073 df, AIC = 141.14) and indicated that child warmth was primarily a function of child-specific environmental influences along with measurement error (A = 0.12, C = 0.00, E = 0.88), whereas maternal warmth was a function of shared and nonshared environmental influence (A = 0.02, C = 0.32, E = 0.66). The nonshared environmental correlation was estimated at .63, suggesting nonshared environmental influences on child warmth overlap with those on maternal warmth. Genetic and shared environmental correlations were not significant (to be expected, given the nonsignificant A and C parameter estimates).
Multivariate models
Consistent with the bivariate analyses, parameter estimates from the multivariate minute-by-minute analyses indicated significant genetic, shared environmental, and nonshared environmental influences on child control (average: A = 0.28, C = 0.15, E = 0.54) and maternal control (average: A = 0.25, C = 0.17, E = 0.56) during each minute of the interaction. Genetic and environmental correlations for control are presented in Table 4. As seen there, genetic and shared environmental influences on child control were highly stable across consecutive minutes (both rs = .99). Nonshared environmental influences on child control were also relatively stable (average r = .74). Genetic, shared, and nonshared environmental influences on maternal control demonstrated nearly identical patterns of stability (average r = .98, .98, and .72, respectively). Genetic and environmental influences on child control within a given interval also overlapped quite a bit with those of maternal control in that same interval (rA = –.94, rC = –.96, and rE = –.70). Genetic and shared environmental influences on child and maternal control were identical (if in opposing directions) within a given minute. However, these cross-sectional links between maternal and child control appeared to more or less persist across consecutive minutes. In particular, cross-time genetic correlations approached unity (r = –.92 and –.94 for child predicting mom and mom predicting child, respectively). Cross-time shared environmental overlap over time was also large (r = –.96 and –.97). Nonshared environmental overlap over time was more moderate (r = –.49 and –.51).
Table 4. Genetic and environmental correlations for control over 1-min time intervals

Note: rA, genetic correlation; rC, shared environmental correlation; rE, nonshared environmental correlation.
*p < .05.
The multivariate results for warmth were also consistent with the overall bivariate results in that child warmth was largely influenced by the nonshared environment (average A = 0.06, C = 0.01, E = 0.92), whereas maternal warmth was both shared and nonshared environmental in origin (average A = 0.05, C = 0.23, E = 0.72). Genetic and environmental correlations for warmth are presented in Table 5. As seen there, genetic and nonshared environmental influences on child warmth were highly stable across minutes (average r = .99 and .84, respectively), although the genetic correlations were not significant. For maternal warmth, genetic, shared, and nonshared environmental influences were also highly stable (average r = .87, .99, and .82, respectively), although, consistent with the nonsignificant contribution of A to maternal warmth, genetic correlations were not significant. Within each minute, genetic and environmental correlations between mothers and children were small to moderate in magnitude (rA = .27, ns; rC = .40, ns; rE = .57). The same pattern of results persisted to consecutive minutes (rA = .32 and .22, ns; rE = .47 and .51).
Table 5. Genetic and environmental correlations for warmth over 1-min time intervals

Note: rA, genetic correlation; rC, shared environmental correlation; rE, nonshared environmental correlation.
*p < .05.
Discussion
The goals of the current study were to confirm the presence of evocative rGE influences on the mother–child relationship in real-time interactions and to begin to identify at least some of the behaviors underlying evocative rGE by examining these processes as they unfold in real time. To do this, we made use of a recently developed momentary assessment of interpersonal behavior in a sample of mothers and twins. Our analyses revealed that interpersonal warmth was primarily influenced by the shared and the nonshared environment (although the former was important only in mothers). By contrast, there were significant genetic influences on both child and maternal interpersonal control. Because of our child-based twin design, any genetic influences on maternal behavior are a function of her response to her children's behavior. Genetic influences on maternal control thus confirm that evocative rGE can influence and shape parental control even during relatively brief interactions between mothers and children, a particularly striking finding considering that estimates of maternal control with her two children were coded by independent observers. Moreover, our results indicated that genetic influences on maternal control are a function of genetic influences on child control-oriented behaviors, both overall and in given minutes of the interaction. Such results represent a critical step forward in specifying the interpersonal processes through which evocative rGE impacts parenting behaviors.
Such findings replicate and extend previous studies examining the etiology of informant-reported parental control within genetically informed child-based designs (Ge et al., Reference Ge, Conger, Cadoret, Neiderhiser, Yates and Troughton1996; Harlaar et al., Reference Harlaar, Santtila, Björklund, Alanko, Jern and Varjonen2008; Herndon et al., Reference Herndon, McGue, Krueger and Iacono2005; O'Connor et al., Reference O'Connor, Deater-Deckard, Fulker, Rutter and Plomin1998; Plomin et al., Reference Plomin, Reiss, Hetherington and Howe1994). Our results are only partially consistent with other twin studies of the etiology of observed parenting. Neiderhiser et al. (Reference Neiderhiser, Reiss, Pedersen, Lichtenstein, Spotts and Hansson2004) reported a lack of genetic influence on observed maternal positivity and negativity (perhaps analogous to warmth) and O'Connor et al. (Reference O'Connor, Hetherington, Reiss and Plomin1995) also reported a lack of genetic influence on maternal warmth, both of which are consistent with our findings. However, neither study found evidence of genetic influences on observed maternal control/assertiveness behaviors. Our results are consistent with the work of Eley and colleagues (Reference Eley, Napolitano, Lau and Gregory2010), who found evidence for significant child-driven genetic effects on observed maternal control. Although it is unclear what may account for these differences across studies, the current findings are thought to provide additional confirmation of the importance of evocative rGE on control in the parent–child relationship.
Despite the growing body of evidence for evocative rGE effects on parenting, few studies have attempted to elucidate the specific processes through which a child's genetic makeup influences parenting behavior. In other words, the majority of evocative rGE studies of parenting simply identify the global presence of child-driven genetic effects on parenting. Although such findings are important, they do not reveal anything about the specific interpersonal processes driving evocative rGE (i.e., we are not able to trace these evocative rGE effects back to specific behaviors of the child). The current findings, by contrast, highlight a specific behavior driving evocative rGE in the parent–child relationship: interpersonal control. The extent to which children behave in a dominant or submissive manner during a given moment is partially a function of their genetic predispositions toward interpersonal control. By responding to these dominant or submissive behaviors in that same moment, mothers are indirectly responding to their children's genetic predisposition toward control. In short, the current study not only confirms the presence of evocative rGE in actual interpersonal processes (something that had not yet been done) but also illuminates at least one of the specific behaviors that underlie this evocative process.
There are several limitations to the current study. First, the joystick video coding method is a recently developed coding methodology and, moreover, one that had yet to be applied to parent–child dyads. The reliability of this method was supported within the current study and has been supported in two additional published studies and in one study under review (Markey, Lowmaster, & Eichler, Reference Markey, Lowmaster and Eichler2010; Sadler et al., Reference Sadler, Ethier, Gunn, Duong and Woody2009; Thomas et al., Reference Thomas, Hopwood, Ethier and Sadler2012). Although the reliability for warmth within the current study was adequate and consistent with other joystick studies and other observational coding schemes of maternal behavior (e.g., Neiderhiser et al., Reference Neiderhiser, Reiss, Pedersen, Lichtenstein, Spotts and Hansson2004), the reliability for warmth was not as high as for control. Therefore, differences between warmth and control within this study should be interpreted with caution. Another limitation is that ratings of warmth and control were obtained from short 8-min slices of behavior, and it could be argued that the behaviors observed during these interactions may not generalize to overall patterns of mother–child interaction. However, research has supported the utility and validity of inferences based upon short slices of behavior, such that ratings of personality based on observations lasting only a few minutes are significantly associated with reports by knowledgeable informants (Borkenau, Mauer, Riemann, Spinath, & Angleitner, Reference Borkenau, Mauer, Riemann, Spinath and Angleitner2004).
Second, the nature of the Etch-a-Sketch task required mothers and children to work together, which may have increased levels of cooperation and warmth. Consistent with this, there was less variance observed for warmth as compared to control. This possible restriction of range for warmth may have contributed to etiological differences between warmth and control observed here. However, the nature of the mother–child relationship often requires that mothers and children work together toward a shared goal, and thus a task that calls for affiliation may resemble many of the day-to-day interactions between mothers and children. In addition, the presence of large nonshared environmental influences on warmth (and particularly child warmth) may indicate the presence of error in these ratings. These findings are consistent with previous research suggesting that warmth behaviors are somewhat more strongly associated with idiosyncratic situational factors (as opposed to traits), at least as compared to control behaviors (Sadler & Woody, Reference Sadler and Woody2003). Therefore, the structured lab task situation may have elicited primarily situation-specific rather than trait-based warmth behaviors, particularly in the child.
Third, the results of this study only apply to typically developing children in middle childhood and their mothers. It is possible, even likely, that the etiology of interpersonal processes in the mother–child relationship is fluid across development and thus shifts with age. It may also differ in father–child dyads. In addition, these results do not specifically address the etiology of mother–child interactions in clinical populations. Research has implicated an important role of child resistance to control in the etiology of conduct problems (Dodge & Pettit, Reference Dodge and Pettit2003), and it is thus possible that child control/autonomy behaviors may not be associated with maternal relinquishing of control in cases where children express clinically significant conduct problems. Other studies suggest that lower levels of parental involvement and monitoring are associated with conduct problems (Wasserman, Miller, Pinner, & Jaramillo, Reference Wasserman, Miller, Pinner and Jaramillo1996), and thus mothers of conduct-disordered youth may respond with excessive relinquishing of control. Supportive evidence for this possibility can be found in common interventions for families of children with conduct problems, which often include helping parents to regain appropriate and consistent control within the parent–child relationship (McMahon, Forehand, & Foster, Reference McMahon, Forehand and Foster2005). Future research is necessary to further elucidate the ways in which parent and child interpersonal control may function in families of children with conduct problems. However, these results do shed light on normative mother–child interactions in an important period of child development during which the mother–child relationship plays a crucial role.
Despite these limitations, the current study has several important implications. First, our results suggest that the interpersonal framework is a useful and integrative way to conceptualize parent–child interactive behaviors. Currently, there is little consistency regarding the operationalization of the parent–child relationship within the parenting literature. Second, the IPC framework is grounded in a rich theoretical and empirical history and can function as a conceptual bridge between developmental models of the parent–child relationship (such as Baumrind's parenting model) and other theories within the broader literature. Third, the IPC is well suited for the identification of bidirectional processes in the parent–child relationship, taking into account the importance of both child- and parent-driven factors. Fourth, the IPC provides a common language that could serve to connect research on parent–child relationships with other key components of the child's family system and social networks (e.g., sibling relationships, marital relationships, friendships, and teacher–student interpersonal processes).
Our results also highlight the utility of momentary assessments of interpersonal behavior for behavioral genetic research. Averaged momentary data estimates provide an equal weighting of all moments across an interaction and thus should allow for better detection of the etiology of interpersonal behavior as compared to traditional global coding schemes. However, analysis of the actual half-second by half-second data provides another, perhaps even more promising, avenue for future investigations of evocative rGE. Visual inspection of these data suggests the presence of cycling over time. This is consistent with research suggesting that familial relationships function as systems that are characterized by attractor states (such that behaviors return to equilibrium) and reciprocal influences, and may thus be best measured using a dynamical systems analytic approach (van Geert & Lichtwarck-Aschoff, Reference van Geert and Lichtwarck-Aschoff2005). Coupled dynamic models may thus be an especially interesting direction for future research. Within coupled dynamic models, the cyclical behavioral patterns of one individual in an interaction can be modeled as they are influenced by the patterns of the other. This enables an examination of process within an interaction, including direction and strength of effects between members of a dyad (Boker & Laurenceau, Reference Boker, Laurenceau, Little, Bovaird and Card2007). The development of such models within a behavioral genetic framework is ongoing and would allow for estimates of genetic and environmental influence on dynamic process aspects of the parent–child relationship.
The current study also has important implications for our understanding of the etiology of interpersonal behavior between mothers and children, a key consideration given that both parental warmth and control are associated with the development of psychopathology (e.g., low parental warmth has been associated with increased adolescent antisocial behavior; Feinberg, Button, Neiderhiser, Reiss, & Hetherington, Reference Feinberg, Button, Neiderhiser, Reiss and Hetherington2007), whereas parental control is associated with childhood anxiety (van der Bruggen, Stams, & Bogels, Reference van der Bruggen, Stams and Bogels2008) and conduct problems (Dishion, Nelson, & Bullock, Reference Dishion, Nelson and Bullock2004). In particular, our findings suggest the presence of minimal genetic influences on observed warmth in the mother–child relationship. The warmth displayed by children toward their mothers was found to be primarily nonshared in origin (i.e., idiosyncratic to each twin), and maternal warmth was influenced by both the shared and the nonshared environment (32% and 66%, respectively). Nonshared environmental influences on child and maternal warmth may include idiosyncratic effects of the task and the assessment environment as well as the unique (nongenetically influenced) characteristics of children. The significant nonshared environmental correlations between child and maternal behavior both within and across minutes also suggest that some of the same unique etiological factors influence behavior in both children and mothers (and could include the influence of idiosyncratic events earlier in the day or more stable nonshared sources of variance such as one twin's physical illness). For their part, shared environmental influences on maternal warmth may include such factors as stable maternal personality characteristics as well as social and culture factors that act to promote more or less consistent levels of maternal warmth. This interpretation is bolstered by shared environmental influences on maternal behavior in child-based twin designs including characteristics of the mother, because these should be more or less constant across children. Moreover, mean levels of maternal warmth were higher than child warmth, collectively suggesting that mothers are perhaps responding to societal or personal expectations that mothers exhibit warmth toward their children (irrespective of how the children respond) or that observers are more likely to perceive maternal figures as warm. In other words, maternal warmth may be particularly influenced by broader societal factors or individual personological factors, at least as compared to maternal control.
By contrast, genetic influences appear to play an important role in even thin slices of mother and child control-oriented behaviors. Maternal control appears to be driven in large part by the individual genetically influenced characteristics of her twins. Such findings not only serve to robustly confirm the presence of evocative rGE in the mother–child relationship but also serve to illuminate at least one of the specific interpersonal behavior(s) that underlie this evocative process. In particular, our findings suggest that the give-and-take balance of interpersonal control between a mother and her child, even in a given moment, is at least partially a function of that child's genetic predisposition toward control. When children expressed autonomy, mothers generally responded by relinquishing some control (although notably, mothers were higher in control than children on average and thus were likely to retain some control even while granting some autonomy). Conversely, when mothers took a dominant position, children generally responded by following the mother's lead. Moreover, this process unfolded during real-time interactions, suggesting that genetic influences on control operate at the level of day-by-day and even moment-by-moment interactions between mothers and children. Such findings represent an important first step toward the identification of processes by which evocative rGE operates in the parent–child relationship and, in this way, begin to bridge the gap between research and practice by elucidating how child-driven genetic factors impact parenting behaviors. Our results suggest that interventions promoting adaptive maternal control should target dyadic relationship patterns and the ways in which mothers respond to their children's unique behaviors in everyday interactions. Future research should continue to examine the mother–child relationship as a bidirectional and dynamic process, influenced by both genetic and environmental factors, and should seek to elucidate the specific child and parent behaviors that underlie evocative rGE effects on parenting.