


Introduction
The eastern larch beetle, Dendroctonus simplex LeConte (Coleoptera: Curculionidae: Scolytinae), is a monophagous bark beetle that attacks and colonises the phloem of its host tree, eastern larch (tamarack) Larix laricina (Du Roi) Koch (Wood Reference Wood1982). The range of the eastern larch beetle is extensive and closely matches that of tamarack, found throughout the Canadian boreal forest as well as the northeastern and northcentral United States of America and Alaska, United Sates of America (Seybold et al. Reference Seybold, Albers and Katovich2002). Historically, the eastern larch beetle has been considered a non-aggressive bark beetle and thus the subject of fewer studies than other tree-killing Dendroctonus Erichson species (Langor and Raske Reference Langor and Raske1989a, Reference Langor and Raske1989b; Hopkins Reference Hopkins1909). Unthrifty tamaracks stressed from flooding, wind-throw, defoliation, drought, pathogen infection, or other predisposing agents are often colonised (Hopkins Reference Hopkins1909). Under favourable conditions, the insect will attack and kill relatively healthy trees, with such infestations typically being of short duration (e.g., two to three years) (Langor and Raske Reference Langor and Raske1988). In recent decades, however, the eastern larch beetle has exerted increasing landscape-scale mortality, affecting millions of hectares of tamarack forest in separate outbreaks in Alaska and the Canadian Maritimes region (Werner Reference Werner1986; Langor and Raske Reference Langor and Raske1989b).
Reproductively mature adult beetles typically emerge in early spring from tamarack material colonised the previous year, disperse, and locate new host trees or material. Pheromone-mediated mass-aggregation (Prendergast Reference Prendergast1991) facilitates mate procurement and enables beetles to overwhelm host defenses, killing the tree and facilitating successful brood production. Eggs of the first larval brood are laid shortly thereafter. Parent beetles often re-emerge from colonised tamaracks and establish a second, and sometimes a third, sibling or “sister” brood in separate host material. Larvae develop rapidly, and by early summer the adult beetles of the first brood begin to emerge from pupal chambers. Depending on the year and sibling cohort, 20–80% of brood adults will exit the host throughout the summer and fall, drop to the lower bole of the natal tree, re-enter the bark, and overwinter in non-reproductive galleries constructed in the base of the tree (Simpson Reference Simpson1929; Werner Reference Werner1986). Remaining brood adults overwinter in the pupal chambers (Werner Reference Werner1986; Langor and Raske Reference Langor and Raske1987b; Seybold et al. Reference Seybold, Albers and Katovich2002). The brood adult is the predominant overwintering life-stage (Langor and Raske Reference Langor and Raske1987a), although both adults and larvae are extremely cold hardy, surviving temperatures as low as −42 °C and −49 °C, respectively (Venette and Walter Reference Venette and Walter2008).
Previous studies report an absence of reproductive efforts by emerging brood adults in the summer and fall such that new host trees are not attacked even though sufficient time often remains in a season for the development of a new generation of beetles (Swaine Reference Swaine1911; Langor and Raske Reference Langor and Raske1987b). Because only one reproductive generation of eastern larch beetles per year has ever been observed historically (Hopkins Reference Hopkins1909; Prebble Reference Prebble1933; Dodge Reference Dodge1938; Werner Reference Werner1986; Langor and Raske Reference Langor and Raske1987b), evidence suggests that the insect possesses an obligate reproductive diapause (Langor and Raske Reference Langor and Raske1987b). An obligate overwintering reproductive diapause reportedly occurs in the Douglas-fir beetle (Dendroctonus pseudotsugae Hopkins (Coleoptera: Curculionidae)) (Furniss Reference Furniss1976), the closest relative of the eastern larch beetle (Wood Reference Wood1982) as well as in other species of bark beetles such as Dendroctonus rufipennis (Kirby) (Hansen et al. Reference Hansen, Bentz, Powell, Gray and Vandygriff2011). However, no published studies have tested whether the eastern larch beetle can reproduce without an overwintering period (i.e., lack of reproductive diapause).
Since 2000, an ongoing outbreak of eastern larch beetles in Minnesota, United States of America, has killed over 75 000 ha of tamarack in the state (Minnesota Department of Natural Resources 2012). Eastern larch beetle activity is also increasing throughout other areas of the Great Lakes region including Wisconsin and Michigan, United States of America, as well as in Manitoba and Ontario, Canada (Ontario Ministry of Natural Resources 2012; Michigan Department of Natural Resources 2013; Wisconsin Department of Natural Resources 2013; Manitoba Conservation Forestry Branch 2014). In Minnesota, the cause of the outbreak is not known, as beetle activity is not associated with any known predisposing conditions (Albers Reference Albers2010; Minnesota Department of Natural Resources 2012). As part of a larger study to elucidate the seasonal phenology and reproductive capacity of the insect, a series of three experiments were undertaken with the following objectives: (i) to determine if eastern larch beetle progeny (F1) from the field could reproduce successfully without an overwintering period, (ii) to determine whether their offspring (F2) were reproductively viable without a subsequent overwintering period, and (iii) to explore potential differences in the reproductive abilities of brood adults (F1) that emerge from the host before winter versus those brood adults that remain in the pupal chambers to overwinter. These studies seek to determine whether a second generation of eastern larch beetles in field settings is hypothetically possible in a single year due to reproduction by beetle progeny before an overwintering period and if such reproduction could be a factor contributing to the current beetle outbreak.
Methods
Experiment 1: Are F1 eastern larch beetle progeny that do not emerge from natal hosts in the fall capable of reproduction without an overwintering period?
To obtain beetles, three infested tamaracks containing the first spring brood of 2011 were harvested on 19 June 2011 from the Red Lake Wildlife Management Area (RLWMA), Lake of the Woods, Minnesota, United States of America (48.5725°N, 95.0616°W). Parent beetles (F0) were re-emerging from the infested trees at the time of cutting and developing offspring were in the egg, first instar, and second instar life-stages. The infested logs were brought to the University of Minnesota, St. Paul, Minnesota. The cut ends of the logs were sealed with molten paraffin wax before placement in rearing tubes. Brood (F1) development continued at 23 °C and a 24:0 light:dark photoperiod. Collecting jars were attached to the rearing tubes to capture emerging progeny.
Because we were interested only in the reproductive ability of beetle progeny (F1) that did not emerge from the infested material, all re-emergent parent (F0) eastern larch beetles (Simpson Reference Simpson1929; Langor and Raske Reference Langor and Raske1987a) and emergent brood (F1) adults were discarded. The infested logs were peeled on 20 August 2011 after a period of 10 days without beetle emergence. At that time, fully sclerotised (i.e., black and maroon) or dark brown brood adults were extracted directly from pupal chambers. Henceforth, such beetles are referred to as “manually extracted” while those that emerged into the collecting jars are referred to as “naturally emergent”. These progeny were separated by sex and natal host, and stored at 4 °C on moist paper towel for 48 hours until use.
To obtain breeding material for these progeny, three green tamaracks were harvested on 19 August 2011 from the RLWMA (48.6268°N, 94.9528°W). The diameter at breast height (DBH: 1.4 m) of the trees were 24.1, 22.4, and 22.7 cm. Eighteen bolts, six from each green tamarack, each 50 cm in length were prepared 20 August 2011. Cut ends of the bolts were sealed with wax.
The female progeny (F1) were removed from 4 °C storage and allowed two hours to warm to room temperature (~23 °C) on 22 August 2011. Ninety vigorous, two-day-old females, 30 from each infested tamarack, were randomly selected and pooled. Five females were introduced to each bolt using starter holes 5 mm wide, drilled into the phloem, and spaced evenly around the bolt circumference, 5 cm from one end. One female was sealed in each hole using aluminum screening. Females were checked after two hours and four females that were not boring into the bolts (i.e., female still present in starter hole and boring frass absent) were replaced. Bolts were placed horizontally in rearing tubes to allow females to excavate the egg gallery.
The next day, the male progeny (F1) were removed from 4 °C storage and likewise warmed for two hours at room temperature. One hundred and eight vigorous, three-day-old males, 36 from each infested tree, were randomly selected and pooled. The aluminum screening was removed from the female entrance holes. Six males were placed on the bark of each bolt and allowed to locate the female egg galleries. Males were checked 6, 12, and 24 hours after introduction. Males that had fallen from the bolts were placed back on to the bark surface. No beetles were found at the 24 hours check indicating that all had joined the female colonisers in the log. Bolts were stored horizontally at 23 °C.
The bolts were monitored for emerging offspring (F2) every two days, beginning 21 days after male introduction. The first emergent offspring was observed on 19 October 2011. Emergent offspring were counted and separated by date, and bolt. The bolts were peeled 4 November 2011 to collect adult offspring (F2) from the pupal chambers. Offspring were counted and separated by bolt and sex and stored at 4 °C for use in a subsequent breeding experiment (see below). Egg galleries of eastern larch beetles tend to meander and quickly intersect one another to form egg gallery networks (Langor and Raske Reference Langor and Raske1987b) making it difficult to attribute specific reproductive success to each female when multiple females are present. Thus, the reproductive success of introduced progeny was expressed as the number of offspring per female by dividing the total number of offspring produced in a bolt by the number of introduced females. This method was also used for experiments 2 and 3 (see below).
Experiment 2: Is the F2 generation of non-wintered eastern larch beetle progeny reproductively viable?
Non-emergent F2 progeny of the non-emergent F1 beetles (see experiment 1) were used to determine if the F2 brood were also reproductively viable without an over-wintering period. Three healthy tamaracks (DBHs: 20.5, 18.9, and 18.6 cm) harvested from the RLWMA on 8 October 2011, and stored as previous, were sectioned into five bolts, each 50 cm in length, on 5 November 2011. Two bolts were cut from each of two tamaracks, and a single bolt was sourced from the third tamarack. The midsection diameters of the bolts were 15.1, 13.9, 14.1, 13.8, and 16.1 cm.
These bolts were infested with F2 female beetles on 7 November 2011. Females were removed from cold storage and warmed for two hours at 23 °C. Fifty vigorous, three-day-old females, five from each of 10 natal bolts of experiment 1, were selected randomly and pooled. Ten females were introduced to each of the new bolts by placing the females on the bark and allowing them to select their own entry points. On 9 November 2011, 60 vigorous, five-day-old F2 males, six from each of the 10 natal bolts of experiment 1, were randomly selected and pooled. Twelve males were placed on the bark of each breeding bolt and allowed to locate the female entrance sites. All beetles were checked at 6, 12, and 24 hours post-introduction, again with fallen beetles being placed back on the bark. No beetles were found at the 24-hour check, indicating that all beetles had entered the bolts. Bolts were kept at 23 °C.
The bolts were checked for offspring (F3) emergence every two days beginning 21 days after male beetle introduction. Emergent offspring were first recorded 27 December 2011. Emergent offspring were collected and separated by emergence date, and bolt. Bolts were peeled on 21 February 2012 to collect offspring from pupal chambers under the bark. Reproductive capacity was expressed as the number of offspring produced per parent female.
Experiment 3: Does the reproductive ability of non-wintered F1 beetles that emerge naturally versus reside in pupal chambers differ?
As stated, and experienced in experiments 1 and 2, a proportion of insects in the field and laboratory choose to reside in place in their pupal chambers putatively for overwintering rather than emerging from under the bark (Werner Reference Werner1986; Langor and Raske Reference Langor and Raske1987a; Seybold et al. Reference Seybold, Albers and Katovich2002). We examined whether the reproductive capabilities of the naturally emergent and manually extracted groups of F1 beetles differed prior to going through the winter. To obtain non-overwintered eastern larch beetle brood adults (F1), two tamaracks infested with the first spring brood of 2012 were cut from the RLWMA (48.6266°N, 94.9537°W) on 28 July 2012. Phenology data of eastern larch beetle activities collected for a separate study (F.R.M. and B.H.A., personal observations) indicated that parent beetles (F0) had completed re-emergence from spring brood trees by 6 July 2012, and brood adults had been emerging since 13 July 2012. The infested logs were brought to the University of Minnesota, and were prepared and placed in emergence tubes as previously described. Brood adults (F1) continued to emerge and were collected every one to two days, separated by date, natal host, and sex, and stored at 4 °C on moist paper towel. After seven days without emergence, the infested logs were peeled and remaining brood adults were extracted by hand from pupal chambers on 15 August 2012.
Three healthy tamaracks (DBHs: 18.1, 18.0, and 18.8 cm) were cut from the RLWMA (48.6269°N, 94.9527°W) on 1 July 2012. Again, ends were sealed with paraffin wax, and the logs were stored at 4 °C. On 6 August 2012, eight bolts, 30 cm in length, were cut from the green logs. Four bolts were prepared for the naturally emerging insects, and four for those insects that were manually extracted from logs upon peeling. For each beetle group, two bolts were made from one tamarack and single bolts were made from each of the other two tamaracks. The ends of the bolts were sealed with paraffin wax before storage at 4 °C until needed. The diameters of the four bolts were 12.8, 13.3, 13.2, and 14.6 cm and 13.3, 12.5, 14.4, and 14.5 cm for the naturally emergent and manually extracted brood adults, respectively.
On 6 August 2012, naturally emergent female beetles (F1) were removed from cold storage and warmed for two hours at 23 °C. Forty vigorous females, 20 from each of the two infested tamaracks were randomly selected and pooled. Ten females were placed on the bark of each bolt. Again, the females selected their own entry sites. On 8 August 2012, naturally emergent male beetles (F1) were removed from cold storage and warmed for two hours at 23 °C. Forty-eight vigorous males, 24 from each of the two infested tamaracks were randomly selected and pooled. Twelve male beetles were placed on the bark of each bolt, and allowed to locate female entry sites. All beetles were ⩽6 days old. The methods for introducing manually extracted beetles to the parallel set of bolts was identical to the naturally emergent beetles, and occurred on 19 August 2012 (females) and 21 August 2012 (males).
All bolts were monitored for emergent offspring (F2) every two days beginning 21 days after beetle introduction. Emergent offspring were first observed 23 September 2012 and 8 October 2012 from bolts infested with naturally emergent and manually extracted beetles, respectively. Emergent offspring were separated by emergence date, and bolt. Bolts were peeled 30 days after the initial emergence of offspring, when the emergence had dropped to near zero, to collect and count any remaining live offspring.
Lengths of parental galleries were recorded for each bolt, as well as the types of galleries. Different types of gallery architecture have been noted in other studies of eastern larch beetles (Simpson Reference Simpson1929; Werner Reference Werner1986), but their functions remain unclear. For our purpose, we characterised the galleries into three types (Table 1).
Table 1 Characteristics of the galleries excavated by eastern larch beetle brood adults when introduced to tamarack bolts.

* Names given for gallery types are unofficial terms used by the authors for convenience.
≅ Name based on field observations of similar galleries excavated in the basal bark of natal trees by emergent brood adults and in which beetles overwinter.
Statistical analyses
Analysis of variance (ANOVA) was used for five analyses: (i) comparing offspring production per female between the F1 and F2 generations of non-wintered brood adults (experiment 1 versus experiment 2), (ii) comparing offspring production per female between the naturally emergent and manually extracted (F1) brood adults (experiment 3), (iii) comparing offspring production per cm of egg gallery between the naturally emergent and manually extracted brood adults (experiment 3), (iv) comparing mean total length per bolt of each gallery type for naturally emergent brood adults (experiment 3), and (v) comparing mean total length per bolt of each gallery type for manually extracted brood adults (experiment 3). Per capita reproduction was calculated by dividing the number of introduced females into the number of offspring produced per bolt. The bolt was the unit of replication for analyses of offspring production per female and total length of each gallery type per bolt. All analyses were performed using R v2.14.1 (R Development Core Team 2014). Variables were transformed as necessary (e.g., √y) to fulfill model assumptions of homoscedasticity and normality of errors.
Results
In both experiments 1 and 2, we observed successful reproduction by the F1 and F2 generations of manually extracted eastern larch beetle progeny that had not been exposed to an overwintering period. Non-wintered F1 brood from the first spring brood of 2011 (experiment 1) produced 6.1±2.6 (mean±SE) offspring per female, and non-wintered F2 brood produced 6.7±3.6 offspring per female (experiment 2) (Fig. 1). These reproductive outputs were not significantly different (ANOVA, F(1,21)=0.17, P=0.68). Thus, it appears that some eastern larch beetles are able to reproduce with some success without an obligate overwintering reproductive diapause. In experiment 2, in which all source beetles were raised in a uniform laboratory thermal environment, we found that 94.9% of the insects emerged naturally. The remaining 5.1% required extraction from their pupal chambers by hand.
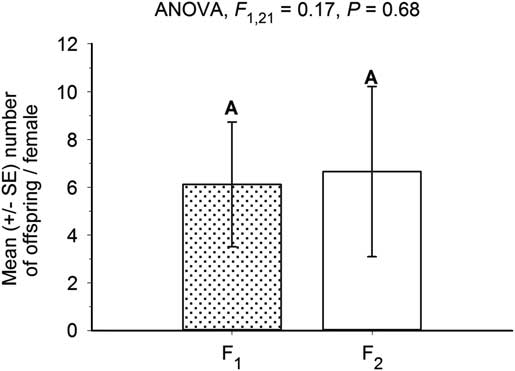
Fig. 1 Number of offspring (mean±SE) produced per non-wintered, manually extracted F1 female (experiment 1) and F2 female (experiment 2). Parent beetles used in experiment 1 were progeny from the first spring larval brood of 2011. Parent beetles used in experiment 2 were offspring from experiment 1.
When we compared insects that emerged naturally versus those extracted manually from pupal chambers in experiment 3, we found successful reproduction by both F1 groups. Naturally emerged beetles produced 43.0±2.7 (mean±SE) offspring per female, whereas the manually extracted beetles produced significantly fewer offspring at 7.4±3.0 per female (ANOVA, F(1,6)=24.7, P=0.0025) (Fig. 2A). Differences in bolt diameter were not responsible for this trend, as the mean±standard error (SE) bolt diameters of 13.5±0.4 and 13.7±0.5 cm for the naturally emergent and manually extracted beetles, respectively, were not significantly different (ANOVA, F(1,6)=0.11, P=0.77).
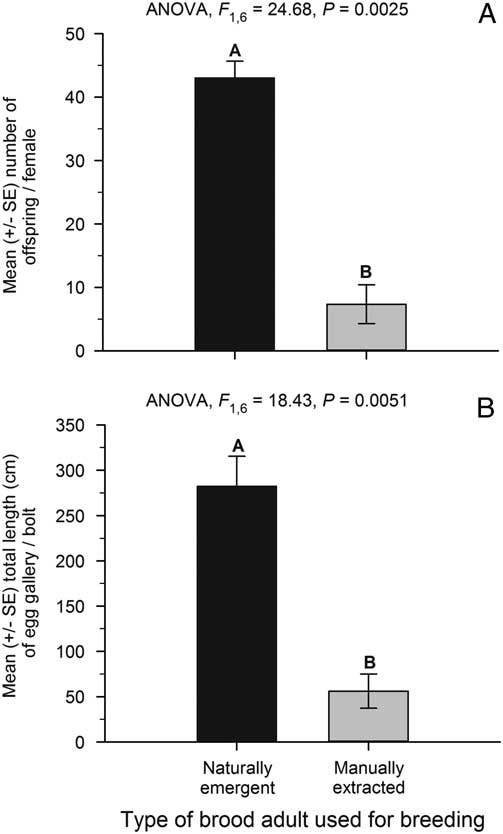
Fig. 2 (A) Number of offspring (mean±SE) produced per female for naturally emerged and manually extracted beetles in experiment 3. (B) Total length (cm) of egg gallery (mean±SE) excavated per bolt by naturally emergent and manually extracted parent beetles. Beetles from both groups had not experienced an overwintering period before the reproductive trials. n=10 females per bolt, n=4 bolts for each of the naturally emergent and manually extracted beetle groups.
Despite the pronounced difference in total reproduction per female, offspring production per cm of egg gallery for naturally emergent and manually extracted F1 brood adults was similar (ANOVA, F(1,5)=0.55, P=0.49). The number of offspring per cm of egg gallery averaged 1.6±0.1 SE and 1.3±0.3 for naturally emerged and manually extracted beetles, respectively. The difference in reproductive success was due to longer egg galleries excavated in the bolts infested with naturally emerged F1 beetle parents (Fig. 2B; ANOVA, F(1,6)=18.43, P=0.0051). The total length of egg gallery in these bolts was more than five-fold those of the bolts infested with F1 beetle parents had been manually extracted directly from pupal chambers before introduction to the bolts. Naturally emergent beetles excavated 282.0±33.3 cm (mean±SE) of egg gallery per bolt while the manually extracted beetles only excavated 56.0±18.9 cm of egg gallery per bolt.
The types of galleries excavated differed between the naturally emergent and manually extracted F1 beetles. Naturally emergent beetles excavated only egg galleries (Fig. 3). In contrast, manually extracted beetles excavated egg, pseudo-egg, and hibernal galleries in equal proportions (Fig. 3) as the total length of each gallery type per bolt did not vary (ANOVA, F(2,9)=0.95, P=0.42). The mean length of individual egg galleries of naturally emergent beetles was 94.1±18.0 cm (n=12). In contrast, the egg galleries of the manually extracted beetles averaged less than half of that amount, only 37.4±11.2 cm in length (n=6). Pseudo-egg galleries (n=6) and hibernal galleries (n=14) averaged 12.7±2.6 and 5.3±1.0 cm in length, respectively.
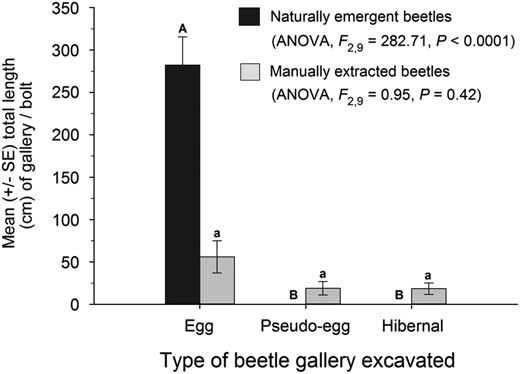
Fig. 3 Mean total length of gallery type per bolt for naturally emergent and manually extracted beetles. Different letters indicate significant differences in total gallery length per bolt for naturally emergent beetles (upper case letters) and manually extracted beetles (lower case letters).
Discussion
While previous studies are consistent with the understanding that a univoltine life cycle for eastern larch beetles is governed by an obligate adult diapause (Hopkins Reference Hopkins1909; Swaine Reference Swaine1911; Simpson Reference Simpson1929; Dodge Reference Dodge1938; Werner Reference Werner1986; Langor and Raske Reference Langor and Raske1987b) similar to the Douglas-fir beetle, the present work suggests that there may be genetic variability in adult diapause capacity within a population. In experiment 2, for example, where all parent beetles were subjected to the same laboratory thermal regime, 95% of the brood were naturally emergent whereas 5% appeared to be in a diapause state. The number of beetles in a putative diapause state may have been higher if the naturally emerging beeltes were actually emerging to seek overwintering sites at the base of a tree (Langor and Raske Reference Langor and Raske1987a). Flight musculature appeared robust, however, suggesting they were reproductively mature and ready to seek new hosts (see below). If diapause capacity is a plastic trait, this minority proportion of brood may have entered diapause with a lower accumulation of thermal units. Such genetic variation in diapuase capacity has also recently been described within the pine weevil Hylobius abietis (Linnaeus) (Coleoptera: Curculionidae). Other than a facultative prepupal diapause, it had been thought that pine weevils needed to overwinter before becoming reproductively viable (Clark Reference Clark1975). In field settings, however, a proporation of adults can mature eggs without overwintering (Tan et al. Reference Tan, Wainhouse, Day and Morgan2010; Wainhouse et al. Reference Wainhouse, Inward and Morgan2014).
Propensity to enter a diapause state is likely triggered by an environmental cue such as temperature during a critically sensitive life stage (Tauber et al. Reference Tauber, Tauber and Masaki1986). The spruce beetle, D. rufipennis (Kirby) (Coleoptera: Curculionidae), for example, exhibits a facultative prepupal diapause and what is thought to be an obligate adult diapause (Safranyik et al. Reference Safranyik, Simmons and Barclay1990). When spruce beetle pre-pupae experience threshold cool temperatures of appropriate duration, a facultative diapause arrests development until the following season. If favourably warm environmental temperatures prevail, then spruce beetles proceed to the adult overwintering diapause stage uninterrupted (Hansen et al. Reference Hansen, Bentz and Turner2001, Reference Hansen, Bentz, Powell, Gray and Vandygriff2011). Differences in environmental conditions (i.e., climate) may partially explain why eastern larch beetle brood adults from previous studies did not attempt immediate reproduction upon emergence (Hopkins Reference Hopkins1909; Swaine Reference Swaine1911; Werner Reference Werner1986; Langor and Raske Reference Langor and Raske1987b), whereas a portion of the brood adults in this study were reproductively viable.
We note two lines of evidence for adult facultative versus obligate diapause states. First, one study reports the existence of eastern larch beetle progeny emerging from host trees in the field in summer with underdeveloped flight muscles (Langor and Raske Reference Langor and Raske1987a), a characteristic of adult bark beetles and weevils in a state of diapause (Ryan Reference Ryan1959; Danks Reference Danks1987; Nordenhem Reference Nordenhem1989; Tan et al. Reference Tan, Wainhouse, Day and Morgan2010). However, emergent progeny collected from rearing jars in the present experiments would often fly rapidly towards ceiling lights, indicating the functionality of the flight muscles. Second, differences in gallery construction behaviour could reflect differences in reproductive maturity. Bark beetles in a state of diapause exhibit barren ovarioles or underdeveloped seminal vesicles (Ryan Reference Ryan1959; Langor and Raske Reference Langor and Raske1987a). In the present work, naturally emergent beetles only excavated egg (i.e., reproductive) galleries, while manually extracted beetles placed equal effort into excavating egg, as well as non-reproductive pseudo-egg, and hibernal galleries (Fig. 3). The construction of feeding and hibernal galleries by non-reproductive brood adults has also been noted in other studies (Simpson Reference Simpson1929; Werner Reference Werner1986). Over-wintering galleries constructed by brood adults in tamaracks in the field are similar to the so-called hibernal galleries observed in this study (F.R.M., personal observation), and may reflect adults in a state of diapause.
This study suggests that eastern larch beetles may be capable of bivoltinism under appropriate conditions that facilitate reproductive maturity in brood adults. Both spring-emergent and summer-emergent reproductive generations of beetles may be possible in a single season when sufficient heat units are available to allow the insects to complete development (McKee and Aukema Reference McKee and Aukema2014). This may be particularly true if climate warming creates a longer window suitable for beetle development due to a warmer and earlier spring as well as warmer and later fall season. A second generation of eastern larch beetles in one summer could contribute substantially to beetle population growth, increase the frequency and severity of beetle outbreaks, and resultant tamarack mortality. The mean number of progeny produced per F1 brood adult female that had emerged naturally from the bolts in the present study (Fig. 2A; 43±2.7 SE insects) is similar to that of spring-emergent beetles in three separate, and independent, laboratory experiments (44.5±5.5 (McKee and Aukema Reference McKee and Aukema2014), 40.8±4.9, and 58±4.8; F.R.M. and B.H.A., personal observations).
The information found in this study challenges traditional assumptions regarding the potential for eastern larch beetles to become a significant forest pest and disturbance agent if it is considered as a strictly univoltine insect colonising moribund trees. Detailed studies to elucidate the mechanisms that govern variation within the reproductive maturation and reproductive capacity of eastern larch beetles are warranted, such as work examining respiratory or developmental rates to quantify potential diapause events in immature stages (e.g., Hansen et al. Reference Hansen, Bentz, Powell, Gray and Vandygriff2011; McKee and Aukema Reference McKee and Aukema2014). Such studies will be particularly important to predict future population dynamics of this insect and its ability to inflict increased mortality to tamarack forests under scenarios of a changing climate.
Acknowledgements
The authors thank Becky Lein and staff (Minnesota Department of Natural Resources – Forestry) for providing access to research material, Gretchen Mehmel (Minnesota Department of Natural Resources – Wildlife), and staff of the Red Lake Wildlife Management Area for providing equipment and accommodations; Jana Albers, Michael Albers, and Valerie Cervenka (Minnesota Department of Natural Resources – Forestry) for logistical support; and Erica Nystrom-Santacruz, and Michelle Cummings (University of Minnesota – Twin Cities) for technical assistance. Funding was provided by the United States Forest Service Evaluation Monitoring Program NC-EM-B-12-01 and McKnight Land-Grant Professorship funds to B.H.A. They thank the two anonymous reviewers who provided helpful comments on an earlier version of this manuscript.




