


Introduction
Amauromyza Hendel was originally erected as a subgenus of Dizygomyza Hendel to accommodate a few black European species with black halteres (Hendel Reference Hendel1931a). The subgenus was later raised to full generic rank by Nowakowski (Reference Nowakowski1962) and this decision was followed by subsequent authors who confirmed the homogeneity of the group based on male genitalic characters (Spencer Reference Spencer1971b). Amauromyza has now a worldwide distribution and consists of 60 species (Table 1). Externally, this genus is highly variable, with colour varying from completely black with partially or completely black halteres (similar to some species of Nemorimyza Frey) to species that are partially to largely yellow with white or yellow halteres (similar to species of Calycomyza Hendel or Phytoliriomyza Hendel). The orbital setulae are usually present and either reclinate or inclinate, but sometimes partially proclinate (as in Amauromyza scleritica Spencer) or absent (as in Amauromyza pleuralis (Malloch)). The costa usually extends to vein M1 + 2 but only to R4 + 5 in two European species: Amauromyza fraxini (Beiger) (previously in Aulagromyza Enderlein) and Amauromyza madrilena Spencer. The presutural dorsocentral bristle is usually present. Correct identification to genus is possible only by using a combination of external (see “Diagnosis”) and genitalic characters (mainly the broadly rounded hypandrium and the large bowl-shaped base of the ejaculatory apodeme). Identification of Canadian species of Amauromyza is also difficult because the most recent key (Spencer Reference Spencer1969b) contained only three species: A. abnormalis (Malloch), A. karli (Hendel), and A. subinfumata (Malloch). Two other species, A. flavifrons (Meigen) and A. pleuralis were included in the genus Trilobomyza Hendel, later treated as a subgenus of Amauromyza (Spencer Reference Spencer1971b) and synonymised with subgenus Cephalomyza Hendel by Spencer and Steyskal (Reference Spencer and Steyskal1986). New species descriptions, nomenclatural changes, and a new concept of the genus supported by genitalic characters have raised the number of Canadian Amauromyza species to 11: A. abnormalis, A. angulicornis Zlobin, A. flavifrons, A. karli, A. luteiceps (Hendel), A. nevadensis Spencer, A. riparia Sehgal, A. scleritica Spencer, A. shepherdiae Sehgal, A. subinfumata, A. pleuralis. Among these, A. nevadensis and A. scleritica previously known from the United States of America and A. luteiceps previously known from the Palaearctic region are here newly recorded in Canada.
Table 1 Checklist of world species of Amauromyza with biogeographic regions.

Note: A. (Annimyzella) maculosa (Malloch), A. (Annimyzella) fuscibasis (Malloch), and A. (Annimyzella) ranchograndensis Spencer were transferred to the genus Nemorimyza and Amauromyza (Annimyzella) lathyroides Spencer was transferred to the genus Liriomyza (Zlobin Reference Zlobin1996); A. (Ceph.) soozi Zlobin is a synonym of A. (Ceph.) chamaebalani (Hering) (Spencer and Martinez Reference Spencer and Martinez1987).
There has been uncertainty in the past on the limit and validity of some subgenera of Amauromyza. The subgenera Amauromyza, Catalpomyza Spencer, and Cephalomyza remain valid but further investigation is needed (Zlobin Reference Zlobin1996). Most Canadian species belong to Cephalomyza (recognised by the spiny membrane associated with the distiphallus), except A. pleuralis, which is in the monotypic subgenus Catalpomyza.
Host plants are known for 26 species of Amauromyza. Among these, 23 species (excluding Amauromyza morionella novakii [Strobl Reference Strobl1902], a synonym of A. morionella [according to Spencer Reference Spencer1966]) are listed in Benavent-Corai et al. (Reference Benavent-Corai, Martinez and Jiménez Peydró2005). Additions to that list include Amauromyza (A.) aliena (Malloch) on Callicarpa formosana Rolfe (Lamiaceae) (Spencer Reference Spencer1986); Amauromyza (A.) clinopodii Sasakawa on Clinopodium chinense grandiflorum (Maximovicz) (Lamiaceae) (Sasakawa and Matsumura Reference Sasakawa and Matsumura1998); and an undescribed species Amauromyza (Ceph.) sp. (Cuba) reared from Commelina erecta Linnaeus (Commelinaceae) (Spencer and Stegmaier Reference Spencer and Stegmaier1973; Spencer Reference Spencer1990). Species with known natural history are either linear-blotch miners or internal stem-borers. In Canada, the host plants have been confirmed for only four species of Amauromyza and belong to the families Elaeagnaceae, Amaranthaceae, Caryophyllaceae, and Bignoniaceae.
Materials and methods
This study is based on specimens housed in the Biodiversity Institute of Ontario, Guelph, Ontario, Canada (BIOUG); the Canadian National Collection of Insects, Arachnids & Nematodes, Ottawa, Ontario, Canada (CNC); the University of Guelph Insect Collection, Guelph, Ontario, Canada (DEBU); Lyman Entomological Museum, McGill University, Ste-Anne-de-Bellevue, Québec, Canada (LEM); the Museum of Zoology, Lund University, Sweden (MZLU); the Royal British Columbia Museum, Victoria, British Columbia, Canada (RBCM); and the Spencer Museum, University of British Columbia, Vancouver, British Columbia, Canada (UBCZ). Morphological terminology follows Cumming and Wood (Reference Cumming and Wood2009) except that orbital and frontal setae are referred to ors and ori, respectively. The clypeus is ∩-shaped and the thickness refers to the width of the sclerite between the outer and inner margins. Abbreviations are as follows: dc: dorsocentral bristles (numbered starting posteriorly); ors: orbital setae; ori: frontal setae. Distribution of species is based on published data and on specimens examined. Asterisks in distribution lists indicate new locality records.
Taxonomic treatment
Amauromyza Hendel
Redia Lioy (Reference Lioy1864): 1313. Preoccupied by Redia Filippi, 1837 (Trematoda). Type species: Agromyza gyrans Fallén (Reference Fallén1823), by subsequent designation of Coquillett (Reference Coquillett1910: 599). Synonymy Frick (Reference Frick1952: 394) (as Calycomyza). Synonymy Spencer and Martinez (Reference Spencer and Martinez1987: 256) (as Amauromyza).
Amauromyza Hendel (Reference Hendel1931a: 59) (as subgenus of Dizygomyza). Type species: Agromyza lamii Kaltenbach (Reference Kaltenbach1858), by original designation.
Phytobia (Amauromyza): Frick (Reference Frick1952: 393).
Amauromyza: Nowakowski (Reference Nowakowski1962: 97); Spencer (Reference Spencer1969b: 157); Spencer (Reference Spencer1971a: 252); Spencer (Reference Spencer1971b: 158); Spencer (Reference Spencer1976: 156); Spencer (Reference Spencer1981: 141); Spencer and Steyskal (Reference Spencer and Steyskal1986: 78); Zlobin (Reference Zlobin1996: 271).
Irenomyia Nowakowski (Reference Nowakowski1960: 421). Type species: Xeniomyza obscura Rohdendorf-Holmanová (Reference Rohdendorf-Holmanová1959), by monotypy. Synonymy Spencer and Martinez (Reference Spencer and Martinez1987: 256).
Melanophytobia Hering (Reference Hering1960: 127) (as subgenus of Phytobia). Type species: Phytobia chamaebalani Hering (Reference Hering1960), by monotypy. Synonymy Spencer and Martinez (Reference Spencer and Martinez1987: 256).
Campanulomyza Nowakowski (Reference Nowakowski1962: 97). Type species: A. gyrans Fallén (Reference Fallén1823), by monotypy. Synonymy Spencer (Reference Spencer1976: 156).
Trilobomyza Hendel (Reference Hendel1931b: 71) (as subgenus of Dizygomyza). Type species: Agromyza flavifrons Meigen (Reference Meigen1830), by original designation. Synonymy Spencer and Steyskal (Reference Spencer and Steyskal1986: 273).
Diagnosis
Canadian species of Amauromyza can be differentiated from other genera of Agromyzidae (subfamily Phytomyzinae) by a combination of characters including: orbital setulae erect, reclinate, or inclinate; vein M1 + 2 closer to wing tip than R4 + 5; wing with costa extending to apex of M1 + 2; foretibial and midtibial bristles absent; calypter fringe usually brown or black (yellow in A. pleuralis); halter usually black or partially darkened (completely yellow in A. flavifrons, A. karli, A. luteiceps, and A. pleuralis); anepisternum usually completely dark (except in A. pleuralis). Genitalic characters include: sperm pump of ejaculatory apodeme with characteristic bowl-shaped base; broad and largely rounded hypandrium; subepandrial sclerites broadly separated from each other; surstylus absent; pregonite without long inner process connecting it to anterior margin of aedeagal hood (Zlobin Reference Zlobin1996). Members of the subgenus Cephalomyza are characterised by having the distiphallus ending in paired sclerites, which are surrounded by a spinulose membrane (Spencer Reference Spencer1981).


Figs. 1–6 Head, lateral: (1) Amauromyza abnormalis (Malloch), (2) Amauromyza angulicornis Zlobin, (3) Amauromyza flavifrons (Meigen), (4) Amauromyza karli (Hendel), (5) Amauromyza luteiceps (Hendel), (6) Amauromyza nevadensis Spencer, (lower ors and lower ori missing). Scale bars = 0.1 mm.

Figs. 7–12 Head, lateral: (7) Amauromyza pleuralis (Malloch), (8) Amauromyza riparia Sehgal (holotype), (9) Amauromyza scleritica Spencer, (10) Amauromyza shepherdiae Seghal (upper ors missing), (11) Amauromyza subinfumata (Malloch). Clypeus (12) Amauromyza abnormalis (Malloch). Scale bars = 0.1 mm.

Figs. 13–18 Clypeus: (13) Amauromyza angulicornis Zlobin (female), inset: variation (male), (14) Amauromyza flavifrons (Meigen), inset: variation, (15) Amauromyza karli (Hendel), (16) Amauromyza luteiceps (Hendel) (male), inset: variation (female), (17) Amauromyza nevadensis Spencer, (18) Amauromyza pleuralis (Malloch). Scale bars = 0.1 mm.

Figs. 19–24 Clypeus: (19) Amauromyza riparia Sehgal (holotype), inset (paratype), (20) Amauromyza scleritica Spencer, (21) Amauromyza shepherdiae Sehgal, (22) Amauromyza subinfumata (Malloch). Head dorsal view: (23) Amauromyza angulicornis Zlobin. forecoxa (arrow – medial bristle). (24) Amauromyza pleuralis (Malloch) (male). Scale bars = 0.1 mm.
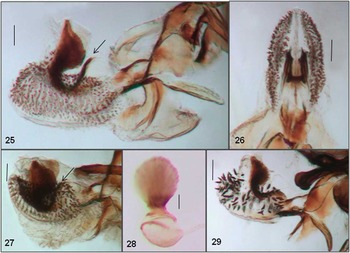
Figs. 25–29 Amauromyza abnormalis (Malloch): (25) phallus lateral, ON, (26) phallus ventral, ON, (27) phallus lateral, NM, (28) ejaculatory apodeme, ON. Amauromyza chenopodivora Spencer: (29) phallus lateral, Sweden. Scale bars = 0.05 mm.

Figs. 30–33 Amauromyza angulicornis Zlobin: (30) phallus lateral, BC (holotype), (31) phallus ventral, BC (holotype), (32) phallus lateral, WY, (33) ejaculatory apodeme, BC (holotype). Scale bars = 0.05 mm.

Figs. 34–36 Amauromyza flavifrons (Meigen): (34) phallus lateral, ON, (35) phallus ventral, ON, (36) ejaculatory apodeme, ON. Scale bars = 0.05 mm.
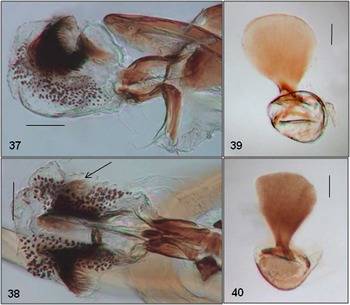
Figs. 37–40 Amauromyza karli (Hendel): (37) phallus lateral, ON, (38) phallus ventral, ON, (39) ejaculatory apodeme, ON, (40) ejaculatory apodeme, AB. Scale bars = 0.05 mm.

Figs. 41–43 Amauromyza luteiceps (Hendel): (41) phallus lateral, NB, (42) phallus ventral, NB, (43) ejaculatory apodeme, NB. Scale bars = 0.05 mm.

Figs. 44–46 Amauromyza nevadensis Spencer: (44) phallus lateral, (45) phallus ventral, (46) ejaculatory apodeme. Scale bars = 0.05 mm.

Figs. 47–52 Amauromyza pleuralis (Malloch): (47) phallus lateral, (48) phallus ventral, (49) ejaculatory apodeme. Amauromyza riparia Sehgal: (50) phallus lateral (holotype, permanent mount), (51) phallus lateroventral (holotype, permanent mount), (52) ejaculatory apodeme (paratype, permanent mount). Scale bars = 0.05 mm.

Figs. 53–56 Amauromyza scleritica Spencer: (53) phallus lateral, (54) phallus ventral, (55) ejaculatory apodeme, (56) abdomen female, with long oviscape. Scale bars = 0.05 mm, except Figure 56 = 0.2 mm.

Figs. 57–60 Amauromyza shepherdiae Sehgal, (57) phallus lateral (holotype, permanent mount), (58) phallus lateral, MB, (59) phallus ventral (also visible, postgonites and hypandrium), MB, (60) ejaculatory apodeme, MB. Scale bars = 0.05 mm.

Figs. 61–63 Amauromyza subinfumata (Malloch): (61) phallus lateral, QC, (62) phallus ventral, QC, (63) ejaculatory apodeme (showing size relative to phallus). Scale bars = 0.05 mm, except Figure 63 = 0.1 mm.
Amauromyza (Cephalomyza) abnormalis (Malloch)
Agromyza abnormalis Malloch, Reference Malloch1913: 320.
Dizygomyza (Amauromyza) abnormalis: Hendel (Reference Hendel1931a: 59). (misidentification = Amauromyza chenopodivora Spencer).
Phytobia (Amauromyza) abnormalis: Frick (Reference Frick1952: 393; Reference Frick1959: 378).
Amauromyza abnormalis: Spencer (Reference Spencer1969b: 158; Reference Spencer1990: 65).
Amauromyza (Cephalomyza) abnormalis: Spencer (Reference Spencer1981: 148); Spencer and Steyskal (Reference Spencer and Steyskal1986: 80).
Specimens examined:CANADA:British Columbia: Robson, 13.vi.1952, H.R. Foxlee (1 ♀: CNC). Manitoba: Brandon, 6.viii.1958, J.G. Chillcott (2 ♀: CNC); same except 10.viii.1958 (1 ♀: CNC); 5 mi. N. Minnedosa, 8.vii.1958, R.L. Hurley (1 ♀: CNC); Winnipeg, St. Charles Rifle Range (49°54.6′N, 97°20.5′W), Malaise trap at poplar forest/tallgrass prairie edge 3–10.vii.1998, Pollock & Roughley (1 ♀: LEM). Ontario: Chatham, 29.viii.1928, A.B. Baird (1♀: CNC); Essex Co., East Sister I. Prov. Nat. Res., 41°49′N, 82°51′W, 30.vii.2003, dry pond bed, yellow pans, S.A. Marshall (9 ♂: DEBU); same except cormorant colony (4 ♂: DEBU); Essex Co., East Sister I. Prov. Nat. Res., 41°49′N, 82°51′W, 30.vii.2003, sweeping, S.A. Marshall, debu00238339 (1 ♂: DEBU); Essex Co., Middle I., 41°41′N, 82°41′W, yell. pans in mud, 11.vi.2003, S.A. Marshall (8 ♂: DEBU); except yellow pans, 4.vii.2003, Marshall, Paiero & Cheung (3 ♂: DEBU); Lambton Co., Port Franks, Watson Property near L-lake, 19.viii.1996, J. Skevington (1 ♂: DEBU); Ottawa, 11.vii.1957, J.E.H. Martin (1 ♀: CNC); Ottawa, Damp second-growth Acer-Betula wood, 5.vii.1994, J.R. Vockeroth (1 ♂: CNC); Marmora, 10.vi.1952, J.R. Vockeroth (1 ♂: CNC). Québec: Abbotsford, 19.vi.1937, G. Shewell (1 ♂: CNC). UNITED STATES OF AMERICA:Arizona: Patagonia, 13.vi.1992, Sonoita Creek Pres. Malaise, Grass/Nettle meadow, JS and AG (2 ♂: DEBU); Santa Cruz Co. Patagonia (31.53′N,110.77′W), Malaise trap, 28.v.1995, B. Brown & E. Wilk (4 ♂: LEM); same except 15.vii.1995 (1 ♂; 2 ♀: LEM); same except 23.vii.1995 (3 ♂: LEM); same except 13.viii.1995 (1 ♀: LEM); same except (31.58′N,110.77′W), Malaise trap, 01.vii.1995 (1 ♂: LEM). New Mexico: Grant Co. 14 mi. N. Silver City, Cherry Ck Cmpgrd (32°54.8′N, 108°13.6′W), 7400’, Malaise trap, 14–16.viii.2007, J.E. O'Hara (2 ♂: LEM); Roosevelt Co. Portales 4000’, 23–26.viii.1993, O'Hara & Jorgensen, Malaise (1 ♂: LEM).
Redescription. Frons width (including orbital plates) 0.27–0.40 mm; usually three inclinate ori (sometimes four) and two reclinate ors (upper ors sometimes directed slightly outwards); orbital setulae short, erect, or reclinate; frons strongly projecting in profile and parafacial broad, forming distinct ring below eye (Fig. 1); gena deep, at midpoint approximately 0.30 times maximum eye height, gena much deeper at rear (Fig. 1); first flagellomere rounded; arista normal, approximately 0.35–0.40 mm long (at least 3.5 times length of first flagellomere); clypeus shiny or subshiny, medium thickness; epistoma broad (Fig. 12). 3 + 1 well-developed dc; acr in approximately five rows. Wing length 1.85–2.20 mm (♂) and 2.20–2.70 mm (♀) (up to 3.20 mm according to Spencer Reference Spencer1969b); last section of CuA1 approximately 2.0–2.3 times penultimate.
Colour. Head usually completely brown, sometimes frons and gena paler, orange brown or yellowish brown; face, orbit, and parafacial sometimes contrastingly darker (black); clypeus black; epistoma pale brown or beige; thorax, abdomen, and legs completely brown; halter mostly brown except white at suture (narrowly or more extensively, sometimes leaving only a black spot apically); calypter pale brown, margin and fringe dark brown.
Genitalia: Phallus with distinct shape in lateral view (Figs. 25, 27); membrane of distiphallus large, covered with numerous small spines; ejaculatory apodeme with broad blade and large bowl-shaped base (Fig. 28).
Geographic distribution: Canada (BC*, MB*, ON, QC); United States (various states from CA to DC); Mexico (Bautista-Martinez et al. Reference Bautista-Martinez, Vejar-Cota and von Tschirnhaus1997).
Host plant and natural history: Larva develops and pupates in stems of Amaranthus spp., (Amaranthaceae) (Spencer Reference Spencer1990).
Comments: Spencer (Reference Spencer1981: 150) mentioned that the male specimen from Québec may represent a different species than the male from California (both illustrated in Spencer (Reference Spencer1981) and Spencer and Steyskal (Reference Spencer and Steyskal1986)). Examination of the Québec specimen (QC: Abbotsford, CNC) confirmed that it is A. abnormalis and that the phallus is not as illustrated in Spencer (Reference Spencer1969b, Reference Spencer1981) but is most similar to specimens examined from New Mexico (Fig. 27). Although this species had been considered Holarctic (Hendel Reference Hendel1931a; Papp Reference Papp1984), records of A. abnormalis in the Palaearctic are not valid (Spencer Reference Spencer1976); they represent A. chenopodivora Spencer (Spencer Reference Spencer1971b), a similar species restricted to the Palaearctic region. Specimens of A. chenopodivora examined from Sweden (3 ♀; 3 ♂: MZLU) are slightly larger: 2.10–2.40 (♂) and 2.75–2.90 mm (♀) than A. abnormalis and the last section of CuA1 is smaller, usually less than twice (approximately 1.85) the length of the penultimate section. The shape of the distiphallus (Fig. 29) is slightly different, especially in lacking the narrow basal extension that curves upwards in A. abnormalis (arrow, Figs. 25, 27). The spines on the membrane of the distiphallus of A. chenopodivora are larger and less numerous (Fig. 29) than in A. abnormalis (Figs. 25, 27).
Amauromyza (Cephalomyza) angulicornis Zlobin
Amauromyza (Cephalomyza) angulicornis Zlobin, Reference Zlobin1997: 88.
Type specimen examined: Holotype ♂. CANADA: British Columbia: Manning Park, 1700 m, 27.vi.1984, leg. R. Danielsson (MZLU) (head missing).
Other specimens examined:UNITED STATES OF AMERICA:Wyoming: Sheridan Co., Twin Buttes, (44.8060°, −107.5401°), 2441 m, sweep alpine meadow, 03.vii.2010, T.A. Wheeler (1♀: LEM); same except: Big Horn NF, Tongue Creek (44.8062°, −107.5399°), 2460 m, sweep alpine meadow, 02.vii.2010, A. MacLeod (1 ♂: LEM).
Redescription. Frons width (including orbital plates) 0.18–0.20 mm; orbital plates widening at level of ors (Fig. 23); four orbital bristles, including possibly three ori and one ors (as mentioned by Zlobin Reference Zlobin1997) or two ori and two ors (division and orientation of ori and ors not clear); orbital setulae slightly reclinate; frons slightly projecting in profile, parafacial weakly developed, forming only narrow ring below eye; gena deepest at midpoint; approximately 0.26–0.30 times maximum eye height; first flagellomere slightly angulate at upper corner (Fig. 2); arista short, 0.18 mm (approximately twice length of first flagellomere); clypeus medium-thick, rounded; width 0.6 mm (arrow, Fig. 13); clypeus sunken deep inside mouth margin; epistoma well developed (female, Fig. 13) or absent (male, inset Fig. 13). At least 4 + 1 dc (possibly more, but weakly developed); acr in approximately 5–6 irregular rows in holotype, 3–4 rows in other specimens. Wing length 1.4–1.6 mm in males,1.5 mm in female; last section of CuA1 approximately 2.6–2.8 times penultimate.
Colour. Head completely dark brown, except gena slightly paler brown; thorax, abdomen and legs all brown; calypter, margin and fringe brown; halter with pale brown stalk, knob mostly white or yellow, but narrowly pale brown at base.
Genitalia: Spinulose membrane of phallus well defined (Figs. 30–32); distiphallus short, membranous, about same length as mesophallus and extending slightly beyond spinulose membrane (most clearly visible in Fig. 32); mesophallus with well sclerotised ridge ventrally; ejaculatory apodeme with narrow blade and moderately large cone-shaped base (Fig. 33).
Geographic distribution: Western Nearctic (BC; WY*).
Host plant and natural history: Unknown.
Comments: This species was previously known only from the holotype, which has lost its head subsequent to the original description. Two additional specimens were identified in this study. The species is easily distinguished by its slightly angulate first flagellomere, short arista, and gena deepest in the centre.
Amauromyza (Cephalomyza) flavifrons (Meigen)
Agromyza flavifrons Meigen, Reference Meigen1830: 184.
Dizygomyza (Trilobomyza) flavifrons: Hendel (Reference Hendel1931b: 71).
Trilobomyza flavifrons: Spencer (Reference Spencer1969b: 160).
Amauromyza (Trilobomyza) flavifrons: Spencer (Reference Spencer1971b: 159); Spencer (Reference Spencer1976: 162); Spencer (Reference Spencer1987: 877).
Amauromyza (Cephalomyza) flavifrons: Spencer and Steyskal (Reference Spencer and Steyskal1986: 85, 275).
Specimens examined:BELGIUM: Welle, Wellemeersen Nature Reserve (50°54′N, 04°02′E), sweep 06.vii.2000, S.E. Brooks (1 ♀: LEM). CANADA:British Columbia: Penticton, West Bench, 11.vii.1986, R.A. Cannings, ENT991-13881 (1 ♀: RBCM); Point Grey, Vancouver, 20.v.1973 (2 ♀: CNC); same except, 22.vii.1973, J.R. Vockeroth (3 ♂; 4 ♀: CNC); same except 12.ix.1972 (2 ♀: CNC); West Bench, Penticton, 4.vi.1983, S.G. Cannings (1 ♂: UBCZ). Ontario: Algoma Distr., Tenby Bay, Lakeshore, Malaise, 30.vii.1992, J.E. Swann, debu00305901 (1 ♀: DEBU); same except 12.vii.1992, debu00298010 (1 ♀: DEBU); same except 13.viii.1992, debu00302367 (1 ♀: DEBU); Algoma Distr., Hilton Beach, 46°15′N, 83°53′W, 23.viii.1992, edge of hardwood forest & field, Malaise, J.E. Swann, debu00297245 (1 ♀: DEBU); same except 5.vii.1992, debu00297180 (1 ♂: DEBU); Cambridge, 27.viii.1979, K. Barber (1 ♀: DEBU); same except 20.viii.1979 (1 ♀: DEBU); Cherry Valley, Pr. Edward Co., 14.viii.1968, J.R. Vockeroth (1 ♂: CNC); Essex Co., Point Pelee Natl. Pk., Visitor Centre, Malaise & pans, O. Lonsdale, 5–26.ix.2000, debu01007104 (1 ♀: DEBU); same except 26.ix-10.x.2000, debu01006835 (1 ♀: DEBU); same except 10–18.x.2000, debu01006739 (1 ♀: DEBU); same except 18.x-19.xi.2000, debu01006439 (1 ♀: DEBU); Forks of the Credit River, 5.vi.1981, J. Cairns (1 ♂: DEBU); Grey Co., Hepworth, Grey Sauble Authority Forests sweep of mature pine, 27.viii.1993, D.C. Caloren (2 ♂: DEBU); Guelph, 27.vii. 1977, P. R. Heels (1 ♀: DEBU); Guelph, 22.vii.1978, B. Menard (1 ♀: DEBU); Guelph Dow backyard, veg sweep, 18.vii.1996, J.M. Dow (1 ♀: DEBU); Halton Co., Milton, 16 mile Crk. & 4th line sweep, 9.vi.2001, S. Paiero, debu00172584 (1 ♀: DEBU); Hilton Twp. Tenby Bay, Malaise at edge of lake & poplar forest, 26.vi.1992, J.E. Swann (1 ♀: DEBU); Lambton Co., Port Franks Watson Property near L-lake, 15–17.vii.1996, Malaise traps, J. Skevington (1 ♀: DEBU); Orleans, Chapel Hill, 28.v.1990, J.R. Vockeroth (1 ♀: CNC); Ottawa, Damp second-growth Acer-Betula wood, 27.vi.1989, J.R. Vockeroth (1 ♀: CNC); same except, 27.vi.1993 (1 ♂; CNC); same except, 10.vii.1989 (1 ♀: CNC); same except, 12.vii.1989 (1 ♀: CNC); same except, 15.vii.1989 (1 ♀: CNC); same except, 12.vii.1990 (1 ♂; 1 ♀: CNC); same except, 13.vii.1989 (1 ♀: CNC); same except, 18.vii.1989 (1 ♀: CNC); same except, 26.vii.1989 (1 ♀: CNC); same except, 18.viii.1989 (1 ♀: CNC); same except, 28.viii.1989 (1 ♂: CNC); same except, 10.ix.1989 (1 ♀: CNC); Owen Sound, 12.viii.1975, J.M. Cumming (1 ♀: DEBU); Pt. Pelee, 7.vii.1980, D.L. Krailo (1 ♂; 2 ♀: CNC); same except, 9.vii.1980, S. Beierl (1 ♂; 1 ♀: CNC); Stittsville, 19.viii.1968, J.R. Vockeroth (1 ♂: CNC); Thornhill, 30.v.1964 (1 ♂: CNC); Wellington Co., Fergus, Malaise, 21.v.1992, Marshall & Cannings (1 ♀: RBCM); Wellington Co., Guelph, Stone Rd. E. 2.ix.1992, Malaise, R.A. Cannings (1 ♀: RBCM); Weston, 22.vii.1978, W. Ralley (1 ♂: DEBU). Québec: Gaspé, Haldimand (48°47′N, 64°22′W), sweep path in forest, 18.viii.2001, H. Varady-Szabo (1 ♂: LEM); Mont St. Hilaire (45°32′N, 73°09′W), sweep vegetation at parking lot, 25.v.2001, T.A. Wheeler (2 ♀: LEM); Mont St. Hilaire Reserve, Rocky trail, sweep trail in forest, 19.vi.2007, Lyman students (1 ♀: LEM); Pointe Fortune Conservation Area (45°34′N, 74°23′W), 7.vii.1999, S.E. Brooks (1 ♀: LEM); Ste-Anne-de-Bellevue, Stoneycroft Pond (45°25.8′N, 73°56.4′W), Malaise near pond, 24.v.1998, S.E. Brooks (1 ♀: LEM). UNITED STATES OF AMERICA:Michigan: Marquette Co. ne/se T51N-R28W S 14, Emgd: 21.iii.2002, Surv: R.J. Priest. Reared EX Silene pratensis, Recv 24.ix.2001, LOT: RJP 1242.5 (1 ♂: LEM); same except, Emgd 8.iii.2002, LOT: RJP 1242.2 (1 ♀: LEM); same except, Emgd 19.iii.2003, Recv 23.ix.2002, LOT: RJP 1353.10 (1 ♀: LEM); same except, Emgd 22.iii.2002, LOT: RJP 1242.17 (1 ♀: LEM). Oregon: Marion Co., 1–5 rest area, 4.5 km NW Jefferson (44°44.41′N, 123°03.30′W), 60 m, 21.vi.2008, C. Borkent (1 ♀: LEM). Vermont: So. Randolf, Hwy 14, 27.vi.1980, B.V. Peterson (1 ♀: CNC). Washington: Mt. Baker, ne slope 900 m, 27.v.1973, J.R. Vockeroth (2 ♂: CNC).
Redescription. Frons width 0.25–0.33 mm; three inclinate ori and two reclinate ors, sometimes two or four ori on one side only; orbital setulae mostly inclinate or reclinate; frons not or very slightly projecting anterodorsally; gena at midpoint approximately 0.16–0.19 times maximum eye height, gena slightly deeper at rear (Fig. 3); first flagellomere rounded, with patch of short white setae apically; arista normal, length 0.30–0.40 mm (at least 3.5 times length of first flagellomere); clypeus thick with upper margin rounded; epistoma variable, moderately broad (Fig. 14), or weakly developed (Fig. 14, inset), upper margin rounded, not projecting anteriorly towards antennae. 3 + 1 well-developed dc; acr in approximately five rows. Wing length 1.8–2.0 mm in males and 2.00–2.25 mm in females (up to 2.7 mm according to Spencer Reference Spencer1969b); cell dm small: last section of CuA1 approximately 2.3–3.0 times length of penultimate.
Colour. Frons and orbit yellow with vertical bristles on dark background; antenna, face, and palpus usually brown, but antenna sometimes paler to almost completely yellow; gena yellow (pale brown in one specimen); clypeus dark brown; epistoma white; thorax, abdomen, and legs brown (rarely rusty orange), foreknee faintly yellow; halter completely yellow; calypter and fringe brown.
Genitalia: Distiphallus consisting of one pair of well-sclerotised, black processes with spinulose membrane small and restricted to apex (Figs. 34, 35); ejaculatory apodeme large, with small bowl-shaped base (Fig. 36).
Geographic distribution: Eastern (ON, QC*) and Western (BC*) Canada; United States (DE, MI*, OH, OR*, PA, VT*, WA*, WI); Palaearctic (Spencer Reference Spencer1976).
Host plant and natural history: Larva forms a white blotch mine on many genera of Amaranthaceae and Caryophyllaceae (Spencer Reference Spencer1990). Mines were illustrated in Spencer (Reference Spencer1976) and Spencer and Steyskal (Reference Spencer and Steyskal1986).
Comments: This species was previously excluded from the subgenus Cephalomyza based in part on the apparent lack of spinules around the distiphallus (Spencer Reference Spencer1976). Although the spinulose membrane is not as developed in this species as in other species in the subgenus, it is still present apically, and the species was transferred to Cephalomyza by Spencer and Steyskal (Reference Spencer and Steyskal1986). This species was previously known from the northeastern Nearctic region. Its presence in Oregon, Washington, and British Columbia represents a considerable range extension.
Amauromyza (Cephalomyza) karli (Hendel)
Dizygomyza Karli Hendel, Reference Hendel1927: 253.
Dizygomyza (Cephalomyza) Karli: Hendel (Reference Hendel1931a: 34).
Amauromyza (Cephalomyza) karli: Spencer (Reference Spencer1969b: 158; Reference Spencer1977: 242).
Specimens examined: CANADA:Alberta: Red Deer River, NW Dinosaur Prov. Pk. (50°50.5′N, 111°36.5′W), 06–09.vii.1997, J.E. O'Hara. Malaise trap (2 ♂; 4 ♀: LEM); S of Onefour (49°00.7′N, 110°26.6′W), Malaise trap, 14–16.vii.1997, J.E. O'Hara (1 ♂: LEM). Manitoba: Aweme, Criddle farm (49°42′N, 99°36′W), sweep prairie N of house, 13.vi.1999, S. Boucher (1 ♀: LEM); near LaSalle, La Barrière Pk., sweep vegetation near river 15.vi.1999, S. Boucher (1 ♀: LEM); Winnipeg, Beaudry Prov. Pk. (49°51.5′N, 97°28.6′W), sweep disturbed meadow near Assiniboine River, 18.vi.1999, T.A. Wheeler (1 ♀: LEM). Ontario: Algonquin Prov. Pk., Arowhon Rd., Simm's Pit, 45°34′19′′N, 78°42′41′′W, yellow pans, 15–16.viii.2002, M. Buck, debu00225814 (1 ♂: DEBU); Elora, 11.viii.1971, G.A. Surgeoner (1 ♀: DEBU); Erin, 25.vii.1979, J. Ernst (1 ♀: DEBU); Manitoulin Island, 10 km W Gore Bay (45°53′N, 82°34′W), Malaise trap in grassland alvar, 21.viii.1996, P. Bouchard (1 ♀: LEM); Metcalfe, 4.viii.1991, B.E. Cooper (1 ♂: CNC); Oxford Co., Black Creek Swamp, near Drumbo, 20.vi.1993, J. Skevington, Black Ash wetland, sweep over body of roadkill snapping turtle (1 ♀: DEBU); Stonecliffe, Driftwood Prov. Pk., 2.ix.−5.x.79, S. Peck, Malaise, open pine plant, sand & heath (1 ♀: DEBU); Thunder Bay Distr., Neys Prov. Pk., Dune Trail, 48°46′52N, 86°36′53W, Malaise, 7–19.vii.2002, M. Buck, debu00215484 (1 ♂: DEBU). Québec: Chemin-du-Lac Bog (47°45′36.8′′N, 69°31′35.5′′W), pan trap, restored site, 21–26.vii.2006, A.G. Taillefer (1 ♂: LEM); Johnville Bog and Forest Park (45°20.7′N, 71°44.5′W), Malaise trap, 23–28.vi.2005, J. Savage & J. Kuchta (1 ♂: LEM); Ste-Anne-de-Bellevue, Stoneycroft Pond (45°25.8′N, 73°56.4′W), Malaise trap, 26.v.−01.vi.1999, S.E. Brooks (1 ♂; 3 ♀: LEM); St.-Charles Bog (46°45′48.0′′N, 70°59′34.4′′W), pan trap, abandoned site, 14–22.vi.2006, A.G. Taillefer (2 ♀: LEM); same except 7–14.vii.2006 (1 ♀: LEM); same except, Malaise trap, 7–9.vii.2006 (1 ♀: LEM); same except (46°44′52.1′′N, 70°59′42.0′′W), pan trap, restored site, 29.vi.–7.vii.2006 (1 ♀: LEM).
Redescription. Frons width 0.25–0.30 mm; three inclinate ori and one reclinate ors (one specimen with extra bristle on one side); orbital setulae inconspicuous, short and pale, mostly erect; frons strongly projecting in profile and parafacial broad, forming distinct ring below eye; gena very deep (Fig. 4), at midpoint approximately 0.35–0.45 times maximum eye height, gena strongly extended at rear; first flagellomere rounded or slightly elongated; arista normal, length 0.36–0.40 mm (at least 3 times length of first flagellomere); clypeus thin with upper margin triangular; epistoma conspicuously broad and projecting anteriorly towards antennae (Fig. 15). Normally 3 + 1 dc, but sometimes 3 + 2 or 4 + 2; acr in two to four irregular rows. Wing length 1.7–2.0 mm in males and 2.0–2.5 mm in females; last section of CuA1 approximately 1.8–2.3 times penultimate.
Colour. Head almost completely yellow, including antenna, face, epistoma, and palpus; at most upper orbit brown, sometimes just below ors; vertical bristles on dark background, but inner vertical sometimes at margin of yellow; clypeus brown; thorax and abdomen brown; legs mostly brown with femora narrowly yellow (about 0.1 times length of femora), tibiae sometimes narrowly yellow adjoining femora, and forecoxa sometimes yellow apically; halter yellow; calypter pale yellow or slightly darker, margin and fringe pale brown.
Genitalia: Distiphallus well sclerotised with spinulose membrane well defined; distiphallus with one tuft of setae present on side of each sclerite, most easily observed in ventral view (arrow, Fig. 38); ejaculatory apodeme with blade uniformly large (Fig. 39) or narrow basally and gradually expanding (Fig. 40).
Geographic distribution: Canada (AB*, MB*, ON, QC*); Palaearctic (Spencer Reference Spencer1969b, Reference Spencer1977).
Host plant and natural history: Unknown.
Comments: This Palaearctic species was first recorded in Canada by Spencer (Reference Spencer1969b) and was restricted to Ontario. Many new Canadian localities are here recorded and the species is now widespread. Amauromyza karli is similar to A. luteiceps, a Palaearctic species newly recorded in Canada (see below).
Amauromyza (Cephalomyza) luteiceps (Hendel)
Dizygomyza luteiceps Hendel, Reference Hendel1920: 133.
Dizygomyza (Cephalomyza) luteiceps: Hendel (Reference Hendel1931a: 34).
Dizygomyza hendeli de Meijere (Reference de Meijere1924: 138). Synonymy Hendel (Reference Hendel1931a: 34).
Amauromyza luteiceps: Spencer (Reference Spencer1969a: 13).
Amauromyza (Cephalomyza) luteiceps: Spencer (Reference Spencer1976: 160).
Specimens examined:CANADA:New Brunswick: St. Andrews, 8.vii.1978, S.A. Marshall (1 ♂: DEBU). Nova Scotia: Crescent Beach (44°13.8′N, 64°23.2′W), sweep dune vegetation, 24.vii.2002, J. Forrest & T. Wheeler (1 ♂: LEM). Québec: Gaspé, Rivière aux Renard, sweep in marsh, 12.viii.2001, H. Varady-Szabo (1 ♀: LEM).
Redescription. Externally similar to A. karli, except clypeus with upper margin not triangular but rather rounded or straight (Fig. 16); epistoma not as broad and not always projecting anteriorly towards antennae (Fig. 16, inset).
Colour. As described for A. karli, except legs more extensively yellow: all femora yellow for about 1/3 of length, tibiae narrowly yellow adjoining femora, and forecoxa yellow apically.
Genitalia: Phallus similar to A. karli, except without tufts of setae on distiphallus (Fig. 42) (compare with Fig. 38: A. karli), and with sclerite of distiphallus having a sharp projection, visible in lateral view (arrow, Fig. 41); ejaculatory apodeme gradually expanding and with moderately large bowl-shaped base (Fig. 43).
Geographic distribution: Eastern Canada* (NB, NS, QC); Palaearctic (Spencer Reference Spencer1969a, Reference Spencer1976).
Host plant and natural history: In Europe this species is a stem borer in Atriplex (Amaranthaceae) (Spencer Reference Spencer1990). Although the host plant is unconfirmed in Canada, specimens were collected near ocean dunes and marshes, which suggests that species of Atriplex may also be the host plant in North America.
Comments: This Palaearctic species is newly recorded in Canada. It is similar to A. karli, but is easily distinguishable externally by the more extensively yellow femora.
Amauromyza (Cephalomyza) nevadensis Spencer
Amauromyza nevadensis Spencer, Reference Spencer1981: 155; Spencer and Steyskal (Reference Spencer and Steyskal1986: 81).
Specimen examined:CANADA:Alberta: Waterton Lakes NP, Red Rock Parkway, (49.0996°, −113.9061°), 1390 m, sweep meadow, 08.vii.2010, T.A. Wheeler (1 ♂: LEM).
Redescription. Frons width (including orbital plates) 0.28 mm; apparently two inclinate ori and two ors (Spencer Reference Spencer1981 mentioned normal arrangement probably three ori and two ors, but number of ori may vary from two to four), lower ors somewhat inclinate and reclinate and upper ors directed posteriorly and outwards; orbital setulae inclinate or erect; frons moderately projecting in profile and parafacial well developed, forming distinct ring below eye; gena at midpoint 0.31 times maximum eye height; gena deepest at rear (Fig. 6); arista short, approximately 0.23 mm long (2.5 times length of first flagellomere), clypeus medium-thick, with upper margin straight; epistoma well developed, broad, and projecting towards antennae (Fig. 17). Apparently 4 + 2 well-developed dc (but number of dc irregular, sometimes 4 + 3 or 3 + 2 [Spencer and Steyskal Reference Spencer and Steyskal1986]); acr in approximately three to four irregular rows. Wing length 1.9 mm (up to 2.1 mm according to Spencer and Steyskal Reference Spencer and Steyskal1986); last section of CuA1 approximately 1.4 times penultimate (up to 2 times according to Spencer and Steyskal Reference Spencer and Steyskal1986).
Colour. Head mostly brown: frons and gena paler brown (both slightly yellowish brown posteriorly), orbit black, antenna dark brown. Mesonotum dark brown, subshiny; pleurites brown, legs all brown; calypter greyish black, margin and fringe brown; halter with knob mostly pure white, stalk and base of knob brown.
Genitalia: Apex of distiphallus elongate and curving ventrally in lateral view (Fig. 44); spinulose membrane of phallus well defined, looping ventrally below distiphallus in lateral view (Fig. 44); ejaculatory apodeme moderately large with small bowl-shaped base (Fig. 46).
Geographic distribution: Western Nearctic (AB*; CA).
Host plant and natural history: Unknown.
Comments: This species has the phallus most similar to A. scleritica (see below). Both species were described from the same locality in California (Spencer Reference Spencer1981) and are here newly recorded in Canada from the same locality in Alberta.
Amauromyza (Catalpomyza) pleuralis (Malloch)
Agromyza pleuralis Malloch, Reference Malloch1914: 311.
Phytobia (Trilobomyza) pleuralis: Frick (Reference Frick1953: 71; Reference Frick1959: 393).
Trilobomyza pleuralis: Spencer (Reference Spencer1969b: 161).
Amauromyza (Catalpomyza) pleuralis: Spencer (Reference Spencer1977: 242); Spencer and Steyskal (Reference Spencer and Steyskal1986: 83); Spencer (Reference Spencer1990: 218).
Specimens examined:CANADA:Ontario: St. Williams, Lake Erie, 21.ii.1962, Catalpa sp. (2 ♂: CNC); same except, 28.ii.1964 (1 ♂; 2 ♀: CNC).
Redescription. Frons width 0.25–0.28 mm; three inclinate ori and one reclinate ors; orbital setulae absent; frons moderately projecting and parafacial weakly to moderately developed (Fig. 7); gena at midpoint approximately 0.36 times maximum eye height, gena slightly deeper at rear; first flagellomere rounded; arista normal, length 0.35 mm (3 times length of first flagellomere); clypeus thin with upper margin rounded; no epistoma. Apparently 2 + 0 dc, additional dc present but about same size as acrostichals, these in approximately five rows. Male forecoxa with strong, white, flattened inner bristle (Fig. 24). Wing length 1.75–1.85 mm in males and 2.25 mm in females; last section of CuA1 approximately 1.3–1.5 times length of penultimate.
Colour. Frons, orbit and antenna yellow, sometimes dusted with grey; both vertical bristles on dark background; face and palpus yellow or partly grey; gena yellow; clypeus yellow; mesonotum grey or brown; pleura completely yellow except katepisternum, meron and katatergite brown; bristles on head, thorax and abdomen pale yellow (one female specimen with slightly darker bristles); legs completely yellow; halter yellow; calypter pale yellow, margin pale brown, fringe yellow or brown.
Genitalia: Distiphallus pale, membranous (Figs. 47–49), cup-shaped with a row of setulae near anterior margin (only visible at high magnification – at least 200×) and without distinct spinulose membrane; ejaculatory apodeme small and narrow with wide bowl-shaped base (Fig. 49); hypandrium broadly rounded with small apodeme at apex.
Geographic distribution: Eastern Canada (ON); eastern United States (IL, OH).
Host plant and natural history: Larva forms a small linear-blotch mine on various species of Catalpa (Bignoniaceae). Illustrations of the mine are in Spencer and Steyskal (Reference Spencer and Steyskal1986).
Comments: The overall pale body colour and pale bristles of this species are atypical of the genus Amauromyza but more commonly found among species of Phytoliriomyza. The enlarged base of the ejaculatory apodeme and low and broad hypandrium are typical of Amauromyza. Its isolated position in a monotypic subgenus will need further investigation. Although not easily visible in the male specimens examined, there may be a remnant of a spinular membrane on the distiphallus (Fig. 48, arrow), a characteristic that would justify its transfer into the subgenus Cephalomyza. An external defining character for this species is the presence of a strong, white, flattened medial bristle on the forecoxa in males (Tschirnhaus Reference Tschirnhaus1991). Spencer (Reference Spencer1969b) recorded this species from Turkey Point, Québec but that locality is, in fact, in Ontario.
Amauromyza (Cephalomyza) riparia Sehgal
Amauromyza riparia Sehgal, Reference Sehgal1971: 326.
Type specimens examined:Holotype:CANADA:Alberta: Edmonton, river bank near University of Alberta campus, 18.v.1969, V.K. Sehgal (1 ♂: CNC [#12066]); Paratype, same except, 14.v.1969 (1♂: CNC [#12066]).
Other specimens examined:CANADA:New Brunswick: Fredericton, 23.v.1958, on A. piceae infested bf, N.R. Brown (1 ♀: CNC); Québec: Bolton Pass, Knowlton, 800’, 5.vi.1963, J.R. Vockeroth (1 ♀: CNC); Old Chelsea, 15.v.1989, J.R. Vockeroth (1 ♀: CNC).
Redescription. Frons width, 0.23 mm; three inclinate ori and two reclinate ors, upper ors directed outwards; orbital setulae reclinate; frons slightly to moderately projecting, parafacial forming narrow ring below eye; gena at midpoint approximately 0.24–0.27 times maximum eye height, gena deepest at rear (Fig. 8); first flagellomere rounded, arista short, 0.18–0.20 mm (less than twice length of first flagellomere), slightly enlarged at base; clypeus medium-thick (slightly narrower in paratype than in holotype) (Fig. 19), with upper margin rounded; epistoma absent; Usually at least 4 + 1 or 4 + 2 dc, decreasing in size anteriorly, but presutural definitely longer than acrostichals (Sehgal Reference Sehgal1971 mentioned only three strong dc); acr in approximately five rows. Wing length 1.5–1.7 mm in males, up to 2.2 mm in females. Last section of CuA1 approximately 1.7 times length of penultimate.
Colour. Head (including antenna, face, and palpus) mostly brown; frons slightly paler yellowish brown above, blackish below; orbit mat, slightly darker than frons; mesonotum mat, pale grey; scutellum also grey except for paler yellow vertical line centrally; legs brown; halter apparently brown or at most dirty white (not “distinctly whitish or paler” as described by Sehgal Reference Sehgal1971); calypter and fringe brown.
Genitalia: Distiphallus narrow, elongated, and in lateral view, curving dorsally at an obtuse angle in basal third (Fig. 50); spinulose membrane closely appressed to distiphallus; ejaculatory apodeme slightly expanded on one side and with enlarged bowl-shaped base (Fig. 52).
Geographic distribution: Western Canada (AB); eastern Canada (NB*, QC*).
Host plant and natural history: The host plant is unknown. This species appears to be active in late spring.
Comments: This species was previously known only from the male types from Alberta. Three additional female specimens from Québec and New Brunswick agree externally with the characteristics of this species.
Amauromyza (Cephalomyza) scleritica Spencer
Amauromyza (Cephalomyza) scleritica Spencer, Reference Spencer1981: 160; Spencer and Steyskal (Reference Spencer and Steyskal1986: 84).
Specimens examined:CANADA:Alberta: Waterton Lakes NP, Red Rock Parkway (49.0996°N, −113.9061°W), 1390 m, sweep meadow, 08.vii.2010, T.A. Wheeler (1 ♂; 1♀: LEM). UNITED STATES OF AMERICA:California: Truckee, 6000’ Tahoe Co., 14.vii.1961, B.H. Poole (1♀: CNC).
Redescription. Frons width 0.22–0.30 mm; four orbital bristles; orientation and division between ori and ors not clear, normal arrangement probably three inclinate ori and one reclinate ors (California female specimen with extra upper ors on both sides); orbital setulae inclinate, some slightly proclinate; frons strongly projecting in profile and parafacial broad (Fig. 9); gena very deep, at midpoint approximately 0.30–0.38 times maximum eye height, gena strongly extended at rear; first flagellomere rounded, with patch of white setae apically; arista short, 0.16–0.20 mm (not more than twice length of first flagellomere); clypeus narrow with upper margin rounded; epistoma broad and projecting anteriorly towards antennae (Fig. 20). Mesonotum with 4 + 2 dc, decreasing in size anteriorly; acr in approximately three to four rows. Wing length 1.4 mm in male and 2.0 mm in females; last section of CuA1 approximately 1.4–1.7 times length of penultimate (see “Comments”). Female with long oviscape, approximately half length of abdomen in dorsal view (Fig. 56).
Colour. Frons, gena and most of orbit yellow; upper orbit brown (above and sometimes at level of upper ors); both vertical bristles on dark background; antenna and palpus brown; face yellowish brown; clypeus black; epistoma pale yellow; mesonotum dark greyish brown, scutellum sometimes slightly paler brown; pleura brown except for notopleuron sometimes paler, yellow or yellowish brown; legs completely brown (at most foreknee faintly yellow in one specimen); halter with knob mostly yellow (partially pale brown basally) and stalk brown; calypter and fringe brown. Female abdomen with apex narrowly yellow (Fig. 56).
Genitalia: Phallus somewhat similar to A. nevadensis, but conspicuously enlarged basally and narrowing apically for only a short distance, not curving ventrally (Fig. 53); ejaculatory apodeme narrow with large bowl-shaped base (Fig. 55).
Geographic distribution: Western Nearctic (AB*, CA).
Host plant and natural history: Unknown.
Comments: This species was previously known only from California (Spencer Reference Spencer1981). The specimens examined differ from Spencer's description by having a shorter wing (1.4 mm in male compared with 1.75–1.90 mm in Spencer's description) and a larger discal cell: last section of CuA1 1.4–1.7 times the length of penultimate (compared with almost twice the length of penultimate in Spencer's description).
Amauromyza (Cephalomyza) shepherdiae Sehgal
Amauromyza shepherdiae Sehgal, Reference Sehgal1971: 327; Spencer (Reference Spencer1990: 141).
Type specimen examined:Holotype ♂:CANADA:Alberta: Edmonton, University of Alberta campus, 5.vii.1968, from leaf mines on Shepherdia canadensis (L.), emerged 25.v.1969, V.K. Sehgal (CNC) (head missing).
Other specimens examined:CANADA: Manitoba: 4 km SE Churchill, Dene Village, (58.734°, −994.112°), Malaise trap, 14.vii.2008, A. Thielman (5 ♂: BIOUG).
Redescription. Frons width 0.22–0.23 mm; frons not projecting in front of eye in profile; normally two inclinate ori (one specimen with three ori on one side) and two reclinate ors, lower ors closer to posterior ori than to upper ors (may appear as three ori and one ors when bristles are lacking); orbital setulae sparse (about 9–10), erect or reclinate; gena at midpoint, approximately 0.20–0.23 times maximum eye height; first flagellomere rounded apically; arista moderately short, 0.25–0.27 mm (approximately 3 times length of first flagellomere); clypeus medium thickness with upper margin rounded (Fig. 21). Epistoma weakly developed or absent (Fig. 21). 3 + 2 dc present (presutural dc smaller); acr in approximately four to five irregular rows. Wing length 1.50–1.75 mm (male); last section of CuA1 approximately 2.20–2.75 times penultimate.
Colour. Frons, gena and face dark brown, orbit and ocellar triangle weakly shiny black, antenna black (Sehgal Reference Sehgal1971). Mesonotum and scutellum dark brown, subshiny; pleura brown, legs all brown; calypter, margin and fringe brown; halter with knob mostly white, stalk and base of knob brown.
Genitalia: Distiphallus mostly of uniform width in lateral view, with spinulose membrane small and restricted to distiphallus area (Figs. 57–58); distiphallus flaring apically in ventral view (Fig. 59); ejaculatory apodeme with small blade and large base (Fig. 60).
Geographic distribution: Western Canada (AB, MB*).
Host plant and natural history: This species forms blotch mines (Sehgal Reference Sehgal1971: Fig. 46) on leaves of S. canadensis (L.) Nuttall (Elaeagnaceae).
Comments: This species was previously known from a single damaged male from Alberta. This small dark species is most similar to A. riparia and A. subinfumata and can only be distinguished by the number of orbital bristles (four in A. shepherdia, five in the other two species) and by the male genitalia.
Amauromyza (Cephalomyza) subinfumata (Malloch)
Agromyza infumata Malloch, Reference Malloch1915a: 15.
Agromyza subinfumata (Malloch Reference Malloch1915b: 108). Replacement name for A. infumata Malloch, preoccupied by A. infumata (Czerny and Strobl Reference Czerny and Strobl1909).
Phytobia (Praspedomyza) subinfumata: Frick (Reference Frick1953: 71; Reference Frick1959: 395).
Amauromyza subinfumata: Spencer (Reference Spencer1969b: 159).
Amauromyza (Cephalomyza) subinfumata: Spencer and Steyskal (Reference Spencer and Steyskal1986: 81).
Specimens examined:CANADA:Alberta: Elkwater, 10.vi.1956, O. Peck (1 ♂: CNC). Manitoba: NE Glenboro, Spruce Woods Prov. Pk., Spirit Sands Trail, spruce forest open dune transition area, 8.vii.1992, T.A. Wheeler (1 ♀: LEM). New Brunswick: Chamcook, 30.vi.1965, G.E. Shewell (1 ♀: CNC). Ontario: Manotick, 22–29.v.1952 (1 ♂: CNC); Mer Bleue, Ottawa, 14.vi.1972, H.J. Teskey (1 ♂: CNC). Québec: Eardley, 14.v.1959, J.R. Vockeroth (1 ♀: CNC); Lac St.François Nat. Wildl. Area, Chemin Fraser (45°02.4′N, 74°28.03′W), Malaise trap beside sedge meadow, 18−26.v.1999, S.E. Brooks (2 ♂: LEM); Lanoraie Bog (45°59′N, 73°17′W), Malaise trap, 02.vi.1987, Sanborne, Genier & Hargreave (1 ♂: LEM); Roundtop Mt., Sutton, 1300’, 5.vi.1963, J.R. Vockeroth (1 ♀: CNC).
Redescription. Frons width 0.23–0.28 mm; three inclinate ori and two reclinate ors; orbital setulae erect or reclinate; frons and parafacial not projecting, or only slightly projecting (Fig. 11); gena at midpoint usually 0.13–0.22 (AB and MB specimens: 0.24–0.30) times maximum eye height, gena deepest at rear (Fig. 10); first flagellomere rounded, arista short, 0.22–0.24 mm (about 2.2–3.0 times length of first flagellomere); clypeus thick with upper margin rounded; epistoma absent or weakly developed (Fig. 21); mesonotum with 3 + 1 strong dc (one female with 4 + 2 dc, anterior presutural smaller); acr in approximately four to five rows. Wing length usually between 1.7–1.9 mm in both males and females (up to 2.2 mm in Spencer (Reference Spencer1969b)); last section of CuA1 approximately 1.8–2.5 times penultimate.
Colour. Head (including antenna, face, and palpus) mostly brown, except frons and gena sometimes slightly paler yellowish brown (gena especially paler in AB and MB specimens); frons sometimes paler posteriorly and black anteriorly (as in A. riparia); orbit well differentiated, usually slightly darker than frons; mesonotum dark brown or blackish grey, mat or weakly subshining; scutellum brown, sometimes slightly paler centrally; legs all brown; halter with stalk and most of knob pale brown, somewhat infuscated, knob dirty white apically; calypter and fringe brown.
Genitalia: Phallus with characteristic shape (Figs. 61–62), somewhat V-shaped in lateral view but with dorsal sclerite asymmetrical, slightly longer and upcurved (Fig. 61); spinulose membrane small, restricted to distiphallus area; ejaculatory apodeme with very large blade and enlarged bowl-shaped base (Fig. 63).
Geographic distribution: Widespread in Canada (AB*, MB*, NB*, ON, QC); eastern United States (IL).
Host plant: Unknown.
Comments: This species was previously considered eastern Nearctic, restricted to Ontario, Québec, and Illinois. The Alberta specimen is a considerable extension of the known range. One specimen from BC not listed above (8 km E Sicamous, 1.vi.1992, A. Borkent, 1 ♀: CNC) agrees mostly with A. subinfumata, except for a slightly deeper gena (as in AB specimen), slightly more developed epistoma, a paler brown, subshiny mesonotum, wing length 2.1 mm, and a paler, ochreous calypter and fringe. It may represent a new species but further material would be needed to confirm the status of this specimen.
Acknowledgements
I thank Brad Sinclair (CNC), Steve Marshall (DEBU), Karen Needham (UBCZ), Claudia Copley (RBCM), Valerie Levesque-Beaudin (BIOUG), Roy Danielsson (MZLU), and Owen Lonsdale (CNC) for loans of specimens. I also thank Chris Buddle (McGill) for access to his compound scope for genitalic photography and Terry Wheeler (LEM) for comments on the manuscript.















