



Introduction
Pyrochroidae Latreille, 1807, known also as the fire-coloured beetles, is a tenebrionoid group predominantly distributed in temperate regions worldwide, with 220 species in 33 genera and 5 subfamilies (Young and Pollock Reference Young, Pollock, Leschen and Beutel2010; Kai and Yoshitomi Reference Kai and Yoshitomi2018). Amongst these subfamilies, Pilipalpinae Abdullah, 1964 is a relatively well-studied component, exhibiting a classic Gondwanan distribution: Australia, New Zealand, Chile, and Madagascar. This group constitutes the sole pyrochroid subfamily in Australia and includes 18 described species classified into four genera (Pollock Reference Pollock1995).
The pilipalpine genus Binburrum was established by Pollock (Reference Pollock1995), comprising only five described species distributed from Tasmania, Victoria, and New South Wales to southern Queensland, Australia. The first three described species were placed originally in Techmessa F. Bates, 1874. This genus is now restricted to New Zealand. During a faunistic study on Australian Pyrochroidae, three undescribed species of Binburrum distributed in South Australia (one species) and northern Queensland (two species) were discovered in the Australian National Insect Collection, Commonwealth Scientific and Industrial Research Organisation (CSIRO), Canberra, Australian Capital Territory, Australia. After careful examination and anatomical comparison, we concluded that they were new to science. We describe them in the present paper. Additionally, B. angusticollis Pollock, Reference Pollock1995 is recorded from New South Wales for the first time.
Materials and methods
The specimens examined in this study are deposited in the collections of the Australian National Insect Collection, CSIRO, Canberra, Australian Capital Territory, Australia (ANIC) and the Museum of Victoria, Melbourne, Victoria, Australia (MV). The lectotype images of Binburrum ephippiatum (Wilson, Reference Wilson1926) are accessed from the Atlas of Living Australia website at https://images.ala.org.au/image/221a298f-d39c-478c-89a2-58acb0754839 [accessed 11 April 2020]. Data from specimens of described Binburrum species listed in Pollock (Reference Pollock1995) are not repeated here. We provide only detailed label data of imaged and mentioned specimens of previously described species.
Terminology used in the descriptions follows that of Pollock (Reference Pollock1995). Genitalia of both sexes were cleared in 10% potassium hydroxide solution, cleaned, and examined using a Leica M205C stereomicroscope (Leica Microsystems, Wetzlar, Germany) in glycerol on temporary slides. Genital structures were eventually stored in microvials in glycerol and pinned beneath the specimens. Images of external characters and male genitalia of B. concavifrons Pollock, Reference Pollock1995 were taken using a Leica DFC500 camera mounted on a Leica M205C microscope (Leica Microsystems). Source images were then aligned and stacked in the software program Leica Application Suite, version 4.9. Images of inner structures (excluding B. concavifrons) were taken using a Leica DFC 7000 T digital camera mounted on a Leica DIAPLAN compound microscope with Leica Application Suite X software (Leica Microsystems). Layers were then aligned in Helicon Focus (Helicon Soft, Kharkiv, Ukraine). Additional edits and plates were made in Photoshop CS6 software (Adobe, San Jose, California, United States of America). Body length was measured from the anterior margin of the clypeus to the elytral apices, and width was measured at the widest part of both elytra; absolute measurements are indicated in millimetres. A double backslash (//) is used to separate data from different labels on type specimens, and a single slash with a space before and after (/) is used to separate data from different lines on the same label. Abbreviations: ex(x). – specimen(s) of unidentified sex.
Genus Binburrum Pollock, 1995
Binburrum Pollock, 1995: 621. Type species, by original designation: Techmessa ruficollis Blackburn, 1895.
Diagnosis. Body small- to medium-sized (usually < 9 mm, ca. 3.5–8.5 mm), distinctly parallel sided. Head and pronotum covered with coarse, umbilicate punctures. Antennomeres elongate filiform; antennomere 4 shorter than combined lengths of antennomeres 2 and 3. Elytral epipleuron distinct, traceable to at least apical fourth of elytron.
Key to adults of Binburrum Pollock, 1995 (modified from Pollock Reference Pollock1995)


Fig. 1. Habitus of Binburrum, dorsal view. A, B. angusticollis Pollock, Reference Pollock1995, holotype; B, B. bifoveicollis (Lea, Reference Lea1917); C, B. concavifrons Pollock, Reference Pollock1995, holotype; D, B. ephippiatum (Wilson, Reference Wilson1926), lectotype; E, B. ruficollis (Champion, Reference Champion1895); B. articuno new species: F, holotype; G, paratype; H, B. moltres new species, holotype; I, B. sp., presumably colour variation of B. moltres new species; J, B. zapdos new species, holotype. Scale bar = 1.0 mm.

Fig. 2. Head of Binburrum new species, dorsal view. A, B. articuno new species: A, holotype and B, paratype; C, B. moltres new species, holotype; D, B. zapdos new species, holotype. Scale bar = 0.2 mm.

Fig. 3. Pronotum of Binburrum new species, dorsal view. Binburrum articuno new species: A, holotype and B, paratype; C, B. moltres new species, holotype; D, B. zapdos new species, holotype. Scale bar = 0.2 mm.
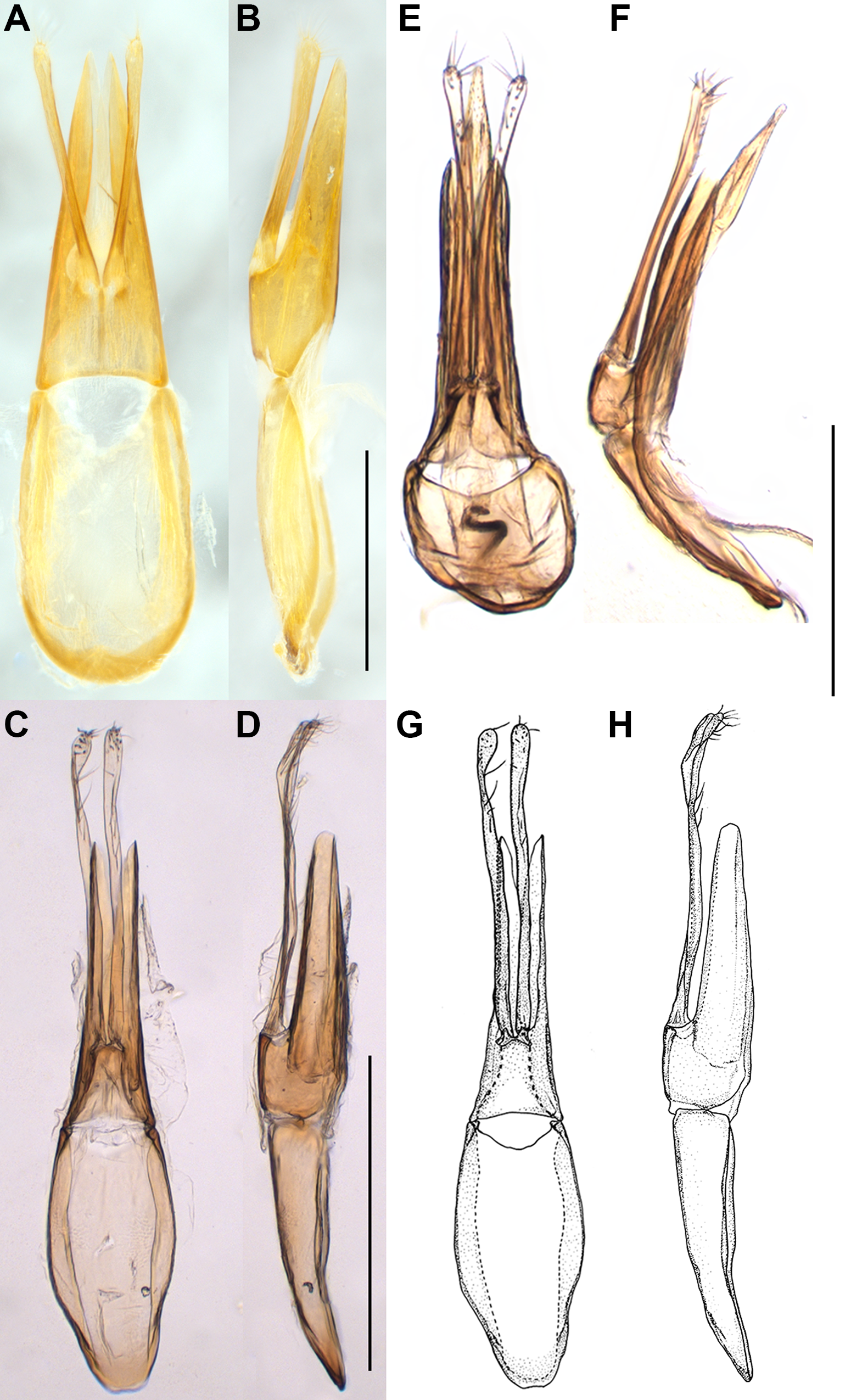
Fig. 4. Genitalic tegmen of Binburrum articuno new species, in comparison with B. concavifrons Pollock, Reference Pollock1995. Binburrum concavifrons Pollock, Reference Pollock1995, holotype: A, dorsal view and B, lateral view; B. articuno new species, holotype: C, dorsal view and D, lateral view; B. articuno new species, paratype: E, dorsal view, F, lateral view; B. articuno new species, holotype, hand drawings G, dorsal view and H, lateral view. Scale bar = 0.5 mm.
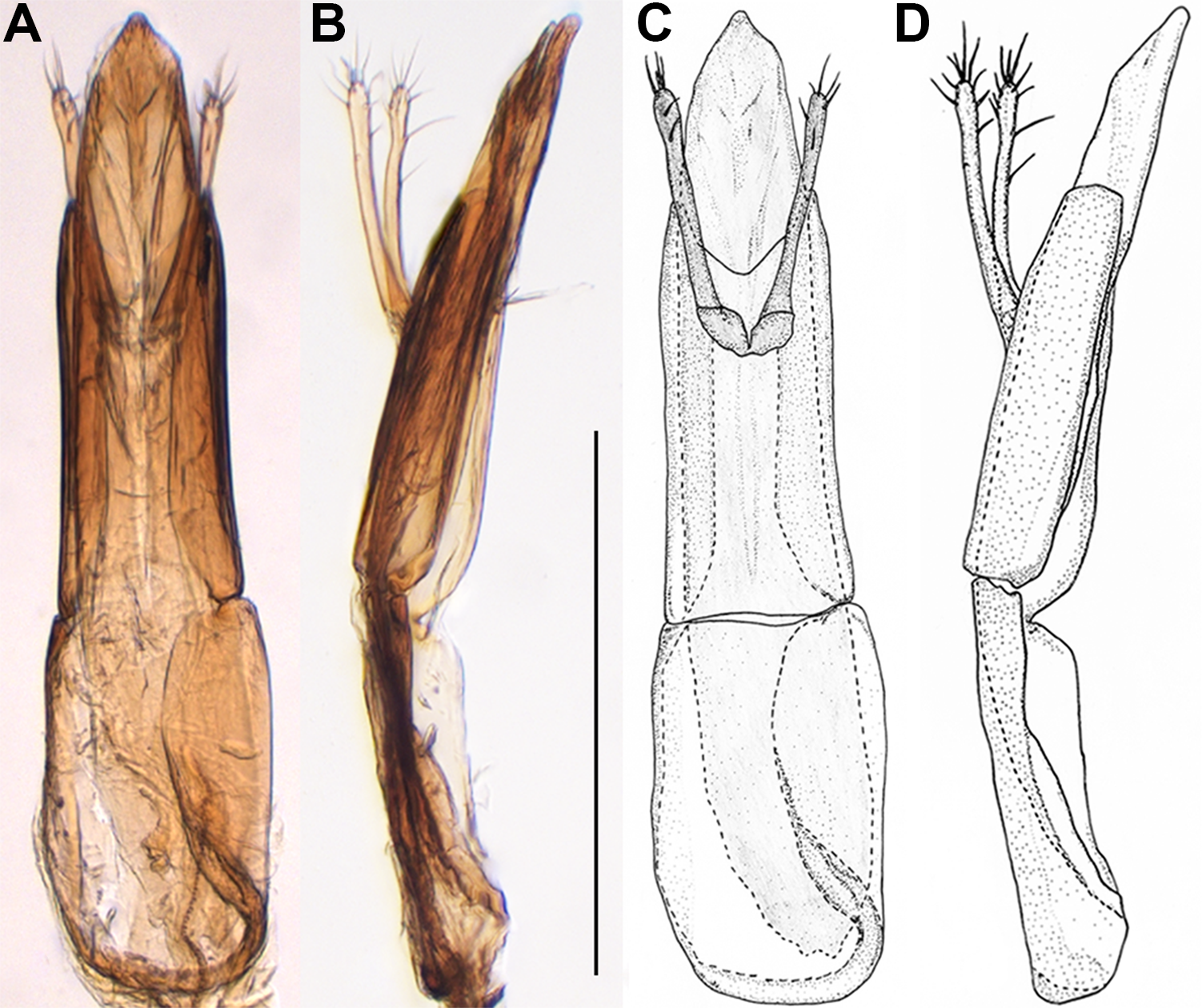
Fig. 5. Male genitalia of Binburrum moltres new species, holotype. A, dorsal view, B, lateral view; hand drawings C, dorsal view and D, lateral view. Scale bar = 0.5 mm.

Fig. 6. Genitalic tegmen of Binburrum zapdos new species, in comparison with B. bifoveicollis (Lea, Reference Lea1917) and B. ruficollis (Champion, Reference Champion1895). Binburrum bifoveicollis (Lea, Reference Lea1917): A, dorsal view; B, lateral view; B. ruficollis (Champion, Reference Champion1895): C, dorsal view and D, lateral view; B. zapdos new species, holotype: E, dorsal view, F, lateral view, and hand drawings G, dorsal view and H, lateral view. Scale bar = 0.5 mm.

Fig. 7. Comparison of median lobes between Binburrum concavifrons Pollock, Reference Pollock1995 and B. articuno new species. Binburrum concavifrons, paratype: A, ventral view and B, lateral view; B. articuno new species, holotype: C, ventral view and D, lateral view. Scale bar = 0.5 mm.
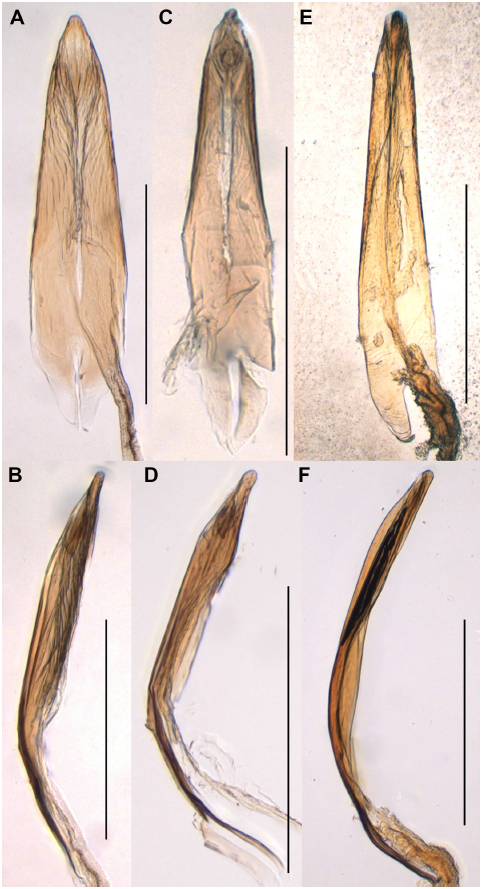
Fig. 8. Comparison of median lobes amongst Binburrum bifoveicollis (Lea, Reference Lea1917), B. ruficollis (Champion, Reference Champion1895), and B. zapdos new species. Binburrum bifoveicollis (Lea, Reference Lea1917): A, ventral view and B, lateral view; B. ruficollis (Champion, Reference Champion1895): C, ventral view and D, lateral view; B. zapdos new species, holotype: E, ventral view and F, lateral view. Scale bar = 0.5 mm.
Binburrum angusticollis Pollock, Reference Pollock1995
Figure 1A.
Binburrum angusticollis Pollock, Reference Pollock1995: 625.
Material examined (see Pollock (Reference Pollock1995) for details of other specimens). Holotype male. Queensland: 28.15 S 152.28 E / The Head, nr Wilsons / Peak QLD 13 Oct / 1984 1. Naumann and / J. Cardale coll. // ANIC // 011 // HOLOTYPE / Binburrum / angusticollis ♂ / Pollock (ANIC). Determined specimen. New South Wales: 30.22 S, 152.43 E, The Glade/Wonga Track, Dorrigo N.P., 13–15.xi.1990, A. Calder leg. (one male) (ANIC).
Distribution. Australia: northeastern New South Wales (new record) to southeastern Queensland.
Binburrum bifoveicollis (Lea, Reference Lea1917)
Techmessa bifoveicollis Lea, Reference Lea1917: 292.
Material examined (see Pollock (Reference Pollock1995) for details on other specimens). Queensland: Binna Burra, Lamington National Park, 27.x.1993, S.A. Ślipiński & J.F. Lawrence leg. (one male) (ANIC).
Distribution. Australia: eastern New South Wales to southern Queensland.
Binburrum concavifrons Pollock, Reference Pollock1995
Binburrum concavifrons Pollock, Reference Pollock1995: 628.
Material examined (see Pollock (Reference Pollock1995) for details on other specimens). Holotype male. Victoria: 37.43 S 145.41 E VIC / Mt. Donna Buang, 1200 m / N of Warburton / 811 26 Jan.-11 Feb. 1987 / A. Newton & M. Thayer // wet scler.-Noth. cunn. / FMHD #87-219 flight / interc. (window) trap // HOLOTYPE / Binburrum / concavifrons ♂ / Pollock (ANIC). Paratype (one male). 38.43 S 143.35 E VIC / Otway NP 390 m Binn / Rd. 4.3 km N. Cape Horn / 808 25 Jan.-8 Feb. 1987 / A. Newton & M. Thayer // wet scler.forest / FMHD #87-210 flight / interc. (window) trap // PARATYPE / Binburrum / concavifrons ♂ / Pollock (one male) (ANIC).
Distribution. Australia: Victoria.
Binburrum ephippiatum (Wilson, Reference Wilson1926)
Figure 1D.
Techmessa ephippiatum Wilson, Reference Wilson1926: 40.
Material examined (see Pollock (Reference Pollock1995) for details on other specimens). Lectotype ex. Queensland: HOLOTYPE & / PARATYPE / Techmessa / ephippiatum / Wilson // F. E. Wilson / Collection // 2135 - Type / 2136 Paraty // Techmessa / ephippiatum / Wilson / TYPE. // BlackallRgs. Q. / Oct. 1920. / F.E. Wilson. (MV).
Distribution. Australia: southeastern Queensland.
Remarks. The type material of Techmessa ephippiatum consists of two specimens on a single cardboard plate, the right specimen of which is marked as “Ty” in red ink. Pollock (Reference Pollock1995) designated this specimen as lectotype (with the other specimen designated a paralectotype); however, the label that was placed on the pin by Pollock indicated a holotype and paratype. This was in error, and a new “lectotype / paralectotype” label will be substituted for the erroneous label.
Binburrum ruficollis (Champion, Reference Champion1895)
Techmessa ruficollis Champion, Reference Champion1895: 248.
Material examined (see Pollock (Reference Pollock1995) for details on other specimens). New South Wales: 34°23ʹ S, 150°50ʹ E, Mt. Keira, 3 km W, 10.xi.1990, Tom Gush leg. (one male) (ANIC).
Distribution. Australia: Tasmania, and Victoria to southeastern New South Wales.
Binburrum articuno Hsiao and Pollock, new species
ZooBank Registration Number:
urn:lsid:zoobank.org:act:CCF17731-0E20-4DF4-8FD3-BE185D47677F
Figures 1F–G, 2A–B, 3A–B, 4C–H, 7C–D.
Type material. Holotype male. South Australia: 34.07S 140.37E (GPS) / 12km SW Calperum HS / S.A. 10 Oct-7 Nov 1995 / A.Lambie & K.Pullen / Flight intercept/trough trap // Calperum Station/ / Bookmark Biosphere / Reserve / invertebrate Survey” (ANIC). Paratype (one male, three ex.): “34.07S 140.37E S.A. / 14km WNW Renmark / 5 Sept-12 Oct 1995 / Flight intercept/ pitfall / trap K.R. Pullen // Calperum Station/ / Bookmark Biosphere / Reserve / Invertebrate survey” (two ex.) (ANIC); “33.53S 140.44E S.A. / 32km N Renmark / 6 Sept-12 Oct 1995 / Malaise trap, mallee / K.R.Pullen // Calperum Station/ / Bookmark Biosphere / Reserve / invertebrate Survey” (one male) (ANIC); 33.53S 140.44E S.A. / 32km N Renmark / 6 Sept- 12 Oct 1995 / Flight intercept/ pitfall / trap K.R. Pullen // CALPERUM STATION/ / BOOKMARK BIOSPHERE / RESERVE / INVERTEBRATE SURVEY (one ex.) (ANIC).
Diagnosis. This species is distinguishable from its congeners by the entirely dark-brown body, depressed frons, elongate antennae, lateral pronotal margins without tubercles, and male genitalia with long triangular apicale slightly longer than basale in lateral view, and elongate accessory lobes largely extended beyond apicale. This new species somewhat resembles B. concavifrons Pollock, Reference Pollock1995 in depressed frontoclypeal area and long triangular apicale of male genitalia in lateral view but can be easily recognised by the entirely dark-brown body (Fig. 1F–G) (head black to light brown; pronotum orange to dark brown; elytra yellowish-brown to black in B. concavifrons; Fig. 1C), frontoclypeal area slightly depressed (distinctly depressed in B. concavifrons), longer antennae, extended to apical fifth of elytra (antennae shorter, extended to elytral midlength in B. concavifrons), punctures on head and pronotum shallower, umbilicate (deeper, nonumbilicate in B. concavifrons), elytra more elongate, with length ca. 6.1–6.3 × pronotal length (shorter, with length ca. 4.4–5.0 × pronotal length in B. concavifrons), genitalic tegmen with apicale nearly parallel laterally in dorsal view and rounded apically in lateral view (Fig. 4C–H) (apicale narrowed apicad in dorsal view and obtusely pointed apically in lateral view in B. concavifrons; Fig. 4A–B), accessory lobes of tegmen largely extended beyond apicale (slightly extended beyond apicale in B. concavifrons).
Description. Measurements. Length: 4.25–5.05 mm (5.05 in holotype); width: 1.20–1.40 mm (1.40 in holotype).
Colour (Fig. 1F–G). Head black, with clypeus dark brown; antennae dark brown; pronotum dark brown to black; meso- and metaventrites, elytra, abdomen and legs dark brown; surface covered with pale, slightly yellowish setae.
Head (Fig. 2A–B). Eyes lateral, small, ratio of eye diameter to interocular space 1.0:2.1–2.5. Antennae elongate, extended to apical fifth of elytra. Punctures large, umbilicate; setae short; frontoclypeal area slightly depressed; frons evenly convex; antennal setae longer than setae on head, recurved apically.
Pronotum (Fig. 3A–B). Narrower than head and distinctly narrower than width across elytral humeri; pronotum wider than long; lateral pronotal margins slightly arcuate, subangulate at widest point, near midlength; margins smooth, without tubercles; median line absent; depressions indistinct, shallow, anteriorly divergent, situated near midlength; punctures moderately deep, umbilicate; punctation contiguous laterally, punctures irregularly distributed, sparsely distributed anteriorly and medially; setae longer than setae on head.
Elytra. Punctures relatively large, rounded, smaller towards elytral apex, with interpuncture distance slightly shorter than puncture diameter; epipleura distinct completely to elytral apex; elytra flat dorsally, sloped towards apex only.
Male genitalia. Apicale slightly longer than basale, cleft deeply apically, nearly parallel laterally in dorsal view and rounded apically in lateral view; accessory lobes very narrow, inserted together in proximal one-fourth of apicale, largely extended beyond apicale (Fig. 4C–H); median lobe distinctly narrowed apicad, spatulate distally (Fig. 7C–D).
Etymology. The specific epithet is named after the Articuno in the Japanese videogame Pokémon, one of the three legendary fictional birds, referring to the presumed rareness of this species that has only five known specimens. The specific name is treated as a noun in apposition. The specific names of new species given in this paper are in memory of this classic videogame, which inspired the first author to become a taxonomist (“real Pokémon trainer”) in his childhood.
Distribution. Australia: southeastern South Australia.
Binburrum moltres Hsiao and Pollock, new species
ZooBank Registration Number: urn:lsid:zoobank.org:act:10D65DC0-F248-4657-9DFC-B9F360FBE279
Type material. Holotype male. Queensland: “17.28S 145.29E QLD / BS1 Longlands Gap / 30Nv1995-7Jan1996 / L.Umback. 1150m / Malaise trap” (ANIC).
Diagnosis. This new species closely resembles B. ephippiatum (Wilson, Reference Wilson1926) in the orange-coloured body, with large, broad piceous fascia on elytra but can be easily recognised by the lighter orange-coloured body, with elytral piceous fascia in posterior half of elytra (Fig. 1H) (body colour darker, with fascia in middle of elytra in B. ephippiatum; Fig. 1D), head and pronotum densely punctate, with interpuncture space much shorter than puncture diameter, punctures deep, coarse (Figs. 2C, 3C) (sparsely punctate, with interpuncture space the same as or longer than puncture diameter, punctures shallow, fine in B. ephippiatum), punctures on elytra large, rounded, with interpuncture space subequal to puncture diameter (small, longitudinally subelliptical, with interpuncture space longer than puncture diameter in B. ephippiatum), genitalic tegmen with apicale subequal to basale in length (longer than basale in B. ephippiatum), concave, not deeply cleft apically in dorsal view and nearly straight, merely slightly deflexed away from accessory lobes in lateral view (Fig. 5) (apicale hoodlike, not cleft apically in dorsal view and sinuate, distinctly deflexed away from accessory lobes in lateral view in B. ephippiatum), accessory lobes of tegmen inserted in distal one-third of apicale, extending beyond apicale (accessory lobes of tegmen inserted in proximal half of apicale, not extending beyond apicale in B. ephippiatum), median lobe slightly narrowing apicad (Fig. 5A, C) (parallel laterally, slightly enlarged, bifid apically in B. ephippiatum).
Description. Measurements. Length: 4.0 mm; width: 1.0 mm.
Colour (Fig. 1H). Head dark orange; antennae with scape and pedicel orange, antennomere 1 dark orange, blackish externally, remaining antennomeres black; pronotum orange; meso- and metaventrites orange; elytra orange, with large, broad piceous fascia in posterior half of elytra; abdomen orange to brown; legs orange; surface covered with yellowish setae.
Head (Fig. 2C). Eyes lateral, small, ratio of eye diameter to interocular space 1.0:3.7. Antennae short, extended to elytral midlength. Punctures large, umbilicate; setae short; frons evenly convex; antennal setae longer than setae on head, recurved apically.
Pronotum (Fig. 3C). Distinctly narrower than head and distinctly narrower than width across elytral humeri; pronotum wider than long; lateral pronotal margins arcuate; margins smooth, without tubercles; median line absent; depressions distinct, oval, anteriorly divergent, situated near midlength; punctures moderately deep, umbilicate, regularly distributed; setae longer than setae on head.
Elytra. Punctures relatively large, rounded, smaller towards elytral apex, with interpuncture distance subequal to puncture diameter; epipleura narrowed abruptly at ventrite 3 or 4, not present to elytral apex; elytra not flat dorsally, distinctly sloped in posterior half.
Male genitalia. Apicale subequal to basale in length, concave, not deeply cleft apically, nearly parallel laterally in dorsal view and nearly straight, merely slightly deflexed away from accessory lobes in lateral view; accessory lobes very narrow, inserted together in distal one-third of apicale, moderately extended beyond apicale (Fig. 5); median lobe nearly parallel laterally, slightly narrowed apicad, spatulate distally (Fig. 5A, C).
Etymology. The specific epithet is named after the Moltres in the Japanese videogame Pokémon, one of the three legendary fictional birds, referring to the presumed rareness of this species that has only one known specimen. The specific name is treated as a noun in apposition.
Distribution. Australia: northeastern Queensland.
Remarks. A female specimen collected from the same area (17.27 S, 145.29 E) exhibits similar colouration, with black head and dark-brown pronotum (Fig. 1I). Given the similarity in distribution and the fact that colour variation frequently occurs in other Binburrum species, this specimen might represent a colour variety of B. moltres.
Binburrum zapdos Hsiao and Pollock, new species
ZooBank Registration Number: urn:lsid:zoobank.org:act:3AE66A1A-FF07-430 A-B4B7-27A86565DDE5
Figures 1J, 2D, 3D, 6E–H, 8E–F.
Type material. Holotype male. Queensland: “17.06S 145.36E QLD / Mt. Haig GS1 1150m / 31 Jul-31 Aug. 1995 / L.Umback / Malaise trap” (ANIC).
Diagnosis. This new species closely resembles B. bifoveicollis (Lea, Reference Lea1917) and B. ruficollis (Champion, Reference Champion1895) in the overall colouration, with blackish head and elytra and orange pronotum (Fig. 1B, E), but can be easily recognised by the rounded lateral pronotal margins, without tubercles (usually with small, setigerous tubercles in B. ruficollis), pronotal disc irregularly and relatively sparsely punctate, with medial region more glabrous (regularly and densely punctate in B. bifoveicollis and B. ruficollis), elytral punctures smaller, with interpuncture distance subequal to puncture diameter (punctures larger, with interpuncture distance slightly shorter than puncture diameter in B. ruficollis), genitalic tegmen with apicale slightly longer than basale (apicale and basale subequal in length in B. ruficollis), distinctly narrowed apicad and moderately cleft apically in dorsal view and broad, straight-sided and distinctly narrowed apicad in lateral view (Fig. 6E–H) (slightly narrowed apicad and moderately cleft apically in dorsal view and broad, straight-sided and distinctly narrowed apicad in lateral view in B. bifoveicollis; Fig. 6A–B) and (slightly narrowed apicad and deeply cleft apically in dorsal view and broad, round-sided and slightly narrowed apicad in lateral view in B. ruficollis; Fig. 6C–D), accessory lobes of tegmen largely extended beyond apicale (slightly extended beyond apicale in B. bifoveicollis), median lobe more elongate (Fig. 8E–F) (stouter in B. bifoveicollis and B. ruficollis; Fig. 8A–D).
Description. Measurements. Length: 4.0 mm; width: 1.2 mm.
Colour (Fig. 1J). Head black, with clypeus orange; antennae with scape, pedicel and antennomere 1 orange, antennomere 4–5 dark orange, blackish externally, remaining antennomeres dark brown to black; pronotum yellow to orange; meso- and metaventrites and elytra black; scutellum orange; abdomen dark brown to black; legs orange; surface covered with yellowish setae.
Head (Fig. 2D). Eyes lateral, small, ratio of eye diameter to interocular space 1.0:3.5. Antennae short, extended to elytral midlength. Punctures large, umbilicate; setae short; frons evenly convex; antennal setae longer than setae on head, recurved apically.
Pronotum (Fig. 3D). Slightly narrower than head and distinctly narrower than width across elytral humeri; pronotum wider than long; lateral pronotal margins rounded; margins smooth, without tubercles; median line absent; depressions indistinct, shallow, anteriorly divergent, situated near midlength; punctures moderately deep, umbilicate; punctures irregularly distributed, sparsely distributed anteriorly and medially; setae longer than setae on head.
Elytra. Punctures relatively large, rounded, smaller towards elytral apex, with interpuncture distance subequal to puncture diameter; epipleura narrowed abruptly at ventrite 3 or 4, not present to elytral apex; elytra flat dorsally, sloped towards apex only.
Male genitalia. Apicale slightly longer than basale, moderately cleft apically, distinctly narrowed apicad in dorsal view and broad, straight-sided and distinctly narrowed apicad in lateral view; accessory lobes very narrow, inserted together in proximal one-third of apicale, largely extended beyond apicale (Figs. 6E–H); median lobe distinctly narrowed apicad, spatulate distally (Figs. 8E–F).
Etymology. The specific epithet is named after the Zapdos in the Japanese videogame Pokémon, one of the three legendary fictional birds, referring to the presumed rareness of this species that has only one known specimen. The specific name is treated as a noun in apposition.
Distribution. Australia: northeastern Queensland.
Discussion
The natural history of Binburrum is poorly known. Pollock (Reference Pollock1995) indicated that larvae can be found under bark of a variety of hardwood trees, or Nothofagus (Nothofagaceae) and Eucalyptus (Myrtaceae), and also from various forest types. Adults are usually collected in flight-intercept traps or by general collecting off vegetation. Pollock (Reference Pollock1995) determined that Binburrum is the sister group to a clade comprising Morpholycus Lea and Cycloderus Solier (Australia and southern South America, respectively). Support for this relationship was based on a single larval character: possession of a low, dome-like antennal sensorium. It was also found that within Binburrum, several species exhibited reversals, specifically the accessory lobes inserted basad of apicale midlength (character 12.0; Pollock Reference Pollock1995) and pronotal depressions absent (character 25.0; Pollock Reference Pollock1995). Relatively few species of the genus were known by Pollock (Reference Pollock1995), and these phylogenetic hypotheses will likely be adjusted upon detailed examination of additional species.
Acknowledgements
The first author wishes to thank Adam Ślipiński (ANIC) for help with literature searches and Zhenhua Liu (Sun Yat-sen University, Haizhu District, Guangzhou, Guangdong Province, China) for his help with specimen photography. The authors are also indebted to the editor and anonymous reviewers for their suggestions in improving the manuscript.








