


Introduction
Nosema bombycis Nägeli (Microsporidia: Nosematidae), Bombyx mori nucleopolyhedrovirus (Baculoviridae: genus Alphabaculovirus) (NPV), and Bombyx mori densovirus (Parvoviridae: genus Iteravirus) (DNV), which cause pebrine, grasserie, and flacherie disease, respectively, are the main causes of reduced cocoon yield and quality in the sericulture industry. Due to a lack of effective means for controlling these diseases, the early detection of these pathogenic microorganisms is important. The development of accurate and sensitive diagnostic tools is essential to prevent the spread of diseases caused by these microorganisms and reduce economic losses. Microscopy is widely used for the detection of silkworm (Bombyx mori (Linnaeus); Lepidoptera: Bombycidae) pathogens but is limited by the specificity and sensitivity of the approach, as well as difficulties in achieving early detection. Furthermore, electron microscopy is necessary for the detection of DNV. An immunoassay is another traditional detection technique that has been extensively used for the development of monoclonal and polyclonal antibodies against microsporidia (Aldras et al. Reference Aldras, Orenstein, Kotler, Shadduck and Didier1994; Shamim et al. Reference Shamim, Ghosh, Baig, Nataraju, Datta and Gupta1997), NPV (Thorne et al. Reference Thorne, Otvos, Conder and Levin2007; Du et al. Reference Du, Zhang, Fu, Chen and Xiao2012; Wangman et al. Reference Wangman, Longyant, Chaivisuthangkura, Sridulyakul, Rukpratanporn and Sithigorngul2012), and DNV (Wangman et al. Reference Wangman, Longyant, Chaivisuthangkura, Sridulyakul, Rukpratanporn and Sithigorngul2009) that infect silkworms and other insects and shrimps. However, the low efficiency and low accuracy of these techniques preclude the routine application of such immunological approaches in the field. In contrast, polymerase chain reaction (PCR)-based assays have undergone rapid development, and the use of this technique has become increasingly prevalent due to its simplicity, speed, specificity, and sensitivity. Some PCR-based approaches have been attempted for the detection of N. bombycis, NPV, and DNV, and some of these assays are well established (Kawakami et al. Reference Kawakami, Iwano, Hatakeyama, Inoue and Canning2001; Chaivisuthangkura et al. Reference Chaivisuthangkura, Srisuk, Rukpratanporn, Longyant, Sridulyakul and Sithigorngul2009; Nimitphak et al. Reference Nimitphak, Meemetta, Arunrut, Senapin and Kiatpathomchai2010; Pan et al. Reference Pan, Gong, Zheng, Guo and Shen2011). In addition, several multiprimer PCRs were developed for the detection of microsporidian infections in B. mori (Hatakeyama and Hayasaka Reference Hatakeyama and Hayasaka2001, Reference Hatakeyama and Hayasaka2003). Real-time PCR, combined with fluorescent detection probe technology, is a simpler, more sensitive, and faster technique than PCR and has been more frequently applied for early pathogen detection. Burgher-MacLellan et al. (Reference Burgher-MacLellan, Williams, Shutler, MacKenzie and Rogers2010) established a duplex real-time PCR assay for the detection of Nosema apis and N. ceranae in Apis mellifera; additionally, Panichareon et al. (Reference Panichareon, Khawsak, Deesukon and Sukhumsirichart2011) developed a multiplex real-time PCR assay and high-resolution melting analysis to simultaneously detect white spot syndrome virus, yellow-head virus, and Penaeus monodon densovirus in Penaeidae (Decapoda) shrimp. Ravikumar et al. (Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011) established a multiplex PCR assay for the detection of microsporidia, nucleopolyhedrovirus, and densovirus simultaneously in silkworms, but the authors did not confirm the efficacy of this approach with artificially inoculated samples. Therefore, the aim of this study was to develop a multiplex real-time PCR method for the early detection of N. bombycis, nucleopolyhedrovirus, and densovirus simultaneously in B. mori.
Materials and methods
Insects, pathogens, and infections
Bombyx mori (Qiufeng strain) larvae were reared on mulberry (Morus Linnaeus; Moraceae) leaves at 25 °C with a 12-hour light and 12-hour dark cycle. The fifth instars (day 1) were reared on mulberry leaves pre-coated with N. bombycis (105 spores/larva), BmNPV (106 virus particles/larva), or BmDNV (Zhenjiang strain). Bombyx mori densovirus (Parvoviridae: genus Iteravirus) infections were performed as previously described (Ravikumar et al. Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011). Midgut tissues and whole larvae were harvested at 24, 48, 72, 96, 120, and 144 hours for DNA extraction after spores and viruses feeding. At 144 hours post inoculation, the mortality rate for BmNPV-infected larvae was 100%; therefore, no samples were collected at this time point.
Several other common pathogens of the silkworm, Bombyx mori cypovirus 1 (BmCPV-1, Suzhou strain), Bombyx mori infectious flacherie virus (BmIFV, Tongxiang strain), Bacillus thuringiensis Berliner (Bacillaceae) (Bt), Beauveria bassiana (Balsamo-Crivelli) Vuillemin (Clavicipitaceae) (Bb), and Nomuraea rileyi (Farlow) Samson (Clavicipitaceae) (Nr), were also used for the primers and probes-specific tests.
DNA extraction
DNA for the PCR experiments was extracted from pure germinated N. bombycis spores, viral particles (BmNPV and BmDNV, Zhenjiang strain), bacteria (Bt), and fungi (Bb and Nr) using a DNA Purification System (FF3750; Promega, Madison, Wisconsin, United States of America) according to the manufacturer’s instructions.
DNA for the artificially inoculated sample test was extracted either from 20 mg of midgut tissue or from 100 mg of whole larva tissue using the DNA Purification System (FF3750). For both sample tissues, at least five infected silkworm larvae were sampled at each time point. Before the DNA extractions, the midgut and whole larvae were pulverised in liquid nitrogen. In addition, the tissue from N. bombycis-infected larvae was homogenised a second time before DNA extraction using a FastPrep®-24 Instrument (MP Biomedicals, Santa Ana, California, United States of America) with glass beads (Φ 425–600 μm; Sigma, St. Louis, Missouri, United States of America) six times at 5500 rpm for 20 seconds (Cai et al. Reference Cai, He, He, Qiu, Li and He2011). DNA templates from untreated (uninfected) silkworms and mulberry leaves were used as controls.
In the artificially inoculated sample tests, mixed suspensions of DNA extracted from individuals infected with a single pathogen type were used as templates in duplex and multiplex PCR assays.
RNA extraction, complementary DNA synthesis
Total RNA from viral particles, BmCPV (Suzhou strain) and BmIFV (Tongxiang strain) was extracted by using UNIQ-10 (Sangon, Shanghai, China) according to the manufacturer’s instructions. The RNA samples were further treated with DNase I (RNase free) (TaKaRa, Dalian, China) to remove any contaminating DNA following the manufacturer’s instructions. One microgram of total RNA was reverse transcribed in a 20 μL reaction mixture with a First-Strand cDNA Synthesis Kit according to the manufacture’s instruction (TaKaRa).
Primer and probe design
The primer and probe sequences for N. bombycis, BmNPV, and BmDNV were selected from small subunit ribosomal RNA (SSUrRNA), a DNA polymerase gene and a putative structural protein gene, respectively. All the primers and probes were designed using the online Primer3 internet-based interface (http://frodo.wi.mit.edu), manufactured by TaKaRa, and checked by using a basic local alignment search tool (BLAST) (http://www.ncbi.hlm.nih.gov/BLAST/) against the GenBank database for significant similarities to sequences of non-target organisms (Supplementary Table S1). In addition, the potential PCR target sequences were also checked with the BLAST program to further confirm the specificities of the primers and probes.
Polymerase chain reaction, cloning, and sequencing
Before performing the multiplex real-time PCR experiments, individual PCRs for each pathogen were conducted in the following reaction mixture: 2.5 μL PCR ExBuff (TaKaRa), 2 μL dNTP (2.5 pmol/μL), 2 μL MgCl2 (25 pmol/μL), 0.5 μL of each primer (10 pmol/μL, Table 1), 0.125 μL ExTaq (5 U/μL), 4 μl DNA, and 13.375 μL H2O in a total volume of 25 μL. The PCRs were performed on a MJ Research PTC-200 PCR system according to the following protocol: initial activation at 94 °C for five minutes, 40 cycles at 94 °C for 30 seconds, 60 °C for 60 seconds, and 72 °C for 45 seconds, and a final extension at 72 °C for 10 minutes. The PCR products were then analysed by 1% gel electrophoresis. Next, the three independent PCR amplifications were cloned into PMD18-T vectors (TaKaRa) and sequenced by TaKaRa. The results of the sequencing were checked using BLAST searches. The individual PCRs were also used for primer-specific tests.
Table 1 Detection of Nosema bombycis (Nb), Bombyx mori nucleopolyhedrovirus, and Bombyx mori densovirus in the corresponding plasmids by real-time polymerase chain reaction (PCR).

Notes:
a Concentration of each corresponding plasmid. The initial concentrations of the three plasmids were uniformed into 8.5×108 copies/μL (2.5 ng/μL). Whether in simplex, duplex, or multiplex PCR, 1 μL of each corresponding DNA template (plasmid) was used. In both the duplex PCR and multiplex PCR, the primer/probe sets between different pathogens were mixed in the ratio of 1:1 and 1:1:1.
NPV, nucleopolyhedrovirus; DNV, densovirus; +, positive result of the detection; −, no positive signal after 40 PCR cycles or the results out of the detection limits; N, negative controls.
Real-time polymerase chain reaction
The simplex (individual) real-time PCR mixtures contained 10 μL Premix ExTaq (TaKaRa), 0.4 μL of each primer (10 pmol/μL), 0.4 μL of probe (10 pmol/μL), 1 μL of each template DNA, and water in a total volume of 20 μL. The individual real-time PCRs were also used for primer-and-probe specific tests.
The duplex and multiplex real-time PCRs were conducted as above in 20 μL reaction mixtures. By adjusting the volumes of each primer and probe according to a certain proportion, the total volumes of the primers (in total, 0.4 μL of forward primers and 0.4 μL of reverse primers) and probes (in total, 0.4 μL) remained unchanged in the mixtures. A mixture of 1 μL of each corresponding DNA template extracted from individuals infected with a single pathogen type was used in these reactions.
Real-time PCR was performed with an initial denaturation of 95 °C for 10 seconds, followed by 40 cycles at 95 °C for five seconds and 60 °C for 23 seconds (LightCycler 480; Roche, Indianapolis, Indiana, United States of America). All the real-time PCR experiments were repeated at least three times with similar results observed.
Although the optical filters for each detection channel of the LightCycler 480 system are optimised for the different emission maxima of the fluorescent dyes, some residual crosstalk between the channels is observed. A colour compensation file was used to correct this crosstalk colour compensation file was generated according to the Roche manual, using the following conditions: (1) 95 °C for 10 seconds, (2) 40 cycles of 95 °C for five seconds and 60 °C for 23 seconds, (3) 95 °C for 10 seconds, 40 °C for 30 seconds, 95 °C for “continuous”, and 40 °C for one second. The real-time PCR mixtures for the colour compensations were the same as those described above for the simplex real-time PCR mixtures. After each multiplex/duplex real-time PCR experiment, the corresponding colour compensation file that was generated was consulted to ensure that no false positives were generated by crosstalk.
Construction of stand curve
To evaluate the detection limit and linear range of the species-specific PCR systems, 10-fold dilution series (in water) of plasmid DNA were used, in simplex, duplex, and multiplex PCR systems. The amplification curve was created by plotting the logarithm of the DNA concentration against the C t values (Pegels et al. Reference Pegels, López-Calleja, García, Martín and González2013). The correlation between the variables, C t and concentration is semilogarithmic: C t =blog[ ]+a, where b is the slope and a the intercept.
Results
Verification of specificity for primers
After the individual PCRs and real-time PCRs for the single or mixed pathogen experiments, each primer pair (and probe) successfully produced the desired PCR product without cross reacting with DNA from the other pathogens or the controls (Supplementary Fig. S1 and Supplementary Table S2). To further confirm the specificities of the primer sets, the respective PCR products were sequenced and analysed by BLAST. The results of the BLASTn are presented in Supplementary Table S3. The 66 base pairs product showed 100% SSU-rRNA homology to N. bombycis, Nosema species, and an additional microsporidian species. The 268 base pairs sequence showed 99–100% polyhedrin gene homology to a number of isolates of NPVs of B. mori and B. mandarina (Moore) and 93–96% homology to NPVs of Plutella xylostella (Linnaeus) (Lepidoptera: Plutellidae), Autographa californica (Speyer) (Lepidoptera: Noctuidae), and Rachiplusia ou (Guenée) (Lepidoptera: Noctuidae). The 171 base pairs sequence showed 100% sequence identity to Bombyx mori DNV Zhenjiang and Bombyx mori DNV3 (VD1 isolate) and 99% identity to Bombyx mori DNV Zhenjiang segment VD1 and a parvo-like virus. These results demonstrated that the three primer pairs were sufficiently specific for N. bombycis, Bombyx mori nucleopolyhedrovirus, and Bombyx mori densovirus, respectively.
The following experiments were all completed using real-time PCR; therefore, in the following passages of results, the phrase “real-time PCR” is abbreviated as “PCR”.
Verification of sensitivity for primers and probes
The sensitivities of the three primer/probe sets were evaluated by PCR using serial dilutions of plasmids (pMD18-T vector) containing the respective PCR products as templates (Table 1, Supplementary Figs. S2–S4). As shown in the Supplementary Fig. S2, linearity was observed for the three simplex PCR systems (N. bombycis, BmNPV, and BmDNV) over 6, 7, and 5 orders of magnitude, respectively. Correspondingly, as Table 3 shows, the detection limits in the simplex PCR experiments for the N. bombycis, BmNPV, and BmDNV plasmids were 8.5×103 copies, 8.5×102 copies, and 8.5×104 copies, respectively. The values also refer to the lowest concentration of target plasmid DNA producing reliably detected fluorescence signals. In the duplex PCRs of the different combinations, the sensitivities of the primers for N. bombycis and BmDNV were the same as those in the simplex reactions (Supplementary Figs. S3A–S3C); however, for BmNPV, the sensitivity was lower at 8.5×103 copies (Supplementary Figs. S3B–S3C). In the triplex (multiplex) PCRs, the detection limits for the N. bombycis, BmNPV, and BmDNV plasmids were 8.5×103 copies, 8.5×103 copies, and 8.5×105 copies, respectively (Supplementary Fig. S4A). Only the primer/probe set for the detection of N. bombycis demonstrated the same efficiency both in the simplex and multiplex PCRs. However, the sensitivities of the primer/probe sets for BmNPV and BmDNV were reduced with the increasing complexity of the PCR mixtures.
To improve the detection limit of BmDNV in the multiplex PCR, the proportions of different primer pairs and probes were optimised (Table 2). After increasing the ratio (in volume) of the primer/probe set for BmDNV, the detection sensitivity for BmDNV improved significantly (Table 2, N. bombycis:BmNPV:BmDNV=2:2:3; 1:1:2; 1:1:3) compared with the sensitivity obtained with the initial primer/probe ratio (N. bombycis:BmNPV:BmDNV=1:1:1); however, only by using a ratio of 2:2:3 (N. bombycis:BmNPV:BmDNV) was it possible to improve the detection capability of BmDNV without compromising the ability to detect the other two pathogens. By increasing the proportions of the primer/probe sets for both BmDNV and BmNPV in the multiplex PCRs (Table 2, N. bombycis:BmNPV:BmDNV=2:3:3; 1:2:2), the detection sensitivities for BmDNV and BmNPV improved to 8.5×104 copies and 8.5×102 copies, respectively; simultaneously, however, the detection sensitivity for N. bombycis decreased to 8.5×104 copies. However, while increasing the proportions for both BmDNV and N. bombycis (Table 2, N. bombycis:BmNPV:BmDNV=3:2:3; 2:1:2), the sensitivities decreased both for N. bombycis and BmNPV. Thus, the ratio of 2:2:3 (N. bombycis:BmNPV:BmDNV) for the three primer/probe sets in the multiplex PCRs was the optimal ratio, and the detection limits for the plasmids of N. bombycis and BmNPV were both 8.5×103 copies and for BmDNV was 8.5×104 copies (Supplementary Fig. S4B).
Table 2 Multiplex real-time polymerase chain reaction (PCR) optimisation by adjusting the proportion of primer/probe sets of different pathogens, using the plasmids as the template DNA.
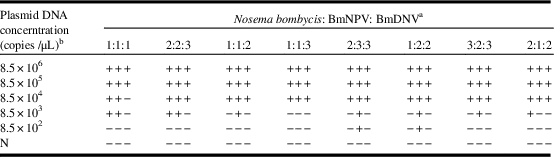
Notes:
a In the multiplex PCR, the proportions of primer/probe sets for detection different pathogens.
b Concentration of each plasmid. In multiplex PCR, 1 μL of each corresponding DNA template (plasmid) was used.
BmNPV, Bombyx mori nucleopolyhedrovirus; BmDNV, Bombyx mori densovirus; NPV, nucleopolyhedrovirus; DNV, densovirus;+: positive result of the detection; −, no positive signal after 40 PCR cycles or the results out of the detection limits; N, negative controls.
Detection of Nosema bombycis, Bombyx mori nucleopolyhedrovirus, and Bombyx mori densovirus in artificially inoculated silkworms
The PCR assay was also used for the early detection of the three pathogens in artificially inoculated silkworms. The BmDNV virus could be detected within 24 hours post inoculation from all the simplex, duplex, and multiplex PCR (N. bombycis:BmNPV:BmDNV=2:2:3) assays using template DNA isolated from either the whole body or the midgut of the infected silkworms (Tables 3–4). Bombyx mori nucleopolyhedrovirus (Baculoviridae: genus Alphabaculovirus) could not be detected at 24 hours post inoculation by multiplex PCR using DNA extracted from the whole body samples but BmNPV-specific amplicons were detected at 48 hours post inoculation. However, the results for the detection of N. bombycis varied. When the DNA was obtained from the whole body, N. bombycis was only detected in samples by day 6 post inoculation by simplex and duplex PCRs; however, no positive result was achieved using multiplex PCR for the duration of the experiment (Table 3). When DNA was obtained from the midgut, the simplex and duplex PCRs showed early detection of N. bombycis on day 3 and day 5 post inoculation onwards, respectively; the detection of N. bombycis from DNA from the whole body occurred three and one day(s) later for the simplex and duplex PCRs, respectively, compared with the findings described for midgut DNA (Table 4). In the multiplex PCR experiments, N. bombycis remained undetectable in the DNA from the midgut throughout the timecourse (Table 4).
Table 3 Early detection of Nosema bombycis (Nb), Bombyx mori nucleopolyhedrovirus, and Bombyx mori densovirus by real-time polymerase chain reaction (PCR), with the template DNA isolated from infected silkworm whole body.

Notes: In the duplex PCR, the primer/probe sets between different pathogens were mixed in the ratio of 1:1, and in the multiplex real-time PCR, the ratio of 2:2:3 (N. bombycis:BmNPV:BmDNV) was used. Whether in simplex, duplex, or multiplex PCR, 1 μL of each corresponding DNA template extracted from silkworm whole bodies infected with a single pathogen type was used.
a No sample.
NPV, nucleopolyhedrovirus; DNV, densovirus; +, positive result of the detection; −, no positive signal after 40 PCR cycles; N, negative controls.
Table 4 Early detection of Nosema bombycis (Nb), Bombyx mori nucleopolyhedrovirus, and Bombyx mori densovirus by real-time polymerase chain reaction (PCR), with the template DNA isolated from infected silkworm midgut.

Notes: In the duplex PCR, the primer/probe sets between different pathogens were mixed in the ratio of 1:1, and in the multiplex real-time PCR, the ratio of 2:2:3 (N. bombycis:BmNPV:BmDNV) was used. Whether in simplex, duplex, or multiplex PCR, 1 μL of each corresponding DNA template extracted from silkworm midguts infected with a single pathogen type was used.
a No sample.
NPV, nucleopolyhedrovirus; DNV, densovirus; +, positive result of the detection; −, no positive signal after 40 PCR cycles; N, negative controls.
As the growth rate of the virus was significantly faster than the rate of spore production in the host cells, the large amount of viral DNA may have inhibited the amplification of the spore DNA in the multiplex PCR experiments, which may have led to the negative results for the detection of N. bombycis throughout the experiment. In addition, the DNA templates isolated from the virus-infected silkworms were diluted. As shown in Table 5, these dilutions enabled the multiplex PCRs to detect N. bombycis on day 6 post inoculation in the DNA samples from both the whole body and the midgut. According to the results of the plasmid tests, the primer/probe sets for BmDNV showed less efficiency compared with the sets for N. bombycis and BmNPV (Table 1). However, in the artificially inoculated silkworm test, BmDNV was detected on day 1 post inoculation in the multiplex PCR, even using samples that were diluted by 10-fold. This detection occurred significantly earlier than the detection of BmNPV (Table 5, column II), which may be due to the differences in the initial inoculation amounts of the viruses and their growth rates.
Table 5 Optimisation of the multiplex real-time polymerase chain reaction (PCR) assay by adjusting the concentration of template DNA.
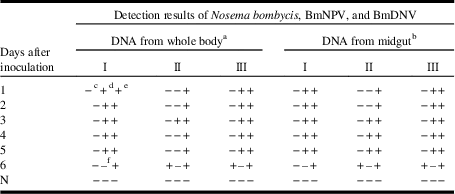
Notes: In the multiplex real-time PCR, the ratio of 1:1:1 (N. bombycis:BmNPV:BmDNV) for the three primer/probe sets was used.
a,b One microlitre of each corresponding DNA template (diluted or not diluted) extracted from individuals infected with a single pathogen type was used.
c,d,e Columns recording the detection results of N. bombycis, BmNPV, and BmDNV, respectively.
f No sample.
I: all the DNA samples were not diluted; II: DNA samples extracted from the silkworms infected with N. bombycis were not diluted, but infected with BmNPV or BmDNV were 10-fold diluted; III: DNA samples extracted from the silkworms infected with N. bombycis were not diluted, if infected with BmNPV were five-fold diluted, and if infected with BmDNV were 10-fold diluted; BmNPV, Bombyx mori nucleopolyhedrovirus; BmDNV, Bombyx mori densovirus; +, positive result of the detection; −, no positive signal after 40 PCR cycles; N, negative controls.
When detecting pathogens in naturally infected samples, the optimisation of template concentrations is not feasible for multiplex PCR assays. Therefore, to improve the detection limit for N. bombycis in the artificially inoculated samples, a series of volume ratios of the three primer/probe sets were evaluated; the original concentrations of the DNA templates were also used in these tests. In the whole body test, the multiplex PCR could detect N. bombycis on day 6 post inoculation when the ratio was changed from 1:1:1 to 3:2:2 (N. bombycis:BmNPV:BmDNV); however, even if the proportion of N. bombycis was further increased, earlier detection could not be obtained (Table 6). In contrast, when the volume of the primer/probe set for N. bombycis was increased, the detection sensitivities for BmNPV and BmDNV were reduced (Table 6). Similar results were observed in the midgut test, in which the ratio of 3:2:2 (N. bombycis:BmNPV:BmDNV) and 2:1:1 enabled the detection of N. bombycis on day 5 post inoculation; however, a further increase in N. bombycis was neither conducive to the detection of BmNPV and BmDNV, nor improved the detection sensitivity of N. bombycis (Table 7). For the detection of N. bombycis, there was at least a two-day delay using the multiplex PCR assay compared with the simplex PCR assay.
Table 6 Optimisation of the multiplex real-time polymerase chain reaction (PCR) assay by adjusting the proportions of primer/probe sets of different pathogens, with the template DNA isolated from the whole body of the infected silkworms.

Notes: In the multiplex PCR, a mixture of 1 μL of each corresponding DNA template extracted from individuals infected with a single pathogen type was used.
a,b,c Columns recording the detection results of N. bombycis, BmNPV, and BmDNV, respectively.
d No sample.
BmNPV, Bombyx mori nucleopolyhedrovirus; BmDNV, Bombyx mori densovirus; +, positive result of the detection; −, no positive signal after 40 PCR cycles; N, negative controls.
Table 7 Optimisation of the multiplex real-time polymerase chain reaction (PCR) assay by adjusting the proportion of primer/probe sets of different pathogens, with the template DNA isolated from the midgut of the infected silkworms.

Notes: In the multiplex PCR, a mixture of 1 μL of each corresponding DNA template extracted from individuals infected with a single pathogen type was used.
a,b,c Columns recording the detection results of N. bombycis, BmNPV, and BmDNV, respectively.
d No sample.
BmNPV, Bombyx mori nucleopolyhedrovirus; BmDNV, Bombyx mori densovirus; +, positive result of the detection; −, no positive signal after 40 PCR cycles; N, negative controls.
In the field, mixed infections with multiple pathogens are less likely to occur compared with single-pathogen infections (Ravikumar et al. Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011). Therefore, assuming that the field sample in this study was only infected with N. bombycis, the DNA isolated from the N. bombycis-infected silkworms was exclusively used in the triplex PCR assay. Without interference from the DNA of the other pathogens, and under the optimal volume ratio for the three primer/probe sets (3:2:2=N. bombycis:BmNPV:BmDNV), N. bombycis could be detected on day 3 post inoculation in the midgut test, as early as the detection of N. bombycis using simplex PCR (Table 8).
Table 8 Early detection of Nosema bombycis by triplex real-time polymerase chain reaction (PCR) assay, with the template DNA isolated from N. bombycis-infected silkworms.
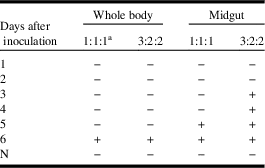
Notes:
a The ratio for the three primer/probe sets (N. bombycis:BmNPV:BmDNV) used in the multiplex real-time PCR.
BmNPV, Bombyx mori nucleopolyhedrovirus; BmDNV, Bombyx mori densovirus; +, positive result of the detection; −, no positive signal after 40 PCR cycles; N, negative controls.
Discussion
The multiplex real-time PCR-based identification of three important pathogens infecting silkworms represents a simple molecular approach for the specific and rapid detection of these species simultaneously. In a previous study, Ravikumar et al. (Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011) established a multiplex PCR assay for the detection of these three pathogens. After comparing these two methods, it appears that the real-time PCR assay, combined with fluorescent detection probe technology, is simpler and faster compared with the latter approach.
The primers and probes that we designed showed great specificity for the target pathogens. To ensure adequate specificities, all the primers and probes were checked by BLAST before being used; additionally, following the PCRs, the amplified fragments were sequenced. The BLAST results from the sequencing presented high degrees of sequence similarities to genes from the three target species (Supplementary Table S3). Similar to N. bombycis and BmNPV, many other microsporidia (Huang et al. Reference Huang, Pan, Zhu, Lin, Luo and Shi2011) and NPVs (Chen Reference Chen2008) can cross-infect B. mori; furthermore, these microsporidian spores and baculoviruses share significant homology in their nucleic acid sequences (Supplementary Table S3, Chen Reference Chen2008; Ravikumar et al. Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011); therefore, it is possible that our detection system could be used to detect these various other pathogens in B. mori or other silkworms, to highlight the usefulness of our study for the detection of such pathogens. However, due to a lack of available pathogens, the potential of our approach for the detection of such pathogens was not tested. Ravikumar et al. (Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011) reported the successful detection of microsporidia in tasar silkworms (Antheraea mylitta (Drury); Lepidoptera: Saturniidae) and DNV in eri silkworms (Samia cynthia (Drury); Lepidoptera: Saturniidae) using primer sets optimised for the detection of B. mori pathogens. Because the Zhenjiang strain was the only DNV variant we had available, the primers and probe for the detection of this pathogen were designed based on the BmDNV Zhenjiang sequence (Supplementary Table S1). The BLAST results of the primer pair, the probe (data not shown), and the PCR product (Supplementary Table S3) illustrate that, in addition to the Zhenjiang strain, BmDNV 3 is also likely to be detected by this assay. In the previous study by Ravikumar et al. (Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011) the primer pair “DNV 2 & DNV 3” were designed based on the sequences of DNV 2 and DNV 3 and could detect four DNV variants, including DNV 2, 3, 6, and Zhenjiang. However, the BLAST results of the primer pair “DNV 2 & DNV 3” show that only the DNV 3 and Zhenjiang sequences share 100% identity with both primers. Even if the PCR product of the primer set shares high similarity (97–99%) with the DNV 2 and 6 sequences, it remains unclear whether the primer set “DNV 2 & DNV 3” has the capability to detect DNV 2 or 6. Due to the minimal sequence homology that exists between the various BmDNVs, it may be challenging to design a universal primer pair that detects all variants of BmDNV.
For the detection of N. bombycis in artificially inoculated silkworms, we found that it was important to homogenise the sample first using the FastPrep®-24 Instrument. If this step was not performed, the PCR could not detect the pathogen at any point in the experiment. However, after six days post inoculation, spores could be observed in the midgut samples using the optical microscope (data not shown). The N. bombycis spore wall consists of an electron-dense, proteinaceous exospore and an electron-lucent endospore composed of chitin, which is helpful in maintaining the normal morphology and resisting adverse environmental conditions. The spore wall, however, increases the difficulty of extracting DNA from the spores (Cai et al. Reference Cai, He, He, Qiu, Li and He2011). Typically, the spores are subjected to polar filament discharge or liquid nitrogen freezing and grinding treatment before extraction, which breaks the spore walls and improves the DNA or protein yield (Xie Reference Xie2007; Pan et al. Reference Pan, Gong, Zheng, Guo and Shen2011). The recently developed method using a FastPrep agitator and glass beads has proven to be effective for DNA and protein extraction from microsporidian spores, fungi, and other microbial organisms with a thick and rigid outer wall (Fredricks et al. Reference Fredricks, Smith and Meier2005).
In the artificially inoculated silkworms test, the multiplex PCR enabled the early detection of BmNPV and BmDNV under optimal conditions on day 1 post inoculation in samples from both the whole body and midgut (Tables 6–7); for N. bombycis, however, positive results were not obtained until day 6 and 5 in the whole body and midgut, respectively (Tables 6–7). The multiplex PCR detected N. bombycis infection more efficiently with DNA templates exclusively from N. bombycis-infected silkworms (Table 8) than with a combination of templates from silkworms individually infected with the three pathogens. As shown in Table 8, the triplex PCR assay could detect N. bombycis on day 3 post inoculation with the individual DNA template, which was as early as the infection was detected using simplex PCR. In a previous report (Ravikumar et al. Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011), the PCR assay detected N. bombycis and BmDNV on day 1 post inoculation and BmNPV on day 2 post inoculation, in which the detection of N. bombycis occurred much earlier than in our study. However, in these two studies the samples were processed differently. In the study by Ravikumar et al. (Reference Ravikumar, Raje Urs, Vijaya Prakash, Rao and Vardhana2011), the DNA for analysis was isolated from 20 mg of host tissue mixed with spores, and the method (simplex or multiplex PCR) used in the assay was unclear. In our study, samples were obtained from either the whole body or midgut of the orally infected silkworms and analysed by multiplex real-time PCR. Therefore, the results of these studies cannot be compared. Using microscopy to identify the pathogens, N. bombycis was not observed until day 6 and was only observed in the midgut samples; BmNPV was observed by day 2 and 3 in the midgut and whole body, respectively (data not shown). These findings illustrate that this newly developed real-time PCR method is more sensitive and allows for earlier detection compared with traditional microscopic methods for the detection of these pathogens.
Under natural conditions, a number of pathogens infect silkworm larvae mainly via the oral route and then enter the body through the intestinal tract. Therefore, the intestines could be an ideal target for the early diagnosis of pathogens. Our findings support that the use of the midgut is superior to the whole body for the detection of silkworm pathogens (Tables 3–8).
In conclusion, the multiplex real-time PCR assay developed in this study is highly sensitive and specific for the simultaneous detection of N. bombycis, BmNPV, and BmDNV. Further evaluation of this newly developed method using artificially inoculated silkworms (B. mori) is necessary to verify its detection efficacy, which would greatly facilitate the early detection of these three major pathogens and aid in reducing crop loss in the sericulture industry.
Acknowledgements
This work was supported by grants from the General Administration of Quality Supervision, Inspection, and Quarantine of the People’s Republic of China (2007IK017) and the Science Technology Department of Zhejiang Province (2010C12005-2).
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.4039/tce.2016.54.









