


Introduction
The application in recent years of increasingly accurate and precise absolute dating methods for Palaeolithic cave art has placed our understanding of the timing and nature of its origins on a firmer foundation. Although the majority of examples remain to be dated independently, the increasing number of dates for cave art—particularly in Spain—have now revealed an antiquity far older than has been accepted hitherto, as well as a thematic development previously unknown. Appearing in Western Europe in non-figurative form >64,000 bp (Hoffman et al. Reference Hoffmann, Standish and García-Diez2018a) and in Indonesia by at least 40,000 bp, ‘cave art’ currently constitutes the earliest known example of art in the world; and the presence of animal depictions by at least 37,000 bp renders it the oldest figurative art in the world. Overall, cave art was produced—probably in fits and starts—by two human species, Homo neanderthalensis and Homo sapiens, and endured until around 13,000 years ago. It therefore spans a considerable portion of the known chronology of human artistic endeavour. Given this longevity, several of its defining features are remarkable, which distinguish the origins of art (and its greater [pre]history) from art of later prehistory onwards: 1) an early phase of non-figurative, peripersonal art created directly with the hands; 2) from ~37,000 years ago the overwhelming thematic dominance of large wild herbivores and carnivores in the figurative repertoire; 3) the relative absence of humanoids and absolute absence of clearly/unambiguously identifiable humans; 4) a lack of thematic diversity, either across space or time, in that large herbivores and carnivores remained the focus of concern (though there were fewer carnivores depicted during the late Upper Palaeolithic); 5) the rarity (or arguably, absence) of composed ‘scenes’ that one could describe as ‘portraits’ or ‘landscapes’ beyond the occasional use of natural ridges and other features as a ground (e.g. in Lascaux's Hall of the Bulls). While specialists no longer believe in the validity of selective and untestable ‘umbrella’ hypotheses that purport to ‘explain’ Palaeolithic ‘art’ (see a useful critique in Bahn Reference Bahn2016), or prioritize western notions of art, we may at least attempt to understand why art evolved in this peculiar way.
One of the interesting and provocative features of Upper Palaeolithic (UP) depictions concerns the incorporation of the cave's natural features into the portrayal of animals. This commonly involved the use of the shape of cracks, concavities and convexities of cave walls and ceilings as elements of the animal, notably serving as dorsal lines, chests and legs, but also seems to have included the projection of shadows (Groenen Reference Groenen2000; Pettitt Reference Pettitt, Dowd and Hensey2016; Pettitt et al. Reference Pettitt, Sakamoto, Lelushko, Papadopoulos and Moyes2017). Although the inclusion of natural features has been well documented for some time, with the exception of Ogawa (Reference Ogawa, Heyd and Clegg2005; Reference Ogawa and Clottes2012) and Brot (Reference Brot2010) it has largely been listed simply as a peculiar characteristic of Palaeolithic art to the extent that its potential for understanding the appearance and development of figurative art has not been fully explored. Moreover, despite Hodgson (Reference Hodgson2003a,b; Reference Hodgson2006a,b; Reference Hodgson2008; Reference Hodgson, Sachs-Hombach and Schirra2013b) and Alpert (Reference Alpert2009) having drawn attention to the phenomenon in relation to visual science, it has still not been sufficiently investigated in the present context (see also Dobrez Reference Dobrez2010–11; Dobrez & Dobrez Reference Dobrez and Dobrez2013; Halverson Reference Halverson1992a,Reference Halversonb; Hodgson Reference Hodgson2000a,Reference Hodgsonb; Reference Hodgson2003a,b; Reference Hodgson2006a,b; Reference Hodgson2008; Reference Hodgson2013a,b; Hodgson & Helvenston Reference Hodgson and Helvenston2006a; Hodgson & Watson Reference Hodgson and Watson2015; Hudson Reference Hudson1998; Kennedy & Silver Reference Kennedy and Silver1974).
We argue here that the deliberate association of non-figurative peripersonal art with natural features of caves and the inclusion of such in animal depictions may not simply be another of the ‘oddities’ of Palaeolithic art, but may prove critical to understanding why ‘art’ arose in the first instance. In order to explore this issue we will use the emerging chronology of cave art (which emerges early relative to portable art) to examine the phenomenon in depth from the perspective of perceptual psychology and the neuroscience of vision. We first examine the role of parietal handmarks (by which we refer to both positive hand prints and the far more common negative hand stencils) as a precursor to the later (younger) animal depictions. We then demonstrate how the suggestive natural features of the cave may have facilitated the representation of animals. We subsequently present and explore a falsifiable hypothesis that handmarks, along with related antecedents such as finger dots and blown discs, provided a precursor to animal depictions which, together with the hunter's hyperactive visual system for detecting fauna and the suggestive features and evocative environment of the cave, led to the depiction of animals and, thus, served as a stimulus for the appearance of figurative art. We recruit a number of concepts from perceptual psychology and visual neuroscience to explore this hypothesis.
Parietal handmarks
In terms of the minimum ages provided by U-Th dating of overlying calcite flowstones (e.g. Hoffmann et al. Reference Hoffmann, Pike, García-Diez, Pettitt and Zilhão2016; Reference Hoffmann, Standish and García-Diez2018a; Pike et al. Reference Pike, Hoffmann, Pettitt, García-Diez and Zilhão2017), handmarks and finger dots currently appear to be the oldest tangible examples of art, apparently predating the depictions of animals in Palaeolithic Europe and Australasia by at least several thousand years. In Europe, minimum ages of 40,000 bp have been obtained from the Great Panel of Hands in El Castillo Cave (Cantabria, Spain), located in proximity to younger animal depictions, and minimum ages of 64,000 bp have been obtained for colour washes on stalactites in Ardales cave (Andalusia), hand stencils in Maltravieso cave (Extremadura) and a scalariform motif comprised of small dots in La Pasiega cave (Cantabria) (Hoffmann et al. Reference Hoffmann, Standish and García-Diez2018a). In Sulawesi, (Indonesia), a minimum age of 39,900 bp has been obtained for a handmark, and a minimum age of 35,400 bp for the representation of a babirusa (‘pig-deer’) (Aubert et al. Reference Aubert, Brumm and Ramli2014), the latter at present being consistent with a later emergence of figurative art ~37,000 bp. Recent critical evaluations of the available chronology of handmarks (García-Diez et al. Reference García-Diez, Garrido, Hoffmann, Pettitt, Pike and Zilhão2015; Pettitt et al. Reference Pettitt, Arias, García-Diez, Bueno-Ramírez and Bahn2015) reveal that, far from being broadly Mid Upper Palaeolithic (MUP) in age, as is usually assumed, they are early MUP at their youngest, and in most dated cases, Early Upper Palaeolithic (or older). By contrast, all dated examples of figurative art have younger minimum ages, for example in Altamira (García-Diez et al. Reference García-Diez, Hoffmann, Zilhão, de las Heras, Lasheras, Montes and Pike2013) and Cosquer (Clottes Reference Clottes, Chippindale and Taçon2000; Clottes et al. Reference Clottes, Courtin and Vanrell2015) as well as at Sulawesi, and Chauvet, taking arguments for an Aurignacian age of ~35,000 cal. bp for its parietal art based on five charcoal samples deriving from three hearths on its floor at face value (Quiles et al. Reference Quiles, Valladas and Geneste2014), whether or not one agrees that its art is well dated (cf. Pettitt & Bahn Reference Pettitt and Bahn2015). Similarly, in the rock art of Arnhem Land, Australia—long thought to be relatively ancient—handmarks pre-date animal depictions, whatever its specific chronology (Chippindale & Taçon Reference Chippindale, Taçon, Chippindale and Taçon2000). In addition, Taçon et al. (Reference Taçon, Tan and O'Connor2014, 1061) note that handprints
… appear very early in rock art sequences wherever they are found (as with Europe and Egypt), but continued to be produced for thousands of years. In Europe and Sulawesi, the oldest hand stencils we know of were made close to the time when modern humans settled those areas …
Chaloupka (Reference Chaloupka and Ucko1977; Reference Chaloupka1984a,Reference Chaloupkab; Reference Chaloupka1993a,Reference Chaloupka, Lorblanchet and Bahnb), as well as Taçon and Chippindale (Reference Taçon and Chippindale1994), make essentially the same point (see Bednarik Reference Bednarik2013 for a critical assessment of this claim, which can now be rejected on the basis of the chronological data noted above). Handmarks, therefore, stand as perhaps the most ancient and universal type of representation, at least in terms of our current understanding of non-perishable art preserved in the archaeological record (Hodgson Reference Hodgson2006a); perhaps because they are a relatively straightforward and accessible means of signalling human presence (Hodgson Reference Hodgson2006a) in close (peripersonal) proximity to the body itself. But how did handmarks originate?
It is not difficult to see how prosaic activities may have left accidental handmarks on natural surfaces. Butchery of animal carcasses would have rapidly covered the butchers’ hands in blood, which may also have led to accidentally made handprints (Hodgson Reference Hodgson2013a). When ochre came to be used—for which there is evidence in Europe by 250,000 bp (as a liquid paint: Roebroeks et al. Reference Roebroeks, Sier, Kellberg Nielsen, de Loecker, Parés, Arps and Mücher2012) and Africa by 300,000 bp (as crayons: Barham Reference Barham2002)—handmarks could have been created fortuitously in deliberately produced pigment. In caves, the act of navigation can leave hand (and for that matter, foot) impressions on surfaces, and, in this respect, handprints of Palaeolithic age still survive in cave earth and mondmilch (e.g. Clottes et al. Reference Clottes, Courtin and Vanrell2005; Pastoors et al. Reference Pastoors, Lenssen-Erz and Ciquae2015). We may assume, therefore, that handmarks most likely originated accidentally in prosaic, messy activities, and subsequently became meaningfully associated with ochre, perhaps in the context of the marking of the body. This would, however, explain only the origin of positive hand prints; the overwhelming majority of handmarks in Palaeolithic art are, by contrast, negative stencils. Although these could have been accidentally created, this seems less likely and, conceptually, they require more complex thought than pressing a pigment-covered hand onto a surface.
There is a further step-change from simply printing or projecting pigment round the fingers or hand to the actual portrayal of animals (Hodgson Reference Hodgson2013a). In this regard, it is notable that some early depictions of animals are composed of finger dots or palm prints, and furthermore that many examples of figurative art were produced by spitting pigment and using the hands as guide templates. Whatever the specifics of the evolution from accidental to deliberate hand marking with pigment to the intentional creation of animal outlines using pigment and hands, we can assume that, at some point in time, handprints were ‘exapted’ for socio-cultural purposes; that is to say, deliberately produced to carry meaning (Dobrez Reference Dobrez2013).
Gravitating from relatively simple handmarks to the first animal depictions may have been conceptually challenging, whatever the time lag between the two. Nonetheless, the ‘trick’ of depicting a hand—in the sense that one thing can stand for another—may have provided the conceptual platform from which the more sophisticated animal depictions could be fabricated. As Pettitt et al. (Reference Pettitt, Arias, García-Diez, Bueno-Ramírez and Bahn2015, 40) articulate:
Is it possible that their very nature at the borders of the figurative and non-figurative, and their apparent appearance just as figurative art is emerging in European caves, suggest they played a role in the recognition that things could be figured in art? If the hand could be represented in outline, then why not animals?
What kind of mechanism could have served as a bridge to link the two depictive strategies? We propose that such a mechanism can be found in the suggestive features of rock surfaces and caves.
Role of natural features
It is widely acknowledged that caves containing Palaeolithic art abound with evocative natural features, which were often incorporated into the art itself. In fact, early rock art incorporating natural features can be found in many parts of the world (e.g. Keyser & Poetschat Reference Keyser, Poetschat, Chippindale and Nash2004; Taçon et al. Reference Taçon, Gang and Decong2010). Such features—elongated cracks, bosses, chimneys and the like—invariably simulate the contours of animals, particularly the cervico-dorsal line. It is, however, unlikely that suggestive natural features alone led to the ability to produce the fully fledged intentional animal depictions. Why should they necessarily be interpreted as parts of animals? Some further element that enhanced the saliency of such natural structures and induced the graphic embellishment seems necessary. Hodgson (Reference Hodgson2003b: Reference Hodgson2008) proposed that the visual imagery of the hunter was especially tuned or primed to detect the salient features of animals for several reasons:
• The critical importance of animals to the evolutionary survival of members of the genus Homo leading to an innately defined area of the visual brain specialized for detecting and encoding animal outlines;
• The existence of a pre-attentive (i.e. implicit preconscious) pathway arising from the retina projecting through the thalamus directly to the amygdala, that affords an instantaneous reaction to decisive diagnostic cues signalling the presence of an animal;
• Visual imagery selectively facilitates perception, so that if subjects imagine a form they will be more likely to detect a degraded version of an object, such as an animal, when presented with alternatives (Kosslyn & Shin Reference Kosslyn, Shin, Farah and Ratcliff1994).
We know that the human brain is finely tuned to detect animals (Hodgson Reference Hodgson2013a and references therein). This is observable in certain perceptual phenomena including illusory lines/shapes, filling in, perceptual closure, line continuation, as well as visual hypotheses testing and projective imagery, which can be observed to various degrees in perceptual phenomena such as the Kanizsa Triangle, Street Figure, Dalmatian Dog test, Rubin's Face/Vase illusion, Rorschach Inkblot (Fig. 1) and the Perky Effect (Perky Reference Perky1910). The first three concern figure completion by mental filling-in of the missing areas, which is an automatic function of the visual brain that takes place in the early to intermediate visual system relating to predictive coding (Van de Cruys et al., Reference Van de Cruys, Evers, Van der Hallen, Van Eylen, Boets, de-Wit and Wagemans2014). The last three relate to visual hypotheses testing and projective imagery (the propensity to see imaginary forms in response to ambiguous material arrays). Interestingly, Alpert (Reference Alpert2012) mentions the relevance of the Rorschach Ink Blot (see also Hodgson Reference Hodgson2003b; Hodgson & Helvenston Reference Hodgson and Helvenston2006a) and Kanizsa Triangle to understanding some of the depictions of animals in Niaux Cave. Compared to non-human primates, humans seem to benefit from an enhanced ability to fill in missing lines/shapes that would have been useful for detecting predator and prey when hominins began to survive on the Savannah (Vyshedskiy Reference Vyshedskiy2014). The raised sensitivity of human perception to contours and edges is, therefore, a predetermined capacity of the visual system which ‘encourages’—and hence facilitates—the detection of critical and specific stimuli such as a partly hidden animal (Alcock Reference Alcock1998).

Figure 1. Top left to top right: Kanizsa Triangle, Street Figure. Middle left to middle right: Dalmatian Dog test, Rubin's Face/Vase illusion. Bottom: Rorschach inkblot.
Recent hypotheses on the effect of milder ‘altered states of consciousness’ (ASCs) on cave art have emphasized the importance of sleep deprivation, fatigue, hunger, prolonged concentration directed at one object, as well as raised levels of arousal stimulated by fear and threat, stress and anxiety levels and traumatic events (Hodgson Reference Hodgson2013a) in stimulating such ‘altered’ states. It is, of course, impossible to ascertain whether, and how frequently, such ‘ASCs’ were experienced, and particularly whether or not they influenced art. While we would want to distance ourselves from hypotheses about more extreme hallucinatory ASCs, we think it pertinent to note that anxiety, fear, tiredness, hunger and prolonged concentration could have potential effects on the human experience of the cave environment. One therefore does not need to invoke extreme ASCs. Thus, the milder states can lead to the misperception of objects, thereby increasing the likelihood of the misinterpretation of natural cave features, which are seen not just as cracks, protrusions or blemishes etc., but as suggestive of animals. We propose that it is reasonable to posit such a mechanism due to the following triggers:
• Caves are unfamiliar, inexplicable and potentially dangerous places where vigilance is essential.
• Caves were frequented by carnivores such as lions, bears and wolves; human visitors would have been on heightened awareness for such animals.
• Caves are replete with natural features that simulate animal outlines.
• The subdued lighting of caves caused internal visual imagery and visual hypothesis testing to intensify.
• Available light (lamps, torches, and occasionally small hearths) caused shadows to move, lending a sense of animacy to the natural features.
The cave as a simulated hunting environment
Beyond the naturally lit zones of cave mouths, where Palaeolithic groups often camped, there is no prosaic reason to enter deep caves. There are no resources to be obtained, with the exception of water, which probably harboured bacteria and other contaminants, and which could be obtained more safely from rivers, lakes and snow. Some caves may have been entered to procure ochre, calcium carbonate (Clottes et al. Reference Clottes, Courtin and Vanrell2005) or suchlike, although these are hardly commonplace activities. So why enter the disorienting and potentially dangerous environment of caves when there were no obvious quotidian advantages to be had? We may speculate that caves came to be imbued with meaning due to the very fact that they were inexplicable, mysterious, liminal places; but what, specifically, drew people into them and stimulated the production of art in their dark recesses? Most cave art—at least from the early Mid Upper Palaeolithic onwards—is situated in deep caves, away from natural light. With the attenuated lighting available in such dark zones it would have been a challenge to distinguish artistic figures from the natural background, and in this sense the cave would simulate the conditions that a hunter normally faced during the act of hunting herbivorous prey while remaining alert to potential predators. This entails the need to detect and identify predator and prey at distance and among amorphous foliage, while accommodating changing atmospheric conditions such as in approaching darkness, all of which could be simulated in the cave by extremely low lighting and reduced colour variability and saturation, unfamiliar topography and orientation, navigational difficulty and potential danger.
The human visual system is poorly adapted to darkness, to the extent that humans become wary at night and, thus, they tend to associate darkness with danger due to the possibility of being easily deceived or a perceived lack of control that comes with night vision (Edensor Reference Edensor2013; Galinier et al. Reference Galinier, Becquelin and Bordin2010). Moreover, for many hunter-gatherer groups the night has negative associations (Schnepel & Ben-Ari Reference Schnepel and Ben-Ari2005). By contrast, predators, such as lions, benefit from excellent night vision. There is no reason to believe that the same wariness and association with danger did not apply to unfamiliar caves, where light conditions were too low to allow the rods in human eyes to adapt to low light (Pastoors & Weniger Reference Pastoors and Weniger2011), and the impenetrable darkness is often associated with other worlds (Arias & Ontañon Reference Arias, Ontañón, Bergsvik and Skeates2012). It is worth noting that, however frequently caves were explored, they would always be unfamiliar due to this impenetrable darkness and the impossibility of lighting all but small pools of light. Depictions were frequently created not on surfaces easy to access, but in difficult, unexpected locations that must have been actively sought and in which depictions are even today difficult to resolve. Why did artists ignore easy-to-access places where depictions were unambiguous, i.e. why was there so little preference for ‘least cost’ decision-making in cave art? This positioning is analogous to the way animals attempt to remain concealed in the natural environment, rather than taking a least-cost but visible route through it. One might say that each scenario corresponds to prospect-refuge theory as outlined by Appleton (Reference Appleton1975), in which humans prefer locales in which they can observe (prospect) but not be seen (refuge) and manifest this preference in art as well as the landscape. In many techniques of depiction, including the build-up of palm or finger discs or multiple striated lines, animals seem to have been deliberately rendered difficult to identify, a task made additionally problematic through subsequent superimpositions or partial obliteration. Why were images not made in clearly viewable positions and rendered in unambiguously identifiable ways? It may be significant that dots and stripes function as camouflage by both predators and prey, by deceiving perceptual systems, and if we approach the visual nature of cave art in terms of camouflage, it becomes clear that they possess an integral set of distracting characteristics. Such distractors will have posed a challenge to the perceptual acumen of Palaeolithic viewers by breaking up an animal's contour. As most animal depictions in cave art take the form of simple outlines (often of shallow engravings difficult to see when not in oblique light), and are often superimposed on uneven, blemished, stained or cracked surfaces, it could be said that they maximized the efficacy of such ‘distractors’ as a form of surrogate camouflage. One example of the use of spots as distractors in UP depictions is the spotted horses panel of Pech Merle, which some believe depicts real dappled horses rather than a convention of design (Pruvost et al. Reference Pruvost, Bellone and Benecke2011). An observation that has been used against this hypothesis is that the dots extend beyond the horse's body lines, although this is a relatively common trait of cave art and could be an attempt to introduce deliberate ambiguity into the depictions; when viewed in the subdued unstable lighting of caves the resulting effect is similar to the Dalmatian Dog figure, both of which present a challenge to the human visual system (Fig. 2).

Figure 2. Similarity between Dalmatian Dog Illusion and Pech Merle Horses (in subdued torchlight).
Darkness and caves
Beyond the immediate entrance area, the darkness of the deeper cave becomes increasingly intense where torches, lamps and hearths provide only limited illumination. To some extent, such darkness simulates the experience of the night, in that the glow of the campfire is surrounded by an enveloping blackness; outside this darkness comes the call of wild animals. In such circumstances human imagination can indulge in flights of fantasy regarding the interpretation of such sounds; a tendency to audial hypothesis testing that is reflected by the proclivity of the visual brain to engage in visual hypothesis testing. The latter is an adaptive mechanism for detecting predators and prey by ensuring that false positives (or false alarms: as an adaptation it is far better to respond to false alarms than to ignore potentially real ones) invariably elicit a response (known as adaptive conservatism). As Bednarik (Reference Bednarik2003a) noted, mistaking an exposed tree root in a forest for a snake is an example of visual ambiguity which, through projection, may have led early humans to realise one thing can stand for something else (but see below for how this is played out in archaeological terms). As vision is the most vital sense for encoding the environment, when this sense is hindered, not only do visual imagery and imagination increase, but other senses, such as touch, come into play. As a result, the viewer is obliged constantly to reformulate what is being perceived (Lenzon-Erz Reference Lenson-Erz, Sachs-Hombach and Schirra2013).
Thus, the disorientating, low-light condition of caves served as proxy for the quotidian environment, especially as firelight intensified the suggestive contours of the caves by making shadows more intense and animated. As one penetrates further into a cave, the natural light fades from liminal to complete blackness, simulating the way daylight transforms into night, and vice versa when leaving a cave. The transition from daylight to twilight to night-time may explain why animals were often portrayed in liminal areas of caves (e.g. Altamira's Polychrome Chamber and Lascaux's Hall of the Bulls) as well as their deeper recesses (Pettitt Reference Pettitt, Dowd and Hensey2016). These insights suggest that, although the caves were, in one sense, detached from everyday life, at the same time, they simulated certain fundamental aspects of that life, with all this implies. As Pettitt (Reference Pettitt, Dowd and Hensey2016) proposed, the darkness of the caves formed part of the art's ‘landscape context’ where touch interacted with vision as a means of navigating the various chambers. Darkness also intermingled with both the abbreviated and more complete depictions in a way that accentuated the coterminous appearance and dissolution of the suggestive natural features and graphic highlights/outlines (Hodgson Reference Hodgson2003a; Pettitt Reference Pettitt, Dowd and Hensey2016). Animals, therefore, seem to emerge from and dissolve into obscurity, passing through the viewer's light source as they progress through the darkness. Thus we seem to see a continuum in the expression of and perception of animals that the darkness helped modulate, where animals might seem to emerge in different ways from the cave environment, whether directly from the natural features, by way of the graphically highlighted natural features, or from the fully realized graphic depictions bereft of natural features; in this way, a horse ‘falls’ out of a crack in Lascaux, or the bison ‘drip’ from ceiling bosses in Altamira's Polychrome Chamber. Ultimately, the suggestive natural features may have been considered as significant to UP visitors as the graphic depictions, in that the perceived ambiguity was self-reinforcing.
Interaction of visual imagery with the perceived world
Somewhere in the middle of the fifteenth century, Leon Battista Alberti suggested in De Statua that the origins of art could be traced to the point when viewers
… happened to see in some tree stumps, or in clay, or in various other materials, some features which could, with a little work, be transformed into something similar to faces made by nature … as these men's studies advanced, they no longer needed to see an initial likeness within their materials in order to express any object they wanted. (Arkles Reference Arkles and Arkles2013, 9)
Lucquet (Reference Lucquet1930) used Alberti's notion to explain the perceived relationship between natural shapes and art in Palaeolithic cave art, a notion subsequently taken up and popularized by Breuil (Reference Breuil1906; Reference Breuil1952), who essentially summarized these previous perspectives by suggesting that UP people first became artists when they realized that natural things could be interpreted as objects, such as figures seen in stones and rocks, and human handprints could be seen to represent hands (see discussion in Lorblanchet Reference Lorblanchet2007). In his seminal Art and Illusion, Gombrich (Reference Gombrich1960), building on Alberti's insight, proposed that in a similar manner to ‘primitive man's’ ability to find animal images in the constellations, the animals portrayed in UP caves derived from the propensity of the human mind to project images onto suggestive surfaces; ‘could it not be that bulls and horses were first “discovered” by man in these mysterious haunts before they were fixed and made visible to others by means of coloured earth?’. This is a well-known phenomenon that has been explored by perceptual psychologists in a number of scenarios (Hodgson Reference Hodgson2008; Rorschach Reference Rorschach1942). Gombrich suggested a gradual progression from the earliest manifestation of that tendency towards the more graphically complex (Magdalenian) depictions, although the relatively naturalistic outline drawings and engravings of animals from the early Gravettian (e.g. Grotte Mayenne-Sciences: Pigeaud et al. 2001; and the caves of the Quercy: Lorblanchet Reference Lorblanchet2010) suggest that the portrayal of animals in caves was relatively sophisticated at least by around 30,000 years cal. bp. The same can be said of some of the sophisticated depictions of animals from Chauvet cave that may pre-date this period (Fritz & Tosello Reference Fritz, Tosello, White, Bourrillon and Bon2015), though this is not accepted by all archaeologists. Moreover, Gombrich's account fails to explain why animals were portrayed in the aforementioned stereotyped ways over such a long period. His ‘projection hypothesis’ can, nevertheless, be accommodated within the present context, in terms of its prediction that animal images were so strongly ingrained in the visual brain of UP hunters that they were liable to be projected on to suggestive natural features as well as fairly uniform surfaces; a tendency that derives from the fact that objects dominating the visual experience of hominins were those that evoked profound desires and fears, which were portrayed in palaeoart because they were ‘imprinted’ more strongly than other objects as part of a taxonomic visual system (Bednarik Reference Bednarik1986; Hodgson Reference Hodgson2003a,b; Reference Hodgson2008; Reference Hodgson2013a). Such projections are referred to as projective hyperimages in the sense that the hunter's visual system is primed or tuned to detect animals in various situations, which especially comes to the fore in dimly lit caves replete with suggestive natural features where the prospect of actual dangerous animals such as bears lurking was real. It should be noted that projective hyperimages are not equivalent to pareidolia [the mind perceiving a pattern where none exists, e.g. faces in clouds] due to the fact that, unlike the latter, the former is not a random process as it relies on the primed hyper-charged visual system that conditions or biases visual experience. Although the tendency for pareidolia is universal, because of its arbitrariness it is unable to account for the depiction of certain types of predators and prey in palaeoart, though it will have provided a platform for the more directed or engaged projective hyperimages (pareidolia is part of a broader psychological phenomenon known as apophenia—that includes other senses, including sound and touch—defined as the propensity to see meaningful patterns or connections in random data: Petchkovsky Reference Petchkovsky, Dowd, San Roque and Petchkovsky2008).
Davis (Reference Davis1986) criticized Gombrich's projection hypothesis for its inability to explain instances where suggestive natural features were not exploited graphically. Conversely, he proposed that, in a series of fortuitously created graphic lines, iconic figures may appear to emerge; in other words ‘objects are not seen as marks; rather, marks are seen as objects’ (Davis Reference Davis1986, 199: Lorblanchet Reference Lorblanchet2007, as well as Lorblanchet and Bahn,Footnote 1 posit a similar scenario). The two approaches, although distinct, are not mutually exclusive. Davis’ forms emerge by way of projection from a series of previously created abstract graphic marks (the line comes first and is then interpreted), whereas for Gombrich, iconic images emanate through projection from existing natural features (Avital Reference Avital2001). The difference, therefore, relates only to whether humans created the particular form from which a projection emerges or whether this was supplied by the existing natural features, which is hardly an important distinction for our purposes.
In addition, Davis (Reference Davis1986) employs the term ‘seeing-as’, which is similar to Wollheim's ‘seeing-in’ except, in the latter, projection is emphasized. ‘Seeing-as’ is, however, an automatic, erroneous and involuntary perceptual interpretation involving mere curiosity when viewing a visual array (Davis Reference Davis1987)—as in pareidolia (Bednarik Reference Bednarik2016)—whereas ‘seeing-in’ is the ‘cultivation of a special kind of visual experience, which fastens on certain objects in the environment for its furtherance’ (Wollheim Reference Wollheim1980, 223) and is therefore more closely aligned with projective hyperimages. Ultimately, Davis (Reference Davis2010, 78, fn 32) came to accept Wollheim's thesis regarding the dynamic relationship between seeing-as and seeing-in and therefore implicitly acquiesced with Gombrich's projective hypothesis.
In Davis's view, because rudimentary non-figurative mark making pre-dated iconic depictions, the latter should have originally derived from such elementary graphic arrays, which he refers to as self-sufficient marks. On this basis, and similar to Gombrich's developmental trajectory, Davis also proposed a (purely hypothetical) development from ‘abbreviated’ or ‘depleted contours’ of animals during the Early Upper Palaeolithic (Aurignacian) towards the more ‘complete’ depictions of the Late Upper Palaeolithic (Magdalenian). As with Gombrich's approach, and apart from misconstruing the Palaeolithic artistic record, one simply cannot maintain such a trajectory from incomplete depictions in the EUP to complete graphic images in the LUP; Gravettian outline images of animals—such as those from the Grotte Mayenne-Sciences or Chauvet—are complete and highly naturalistic well before the Magdalenian. Davis’ hypothesis can, therefore, also be rejected on the same grounds, plus the fact that it is possible to see a diversity of objects in a matrix of lines, whereas animals were almost exclusively depicted in a specific orientation and style (Ego Reference Ego2016; Hodgson Reference Hodgson2003b; Reference Hodgson2003b; Hodgson & Helvenston Reference Hodgson and Helvenston2006a).
Gombrich's and Davis’ hypotheses may, however, still be compatible (as originally suggested by Hodgson & Helvenston Reference Hodgson and Helvenston2006a), not only because one might argue that rudimentary graphic lines pre-date the UP by a considerable margin (e.g. those engraved onto the ochre crayons at Blombos Cave, South Africa: Henshilwood et al. Reference Henshilwood, d'Errico and Watts2009) but also because the ability to project images onto suggestive rock surfaces similarly pre-dates the UP—as exemplified by the Berekhat Ram and Tan-Tan pierres figures where natural features of cobbles resembling the human form were minimally modified by engraving in order to mark or ‘draw out’ certain anatomical areas (e.g. Bednarik Reference Bednarik2003a; d'Errico & Nowell Reference d'Errico and Nowell2000). Moreover, recent investigation of the Roche-Cotard ‘mask’ found that it was intentionally modified—if only minimally—by Neanderthals to produce a cat-like face now thought to date as old as ~75,000 bp (Marquet in press, cited in Lorblanchet & Bahn Reference Lorblanchet and Bahn2017). This strengthens the argument for Neanderthals possessing and deploying the ability for ‘seeing as’ or pareidolia long before the first figurines of the Swabian Jura and the two-dimensional depictions of Spain and France. Such ‘sculptures’ are referred to as ‘passively conceived’ (Hodgson Reference Hodgson, Sachs-Hombach and Schirra2013b), as they depend essentially on the projective capacity of the human visual system with most of the original areas of the natural rock left intact (demonstrating that the full potential of the visual image was not realized in the actual materials). In effect, such figurines depend largely on pareidolia or seeing-as for their realization. Such observations indicate that objects of this genre were mainly created at the level of ‘seeing-as’, as were handmarks, simple outlines, collection of naturally occurring rocks with incidental likeness to animals or the human form, etc., which were, nevertheless, gravitating towards fully proactive graphic iconicity (Hodgson & Helvenston Reference Hodgson and Helvenston2006a). Bednarik (Reference Bednarik2003b, 127), however, goes on to state that the capacity for iconicity, as manifest in the first representational depictions, is a ‘… “managed”, intentional use of visual ambiguity’. Thus, the combination of 1) an ability to make non-iconic marks with 2) the enhanced projective capacity of the visual system and 3) the suggestive natural features of caves, provided a mechanism for the eventual realization of fully iconic depictions. Caves could then serve as a scaffold for directing the specific placement of lines/marks. Thus, a subtle dynamic interaction occurred between the material world, mental imagery, and the process whereby the materials utilized for depiction were actualized (Hodgson Reference Hodgson2003a; Reference Hodgson2006a; Laursen Reference Laursen1993). We believe that these observations go a long way in resolving Davis’ misgivings as to the relevance of projective imagery to the origins of iconic depictions.
Archaeological evidence for projective hyperimagery
Natural fissures (‘cracks’), which simulate engraved and drawn lines, were often employed in cave art as a starting point for a depiction, and were sometimes the main constituent of an image (Fig. 3). In La Garma and El Castillo caves (Cantabria), Pettitt et al. (Reference Pettitt, Castillejo, Arias, Ontanon Peredo and Harrison2014), for example, found several examples of hand stencils that had been ‘matched’, that is, deliberately placed in specific relation to natural cracks, including stencils in which cracks acted as the line of one side of the hand stencil's arm (Fig. 4), an association which is particularly meaningful as these handmarks are, at their youngest, EUP in age. Lines are a perceptual feature to which the human visual system is particularly sensitive, as they act as the bounding contour of objects and are therefore the subject of considerable neural investment from the retina to higher visual areas (Mijović et al. Reference Mijović, De Vosa, Vanderperrena, Machilsenc, Sunaerte, Van Huffela and Wageman2014; Walther Reference Walther, Chai, Caddigan, Beck and Fei-Fei2011). As we can now be fairly sure that handmarks pre-date iconic depictions, they may have served as a forerunner to the more elaborate animal depictions where the stains, cracks and fissures of the cave wall provided a further decisive stimulus for the realization of the naturalistic animal depictions. It is notable that self-sufficient marks, such as red discs and other motifs (rectangle and claviform), are commensurate in time depth with the hand prints, dating in some caves to a minimum of 40,800 bp and in others to a minimum of 64,000 bp (Hoffmann et al. Reference Hoffmann, Standish and García-Diez2018a; Pike et al. Reference Pike, Hoffmann and García-Diez2012). As Pike et al. (Reference Pike, Hoffmann and García-Diez2012, 1412) concluded:
Our earliest dates (pre-Gravettian) are for art that is nonfigurative and monochrome (red), supporting the notion that the earliest expression of art in Western Europe was less concerned with animal depictions and characterized by red dots, disks, lines, and hand stencils.
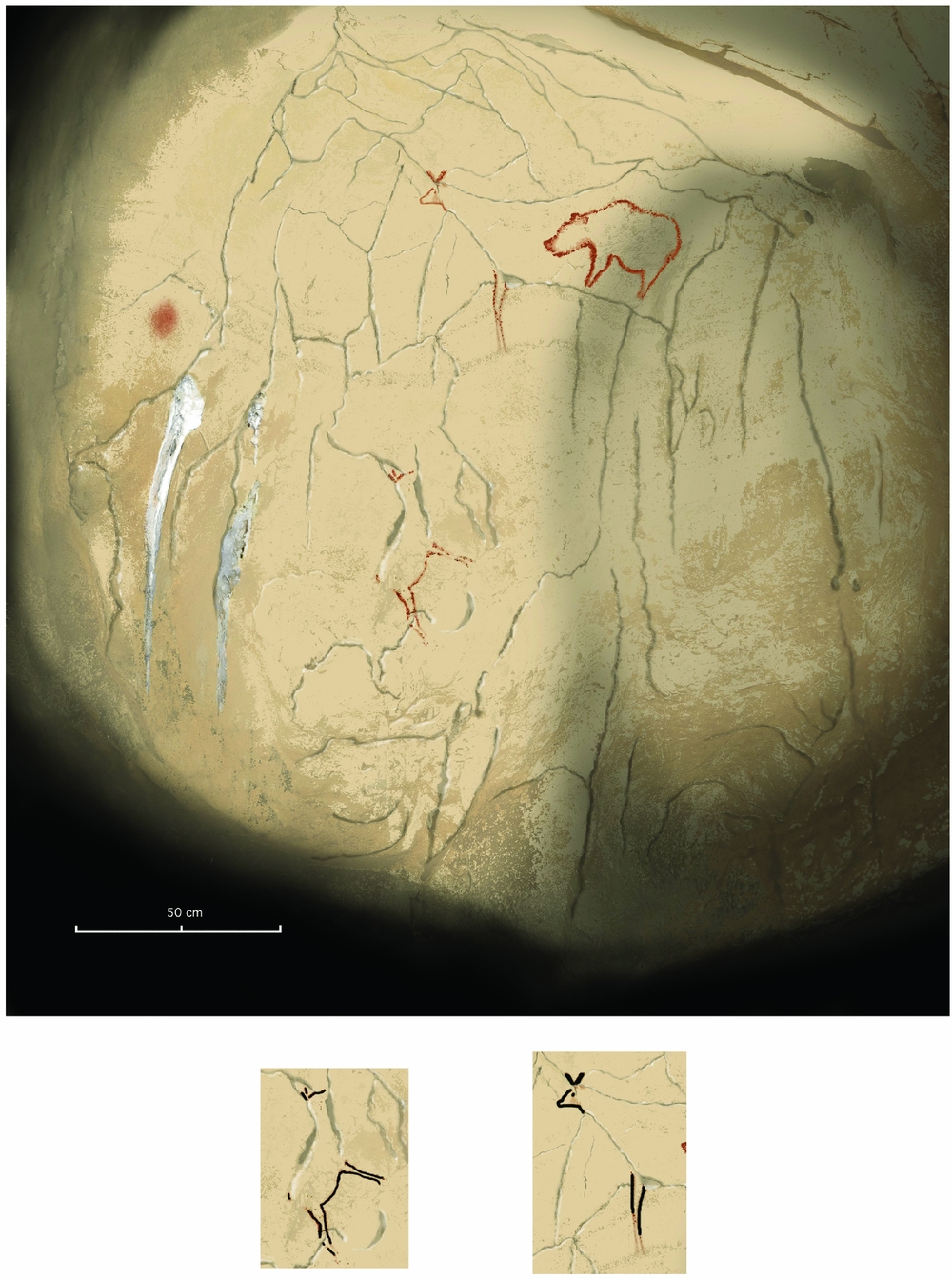
Figure 3. In Chauvet cave, cracks were exploited to create the two deer profiles (highlighted). Note that a minimum of graphic features were added to the two figures in order to complete the depictions. (Photograph: Carole Fritz and Gilles Tosello, CNRS and Équipe Chauvet. Courtesy of French Ministry of Culture and Communication.)

Figure 4. Hand stencils from the Great Panel of Hands in El Castillo (Cantabria), showing (a) a fissure to the right that seems to guide the line of the hand and possibly represent an adjoining arm; and (b) a fissure to the left that clearly matches the edge of the stencil. (Photographs courtesy Roberto Ontañon and Gobierno de Cantabria.)
This observation accords well with Hodgson's (Reference Hodgson2000a,b) insights regarding the primacy of ‘perceptual primitives’ that are prioritiszed in the early visual cortex in a way that gives rise to self-sufficient marks that pre-date the UP by a considerable margin. The creation of simple graphic outlines of the hand may therefore be evidence of what Ego (Reference Ego2016) calls the ‘hands eye’—a novel collaboration between the internal and external, which, as Hodgson (Reference Hodgson2011) proposed, is an external realization of the interaction between the ‘what’ (recognition, imagery and visual memory) and ‘where/how’ (visuo-motor) cortical pathways that are fundamental to making ‘art’.
Transforming the propensity to produce handmarks to depicting the quite complex features that characterize animal outlines is challenging, however, which is borne out by the long time period that seems, on current evidence, to have elapsed from when the first hand marks appeared before 64,000 bp to the first appearance of animal depictions by around 37,000 bp. The question arises as to the nature of the predisposing factors that facilitated such a transition, to which we now turn.
Implicit and explicit trigger cues, synecdoche and pars pro toto
One of the intriguing discoveries of visual science is the fact that human behaviour can be biased or influenced by external stimuli that is not registered consciously (see Hodgson Reference Hodgson2003a and references therein). Consequently, even though an object in peripheral vision may not be consciously perceived, it may still cause assemblies of neurons to fire in the early to intermediate visual cortex. Eagle et al. (Reference Eagle, Wolitzki and Klein1966), for example, presented a tree as figure and the suggestion of an animal as ground (Fig. 5) to individuals so rapidly they were only able to discern the tree and not the embedded animal. On a subsequent drawing task, the subjects were nevertheless influenced by the implicit animal in that they made reference to the animal in their drawings, thereby demonstrating that consciously unregistered information can influence later behaviour. The Perky Effect (Perky Reference Perky1910; Reeves Reference Reeves1982; Segal & Fusella Reference Segal and Fusella1970) demonstrates the same phenomenon, whereby an undetected image projected onto a screen surreptitiously influenced conscious visual imagery (see Hodgson Reference Hodgson2008 for an explanation of this effect as applied to UP art). These two studies illustrate how implicit perceptual processes influence behaviour, of which individuals remain unaware. It is therefore justifiable to assume that suggestive natural features of caves served as unconscious/implicit cues that triggered either the perception and production of animal outlines in various parts of a cave, or the addition of one or two graphic lines in order to accentuate the resemblance of a crack or fissure to an animal, and by so doing turning an implicit signal into an explicit cue.

Figure 5. Effect of a concealed figure in a stimulus. (Reproduced from Eagle et al. Reference Eagle, Wolitzki and Klein1966. Permission of the American Association for the Advancement of Science. License No. 4087750268521.)
The Rubin's face/vase illusion is relevant here (Fig. 1), which shows that, given an unstable image that depicts a vase or faces depending on which is seen as figure or ground, the image tends to flip automatically from the face to the vase and vice versa, on average every six seconds (Windmann et al. Reference Windmann, Wehrmann, Calabrese and Güntürkün2006). This is an automatic process, linked to the hypothesis testing and predictive coding discussed above. This procedure would have been facilitated by the suggestive cave features, whereby they are initially seen as mere natural features, but are then ‘flipped’ to be viewed as animals. It is easy to see this process occurring as a participant moves through a cave, with the light of lamps illuminating the topography as they pass, and their perceptual systems ‘flipping’ between wall–animal–wall. Such reversals are also observable in the Rorschach (Reference Rorschach1942) Ink Blot test, which consists of an amorphous visual array where various objects in the form of visual imagery are projected, or flipped, onto the array (Fig. 1); similarly with the Dalmatian dog test (Fig. 1), where a camouflaged dapple-coated canine is utilized to assess which areas of the brain become active when an observer differentiates an animal from distractors (Ludmer et al. Reference Ludmer, Dudai and Rubin2011). The amygdala was found to be particularly active together with area LO (lateral occipital in the mid-level visual cortex) as well as the medial frontal cortex. The authors concluded that the sudden, internally mediated, solution to a perceptual problem induces long-term memory benefits, because activation of the emotional areas consolidates the processing of information. In other words, a sense of reward/satisfaction is induced, probably by the release of dopamine into neural channels (Petchkovsky Reference Petchkovsky, Dowd, San Roque and Petchkovsky2008). A sense of gratification in this context is confirmed by the finding that when an individual disambiguates—through induced insight—an amorphous graphic array (equivalent to camouflage and similar to a Rorschach ink blot) in which an object is embedded, the reward system of the brain becomes activated, leading to an ‘aha!’ moment facilitated by successfully identifying the hidden figure (Ludmer et al. Reference Ludmer, Dudai and Rubin2011). In fact, brain scans conducted by Asari et al. (Reference Asari, Konishi, K., Chikazoe, Nakamura and Miyashita2010) showed that when subjects see an image in a Rorschach inkblot, the anterior prefrontal area, along with the amygdala (for emotion), becomes more active (Liu et al. Reference Liu, Li, Feng, Li, Tian and Lee2014 found a similar scenario with faces). This suggests the matching of internally generated images with external visual stimuli whereby fragmentary bottom-up signals (such as in the Rorschach inkblot) originating from the occipito-temporal area are combined in the temporo-polar region with top-down information (for visual imagery) from the prefrontal area by way of the amygdala. This, in effect, ‘colours’ the experience with emotional valence. Moreover, the Rorschach inkblot has been found to activate the mirror neuron system in a way that induces a feeling of movement and appears to interact with the mirror neuron system for touch (Girimoni et al. Reference Giromini, Porcelli, Viglione, Parolin and Pineda2010), thereby providing a link between the suggestive cave features (implied Rorschach) and active use of palpation suggested by Pettitt et al. (Reference Pettitt, Castillejo, Arias, Ontanon Peredo and Harrison2014) (see also Dobrez Reference Dobrez2017), the latter of which may also derive from the apophenia discussed above. Thus, the observed cave handmarks will have led to a sense of identification by accentuating the agency experienced in the immediate environment.
These findings suggest that, when cave artists were confronted with the challenging visual environment of the cave, which, we recall, simulates the amorphous landscape of the everyday hunting environment, the sudden perception of animals in the natural cave features would have evoked a similar sense of self-induced reward, especially when this experience was obviously accompanied by emotional arousal related to being in a dangerous and threatening location. The sense of accomplishment that ensues may therefore have motivated the repetition of the experience. Moreover, when viewing potentially ambiguous scenes suggestive of danger, the visual system becomes more sensitive to detail and discerns objects more rapidly compared to viewing neutral stimuli (Kobayashi et al. 2016; Lane et al. Reference Lane, Chua and Dola1999), which is particularly enhanced with partly concealed animal-like stimuli (Altman et al. Reference Altman, Khislavsky, Coverdale and Gilger2016). It is easy to see how, once particular animals have been projected onto cave walls, such repetition could morph, culturally, into behavioural rituals in specific environments such as caves/rock-shelters, and hence to ritual behaviour. Interestingly, religious individuals and those who believe in the supernatural see objects in amorphous arrays more frequently and to a greater degree than sceptics (Riekki et al. Reference Riekki, Lindeman, Aleneff, Halme and Nuortim2013). Hodgson (Reference Hodgson2003a) provides further cogent examples of similar relevant perceptual processes including the basic principles of how unconscious priming influences the way the world is perceived.
Evidence from semiotics
The diagnostic anatomical cues for animals, notably the head, eyes, neck, back and legs, are processed pre-attentively in the early to intermediate visual cortex (Delorme et al. Reference Delorme, Richard and Fabre-Thorpe2010). Such cues relate to Peirce's indexical signs (Walther et al. Reference Walther, Chai, Caddigan, Beck and Fei-Fei2011), whereby only minimal cues are necessary to signal an object through association. However, the indexical appears to take precedence over the iconic, as the former is a preconscious reaction based on fragmentary information that requires an immediate response through ‘blind compulsion’ in order to avoid danger. Blind compulsion refers to signs that have a direct physical association with an object that spontaneously directs attention. Thus, indexical signs exemplify the commonest means by which animals (including humans) respond to stimuli (Sebeok Reference Sebeok and Ketner1995) by way of simple features that incite a response to criteria specific to a species’ evolutionary past. Indexical signs are therefore more naturally assimilated than the iconic (Kull Reference Kull, Cobley, Deely, Kull and Petrilli2011), as the latter rely more on interpretation and tend to be acquired later. Indexical signs can be also regarded as more fundamental (Cheyne Reference Cheyne1993), because they can signal where an animal may be concealed. Diagnostic/indexical cues of animals thereby served to indicate aspects of animal behaviour to which early humans needed to remain vigilant in order to survive (Sebeok Reference Sebeok and Ketner1995). In effect, the indexical served as an important pars pro toto, or synecdoche, by providing the initial clues as to the status of an animal.
Interestingly, the full range of indexical to fully iconic cues can be seen in Gargas cave (Barrière Reference Barrière1976; Foucher et al. Reference Foucher, San Juan-Foucher and Rumeau2007), beginning with the indexical around 27,000 bp and proceeding, eventually, to the fully iconic. Thus, the first depictions of animals at Gargas consist of the outline of the head, neck and withers, followed later by the inclusion of the forward areas and hindquarters and, finally, the complete outline. A similar scenario can be found at Chauvet (Fritz & Tosello Reference Fritz, Tosello, White, Bourrillon and Bon2015). Despite the fact that the depictions were probably completed in one phase, Fritz (Reference Fritz1999; Fritz & Tosello Reference Fritz and Tosello2000) found similar preferences in the sequence of animal contours during the Magdalenian as well as during the Aurignacian/Gravettian. These insights suggest that, despite relatively complex depictions of animals existing at an early date, it was the abbreviated outlines that predominated. This indicates that the attenuated outlines were of greater significance to the instigators, otherwise they would not have persisted with such a format during the whole of the UP.
Discussion
An ability to produce non-iconic graphic primitives, when merged with the highly charged and ‘hair-triggered’ projective capacity of the visual system for detecting animals in certain environmental situations, suggests that iconic depictions can be regarded as both ‘invented’ and ‘discovered’. We believe that our suggestions go some way to answering the question posed by Davis (Reference Davis1987; Reference Davis2010) as to how, when there were no previous informants to rely upon, the original image maker came to conceive the significance of the first iconic depiction. In effect, this required various factors to coalesce in the way described. Handmarks pre-date the depictions of animals—probably by a considerable margin and apparently on several continents—implying that they acted as an important prerequisite to the faunal images. That process probably began simply with hands touching suggestive natural features as part of a process of exploration, which is supported by the fact that many handmarks are located on or near specific natural features, rather than placed randomly on suitable surfaces (Pettitt et al. Reference Pettitt, Castillejo, Arias, Ontanon Peredo and Harrison2014). Moreover, as in Gargas cave and elsewhere, some early animal outlines were made in clay (mondmilch) using fingers. In both cases, there is a strong element of palpation involved in the placement of the art on the natural surface. Nevertheless, although handmarks demonstrate that iconicity was possible, a further device was required to facilitate the actual animal depictions. The natural suggestive features of the cave together with the aroused hyperactive visual system for perceiving animals appear to fulfil this role. Depictions were related to a hunting adaptation in that they reflected a preoccupation with herbivorous prey and dangerous predators, which, although they may not always have directly corresponded to diet and survival, nevertheless reflect a concern with critical species as a necessary perceptual ability which allowed fauna to be differentiated from their environment. Thus, hunting, in the wider sense of the term, would have required a holistic understanding of fauna that encompassed not only those taxa directly related to diet, but also those threatening and ‘neutral’: in other words, animals the hunters ate, those animals that ate hunters, and animals with whom hunters competed. By linking Palaeolithic art with the everyday imperatives of UP hunter-gatherers—that is, the need to detect animals in various changing environmental and ecological conditions which, in some respects, simulates the cave environment—we are much better placed to avoid the impasse of first causes cited by Davis (Reference Davis1987; Reference Davis2010).
In addition to handmarks, a further important antecedent to animal depictions may have been the ability of some hunters to exploit disguises by wearing animal skins in order to approach game. Such strategies could have been so effective that, from a distance, disguised humans may have been mistaken for animals by their fellow hunters (Hodgson & Helvenston Reference Hodgson and Helvenston2006a). Additionally, clothing was, of course, constructed from animal skins and furs. This suggests that, as well as handmarks and the other factors already alluded to, a number of criteria interacted to promote the depiction of animals, including animal footprints/tracks that require complex cognitive processes in order to find and pursue animals (Avital Reference Avital1998; Hodgson & Helvenston Reference Hodgson and Helvenston2006b). It is well known that some of the ‘signs’ usually interpreted as ‘vulvar’ imagery probably represent animal prints and tracks (Bahn Reference Bahn1986), e.g. the early Aurignacian engravings of Abri Cellier and Abri Blanchard in the Dordogne that more closely resemble horse and bison tracks (see Figure 6). In sum, although handmarks may have acted as a bridge towards realizing the more complex animal depictions (Hodgson Reference Hodgson2006a; Reference Hodgson2013a; Malotki Reference Malotki, Sachs-Hombach and Schirra2013: Pettitt et al Reference Pettitt, Arias, García-Diez, Bueno-Ramírez and Bahn2015), they needed to act in tandem with other precursors for this to occur. The vital final link seems to have been provided by the affordances of the suggestive natural rock surfaces together with the human perceptual system's sensitivity to lines/contour and inherent projective capacity.

Figure 6. Top, left to right: Abri Cellier and Abri Blanchard blocks with purported engravings of vulvae iconography (Abri Blanchard and Abri Cellier illustrations with permission of Quaternary International). Bottom left: Track made by bison in snow (kind permission of Dennis Deck). Bottom right: Tracks made by horse with unshod hooves in snow showing the front and back footprints (public domain)—note, with horses, the front print is more circular than the back print, which simulates the round and more ellipsoid engravings of the engraved blocks. Such tracks would have been salient in the snowy ice age conditions of the Upper Palaeolithic. (Reprinted from White et al. Reference White, Bourrillon and Mensan2017; © 2017 with permission from Elsevier.)
The point should be reinforced here that, although the caves provided a particularly resonant environment where natural features were exploited for depictive purposes, the fact that such features were also incorporated into animal representations throughout the world in rock- shelters and outdoor sites (Taçon et al. Reference Taçon, Tan and O'Connor2014)—which are also prone to shifting light conditions—suggests that the same dynamic interaction between the material world and the highly sensitive visual system of hunters for perceiving animals was at work in the wider landscape (Hodgson Reference Hodgson2008; Hodgson & Watson Reference Hodgson and Watson2015). A large number of engravings of animals from the open-air sites of Foz Côa and Siega Verde have also been found that incorporate predisposing natural features from the earliest (probably Gravettian) phases (Fernandes et al. Reference Fernandes, Reis, Remirez and Marcos2017). This is not surprising; the hunter's visual system needed to remain acutely primed to detect animals outdoors in order to discriminate between predators and prey. That the first complex depictions are specifically of megafaunal prey in almost all rock-art sites in caves, rock-shelters and open-air contexts throughout the world further supports this notion (Chaloupka Reference Chaloupka1993a; Taçon & Chippindale Reference Taçon and Chippindale1994; Taçon et al. Reference Taçon, Tan and O'Connor2014).
The proposition that this art can be regarded as ‘naturalistic’ has, however, to be tempered by the fact that it was largely devoid of the hallmarks of a true realistic art, namely, unusual, off-axis views. In addition, twisted perspective tended to be employed more frequently after the Aurignacian (Bourrillon et al. Reference Bourrillon, White and Tartar2017). By contrast, the sideways stereotypical view remained the prime concern of Palaeolithic artists over a remarkably long period, no doubt as it adequately fulfilled the needs of a hunter-gatherer for whom hunting of megafauna was critical to survival. As a by-product, it may have helped to sharpen perception and recognition skills, facilitating the rapid identification of certain animal taxa (Hodgson & Helvenston Reference Hodgson and Helvenston2006a).
Cave walls therefore provided just one opportunity for the graphic realization of animals, as many examples of animal depictions on portable objects similarly derive from existing natural features (Hodgson Reference Hodgson, Sachs-Hombach and Schirra2013b; Sauvet Reference Sauvet, Lejeune and Welte2004). A number of blocks from the Aurignacian, such as the Abri Cellier, are engraved with large herbivores that seem to have been inspired by the suggestive natural edge of the blocks that simulate the animals’ cervico-dorsal line (White et al. Reference White, Bourrillon and Mensan2017).Footnote 2 Some have argued that the art of the Abris Cellier, Blanchard, Castanet and corresponding sites in the Vézère valley and elsewhere may have had significant cultural links with Chauvet cave and the Swabian Jura (e.g. Bourrillon et al. Reference Bourrillon, White and Tartar2017; Floss Reference Floss2017). However, some commentators insist that the earliest figurines from Swabian Jura appeared suddenly and were sophisticated from the outset and represent the earliest figurative art at as much as 40,000 years old (Conard Reference Conard2009; Dutkiewicz Reference Dutkiewicz and Nuria2015; Floss Reference Floss2017). A parsimonious reading of the existing dates for these objects, however, would place them between 37,000 and 36,000 bp, and one of us (PP) sees no convincing thematic, stylistic or chronological link between these disparate regions and their localized art traditions. However, as stated by Hodgson (Reference Hodgson, Sachs-Hombach and Schirra2013b, 409):
… most of the sculptures of the Swabian Jura consist of thoroughly worked and highly finished artifacts (Conard Reference Conard2003) making it difficult to gauge whether they were initially inspired by the original natural features from which they were made. An elongated 28,000 year old siltstone has, however, been recovered from Hohle Fels where the natural resemblance to a phallus was enhanced using a few artificially produced scratches (Conard/Kieselbach Reference Conard and Kieselbach2006).
The phallic object confirms that projective imagery was influential in the iconography of Swabian Jura. If a link between the Aurignacian sites of France and the Swabian Jura were demonstrable, it would point to the possibility of codetermination. We need also to take account of the fact that the number of figures from Germany amounts to only around 50 objects—of which most are quite small depictions of large herbivores and carnivores—whereas the depictions of animals in France and Spain run into thousands. Recall that projective imagery was necessary for the Berekhat Ram and Tan-Tan pierres figures as well as the Roche Cotard Neanderthal ‘mask’ that pre-date the Upper Palaeolithic by a considerable period. The above examples show that projective hyperimagery is relevant to various scenarios that attest to the compelling nature of the phenomenon. It should be added that this compulsion was reinforced, not only by evolutionary instantiated regions dedicated to detecting animals in the human brain, but also the fact that focusing on particular objects leads to the recalibration of the area concerned that improves detection thresholds (Draganski et al. Reference Draganski, Gaser, Bush, Schuierer, Bogdahn and May2004; Sunday & Gauthier Reference Sunday, Gauthier, Hambrick, Campitelli and Macnamara2017). Notwithstanding mobiliary and outdoor art, cave depictions offer a vital clue as to the derivation of UP iconicity due to a number of interacting criteria dominating that environment. In sum, those constraints which had determined the functional outcome of the hominin perceptual system also became important as a trigger for the motifs to be found in UP ‘art’ through a sharing and exploitation of the same visual mechanisms (Hodgson Reference Hodgson2003b). Thus, the same visuo-neural system, together with associated emotional circuits, became active when a hunter viewed animals in the wild as much as when the suggestive rock contours were viewed, thereby motivating the addition of graphic marks to bring out the likeness of an animal.
A falsifiable model for the origin of figurative art
We propose the following hypothesis for the first appearance of figurative depictions. We are aware that the material of concern is so scarce and remote that it would be impossible to advance predictions that in their entirety should be falsifiable under current circumstances. With such limiting conditions, all we can do is to advance the most logically sensible, holistic and clearly stated hypotheses, in the hope that even if we cannot advance falsification criteria that can be met now, they may be in the not too distant future. We hope at least that our model carries with it the seeds of its own falsification. Thus:
1. Humans evolved to ‘think about’ animals due to their critical importance to the survival strategies of Pleistocene hunter-gatherers. As a result of this, the brain established a ‘hair-trigger’ response with a tendency to interpret natural features as animals.
2. Art probably evolved as decoration of the body and processes associated with it, providing the means (pigment, engraving) for creating marks, e.g. the use of ochre and shell necklaces that pre-date the Upper Palaeolithic (Hoffman et al. Reference Hoffmann, Angelucci, Villaverde, Zapata and Zilhão2018b; Pettitt Reference Pettitt, Henshilwood and d'Errico2011; Vanhaeren et al. Reference Vanhaeren, d'Errico, van Niekerk, Henshilwood and Erasmus2013; Watts Reference Watts, Botha and Knight2009). It makes sense that the earliest manifestation of art should be peripersonal.
3. Caves and other stimulating environments activated the brain's hair-trigger mechanisms, acting on its projective ability in terms of elements of the body (e.g. handmarks) and animals, stimulating the transferral of images from the body to objects and places.
4. The earliest of such ‘transferred’ art related to the body, i.e. non-figurative hand and finger marks, representing a direct extension of the body (or if preferred, self) to the external world. This was done by Neanderthals and, later, by Homo sapiens, although whether the two are connected remains to be seen.
5. Later, caves, rock-shelters and open-air landscapes triggered inherent projective capacities—and hence the completion/creation of animal depictions.
We are aware that there are too many unfalsifable hypotheses in the literature about the nature of Palaeolithic art. Given the scientific nature of this field and related areas such as visual science, we do not think this is a healthy situation. We suggest, therefore, that our hypothesis can be falsified according to the following criteria.
Proposal 1: Art evolved from bodies – to things – to places
Predictions
If the archaeological record cannot demonstrate art on the body itself (due to its perishable nature), then the earliest themes should reflect the body, e.g. hand and finger marks. This reflects the most parsimonious route from body decoration to permanent art on the external world. Furthermore, the earliest art should only appear on objects closely associated with the body, probably those connected with other tasks (or on by-products associated with those tasks). Depictions of animals should therefore appear later than earlier (non-figurative) extensions of the body. As an example, the rock art of Kimberley, Australia, perhaps dating back to 50,000 bp, consists of cupules (self-sufficient marks), that are later followed by outlines with some infill of large marsupials (Veth et al. Reference Veth, Myers, Heaney and Ouzman2017), perhaps around 36,000 bp. Although plants are depicted during the animal outline phase, the rock art is dominated by animals (75 per cent) compared to plants (25 per cent). Similarly, during the European UP the depiction of plants does very occasionally occur, at least in the Late Upper Palaeolithic, but animals remain the overwhelmingly dominant theme. Except for handprints, in most cases, however, prey animals pre-date other kinds of objects in Europe, Australia and other parts of the world as discussed above.
Falsification criteria
The appearance of figurative art—particularly animal depictions—noticeably before body extensions such as hand and finger marks would falsify the hypothesis of an earliest, non-figurative, body-centred and peripersonal phase. The depiction of things other than animals—plants, trees, landscapes, etc., predating the animal depictions would falsify the hypothesis that the first figurative images were of prey animals.
Proposal 2: Figurative art should clearly emerge after hand and finger marks
Predictions
In cases of unambiguous dating, examples of non-figurative art—particularly extensions of the body noted in Proposal 1—should pre-date (or their minimum ages should be consistent with) the dates for figurative art.
Falsification criteria
Any clear and unambiguous demonstration that figurative art clearly emerged in a given region before hand and finger marks. Although we note the Middle Pleistocene examples of ‘figurines’ from Berekhat Ram and Tan-Tan (and probably the Roch-Cotard ‘mask’) that obviously pre-date UP figurative art here, these are passively conceived as discussed above, meaning that most of the surface remains unmodified with only certain areas intentionally engraved. In no way has the full figurative potential of these objects been realized, and the majority of the effort involved turning the objects into figurines relies solely on visual imagery related to pareidolia. As a result, such objects can be regarded as idiosyncratic examples of the ability to indulge in projective imagery, rather than figurative art per se. It should not, however, be difficult to identify figurative images of an entirely or mostly artificial nature. Evidence from various global archaeological contexts in fact suggests that fully fledged figurative art emerged subsequent to both self-sufficient marks and handmarks.
Proposal 3: Figurative art should emerge in suggestive environments, whether they are in deep caves or the open air
Predictions
Although caves provide an especially potent environment where a number of criteria intersect that makes the probability of figurative art occurring more likely, this does not mean that figurative art should evolve later in the open-air environments in which hunting was practised. Open-air environments offer a similar interactive dynamic to caves, albeit with reduced saliency/resonance. We would, however, expect a comparable use of rock topography, depicted in a similar style to cave art.
Falsification criteria
The existence of open-air art that does not exhibit evidence of pareidolia or projective hyperimagery in that the depiction of prey animals similar to cave art does not include the exploitation of the natural topography.
Proposal 4: Hand marks and early figurative art will be associated with surface topography and/or pareidolia/seeing-in/hyperimages
Predictions
In most cases one would expect the majority of early art to exhibit meaningful relationships with its surface ‘canvas’, the latter having triggered the brain's projective capacity.
Falsification criteria
The earliest non-figurative and figurative art should not generally be associated with topography or pareidolia/hyperimages; there should be no demonstrable associations of such for the majority of examples.
Proposal 5: The tendency of the brain to engage in synecdoche predicts that incomplete images should play the dominant role in the origins of figurative art
Predictions
Evidence of synecdoche in the form of incomplete depictions should be evident in the earliest figurative art and most images should be incomplete, or at least sites with early figurative art should contain some examples of incomplete images and synecdoche use.
Falsification criteria
Most early images should be complete, i.e. with no requirement for synecdoche. The depictions of animals from Sulawesi seem fairly complete, where most of the outline is included, with the addition of some infill. On close scrutiny, however, one example is defined by a natural fissure in the form of an arch that closely simulates the depicted animal's dorsal contour—an implicit cue perhaps? Additionally, a lower fissure has been incorporated into the outline of the animal's abdomen, which probably served as an explicit cue (see figures 6a and 6b in Aubert et al. Reference Aubert, Brumm and Ramli2014). Furthermore, the caves in which these animals depictions exist are full of suggestive natural contours (Marchant Reference Marchant2016; Taçon et al. Reference Taçon, Tan and O'Connor2014), which were fully exploited for depictive purposes. Thus, there seems to be a continuum here, from natural features with minimal graphic intervention to more complete outlines with infill lacking natural features that originally derived from the natural features.
Conclusion
We believe that our hypothesis about the origins of non-figurative and figurative art accounts—in as parsimonious a way as is possible—for the current understanding of the archaeological record regarding the emergence of ‘art’, as well as the visual brain's role in the perceptual correlates specified. We suggest that our first hypothesis (that handprints, along with other precursors—footprints/tracks and hunting disguises—and the ability to make relatively simple graphic marks pre-dated and encouraged the production of animal depictions) is supported by these fields as they currently exist, as is our second hypothesis—that the highly tuned visual system of UP hunters for detecting animals interacted with the suggestive rock features and induced the addition of minimal graphic marks to complete an iconic depiction. Palaeoart studies need long-term models if we are to contribute to the understanding of the origins of ‘art’ in the wider social sciences, and we also need models that are, in theory at least, testable. We have accordingly proposed a means to falsify our own long-term model in this light. It is a coarse beginning, but we suggest it points in the right direction.






