



Introduction
A human community may be described as a particular biological population embedded in its own ecosystem (Clarke Reference Clarke1968). In these terms it is subject to the ecological conditions of the biotope, draining resources from it and influencing relationships between the species present. From such a perspective, studying an archaeological site and its remains can be viewed as a palaeoecological analysis of an anthropized ecosystem. Only a multi-disciplinary approach will thus be effective in achieving significant results.
Cultural factors—sometimes related to specific religious prescriptions or social conventions—influence relationships between human populations and their ecosystems and can affect the range of resources exploited. Medieval monastic communities are particularly interesting in this respect because the Orders’ Rules often impose upon them abstention from some commodities, such as some specific food or drink types. How did such monastic communities react to these artificial limitations to—or imposition on—available resources?
The aim of this research is to explore whether archaeological data can provide an answer to this question, examining a key early medieval monastic site in Italy whose sudden destruction ‘froze in time’ the very last moments of the monks’ daily lives. Accordingly, this paper sets out to reconstruct:
• the ecology of a particular human population in the past
• the monastic diet and the economy of an early-medieval Benedectine abbey in a specific moment of time
• the function of the different areas of the abbey's kitchens complex and the organization of the work related to food storage, processing and cooking in the abbey.
Historical and site context: San Vincenzo al Volturno. Rise and fall of a powerful early medieval monastic community in central southern Italy (SC, AC)
San Vincenzo al Volturno abbey, in the Molise region of central southern Italy (Fig. 1), is a key site for the study of these topics from a palaeoecological point of view, having been the focus of extensive excavation campaigns since the 1980s.
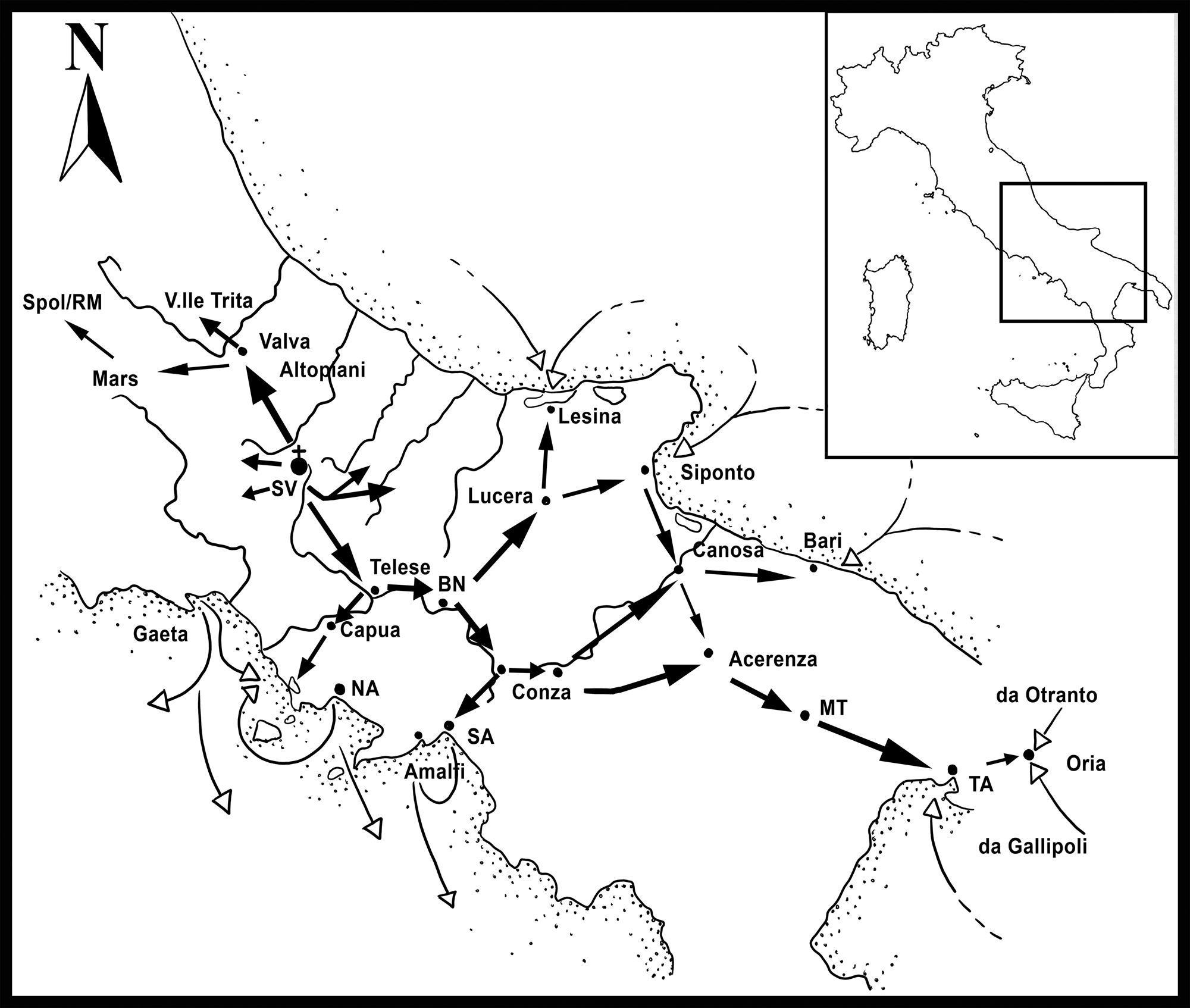
Figure 1. Location of San Vincenzo al Volturno and system of connection with roads and harbours. (Adapted from Marazzi Reference Marazzi and Marazzi1996, 64, fig. 4).
The abbey, founded by Langobard noble brothers in the first half of the eighth century ad near the sources of the River Volturno on the Molise Apennines (c. 550 m a.s.l.), quickly became one of the most important monastic power centres in Italy (Hodges Reference Hodges1997). It lay at the centre of political power-plays between the Carolingian Frankish kingdom established in northern Italy (after 774) and the Langobard principalities that ruled most of the Campania region (Marazzi Reference Marazzi and Marazzi1996; Reference Marazzi and Massullo2000, 21–9) which once included the Molise area. Later the abbey evolved into a virtual monastic ‘city’, favoured by Carolingian emperors, whose possessions extended all over central-southern Italy.
The wealth and role of San Vincenzo al Volturno ended dramatically when the abbey was sacked and set on fire during a Saracen raid in the autumn of ad 881 (probably 10 October), as recorded in the Chronicon Vulturnense (the main source for the abbey's history, written by the monk Iohannes around ad 1130). This sudden destruction caused the roofs of several structures to collapse, which sealed diverse contexts preserving remains related to daily monastic life, providing a freeze-frame of the very last moment of the early medieval monastery's life (Marazzi Reference Marazzi2002, 255).
Archaeological excavations began in 1980 under the direction of Richard Hodges (then at the University of Sheffield). From 1999, direction moved to Federico Marazzi (University of Naples ‘Suor Orsola Benincasa’). The excavations brought to light the destroyed structures of the abbey, including several churches, workshops and living areas (Fig. 2). These latter structures were articulated around a vast court. A large circular lavatorium—a place devoted to the monks’ hygiene—occupied the northeast corner of this area, where those rooms related to subsistence activity were also sited. The presence of the lavatorium, connected to the River Volturno by a drain running through the kitchen complex, suggests the existence of a nearby spring that supplied clean water for the daily needs of the early medieval monastic community.

Figure 2. Reconstruction of San Vincenzo abbey in the ninth century ad: (1) Basilica major; (2) workshops; (3) main court; (4) lavatorium; (5) refectory; (6), kitchen complex; (7) River Volturno; (8) Basilica minor. (Drawing: Simona Carracillo.)
A large refectory—with a capacity for up to 300 people—formed the north border of the court near the lavatorium; it was connected to the kitchen complex by an access room (for an exhaustive description of the zone, see Marazzi et al. Reference Marazzi, Filippone, Petrone, Galloway and Fattore2002, 255–7). The kitchen complex bordered the court on the eastern side. This set of rooms—devoted to food processing and cooking meals—lay between the court on one side and the left bank of the River Volturno on the other (Fig. 3). The Volturno—whose source lies just 2.4 km away—at the time closely bordered the eastern walls of the kitchen complex, although its course later migrated about 10 m further eastwards. Its original position was very practical both for drawing water and for the disposal of waste. The 881 destruction also involved the kitchen complex, which comprised two rooms, here called the ‘kitchen’ and the ‘ante-kitchen’ (Fig. 4). In each, excavation brought to light large amounts of bioarchaeological remains, plus vessels for cooking, etc. The abundance of these, recognised as largely connected with the monastic diet, suggested the need for detailed collection and analysis of both rooms and finds.
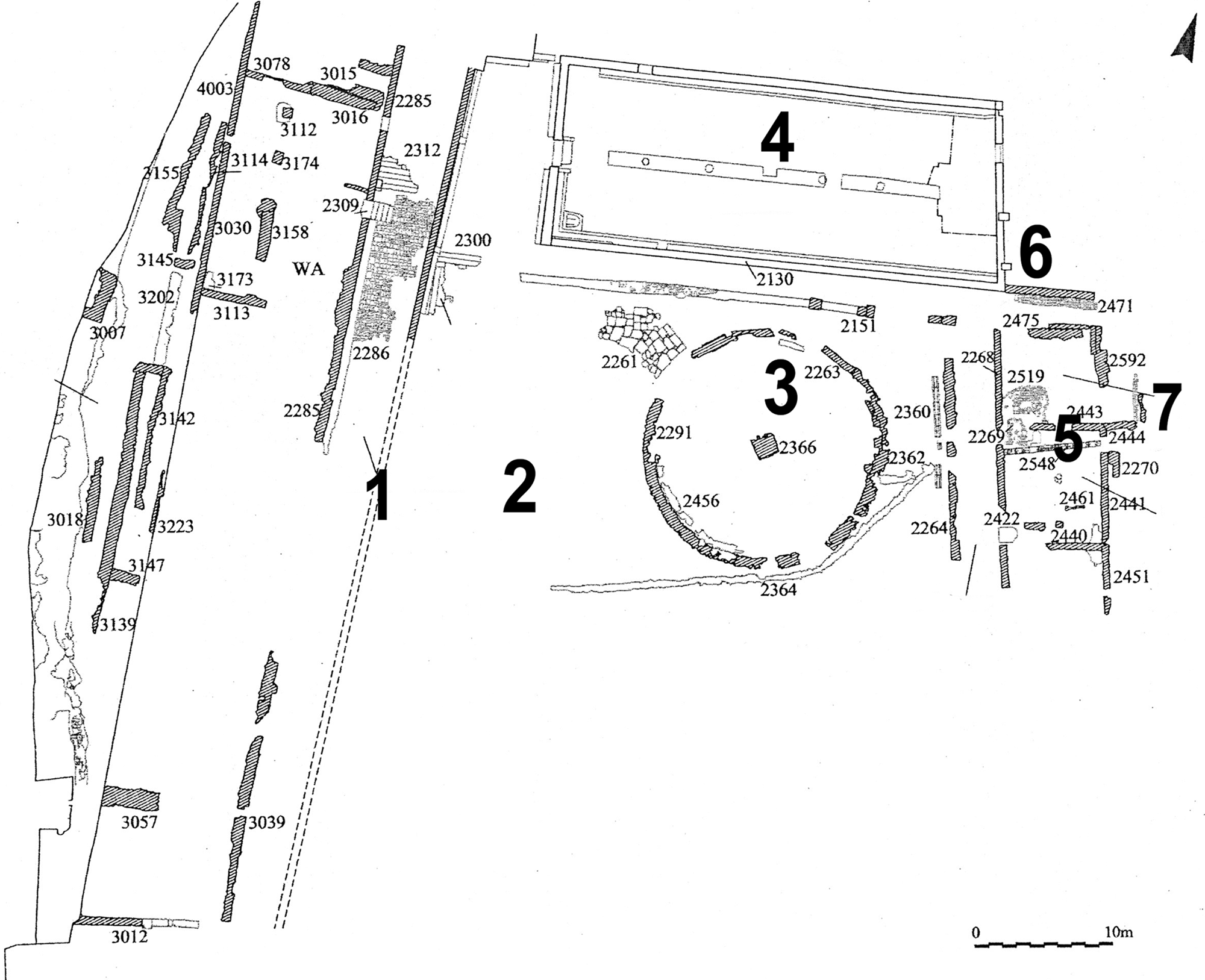
Figure 3. San Vincenzo: general plan of the main court area. (1) western corridor and portico; (2) main court; (3) lavatorium; (4) refectory; (5) kitchen complex; (6) access room of the refectory; (7) River Volturno. (Adapted from Marazzi et al. Reference Marazzi, Filippone, Petrone, Galloway and Fattore2002, 253, pl. 13.)
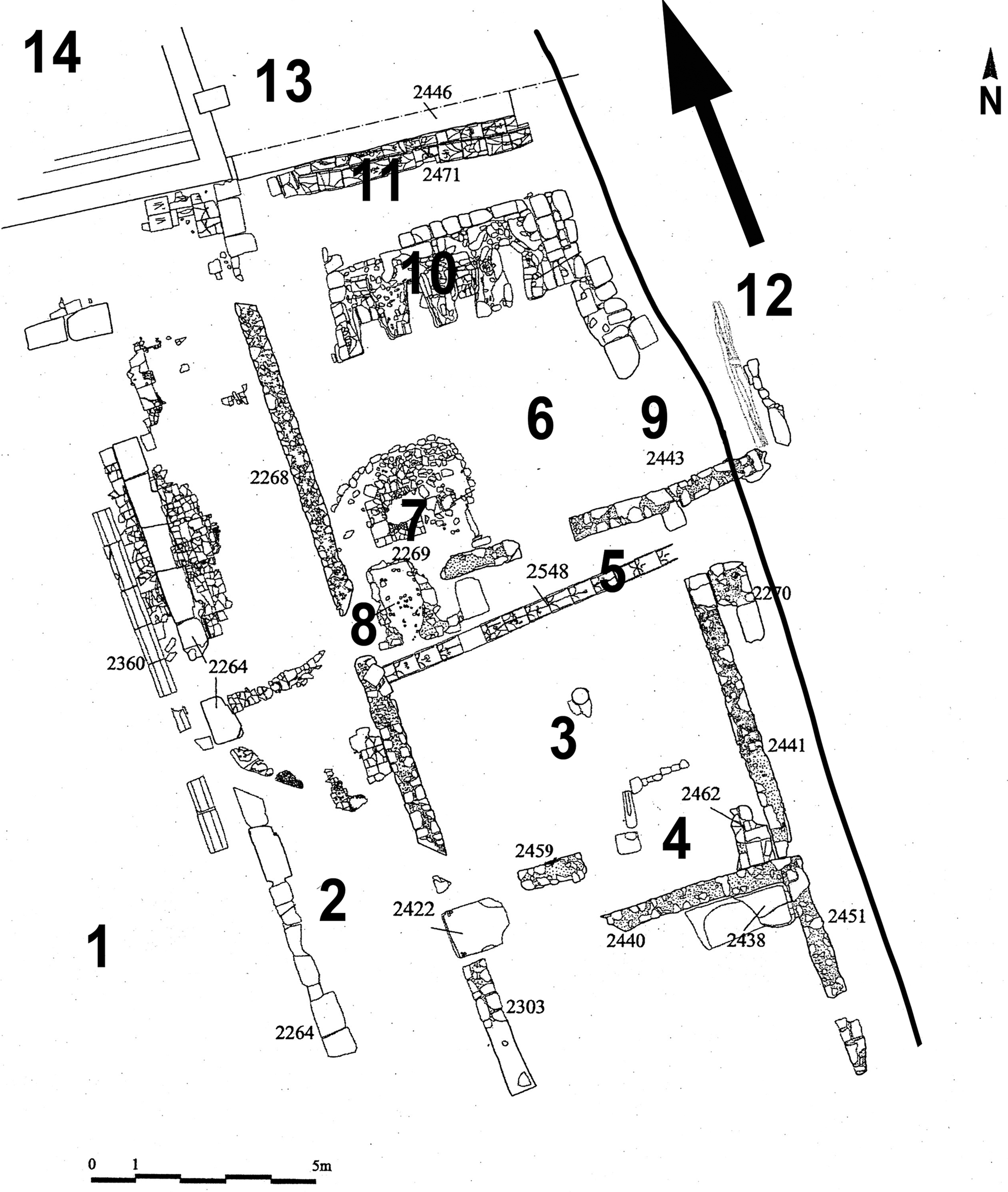
Figure 4. San Vincenzo: general plan of the kitchen complex. (1) Main court; (2) eastern corridor; (3) ante-kitchen; (4) larder; (5) drain pouring water from the lavatorium to the river; (5) kitchen; (7) hearth; (8) mantel/smokehouse; (9) pit over the river; (10) built-in cooker with four stoves; (11) slide (12) River Volturno; (13) access room of (14) the refectory. (Adapted from Marazzi et al. Reference Marazzi, Filippone, Petrone, Galloway and Fattore2002, 255, pl. 14).
The ante-kitchen was closely connected to the kitchen, but lacked structures related to the cooking of meals. Its southeast corner showed traces of a partition; the function of this delimited space was initially enigmatic, due to the lack of any archaeological remains. A simple drain, cut in the floor, ran along the inside of the northern wall: it collected water from the lavatorium and discharged into the River Volturno. The drain was partially filled with sediments and animal bones.
An opening in the same wall gave access to the actual kitchen, where two structures devoted to cooking meals were found: just left of the entrance, a large, elevated hearth in the southwest corner was bordered by a kerb of travertine blocks and paved with terracotta tiles. Right of the entrance was a sort of ‘pit’, which was originally situated over the river and covered by a wooden grating. The pit was filled with blackish sediments rich in bioarchaeological remains, including lots of egg-shell fragments. The other main feature was a large built-in cooker with four stoves set just in front of the door. Ash conoid deposits containing abundant botanical remains lay immediately at the foot of each stove; they were the result of brief accumulation, as their frequent removal was necessary for the use of the structure.
Just behind the stoves, a ‘slide’ was built from a double row of terracotta tiles which sloped down from the kitchen towards the river. A large heap of bones was found on this slide, held in place and sealed by a partially burned wooden roof beam whose collapse clearly resulted from the destruction of the abbey.
A second opening in the north wall of the kitchen close to the slide led to the access room of the refectory. Both the rooms described possessed beaten dirt floors.
Materials and methods (SC, AC, FS)
The rooms in the kitchen complex were excavated with particular care to avoid the loss of any relevant information and data. All the bioarchaeological remains detectable by eye were collected by hand in each stratum; in addition, whole deposits from particular contexts—such as the ante-kitchen drain, the pit, the slide and the stove conoids—were water-sieved (2 mm mesh) and floated to recover any smaller remains. A 5 per cent sample of the sediment from the earth floors was subjected to the same careful procedure. Moreover, a sampling grid (on a 1 m mesh) was set out in both rooms to take samples of the floors’ residual sediments for chemical analysis. In particular, concentrations of fatty acids, proteins and phosphates were sought (Carannante et al. Reference Carannante, Chilardi, Fiorentino, Pecci and Solinas2008).
The main steps of the bioarchaeological analyses were:
• Taxonomical and anatomical identification of all animal bones and botanical remains;
• Analysis of their stratigraphical and spatial distribution;
• Reconstruction of the ecology of each species;
• Determination of the age of death of the mammals represented by the animal bones, where possible;
• Identification of seasonal patterns in butchery, capture or gathering;
• Review of any cut marks and other anthropic modifications on the bones;
• Cross-comparisons with ancient textual sources.
In particular, a sample of 100 marine fish vertebrae and 50 opercular bones (i.e. the bony plates developed in and supporting the gill cover in most fishes) was analysed to assess their last growth rings (Wheeler & Jones Reference Wheeler and Jones1989) which allows definition of the season in which they were caught.
Bioarchaeological data
Several thousand bioarchaeological remains were collected from the different strata of the kitchen complex. Fish, bird, mammal and botanical remains were sorted and then examined separately by specialists. These remains are quantitatively described below. The composition of the vertebrate faunal assemblage shows that fish and bird remains represent 62 per cent and 25 per cent respectively of the studied bone fragments, whereas mammals amount to only 13 per cent. As will be argued, this unusual pattern of distribution is presumably connected with the diet permitted to the monks, who adhered to the Benedictine Rule, which forbids the consumption of red meat other than in exceptional cases of infirmity (Audoin Reference Audoin1986, 35; Marazzi Reference Marazzi, Marazzi and Gobbi2007, 21).
With regard to the archaeobotanical assemblage, 2981 carbonized seeds and charcoal fragments were identified. Nearly half (48 per cent) were grape pips, followed by elder/danewort seeds, legumes, cereals, and walnut and hazelnut shell fragments. Other taxa are each represented by less than 1 per cent of the overall assemblage.
Fish and mollusc remains (AC)
Several thousand fish remains were collected in the kitchen complex, mostly deriving from the rubbish-disposal devices such as the slide and the pit. Among the bony fish (Osteichthyes) remains, 2517 were identified, excluding spines, scales and the smallest fragments: 84.6 per cent pertain to fresh-water taxa (2131 NISP = Number of Identified Specimens), whereas a remarkable 15.4 per cent (386 NISP) are from marine or transitional waters species.
Cyprinidae form by far the best represented taxon (83.9 per cent of the whole Osteichthyes assemblage), with 2112 remains corresponding to 59 MNI (= Minimum Number of Individuals). Most of these pertain to tench (Tinca tinca, 400 NISP, a lacustrine and slow-flowing river species), but one Barbus sp. (a middle–high river course genus) is also attested. Remains of 18 trout (Salmo trutta, 3 MNI) and a single vertebra of large sea-eel (Anguilla anguilla) were also identified.
Among the marine and transitional water bony fish, Mugilidae (mainly Liza sp., grey mullet) are the most represented, with 321 remains corresponding to 22 MNI, followed by sea-bass (Dicentrarchus labrax, 60 NISP, 6 MNI) and Sparidae (5 NISP, 2 MNI, one at least Sparus aurata), such as gilt-head. All these species are typical of a brackish water environment, such as Mediterranean lagoons and estuaries. Chondrichthyes remains are also well represented in the fish assemblage, comprising mostly dermal plates of thornback ray (Raja clavata) (49 NISP, 9 MNI), found in several strata. Two large shark vertebrae were also found.
Few very small shellfish remains were recovered: three apical portions of Pirenella conica, one umbonal portion of Cerastoderma glaucum and one of Mytilus sp. All these species are characteristic of transitional water environments, especially of brackish Mediterranean lagoons. One of the three P. conica and the Cerastoderma were found in the slide heap deposit.
While only the bioarchaeological assemblage from the kitchen complex is considered in this paper, we must note that large quantities of fish bones excavated from the ancient river-bed just outside the kitchens—representing decades of abbey waste—are currently under analysis. Very preliminary data from this latter analysis confirm the findings above, although further species, such as well-represented sturgeon (Acipenser sp.), greater amberjack (Seriola dumerilii), grouper (Epinephelus sp.), gurnard (Trigla sp.) and John Dory (Zeus faber) were also found. Among the Chondrichthyes, in addition to thornback ray, stingray (Dasyatis sp.), angelshark (Squatina sp.) and at least five shark species are attested (Carannante Reference Carannante2015). Diverse ray and shark species are still used as food in southern Italy, while angel shark is no longer part of the diet.
Bird remains (SC, AC)
More than 1000 bird remains were collected in the kitchen complex; most came from the kitchen slide. Of these, 812 have been taxonomically identified, pertaining to nine taxa. The domestic fowl (Gallus sp.) is by far the most well-represented taxon, both in terms of NISP and MNI, with 639 remains corresponding to 46 individuals, followed by the pheasant (Phasianus sp.,152 NISP, 27 MNI). Other bird taxa are less well represented: rock ptarmigan (Lagopus muta, 8 NISP, 3 MNI), red-legged partridge (Alectoris rufa, 2 NISP, 1 MNI), domestic goose (Anser sp., 2 NISP, 2 MNI), brent goose (Branta bernicla, 2 NISP, 2 MNI), mallard (Anas platyrhynchos, 2 NISP, 1 MNI) and curlew (Numenius arquata, 2 NISP, 1 MNI). In addition, just three bone fragments of Passeriformes were found in the pit, representing just one individual of a small size, possibly a songbird, which presence could be accidental.
Most of the bird taxa are domestic or semi-domestic, but wild species are attested too. The only game birds attested are those found in areas ecologically similar to the site and its immediate surroundings—preferred habitats for the common partridge being rocky/sandy hills and mountainous areas, while curlews prefer wetlands, marshes and ponds. The presence of the rock ptarmigan is interesting since this species is nowadays limited to northern Europe, with just an isolated population found within the Italian Alps.
Only 11 bird bone fragments show cut marks, but this is not surprising as birds can often be disarticulated and defleshed without the use of blades. Only 17 bird fragments show traces of having been burnt on a fire, mainly on the distal parts of wings (radii and ulnae) and legs (tibiotarsii).
Interestingly, numerous eggshell fragments were also found in the kitchen complex, mainly concentrated in the pit, although some were also present in the floor layers.
Many other bird remains have come in excavation of the river bed. They are still under analysis but preliminary results show the presence of swan (Cygnus olor), common wood pigeon (Columba palumbus), grey partridge (Perdix perdix), common snipe (Gallinago gallinago) and grey heron (Ardea cinerea), in addition to bird species identified in the kitchen complex assemblage (De Falco et al. Reference De Falco, Pugliese, Soriga, Marazzi and Luciano2015).
Mammal remains (SC)
In contrast to fish and bird remains, few mammal bone fragments were present in the faunal assemblage of the kitchen complex. Despite sometimes being greatly fragmented, it was possible to identify 497 bone fragments, both taxonomically and anatomically; the remaining 62 could only be anatomically identified and were generically determined as belonging to mammals, using dimensional criteria (e.g. small, medium and large mammals).
Pig remains dominate the mammal assemblage, both in terms of NISP (310 fragments) and MNI (6 individuals), followed by Ovis/Capra (162 NISP, 4 MNI) and cattle (20 NISP but only 1 individual). Four fragments of equid bones and a single dog tooth—probably loose—complete this assemblage.
Pigs were butchered when young. Epiphyseal fusion in the limb bones shows that no individual older than three years is present in the assemblage; deciduous teeth are very frequent; no lower third molars were found; and only two third upper molars are present (both unworn and with roots unfused, hence attributable to individuals younger than 18–20 months).
Butchery marks are present on 36 bone fragments (6.4 per cent of the identified bones): both thin cut marks and heavier chop-marks are evident. Pig bones are the most useful for reconstructing meat-processing procedures, due to the size of the available sample. Cut marks show that the head was removed and the mandible separated from the rest of the skull. Butcher's knives and cleavers have left traces on the proximal and distal ends of the femur and on the tarsal bones, from when the hindlimbs were separated from the trunk and then the leg disarticulated from the foot. A similar pattern can be observed in the forelimbs, where cut marks are present on the proximal joint of humeri and the distal joint of radii. None of the vertebrae carries the typical chop-mark required to split them longitudinally, so the meat was probably removed as a piece on both sides of the vertebral column—a very common feature of ancient butchering in a rural context.
Fire traces from cooking are negligible and these data, combined with the butchery marks detected, seem to attest that the meat was usually stewed rather than roasted. The carcass was reduced to portions suitable to be put into pots or pans.
Roe deer (Capreolus capreolus), red deer (Cervus elaphus), wild boar (Sus scrofa) and hare (Lepus sp.) remains were found in the sediments from the river-bed excavation, pointing to an important role for game in the monastic economy (Chilardi Reference Chilardi, Marazzi and Luciano2015). In fact, several of these bone fragments showed some cut marks and were clearly the result of the disposal of kitchen waste over a longer time span. However, the game attested in the river dump, but absent among the remains of the last monastic meal, seems rather to suggest a role in banquets offered on special days to distinguished guests of the abbey who would not have been obliged to abide by monastic rules.
Botanical remains (FS)
An assemblage of 2981 carpological remains was collected from the kitchen complex (Carannante et al. Reference Carannante, Chilardi, Fiorentino, Pecci and Solinas2008). Most of these came from the waste disposal structures such as the drain and the slide, the pit, and the ash deposits at the feet of the stoves. Although there were 18 identified taxa—showing a good taxonomic variability—only eight of these reach a percentage higher than 1 per cent.
Vitis vinifera was by far the best represented (47.9 per cent) taxon with 1428 grape pips, followed by Sambucus sp. seeds (22.6 per cent). Pulses are also abundant with 339 remains. Most of these (10 per cent of the whole carpological assemblage) are faba beans (Vicia faba var. minor)—a small-seeded relative of the common broad bean—although lentils (Lens culinaris 1.2 per cent) and peas (Pisum sativum 0.1 per cent) were also identified.
Cereals were present with a total of 110 remains. Among these, emmer wheat (Triticum dicoccum) is the best represented (1.6 per cent of the whole carpological assemblage), followed by both the commonest varieties of naked wheat (T. aestivum/durum, 1.3 per cent) and barley (Hordeum vulgare, 0.8 per cent).
Nuts are also well represented in the carpological assemblage. Carbonized fragments (320) of walnut shells (Juglans regia, 10.7 per cent) are the most abundant, but hazelnut (Corylus avellana, 1.6 per cent), pine-kernels (0.3 per cent) and acorn fragments (only two) were also recovered.
Some olive-stones (0.5 per cent of the whole carpological assemblage) record the presence of this drupe at San Vincenzo. Other fruits are represented by 12 cherry-stones (Prunus avium), nine peach-stones (P. persica) and seven plum-stones (P. domestica). Three wild pear (Pyrus pyraster) pips were also found; wild pear trees still grow in the mountains around the abbey site.
The only green vegetable remains are some seeds of Atriplex hortensis—orache or mountain spinach. This wild edible plant is still collected in the Molise mountains where it is known as ‘orapi’ or ‘spinacio di montagna’ and used in the preparation of traditional rustic recipes.
To the archaeobotanical picture of the kitchen are added the preliminary data from the river-bed excavation, further attesting almonds and chestnut (Fiorentino & Solinas Reference Fiorentino, Solinas, Marazzi and Luciano2015).
The wood remains recovered belong to the carpentry structures of the pier and the jetty. This was composed of Sorbus domestica (1 per cent), Juglans regia (2 per cent), Ostrya carpinifolia (11 per cent), Ulmus minor (15 per cent), Quercus type deciduous (63 per cent) and 8 per cent of indeterminate fragments.
Results and discussion
Ecology of the monastic community (SC, AC, FS)
How was the monastic community of San Vincenzo able to obtain the different kinds of identified foods, from an ecological perspective? Some of them were obviously cultivated/bred in the immediate surroundings of the abbey, and this provides important information in itself.
Considering historical sources (cf. Oldoni Reference Oldoni2013), the Chronicon Vulturnense reports the long list of donations and properties bequeathed to the abbey before its 881 destruction. Among them are numerous fields with cereals and legumes, vegetable gardens, vineyards and olive groves with mills, woods for pig grazing and farms for breeding, fish ponds and structures used for fishing. The evidence reveals a vast network of possessions extending throughout southern Italy that provided animal and plant resources to the monastic community (see also Wickham Reference Wickham, Hodges and Mitchell1985).
Study of the territory in which the abbey is located can provide further additional inferences about the ecology of its community. San Vincenzo al Volturno lies on a fluvial terrace crossed by the upper reaches of the River Volturno and surrounded on the north and west by the uplands of the Mainarde Massif with Mt Marrone (1800 m asl). The terrace looks east onto a valley that leads to the Venafro plain (Fig. 5).

Figure 5. Map of the San Vincenzo hinterland. (1) The abbey's terrace with the upper reaches of the River Volturno; (1) the Mainarde massif; (3) the Venafro plain; (4) the Montecassino abbey; (5) River Volturno.
The monastic site, according to the phytoclimatic data of the Molise Region, (www.regione.molise.it/pianoforestaleregionale/sezione1b/ambiente_forestale_vegetazionale), is located in the temperate zone of the mountain-subalpine type; phytogeographically, the linked taxa are Quercus cerris (Turkey oak), Q. pubescens (downy oak), Q. frainetto (italian oak or farnetto), Acer (maple), Ostrya carpinifolia (black hornbeam), Fagus sylvatica (beech), Tilia plathyphyllos (linden) and Abies (fir).
Distributed along the banks of the main waterways (in this case, the Volturno river) we find hygrophilous and riparian species such as: willows (Salix purpurea, S. eleagnos, S. alba, S. triandra), poplars (Populus alba, P. canescens, P. nigra), field elm (Ulmus minor), dogwood (Cornus sanguinea) and hop (Humulus lupulus).
Some of the taxa present in the anthracological remains—in particular, Quercus, Ostrya carpinifolia and Ulmus minor—are also present in the present-day vegetation. However, it should be kept in mind that the anthracological assemblage is the result of human selection.
Undoubtedly, what has changed over time is the size of the forest cover of these species, since for centuries, many of these areas have become deforested and used for agriculture.
The area of the abbey is today characterized by riparian vegetation and by a Daphno laureolae/Quercetum cerridis vegetal association: mixed oaks wood including species such as Quercus cerridis, Q. pubescens and Q. ilex. This association corresponds to a temperate climate with snowy winters and relatively warm summers (Taffetani & Biondi Reference Taffetani and Biondi1993). These conditions are probably a bit different from the early medieval ones, since available palaeoclimatic data attest the occurrence of an early medieval cold phase (Lamb Reference Lamb1982, 150–52). This is confirmed by the presence of Lagopus muta in the kitchen complex faunal assemblage, showing that the distribution of this species extended into the central Apennines in the ninth century. In this scenario, vegetables and fruits—such as A. hortensis, P. pyraster, C. avellana and J. regia—would have been available nearby, as were most of the wild birds and mammals.
Domestic or semi-domestic birds—such as Gallus, Anser and Phasianus—and their eggs were indeed also local resources, as were the domestic mammals, not considering the supplies from the monastery's land ownership network.
The high percentage of pigs can be explained if we consider the site's geographical and ecological features. The breeding of swine does not need large open spaces for pastures. Various kinds of pork, cold meat and salami remain important goods in the local economy even nowadays. Oak woods are in fact a most suitable environment for pigs, which are natural forest dwellers, making their keeping the best option. Therefore, pork was the most common source of animal protein during the Roman period (King Reference King1999, 170) and perhaps the composition of the early medieval faunal assemblage can reflect the persistence of older alimentary practices. The S. trutta and Barbus also testify to the exploitation of the upper reaches of the river Volturno. However, other resources reached the abbey from ecosystems quite different from those of the monastic complex's immediate hinterland.
Mountainous areas over 1000 m—such as the Mainarde heights—provided habitats for L. muta, whereas lowlands contributed other resources to the abbey economy. Lakes, swamps and slow-flowing rivers provided habitats for N. arquata and for A. anguilla and T. tinca fishing. These kinds of ecosystems were abundant less than 20 km away in the Venafro plain, and several lakes such as the Gallo and Matese could be exploited at a greater distance (Fig. 6). Eel and tench could be also ‘farmed’ and temporarily kept alive in appropriate structures in the nearby river, although neither archaeological finds nor written sources attest to their existence in early medieval southern Italy.

Figure 6. Map with the main fishing/fowling areas. (1) Location of San Vincenzo al Volturno abbey; (2) Lake Gallo; (3) Lake Matese; (4) the Lesina lagoon; (5) the Varano lagoon; (6), Siponto; (7) the Patria lagoon.
The Venafro area was probably also the source of olives and grapes. Venafro is renowned, since Roman times, for its production of one of the best olive oils, as attested by many ancient authors such as Pliny (Naturalis Historia XV, 1–8), Martial (Epigrammata XIII, 108), Juvenal (Satyrae I, 5) and Horace (Odes et Epodes II, 6). Cereals, pulses and pine kernels were, in all probability, other foodstuffs available in the Venafro territory.
Some of the species identified in the faunal assemblage would, however, have to have come from further afield. Thus, a considerable percentage of marine/transitional water fish is attested in the faunal assemblage; this is interesting, considering that the site lies at about 560 m asl and over 70 km from the Tyrrhenian Sea and 100 km from the Adriatic coast.
As stated above (Fish and mollusc remains), most of the identified marine fish species are typical of brackish waters, as are the few molluscs species found in the kitchen complex. The presence of P. conica and C. glaucum—distinctive of shallow, brackish waters in particular—suggests that coastal lagoons were the most exploited ecosystems.
Historical evidence corroborates the archaeozoological record. The Chronicon Vulturnense refers to some fishing structures and areas situated in the Apulian lagoons of Lesina (160 km away) and Siponto (190 km away) being (with the fishermen too) among the abbey's properties, providing both cuttlefish and fish for the monastic meals—‘ad sippias prendendas pro refectione monachorum’ [destined to catch cuttlefish for the monks’ meal] (Oldoni Reference Oldoni2013, 164). Fishing areas in the Patria lagoon (100 km distant) on the Campania coast were also units of abbey land.
How did the marine fish reach San Vincenzo al Volturno? Some of them were clearly transported dried and/or salted. Cuttlefish, reported in the Chronicon as one of the most important marine products, were probably imported dried, according to Fiorillo (Reference Fiorillo, D'Arienzo and Di Salvia2010, 497).
The Chronicon in fact reports how the monastic community imported hundreds of dried fish, dried eels and prized botargo (dried fish roe, ‘ova tareca et secca’) from the Apulian lagoon of Lesina (Bresc Reference Bresc and Musca1987, 284). Mediterranean lagoons are the ecosystems where—at least since the second millennium bc—grey mullet was fished to extract the fish roes, then salted and dried to produce the highest-quality botargo. The archaeozoological evidence for the prevalence of mullet remains among the marine fish at San Vincenzo complements the importance of botargo among the abbey's resources attested by the written sources. Grey mullet could have reached the abbey as dried fish, as the Chronicon attests. On the other hand, the presence of very small mollusc remains (a few mm) suggests something different, since they had no nutritional value and furthermore showed the peculiar fragmentation pattern typical of molluscs digested by malacovore (shellfish-eating) fish such as gilt-head bream. S. aurata remains have indeed been found in the same strata. This means that shell fragments were passively transported to the abbey as the stomach contents of ungutted fish, which strongly suggests that some marine species—at least gilt-head—arrived at the abbey as fresh fish. The abundance of scales and cranial bones among the marine fish remains strengthens such a hypothesis, since, in order to preserve fish by salting and/or smoking, it is better to remove heads, scales and guts.
The lagoon fishing area owned by the abbey closest to San Vincenzo al Volturno was at Lake Patria, near Naples, over a 20-hour walk but much less on the back of a donkey or horse. The Apulian fish ponds on the Lesina Lagoon and on the Siponto Lagoon to the north and south of the Gargano Promontory were much more distant.
We can argue that, to supply the early medieval monastery with fresh fish, even on non-special days, required that loads departed weekly or monthly from the coastal possessions to the abbey, covering over 100–200 km in no more than three or four days (Fig. 6).
Monastic diet and economy (AC)
It seems clear that, based on the composition of the faunal assemblage being dominated by fish and birds, the Benedictine Rule heavily influenced the diet of the San Vincenzo monks.
The key to understanding the original spirit of the Rule lies in the ‘humoral theory’ of classical medicine (Adamson Reference Adamson2004; Mazzini Reference Mazzini, Flandrin and Montanari1996). This theory stated that foods are characterized by four basic qualities—cold, warm, moist and dry—that influence the humours of the human body—blood, phlegm, black bile and yellow bile—so determining a person's temperament—sanguine, phlegmatic, melancholic or choleric.
According to the view of classical medicine, a balance must be maintained between the four humours and diet could temper their disequilibrium; a varied diet was therefore desirable. This philosophy changed in medieval times as a consequence of a new social structure based on three classes: the warrior aristocracy, the clergy and the plebs. The assertion of class values determined diet, rather than the pursuit of equilibrium and health (Marazzi & Carannante Reference Marazzi, Carannante, Volpe, Buglione and De Venuto2010). Mammal meat—and particularly game—became the typical food for the aristocracy because of the association with blood, courage, aggressiveness and sexuality. This led to the renunciation of this kind of food by the members of the clergy to avoid aggressiveness and pride among its members, who saw in fish flesh the cold quality that enhanced the phlegmatic temperament required for a life of study, work and prayer. This explains why most monastic rules forbad mammal meat, although the Rule of St Benedict (Regula, 39) does allow fish and birds (Marazzi & Carannante Reference Marazzi, Carannante, Volpe, Buglione and De Venuto2010).
The archaeozoological data from San Vincenzo al Volturno demonstrate that the monks respected these prescriptions of the Rule. Nevertheless, the original Benedictine spirit was also to prescribe a simple way of life. So, how was this interpreted by the elitist class of the monastery, since, after all, the Chronicon reveals that many of the monks in San Vincenzo abbey once belonged to aristocratic, powerful and influential families? The presence of food such as fresh marine fish, botargo, cuttlefish, game birds, pheasants and piglets is indicative of a high social status—also corroborated by the preliminary data from the river-bed excavation (cf. Carannante Reference Carannante2015; Chilardi Reference Chilardi, Marazzi and Luciano2015; De Falco et al. Reference De Falco, Pugliese, Soriga, Marazzi and Luciano2015), which reveal that luxury foods such as sturgeon, large marine fish, heron and swan were processed in the monastic kitchens. The archaeozoological data therefore reveal that part of the monastic community kept its privileged way of life and ate the best luxury foods, even while respecting the Rule. In addition, red deer, roe deer and hare, attested in the river-bed assemblage, testify that distinguished guests—i.e. not clerical aristocrats, as attested by Chronicon Vulturnense— were honoured with luxury food permitted to them.
Topography and food processing (SC, AC)
How did the monastic community process the different kinds of resources?
As the bioarchaeological remains were not uniformly represented in the whole complex, it is necessary to examine their distribution patterns in detail to determine the function of each structure of the kitchens as well as each phase of food processing to which it related.
With regard to the ante-kitchen room, only a few mammal bone fragments were found, embedded in its floor. Several mammal remains (almost 21 per cent of the mammal assemblage) were, however, found in the drain that collected the water from the lavatorium and discharged it into the river, together with just a few bird and fish bones. Carpological remains (27.6 per cent of the botanical assemblage) are also well represented (Table 1).
Table 1. Number of Identified Specimens for each biological taxon in the main disposal devices of the San Vincenzo kitchens.

In the kitchen, bioarchaeological remains are very scarce in the contexts related to the hearth and stoves and also in the beaten floor strata, but 9.6 per cent of the botanical remains were recovered in the ash conoids at the base of the stoves. Meanwhile, 18.6 per cent of the identified fish remains, 47.8 per cent of mammal remains and 49.6 per cent of carpological remains come from the pit, together with the 2.6 per cent of the birds. Almost all the eggshell fragments were found in this unit.
The slide's faunal assemblage, to be viewed as relating to the last meal of the monks before the Saracen attack, is the richest in fish (76.4 per cent) and bird (85 per cent) remains whereas 15.2 per cent of the mammal remains come from this functional unit. Only 3.7 per cent of the carpological remains come from the slide (Table 1).
Interesting distributional patterns also emerge when considering the relative abundance of different anatomical segments in the kitchens’ contexts. For this purpose, among mammals, pig remains were chosen because of their statistical abundance. Using Binford's approach (Reference Binford1978; Reference Binford1984), the Minimum Animal Unit percentage was calculated (the results are summarized in Figure 7): the graph shows that anatomical distribution is similar in both drain and pit, while in the slide the pattern is different, almost complementary. Phalanges, metapodials, lumbar vertebrae, the atlas and axis were found only in the drain and the pit; conversely, femurs, humeri, radii, ulnae, tibiae, skull and mandibles were well represented among the slide remains (Fig. 7).

Figure 7. MAU percentage of the pig bones in the main disposal devices of the San Vincenzo kitchens: note the almost complementary distribution of the anatomical parts.
Fish remains show a comparable distribution. Excluding the drain, where they were poorly represented, 85.7 per cent of the vertebrae were concentrated in the slide, along with skull fragments (92.7 per cent), whereas other fish cranial bones and almost all the fish scales and ray-fish dermal plates came from the pit, with just 10.3 per cent of the fish vertebrae recovered therein.
The distribution of the anatomical segments of bird skeletons shows no significant pattern, being almost all concentrated in the slide. But it is worth noting that the few bird remains in the pit were essentially beaks and distal ends of wings.
How can these patterns of distribution be explained? Answering this question permits a reconstruction of both the function of each structure of the kitchen complex and the processes of meal preparation associated with its different areas.
In the northern area of the ante-kitchen, roughly dismembered mammals were further filleted and chopped in order to make smaller portions. The waste from these operations—bones and anatomical pieces with a low nutritional value, such as feet and filleted lumbar vertebrae—were disposed of in the drain. The archaeometric analyses of the floor samples confirm such a hypothesis. Different concentrations of fatty acids, proteins and phosphates in the beaten-earth floor reflect areas where meal preparation activities took place (Carannante et al. Reference Carannante, Chilardi, Fiorentino, Pecci and Solinas2008, 505). For example, the northeast corner showed a high concentration of protein close to the drain where mammal bones were disposed of. It reveals that the floor in that corner adsorbed abundant blood, thus confirming the butchery activities in the room.
The ante-kitchen was not only used for butchery. Archaeometric analyses (Carannante et al. Reference Carannante, Chilardi, Fiorentino, Pecci and Solinas2008, 505) revealed the probable use of the mysterious delimited space in the southern part of the room. Here, the southeast corner showed a second very high concentration of proteins, suggesting that it was used to hang fresh meat (letting the blood drip down). Conversely, very high concentrations of fatty acids and very low protein content characterize a narrow strip of the floor along the southern and the eastern side walls (Fig. 8). Our hypothesis is that this delimited space, protected by wooden partitions, was a sort of larder to preserve matured meat and where cheeses were shelved along the walls, oozing fat.

Figure 8. San Vincenzo: chemical analyses of the dirt-floor of the kitchen complex. Distribution of the concentrations of (a) protein; (b) fatty acids. (Adapted from Carannante et al. Reference Carannante, Chilardi, Fiorentino, Pecci and Solinas2008, 505, figs 6–7.)
Fish and birds do not seem to have been processed in the ante-kitchen, since their remains are barely represented here; instead, their preparation probably took place directly in the main kitchen.
Dirty food-processing activities took place over the pit below which ran the river. Here the fish was scaled, often beheaded and, less frequently, depending on the recipe, filleted. Here, too, mammal meat processing took place, and birds were sometimes decapitated and their wing-tips removed. Eggs, fruits and nuts were also often processed above the pit as attested by eggshells, walnut shells and fruit seeds found in its deposits.
The processed foods were then cooked on the stoves or on the hearth. The floor around these structures, as expected, showed the highest concentration of proteins and fatty acids.
Food was then served into the refectory. The bones found in the slide, which corresponded to the choicest parts of the vertebrates, suggest that the general waste from the refectory was discharged there. The slide was cleaned each day, by pushing the waste into the river, but—luckily for our archaeological understanding—the garbage was not thrown away on the day of the monks’ massacre (Fig. 9).

Figure 9. Reconstruction of kitchen activities in proximity to the slide and the built-in cooker at San Vincenzo in ad 881. (Drawing: Simona Carracillo.)
The last meal (SC, AC, FS)
The collapse of the roof beam which occurred during or just after the abbey's destruction froze in time the last moments of the kitchens’ life. The bone heap found on the slide consequently represents the ‘leftovers’ of the final meal of the monks, allowing us to reconstruct the meal of an early medieval monastic community on what would otherwise have been an ordinary day.
Several pulse and cereal remains suggest that a soup of wheat, emmer and faba beans was eaten during the meal. The main course apparently comprised both domesticated and wild birds (mainly fowl and pheasant, but also goose, duck, partridge and rock ptarmigan) and fish, both freshwater (mainly tench but also some trout and a barb) and marine/transitional (mainly grey mullet, with several sea-bass and also some gilt-head bream and two slices of a large shark). A small amount of mammal meat was seemingly also served, mainly pork.
Obviously, the most prestigious—but least attested in the faunal assemblage—birds and fish (wild feathered game and marine fish) were destined for the table of the abbot and the most powerful members of the monastic community and elite visitors, whereas the more common tench and poultry were the main course for the rest of the monks.
Finally, archaeobotanical remains show that pine kernels, hazelnuts and grapes completed the meal.
The presence of several elder seeds among the last supper remains, in addition to the large amount found in the kitchen pit, suggests that the monks used elderberries in various recipes (e.g. syrup, jelly, pies, sauces for duck and game).
Eggs, cheese, bread and green vegetables probably accompanied the meal, although remains of those could not be traced in the slide.
Valuably, some bioarchaeological remains found in the slide also allow us to hypothesize the season when the last meal was eaten. Seasonal analysis carried out on a sample of fish bones revealed that fish were caught during the autumn. This fact is corroborated by the presence of a brent goose: this is a migratory species that occasionally visits the Italian peninsula during very cold years in the autumn–winter (Brichetti & Fracasso Reference Brichetti and Fracasso2018). The presence of elderberries, grapes and hazelnuts matches well with the same season.
This fits the evidence of the Chronicon Vulturnense, which states that the Saracen raid happened during October of ad 881: this substantially confirms that the remains deriving from the slide are actually the waste of the last meal of the San Vincenzo monks.
Conclusions (SC, AC, FS)
This study can well represent what a sealed archaeological context, appropriately analysed with a multi-disciplinary approach, may offer when compared with ecological data and written sources, depicting the complex interaction between ecology, social status and religious prescriptions in a past human community.
The unique conditions of the site make the early medieval kitchen complex of San Vincenzo al Volturno a case study of general interest. The monastic community, in fact, was able to keep its wealthy and elitist status, while formally respecting the Benedictine Rule according to humoral theory. This was achieved by increasing the range of ecosystems from where food resources were obtained and also widening the spectrum of nutritional resources. The final result of this ecological strategy was a varied diet including some luxury foods, despite known religious restrictions. Moreover, the possibility of obtaining fresh marine fish, even on a non-festive day, at such a great distance from the coast, also highlights the wealth and ‘pull’ of the abbey.
However, the presence of fresh marine fish (often large in size), sturgeon, eel, cuttlefish, of wild birds and game is but a fraction—and a small one at that—of the whole extent of the identified foodstuffs. This pattern reflects the internal social composition of the monastic community.
Furthermore, the careful and rational space management of the kitchen complex indicates a well-organized community. A precise logic governed the management of the kitchens, from the food preserved in the larder and the butchery work, to the dirty operations of food processing, cooking and the final disposal of the waste. The condition of the earth floors, in which scant remains were found, reveals that the kitchens were cleaned regularly. The position of the lavatorium, in fact, placed between the kitchens and the refectory, confirms that hygiene was viewed as an important aspect of monastic life.
In summary, the bioarchaeological data related to the last meal at early medieval San Vincenzo reflect—even in its richness and complexity – what was in truth an ordinary day in these monks’ lives. The menu of the ‘last supper’ before the abbey's destruction thus gives us a unique image of daily life (from food procurement to consumption to discard) in what was one of the richest and most powerful monasteries in early medieval Europe.
Acknowledgements
We want to thank Silvia Cernuschi and Guido Tononi for the careful excavation of the kitchen complex and biological remains recovery that allowed detailed analysis of the contexts. This paper has also profited from the work of two anonymous reviewers who carefully read the manuscript. Their kind suggestions and their revision of the text were very important in improving the paper.










