Introduction
Our understanding of the impact of invasive plants has increased greatly with recent studies that have quantified their effects on plant communities (Houston & Duivenvoorden, Reference Houston and Duivenvoorden2002; Myers & Bazely, Reference Myers and Bazely2003; Schooler et al., Reference Schooler, McEvoy and Coombs2006; Gabbard & Fowler, Reference Gabbard and Fowler2007; El-Keblawy & Al-Rawai, Reference El-Keblawy and Al-Rawai2007; Pratt et al., Reference Pratt, Rayamajhi, Silvers and Ferriter2007) and ecosystem processes (Vitousek & Walker, Reference Vitousek and Walker1989; Bunn et al., Reference Bunn, Davies, Kellaway and Prosser1998; Allison & Vitousek, Reference Allison and Vitousek2004; Brooks et al., Reference Brooks, D'Antonio, Richardson, Grace, Keeley, DiTomaso, Hobbs, Pellant and Pyke2004; Yelenik et al., Reference Yelenik, Stock and Richardson2004). However, few studies have documented the negative effects of invasive plants on related biotic communities at higher trophic levels (Houston & Duivenvoorden, Reference Houston and Duivenvoorden2002; Herrera & Dudley, Reference Herrera and Dudley2003; Ernst & Cappuccino, Reference Ernst and Cappuccino2005; Willis & Memmott, Reference Willis and Memmott2005), and not all studies have detected a negative effect in terrestrial (Samways et al., Reference Samways, Caldwell and Osborn1996; Harris et al., Reference Harris, Toft, Dugdale, Williams and Rees2004) or wetland ecosystems (Douglas & O'Connor, Reference Douglas and O'Connor2003). In addition, to our knowledge, no prior study has compared the effects of two invasive plant species on these communities.
To respond effectively to invasive species problems, we require quantitative measures of the impact of invaders on diversity. The impact (I) of an invader depends on the invaders abundance (A), distribution (D) and per capita effect (E) according to the equation: I=A×D×E (Parker et al., Reference Parker, Simberloff, Lonsdale, Goodell, Wonham, Karieva, Williamson, Von Holle, Moyle, Byers and Goldwasser1999). Estimates of the abundance and distribution of invaders are widely reported, but estimates of per capita effects are rare (Parker et al., Reference Parker, Simberloff, Lonsdale, Goodell, Wonham, Karieva, Williamson, Von Holle, Moyle, Byers and Goldwasser1999; Schooler et al., Reference Schooler, McEvoy and Coombs2006), particularly for effects across trophic levels. Quantitative measures of the sign and strength of per capita effects are necessary to: (i) understand how invasive organisms affect natural environments, (ii) prioritize weed management actions and (iii) determine what levels of control are needed to achieve an acceptable level of impact. Here, we report results of an observational study assessing how severely two invasive plants, purple loosestrife (Lythrum salicaria L.: Lythraceae) and reed canary grass (Phalaris arundinacea L.: Poaceae), reduce wetland herbivore diversity with increasing invader abundance.
Purple loosestrife and reed canary grass are two invasive emergent plants that are spreading and increasing in abundance in many wetlands across temperate North America (Galatowitsch et al., Reference Galatowitsch, Anderson and Ascher1999; Magee et al., Reference Magee, Ernst, Kentula and Dwire1999; Blossey et al., Reference Blossey, Skinner and Taylor2001). Purple loosestrife is native to Europe (Mal et al., Reference Mal, Lovett-Doust, Lovett-Doust and Mulligan1992), whereas the origin of reed canary grass is in dispute. It is likely that the invasive variety of reed canary grass is a hybrid of North American and European stock (Galatowitsch et al., Reference Galatowitsch, Anderson and Ascher1999; Lavergne & Molofsky, Reference Lavergne and Molofsky2004). Both species are known to form dense monospecific stands that negatively affect local wetland plant communities (Galatowitsch et al., Reference Galatowitsch, Anderson and Ascher1999; Merigliano & Lesica, Reference Merigliano and Lesica1998; Blossey et al., Reference Blossey, Skinner and Taylor2001; Schooler et al., Reference Schooler, McEvoy and Coombs2006). However, prior studies have not examined the potential impact of purple loosestrife and reed canary grass on biotic diversity at higher trophic levels.
We use the moth community (Lepidoptera) as a biological indicator of the effect of two invasive plant species on the herbivore trophic level in wetland communities. Characteristics of an ideal biological indicator species are: (i) practical to monitor, (ii) sensitive to the stressor of interest, (iii) ubiquitous, (iv) short generation times and (v) play a key role in the functioning of the community (Parker et al., Reference Parker, Simberloff, Lonsdale, Goodell, Wonham, Karieva, Williamson, Von Holle, Moyle, Byers and Goldwasser1999). Moths exhibit the first four of these traits and arguably the fifth (Foote et al., Reference Foote, Kadlec and Campbell1988; Goyer et al., Reference Goyer, Lenhard and Smith1990). Moths are efficiently sampled by light traps; and, as consumers of living plants, they are likely to be sensitive to the vegetation changes caused by plant invaders (Janzen, Reference Janzen1987; Pinheiro & Ortiz, Reference Pinheiro and Ortiz1992). They have previously been found to be good indicators of environmental conditions (Kremen, Reference Kremen1992; Luff & Woiwod, Reference Luff, Woiwod, Harrington and Stork1995) and are known to be almost exclusively herbivorous and relatively host specific (Brues, Reference Brues1920; Brues, Reference Brues1924; Dethier, Reference Dethier1952; Ehrlich & Raven, Reference Ehrlich and Raven1964; Janzen, Reference Janzen1987; Mitter & Farrell, Reference Mitter, Farrell and Bernays1991; Young, Reference Young1997). In addition, prior studies have found a positive correlation between the species richness of plants and Lepidoptera (Thomas & Mallorie, Reference Thomas and Mallorie1985; Hawkins & Porter, Reference Hawkins and Porter2003).
We expected moth populations to respond to changes in local plant community composition. Since herbivore diversity generally increases with plant diversity, it is expected that increasing abundance of an invasive plant will reduce plant diversity and result in a reduction in herbivore diversity. Therefore, we expected that herbivore diversity would decline with increasing abundance of the invasive plant species. There are two components to this hypothesis: (i) moth diversity will be positively correlated with plant diversity and (ii) invasive plant abundance will be negatively correlated with moth diversity. We were particularly interested in how the two plant species might differ in their effects on the diversity of the herbivore community.
Biotic diversity is a complex measurement; and different invasive species may have a different effect on the two components, the number of species (species richness, S) and the equitability in abundance among species (evenness, J) (Whittaker, Reference Whittaker1972; Magurran, Reference Magurran1988). These descriptors can be measured independently or combined into a diversity index. Two commonly used indices are Brillouin's index (H) and Simpson's index (D) (appendix 1; Magurran, Reference Magurran1988). Brillouin's index is a measure of the information content of the sample and is most strongly affected by the species in the middle of the sequence. It is, therefore, not as sensitive to the abundances of the first few dominant species as compared with Simpson's index (Whittaker, Reference Whittaker1972). Simpson's index is similar to Brillouin's index in that it integrates species richness and abundance but is strongly influenced by the abundances of the first few dominant species and can, therefore, be regarded as a measure of dominance (Whittaker, Reference Whittaker1972). We used these four diversity measures (S, J, H, D) to examine herbivore communities at wetland sites at various stages of invasion by two invasive wetland plants.
Material and methods
Study sites
Twenty wetland field sites were selected in the Pacific Northwest USA. Sixteen sites were established in the Willamette Valley (Oregon) and four sites were located east of the Cascade Mountain Range along the Columbia and Snake rivers, two in Oregon and two in Idaho (fig. 1). The 20 sites were seasonally flooded emergent palustrine wetlands. The vascular plant community at each site was dominated by herbaceous vegetation.

Fig. 1. Map of locations for the 20 study sites in and around Oregon, USA. Thin lines are county boundaries. Symbols denote the dominant plant species within each study site (■, purple loosestrife; •, reed canary grass; ▲, other).
Plant sampling
Percent plant cover was measured for each vascular plant species at each of the 20 sites during July 2001. Each sample universe was circular with a 50 m radius (7854 m2). We based the size of the area of vegetation sampled on the effective radius of the light trap. Effective radius differs among Lepidoptera species and gender; however, several prior studies estimate the effective radius to be less than 50 m for various moth species, including Spodoptera littoralis, Manduca sexta and Heliothis zea (Plaut, Reference Plaut1971; Baker & Sadovy, Reference Baker and Sadovoy1978; McGeachie, Reference McGeachie1988; Young, Reference Young1997). For example, only 1.4% of S. littoralis released from 50 m distance were recaptured in the light trap (Plaut, Reference Plaut1971). At each site, thirty 1-m2 plots (square) were randomly selected from within 50 m of the center point. This was done by randomly selecting a direction (1–360°, with North at 360°) and a random distance (0–2500) of which the square root of the result was the radial distance (m) to the plot. This permitted sampling the circular site in proportion to area. The percent cover of each plant species was measured in each plot. Species identifications and nomenclature follow Hitchcock & Cronquist (Reference Hitchcock and Cronquist1973). A voucher collection of the plant species sampled was assembled and specimens were deposited in the Oregon State University Herbarium.
Moth sampling
The moth community at each site was sampled using an 8-watt blacklight trap (Ward's Natural Science, Rochester, NY) with a diclorvos fumigant strip (Revenge®: 2,2-Dichlorovinyl dimethylphosphate, BioQuip, Rancho Dominguez, CA). The light traps were located at the center of the site and elevated on a metal post so that the light was above the vegetation canopy. Each light trap sample was collected the following day to prevent decay of moth specimens. Sites were sampled synchronously for three moonless nights each month from May through August, resulting in a total of 240 samples collected in 2001. After collection, the moth samples were frozen and transported to Corvallis for identification. Species identifications were made using the Oregon State University Arthropod Collection (OSAC) and verified by P. Hammond. The OSAC collection identifications are based on various taxonomic authorities in various groups, technical publications (e.g. Miller & Hammond, Reference Miller and Hammond2000) and Moths of America North of Mexico including Greenland Series (1971–2004). A voucher collection of the moths was assembled and deposited in the OSAC.
Moth community analyses
The effect of invasive plant abundance on the moth community was examined in six steps. First, we visually examined the structure of the moth communities using rank-abundance curves (Whittaker, Reference Whittaker1972). Rank abundance curves visually integrate the two components of biotic diversity; the number of species and the relative abundance among species. Second, sampling adequacy was examined by constructing species-area curves for each site (McCune & Grace, Reference McCune and Grace2002). Third, we examined the relationship between plant species richness and the species richness of the moth community using linear regression (S-plus, version 6.1, Insightful 2002 and Excel, Office XP, Microsoft 2002). Fourth, we assessed whether the effect of the two plants on moth diversity differed using extra-sum-of-squares F-tests (Ramsey & Schafer, Reference Ramsey and Shafer1997). This test compares the variability of a full model (separate regression models, one for each the two focal invasive plant species) with the variability of a reduced model (single regression model combining data for both plants). If the residual variability does not differ between the full and reduced models, then the effect of the plants on moth diversity was considered to be similar. Where no difference was found, we pooled the data for the two species to analyse the significance of the effect (thereby gaining power to determine significance of impact). Fifth, we examined the association between invasive species abundance and four standard diversity descriptors: species richness (S), species equitability (J), Brillouin's index (H), and Simpson's index (D) (mathematical definitions in appendix 1 and Magurran (Reference Magurran1988) ) using regression analyses. Sixth, we examined the effect of the invasive species on individual moth species, using canonical correspondence analysis (CCA) (PC-ORD, version 4.17: McCune & Mefford, Reference McCune and Mefford1999). CCA constrains an ordination of one matrix (moth species abundance) by the variables in a second matrix (invasive plant abundance). Final scores from the analysis are an indication of the association between the axes (invasive plant abundance) and moth species abundance. Transformations were used to improve normality and homogenize variance where necessary (described below). Success of transformations was determined by examining residual and normal probability plots.
Environmental measurements
Measurements of the species richness and diversity of moth communities are influenced by both environmental variables and sampling bias. Light trap catch is shown to be influenced by: (1) species of insect (Stewart et al., Reference Stewart, Lam and Blythe1969; McGeachie, Reference McGeachie1988), (2) sex of insect (Stewart et al., Reference Stewart, Lam and Hoffman1967; Persson, Reference Persson1976), (3) design of trap (Intachat & Woiwood, Reference Intachat and Woiwood1999), (4) light intensity (Taylor & Brown, Reference Taylor and Brown1972; Young, Reference Young1997), (5) light source (Bowden, Reference Bowden1982; Nabli et al., Reference Nabli, Bailey and Necibi1999), (6) distance to trap (Stewart et al., Reference Stewart, Lam and Blythe1969; Plaut, Reference Plaut1971), (7) ambient light (Yela & Holyoak, Reference Yela and Holyoak1997), (8) meteorological conditions (wind speed, wind direction, rainfall and cloud cover) (Young, Reference Young1997) and (9) temperature (Yela & Holyoak, Reference Yela and Holyoak1997). Influential sampling factors 3–7 were minimized by using the same type of trap during synchronous new moon sampling periods for all sites. In addition, we measured the most important environmental variables (factors 8 and 9): wind speed, cloud cover, rainfall, temperature, ambient light and land-use, using the following methods.
Temperature is likely to be the most influential factor that affects light trap catch. Air temperature was recorded when the traps were set and again when samples were collected. A max.–min. thermometer (Taylor 5458, Oak Brook, IL, USA) was attached to each trap (north side in shade of trap), and these temperatures were recorded when the traps were serviced the following day. Wind speed was measured at dusk on three trap nights for each site using a digital wind speed meter (Kestrel 1000, Nielsen-Kellerman, Chester, PA, USA). In addition, presence or absence of rainfall and an estimate of percent cloud cover were recorded for each trap event. Light intensity measurements were taken on a new-moon night in August 2001 for sites in eastern Oregon and in September 2001 for Willamette Valley sites. A standard photographers' light meter (Gossen Luna-Pro, Bogen Inc., London, UK) was used to measure ambient light intensity (lux) of the night sky (22:00–02:00) at nine points (horizon (N, E, S, W), 45° (N, E, S, W) and 90° (directly overhead). The mean lux of these nine locations was used as the measure of nocturnal light intensity at each site. The correlation among illumination, temperature and invasive species abundance was examined using regression analyses.
Land-use using GIS spatial datasets
Aside from the composition of the host plant community, land-use surrounding the sites may influence local moth populations (Ricketts et al., Reference Ricketts, Daily, Ehrlich and Fay2001). In particular, increasing residential and urban percent cover is likely to decrease moth species richness and diversity by decreasing surrounding host plant habitat, inhibiting colonization from neighboring sources, and causing sampling and behavioral interference due to artificial night illumination (Luff & Woiwod, Reference Luff, Woiwod, Harrington and Stork1995). Ricketts et al. (Reference Ricketts, Daily, Ehrlich and Fay2001) found that a ‘halo’ of increased moth species richness extended from 1.0–1.4 km from the forest edge into agricultural fields; and, therefore, agricultural sites within 1.0 km of a forest fragment had significantly greater species richness and abundance than sites farther than 3.5 km from the forest edge. We expected the local moth communities to react at similar scales and have, therefore, quantified land-use composition in a series of ten concentric radii up to 3000 m around each site. We used a GIS-based land-use data layer (imagery collected in 1993) to calculate percent urban land-use in a series of ten concentric circles with increasing radii (100, 200, 300, 400, 500, 1000, 1500, 2000, 2500 and 3000 m) centered about each light trap location. At each scale, the percent of the area in each of four land-use categories was summarized as forest, natural non-forest, agricultural and urban (after Shaffer et al., Reference Shaffer, Kentula and Gwin1999) for each site. Spatial data were collected using a Trimble Surveyor (XR Pro) global positioning system receiver (GPS) with real-time and post-processing differential correction capable of sub-meter spatial accuracy. ArcView (version 3.2, ESRI 1999) was used to summarize land-use spatial datasets. The relationship between land-use and invasive species abundance was analysed using canonical correspondence analysis CCA (PC-ORD, version 4.17: McCune & Mefford, Reference McCune and Mefford1999).
Results
Plant community
The overall plant community composition of the 20 wetland sites consisted of 162 vascular plant species distributed within 35 families (Schooler et al., Reference Schooler, McEvoy and Coombs2006). Seven plant species were dominant (most abundant in the community) across the 20 wetland sites. Purple loosestrife was the dominant plant at six sites (13.8–91.7% cover), reed canary grass was dominant at six sites (18.8–94.7%) and the remaining eight sites were dominated by five other plant species (12.5–47.8%) (fig. 2a). The other dominant species consisted of three monocots (Alopecurus pratensis, Juncus effusus, Typha latifolia) and two dicots (Oenanthe sarmentosa and Veronica americana).
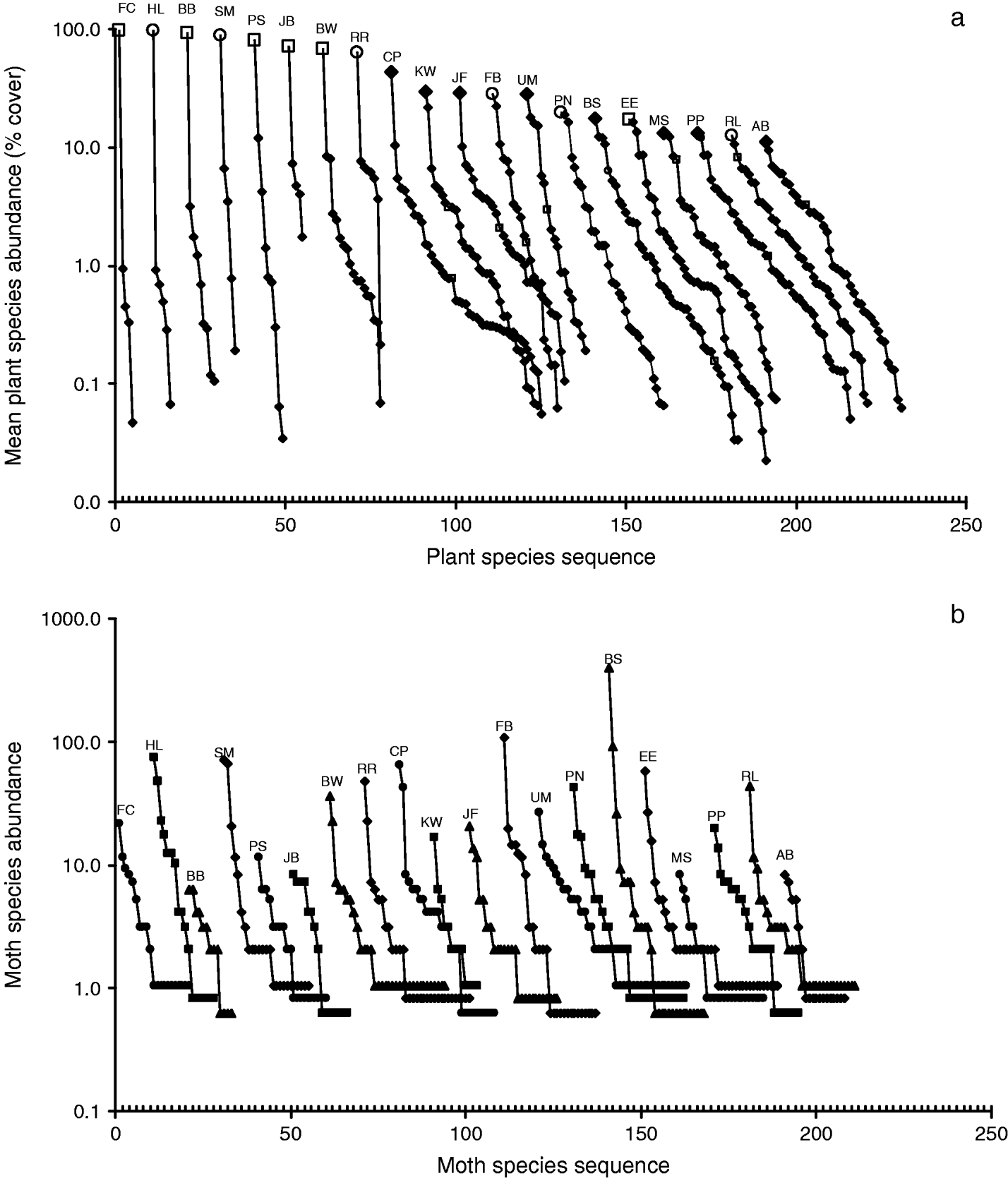
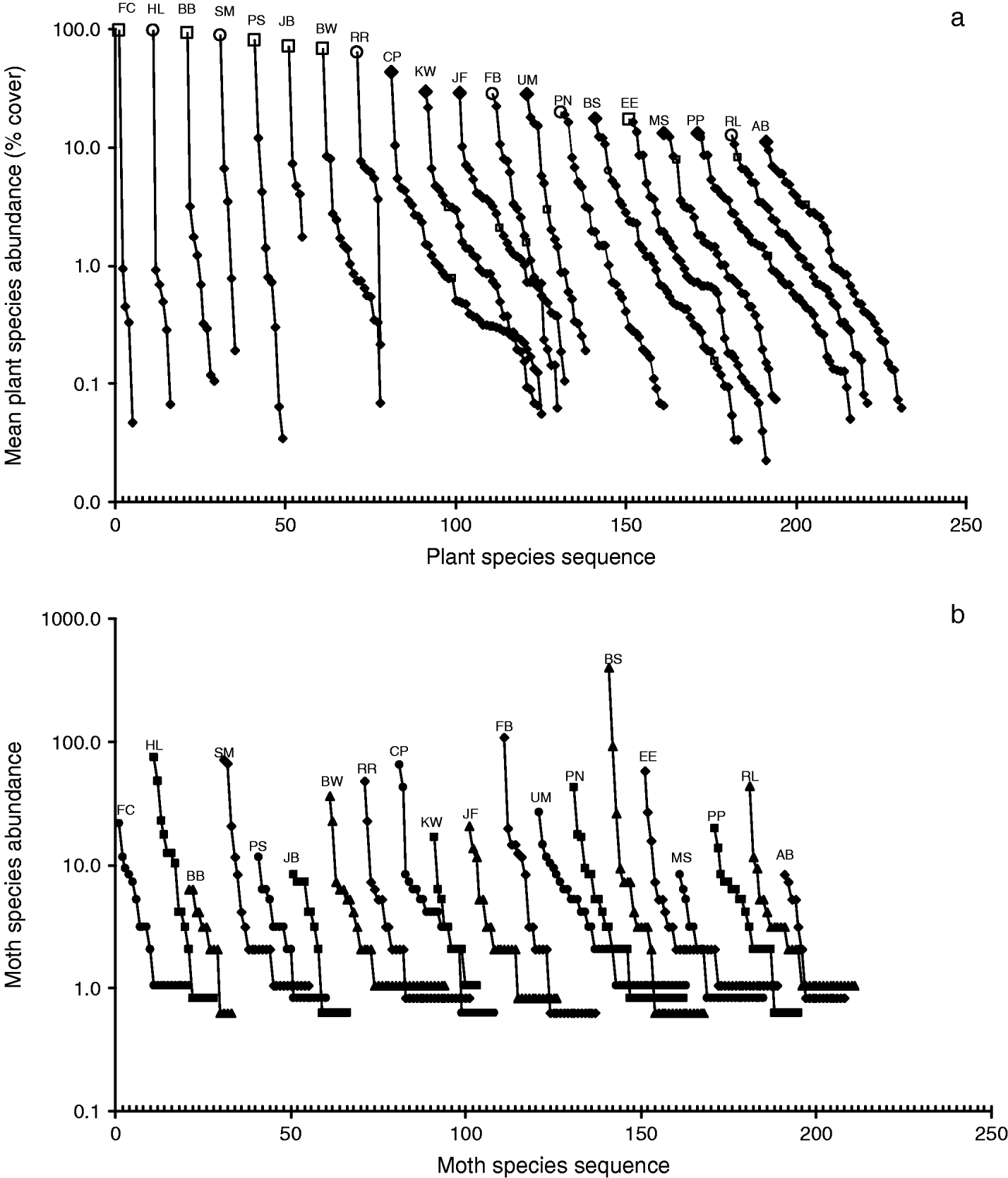
Fig. 2. Rank-abundance curves for the (a) plant and (b) moth communities at 20 wetlands in the Pacific Northwest. The curves, separated by ten unit intervals, start at various points on the species sequence in order to fit them into the same figure. (a) Mean percentage cover values of plant species are plotted for the sequences from the most to the least abundant species for each site. Curves are labeled according to site and are arranged in the order of decreasing abundance of the dominant plant species. ○, purple loosestrife; □, reed canary grass; ◆, other species. (b) Moth community sequences are arranged by site in the same order as plant sequences. Symbols are varied and abundance values about one (single individuals of a species at each site) are varied about one to discriminate among sites.
Moth community
The regional wetland moth community sampled at the 20 sites consisted of 156 moth species (fig. 2b, appendix 2) distributed across 14 families (see Schooler, Reference Schooler2003, for raw data). A total of 2744 individual moths were collected. Eight individuals could not be associated with a particular moth species, although they were identified to either genus (4) or family (4). The four families with the most species, including 88% of all the moth species, were the Noctuidae (88 species), Geometridae (31), Pyralidae (26) and Arctiidae (10). Willamette Valley collections consisted of a total of 2048 moths (75%), while eastern Oregon sites yielded 696 moths (25%). In the Willamette Valley, Aletia oxygala (a grass feeding noctuid) was the most abundant species at seven sites (22% of moths sampled on the Willamette Valley) and Euchromis ocelleus (a grass feeding pyralid) was most abundant at five sites (32%). Other abundant species at the Willamette Valley sites included: Chrysoteuchia topiaria (4%), Crymodes devestator (4%) and Lithacodia albidula (3%). The most abundant species sampled at the eastern Oregon sites included: Petrophilia confusalis (24% of moths collected at eastern Oregon sites), Tehama bonifatella (10%) and Xestia c-nigrum (8%).
Native species comprised 91% of the regional wetland moth taxa (142 species) while 4% (6) were exotic moth species. The native status was not determined for the eight remaining taxa (5%) because they could not be identified to species. Exotic moth species collected were: Apamea ophiogramma (0.4% of total), Caradrina morpheus (0.07%), Idaea dimidiata (0.9%), Noctua comes (0.1%), Tyria jacobaeae (0.3%) and Xestia xanthographa (0.5%).
Moth community analyses
We first qualitatively compared the diversity of the plant and moth communities using rank abundance curves (fig. 2). Plant species richness and evenness decreased with increasing abundance of the dominant plant species (fig. 2a). Moth community curves exhibited a negative exponential shape with a few very abundant species and many rare species (fig. 2b) with 44 species (28%) represented by a single individual.
Second, species-area curves were plotted for the moth data to determine whether the sampling effort was adequate to accurately sample the species diversity of the local moth community. The accumulation of species did not reach a plateau, although the rate of accumulation decreased with increasing sampling effort (fig. 3). The curves were generally the same shape with each curve majorized by another, with the exception of three sites that crossed the values of neighboring curves. During sampling, we noted that one site (Knez Wetland; KW) was near a large illuminated billboard. Ambient night illumination at this site was 2.0 lux, which was 13.3 times greater than the mean night illumination of the other 19 sites (mean=0.15 lux, SD=0.06). This probably decreased the trap catch. A total of 45 individual moths were captured at this site during 2001, while the mean trap catch among the other sites was 3.2 times greater (mean=142.1 moths, SD=117.4). Therefore, we removed the data from this site from the quantitative diversity analyses.

Fig. 3. Species accumulation curves for moth collections. Each curve represents the number of new species found with increasing sample effort over the 12 sampling events at each site.
Third, we examined whether plant species richness was linked with moth species richness. We found that the number of plant species was positively correlated with the number of moth species among the 19 study sites (fig. 4) (t 18=2.20, P=0.04). Because the single individuals representing a moth species may be migrants passing through our study area, we analysed a second dataset with these species removed to determine whether they affected the result. The relationship between species richness of plants and herbivores was similar with and without these single representatives of a species included in the analysis (R2=0.15, t 18=2.59, P=0.02).

Fig. 4. Relationship between moth species richness and plant species richness at the 19 wetland field sites. The number of plant species is the total number of vascular plant species recorded at each 7854 m2 site. The number of moth species is the cumulative number of species trapped over 12 sampling events (three nights each month over four months, May–August 2001).
Fourth, we determined whether there was a difference in effect between the two invasive plant species. Comparison of the regression models indicated that regressions of diversity on invader abundance, pooling invader species, performed as well as regressions separating the two invader species for the four moth diversity measures (table 1). This suggests that the two plant species are similarly affecting moth diversity. Finding no difference between invader species in the effect of invader abundance on moth diversity, we pooled data for the two invasive plants in subsequent analyses. Data were transformed (S and D, natural log; H and J, square root) before analysis, which homogenized variances.
Table 1. Test of difference in relationships between loosestrife and canary grass abundance and moth diversity.

S, species richness; H, Brillouin's index; D, Simpson's index; J, community equitability.
The P-value indicates whether the effect was different between the two plant species (ln, natural log transformation; sqrt, square root transformation).
Fifth, we examined whether abundance of the invasive plant species was correlated with any of the four measures of moth diversity. We found that moth species richness decreased with increasing abundance of the invasive plant species (fig. 5a) (t 18=2.86, P=0.01). However, we found no relationship between invasive plant abundance and the other three measures of moth diversity (J: t 18=0.51, P=0.62, fig. 5b; H: t 18=−0.87, P=0.39, fig. 5c; D: t 18=−0.19, P=0.85, fig. 5d). Simpson's index (D) required a natural log transformation to homogenize variance.

Fig. 5. Relationship between diversity of the moth community and abundance of the invasive plant species. The abundance of the dominant plant is the mean percent cover of the most abundant plant species. Moth community diversity is described as (a) the number of species, (b) the equitability among species, (c) Brillouin's index of diversity and (d) Simpson's index of diversity. Simpson's index was natural log transformed to improve homogeneity of variance (□, purple loosestrife; ○, reed canary grass; △, other plant).
Sixth, we examined which moth species were most affected by the abundance of the two invasive plants. The CCA consisted of two matrices, one containing the abundance data of the 156 moth species (natural log transformed) by site and the second contained the percent cover of the two invasive plant species (arcsine square root transformed). The first axis was primarily correlated with purple loosestrife (r=0.99) and the second was primarily correlated with reed canary grass (r=0.93). We found that the abundances of 104 of the moth species (66%) were negatively correlated with the cover of purple loosestrife (axis 1) while 93 species (60%) were negatively correlated with the cover of reed canary grass (axis 2). In addition, 65 moth species (42%) were negatively correlated with both invasive plant species (appendix 2). Many of the moth species (44) were represented by a single individual and are possibly transient species. However, removing these species did not change the ratios. With these 44 species removed, 72 species (64%) were negatively correlated with purple loosestrife and 67 species (60%) were negatively correlated with reed canary grass. Disregarding the species represented by a single individual, the species most negatively correlated with purple loosestrife were Heliothis phloxiphaga (Noctuidae), Lacinipolia patalis (Noctuidae) and Clostera apicalis (Notodontidae). The species most negatively correlated with reed canary grass were Acronicta marmorata (Noctuidae), Cosmia calami (Noctuidae) and Cyclophora dataria (Geometridae).
We also selected six moth species to examine more closely, based on their feeding habits. Two species of noctuid grass feeders were common in the study: Aletia oxygala and Crymodes devastator. Although both feed on grasses, A. oxygala was present in greater abundance (452 individuals, 16% of individuals sampled) than C. devastator (90 individuals, 3%). We also noted that four rare species of moths (Archanara alameda (8 individuals, 0.3%), A. oblonga (eight individuals, 0.3%), A. subflava (20 individuals, 0.7%) and Bellura obliqua (one individual, 0.04%)) are known to specialize on cattails (Typha spp.), rushes (Juncus spp.) and sedges (Carex spp.). These moth species were only found in reference sites with very low abundance of the two invasive species.
Environmental variables
To help isolate the influence of invader abundance on moth diversity, we examined whether the abundance of the two invasive plant species covaried with potentially confounding variables, including temperature, night illumination, precipitation, wind speed and land use. Minimum monthly mean night temperature during trap events (n=3 nights per month) varied from 2.9 to 17.2°C depending on the month and site. There was no relationship between invasive species abundance and minimum night temperature for any of the four months sampled (May: t 18=0.09, P=0.93; June: t 18=0.27, P=0.79; July: t 18=0.69, P=0.50; August: t 18=0.39, P=0.70). Illumination varied from 0.10 to 0.32 lux with a mean of 0.15 lux (SD=0.06). Most sites had very low ambient illumination during new moon nights (except KW as described earlier). No relationship was found between illumination and invasive species abundance (t 18=0.24, P=0.81). There was no precipitation during the nights when moths were sampled. Wind speed was measured at the sites when the traps were set in the evenings and at dawn when the moth samples were collected. All wetlands were in depressions in the landscape. This topography caused winds to be so slight that they rarely registered on the wind speed meter (<0.3 m sec−1).
The land cover surrounding the wetland sites is expected to influence the species richness and abundance of the moth community. We used CCA to determine whether land-use trends were correlated with the abundance of the invasive plant species (McCune & Grace, Reference McCune and Grace2002). The main data matrix consisted of the 19 sites, each with four surrounding land-use categories (forest, natural non-forest, agricultural and urban) at the ten spatial scales (100, 200, 300, 400, 500, 1000, 1500, 2000, 2500 and 3000 m) for a total of 19 sites and 40 variables. The second matrix contained the mean proportional cover of purple loosestrife and reed canary grass for the sites (2 species and 19 sites). Normality was improved using an arcsine square root transformation on the proportional cover data. The abundances of the two invasive species comprising the second matrix were negatively correlated (r=−0.41). Axis one was primarily correlated with the abundance of purple loosestrife (r=−0.98), while axis two was correlated with the abundance of reed canary grass (r=−0.82). The Monte Carlo test statistic indicated that the relationship between loosestrife abundance (axis 1) and the land-use variables was not significantly greater than expected by chance (P=0.12). The relationship between reed canary grass abundance (axis 2) and the environmental variables was also not significant (P=0.49).
Discussion
We found that moth species richness was positively correlated with plant species richness. In addition, we observed a negative correlation between moth species richness and the abundance of the invasive plant species. This is most likely the result of the reduction in the diversity of the plant community caused by the increasing abundance of the invasive plants (Schooler et al., Reference Schooler, McEvoy and Coombs2006). The negative association between invasive plant abundance and moth species richness was similar for both plant species. The linear relationship between invasive plant abundance and moth diversity has a constant, negative slope (per capita effect), meaning that any management actions that reduce the abundance of the two invasive plant species will increase the species richness of the moth community.
However, invasive plant abundance appeared to have no effect on three other measures of moth diversity: community evenness, Brillouin's index and Simpson's index. These measures of moth community diversity include moth abundance in order to estimate the equitability (or evenness) among the species of the community (appendix 1). Samples of many of the reference wetlands had a large number of individuals of a single moth species that dwarfed the abundance of moths of the other species, thereby decreasing measurements of equitability (fig. 2b). The extreme case was the Baskett Slough site (BS), on a US Fish and Wildlife Service National Wildlife Refuge. We sampled 562 individuals distributed among 28 moth species. The grass feeding moth Euchromius ocelleus (Noctuidae) was very abundant, such that we trapped 385 individuals over the sampling period (69% of all moths collected at that site) (fig. 2b). The next most populous moth species were Aletia oxygala (89 individuals, 16%) and Crymodes devastator (25 individuals, 4%). The remaining 25 species added a total of 63 individuals (11%) to the samples, resulting in an evenness score of 0.38 (evenness scores vary from 0–1, with 1 being equal abundance across all species) (fig. 5b). These trends made the abundance of moth species within the communities very uneven when compared to communities that had fewer individuals and rare species. For example, at the Bird Blind site (BB), with a mean of 87.4% reed canary grass, we trapped a total of 36 individuals of 13 species (fig. 2b), resulting in an evenness score of 0.93 (fig. 5b). These moth community patterns resulted in estimates of evenness that showed no trend with increasing abundance of the invasive plant species. The Brillouin and Simpson indices of diversity showed similar results (fig 5c, d), probably because they both incorporate species equitability into the calculation.
Our results are consistent with those of prior studies that document the expected positive relationship between the species richness of plant and herbivore communities (Murdoch et al., Reference Murdoch, Evans and Peterson1972; Strong et al., Reference Strong, Lawton and Southwood1984; Thomas & Mallorie, Reference Thomas and Mallorie1985; Erhardt & Thomas, Reference Erhardt, Thomas, Collins and Thomas1991; Kremen, Reference Kremen1992; Kremen et al., Reference Kremen, Colwell, Erwin, Murphy, Noss and Sanjayan1993; Luff & Woiwod, Reference Luff, Woiwod, Harrington and Stork1995; Siemann et al., Reference Siemann, Tilman, Haarstad and Ritchie1998; Hawkins & Porter, Reference Hawkins and Porter2003; Zhao et al., Reference Zhao, Fang, Peng and Tang2006). Fewer studies have examined negative associations between invasive plant abundance and herbivore diversity, and prior results are sometimes contradictory. A study of an invasive vine (Vincetoxicum rossicum) in northeastern North America found fewer arthropods in most feeding guilds, including herbivore guilds, than those found on nearby stands of three common native plants (Ernst & Cappuccino, Reference Ernst and Cappuccino2005). In addition, a study on the impact of giant reed (Arundo donax) found a reduction in the species richness of aerial invertebrate communities within stands of the invasive plant when compared with collections from native vegetation (Herrera & Dudley, Reference Herrera and Dudley2003).
However, a study on an invasive shrub, gorse (Ulex europaeus), in New Zealand found that the gorse habitat was more species-rich for Tachinids, fungus gnats and Malaise-trapped beetles when compared with the habitat of a native shrub (Kunzea ericoides) (Harris et al., Reference Harris, Toft, Dugdale, Williams and Rees2004), perhaps reflecting the poverty of island relative to mainland faunas (D'Antonio & Dudley, Reference D'Antonio, Dudley, Vitousek, Loope and Adersen1995). In addition, although Toft et al. (Reference Toft, Harris and Williams2001) found that the species richness of beetles and fungus gnats in New Zealand forests was positively correlated with the species richness of the vascular plant community, they did not detect a negative association of arthropod species richness with increasing abundance of the invasive herbaceous plant spiderwort, Tradescantia fluminensis (Commelinaceae). A study on the effect of an exotic grass (para grass, Urochloa mutica) in Australian floodplain habitats found no difference between benthic and epiphytic macroinvertebrate communities in stands of the exotic plant vs. stands of native plants (Douglas & O'Connor, Reference Douglas and O'Connor2003). They suggest that the structure of the vegetation is more important than plant identity to the diversity of these communities (see also Murdoch et al., Reference Murdoch, Evans and Peterson1972).
Although not all prior studies detected a negative association between invasive plant abundance and herbivore diversity, a common pattern found in these prior studies is that the arthropod communities formed distinct groupings among the different vegetation types, often with unique species inhabiting a specific habitat. Therefore, even though the species richness among the communities is indistinguishable, as the invasive vegetation type displaces the native vegetation, the overall expected result will be regional declines in species richness. An example of this is our observation that several moth species (Archanara alameda, A. oblonga, A. subflava and Bellura obliqua) known to specialize on cattails, rushes and sedges were not present at sites with high cover of the invasive plant species.
Three moth species were of particular interest due to their feeding behavior and area of origin. Two species of noctuid moths were prominent among the grass-feeding species in this study. Aletia oxygala, found at all of the study sites, has larvae that climb up vegetation to elude rising waters in seasonal wetlands with emergent vegetation such as reed canary grass. By contrast, Crymodes devastator is a soil-surface feeding species. It favors coarse grasses as larval foodplants, such as Dactylis glomerata or Zea mays, and is frequently an economic pest in various agricultural crops. We hypothesized that this species would be particularly abundant in wetlands dominated by reed canary grass. However, it was rarely found at reed canary grass sites, perhaps because the larvae feed at the soil surface and the species may be intolerant of seasonal inundation. A third grass-feeding species of interest is Apamea ophiogramma. It is an exotic species native to Europe where it is known to specialize on reed canary grass and other wetland grasses (South, Reference South1961). The species is thought to have been introduced to North America through the Port of Vancouver, British Columbia in the early 1990s. It has since spread south through western Washington in the 1990s and was first found in Oregon in this study. The species was frequent at one of the northern canary grass sites (FC) and is predicted to spread south through Oregon in the future.
Species-area curves (fig. 3) did not distinctly plateau with increasing sample effort. This result may be the effect of sampling a community that is temporally dynamic. The active flight periods of moths wax and wane throughout the growing season, so samples are not drawn from the same assemblage of species for each sample event. However, while the curves do not level off, they are generally the same shape (although three curves do show a different response). This suggests that, although actual diversity may be greater than our estimate, the diversity hierarchy is unlikely to change with more sampling, and the moth communites at the different sites can be reliably compared.
We excluded the abiotic variables that may have contributed to the associations that we observed. Ambient light, minimum night temperature, wind speed, rainfall and surrounding land use were not related to the abundance of the invasive plant species and, therefore, are not an underlying cause of the patterns we observed. The observed decline of moth species in these wetland sites was most likely caused by the negative effect of the invasive plants on the diversity of the wetland plant community.
In this study, we found that the negative effect of invasive plant abundance on plant diversity flows on to negatively impact the species richness of a higher trophic level. Arthropod herbivores and detritivores are known to influence primary production and nutrient cycling (Mattson & Addy, Reference Mattson and Addy1975; Seastedt & Crossley, Reference Seastedt and Crossley1984; Mikola & Setälä, Reference Mikola and Setälä1998; Jonsson & Malmquist, Reference Jonsson and Malmqvist2000). In addition, delivery of the ecosystem service of biological pest/weed control has been shown to increase with the number of herbivore species (Hoffman & Moran, Reference Hoffman and Moran1998); and changes in herbivore species composition, resulting from plant invasions, can have significant direct and indirect consequences to food webs (Willis & Memmott, Reference Willis and Memmott2005). Therefore, reductions in the species richness of these communities may affect ecosystem processes and services. Additional research is needed to determine (i) if a similar per-capita negative effect is realized for other invasive plant species and other herbivore communities, (ii) if carnivore trophic levels are similarly affected by invasive plants and (iii) how these reductions of plant and herbivore diversity will affect ecosystem function.
Acknowledgements
We thank the Oregon Department of Agriculture: Noxious Weed Board, USDI-FWS, Port of Portland, Portland Metro Parks and Greenspaces, Bonneville Power Administration and the Columbia Slough Watershed Council for funding. Thanks to our field assistants: K. Sivam, M. Schat, J. Fuller, C. Buhl and C. Meston. A. Brower, R. Garono, M. Kentula, V. Lesser, P. Rossignol and two anonymous reviewers provided valuable constructive reviews.
Appendix 1. Diversity formulas used in this study (Magurran, Reference Magurran1988)
(a) Species richness (S)=total number of species.
(b) Species equitability (J)=H/Hmax, where H is the Brillouin index of the sample, and Hmax is the potential maximum Brillouin index of the sample.
(c) Brillouin's index (H)=(ln N!−Σ ln n i!)/N, where N is the total number of individuals sampled, and n i is the number of individuals of the i th species.
(d) Simpson's index (D)=1−Σ[(n i(n i–1))/N(N–1))], where n i equals the number of individuals in the i th species, and N equals the total number of individuals.
Appendix 2
Moth species abundance and relationship with invasive plant cover. Axes refer to the final scores generated in the CCA. Axis 1 is primarily correlated with purple loosestrife cover and axis 2 is primarily correlated with reed canary grass cover.