


Introduction
Climate change is taking place all over the world (IPCC, Reference Houghton, Ding, Griggs, Noguer, van der Linden, Dai, Maskell and Johnson2001; Rosenzweig et al., Reference Rosenzweig, Karoly, Vicarelli, Neofotis, Wu, Casassa, Manzel, Root, Estrella, Seguin, Tryjanowski, Rawlins and Imeson2008). Global temperature as predicted by Intergovernmental Panel on Climate Change (IPCC) will increase between 1.1 and 5.4 °C by the year 2100 (IPCC, Reference Houghton, Ding, Griggs, Noguer, van der Linden, Dai, Maskell and Johnson2001). Many countries in the tropical and subtropical regions are expected to be more vulnerable to climate change because additional warming will affect, among other things, the agricultural sectors (Dinar et al., Reference Dinar, Benhin, Hassan and Mendelsohn2012). This is projected to worsen the already bad food situation in sub-Saharan Africa, where a large percentage of the population depends upon agriculture for their livelihood (Slingo et al., Reference Slingo, Challinor, Hoskins and Wheeler2005; Hellmuth et al., Reference Hellmuth, Moorhead, Thomson and Williams2007; IPCC, Reference Pachauri and Reisinger2007; McIntyre et al., Reference McIntyre, Herren, Wakhungu and Watson2009). Maize is one of the important staple crops produced by both small and large-scale farmers in sub-Saharan Africa (De Groote et al., Reference De Groote, Bett, Okuro, Odendo, Mose and Wekesa2004; Tefera et al., Reference Tefera, Mugo, Beyene, Karaya and Tende2011). However, the farming system faces a wide range of challenges and the resulting crop produce rarely meet the local demands. Some of the challenges include low soil fertility and losses associated with field and storage pests (De Groote, Reference De Groote2002; Kfir et al., Reference Kfir, Overholt, Khan and Polaszek2002). The lepidopteran stem borer pests are among the field pests that affect maize crop production and are currently estimated to cause field crop losses ranging between 15 and 50% (De Groote, Reference De Groote2002; Kfir et al., Reference Kfir, Overholt, Khan and Polaszek2002; Ong'amo et al., Reference Ong'amo, Le Ru, Dupas, Moyal, Calatayud and Silvain2006; Tefera et al., Reference Tefera, Mugo, Beyene, Karaya and Tende2011). A study conducted by Jones & Thornton (Reference Jones and Thornton2003) predict an overall reduction of about 10% in maize production by 2055 in sub-Saharan Africa, equivalent to losses of up to $2 billion per year, due to climate change. One of the main factors linked to the increase in global and local temperatures, is a change in stem borer infestation status. Climate, especially temperature, has a strong and direct effect on insect development, reproduction and survival (Bale et al., Reference Bale, Masters, Hodkinson, Awmack, Bezemer, Brown, Buttefield, Buse, Coulson, Farrar, Good, Harrington, Hartley, Jones, Lindroth, Press, Symrnioudis, Watt and Whittaker2002). A change in temperature has a number of implications on the above life history parameters, which may eventually change insect pest population (Ladányi & Horváth, Reference Ladányi and Horváth2010). With such change, insect pests may experience variations in population growth rates, increase in the number of generations, extension of the development season, changes in geographical distribution, crop–pest synchrony and interspecies interactions (Porter et al., Reference Porter, Parry and Carter1991; Roy et al., Reference Roy, Rothery, Moss, Pollard and Thomas2001; Bale et al., Reference Bale, Masters, Hodkinson, Awmack, Bezemer, Brown, Buttefield, Buse, Coulson, Farrar, Good, Harrington, Hartley, Jones, Lindroth, Press, Symrnioudis, Watt and Whittaker2002; Hance et al., Reference Hance, Van Baaren, Vernon and Boivin2007). Studies on the influence of temperature on insect development rates show no development at very low temperatures but development rates are found to gradually increase as temperature rises until an optimal development temperature is reached. Beyond the optimal temperature, development rates reduce and drop fast as they near the upper survival temperature limit (Stinner et al., Reference Stinner, Gutierrez and Butler1974; Logan et al., Reference Logan, Wollkind, Hoyt and Tanigoshi1976; Sharpe & DeMichele, Reference Sharpe and DeMichele1977; Wagner et al., Reference Wagner, Wu, Sharpe, Schoolfield and Coulson1984; Dixon et al., Reference Dixon, Honěk, Keil, Kotela, Šizling and Jarošík2009). Estimating the relationship between temperature and development rate, survival and reproduction is thus important in predicting the areas most suitable for species establishment under different climate change scenarios (Kiritani, Reference Kiritani1988; Cammell & Knight, Reference Cammell and Knight1992; Bale et al., Reference Bale, Masters, Hodkinson, Awmack, Bezemer, Brown, Buttefield, Buse, Coulson, Farrar, Good, Harrington, Hartley, Jones, Lindroth, Press, Symrnioudis, Watt and Whittaker2002; Honek et al., Reference Honek, Jarosik and Martinkova2003; Sporleder et al., Reference Sporleder, Kroschel, Quispe and Lagnaoui2004; Estay et al., Reference Estay, Lima and Labra2009; Régnière et al., Reference Régnière, St-Amant and Duval2012).
Predicting the potential distribution and abundance of agricultural pests could help governments adapt to climate change by developing and equipping farmers with adequate pest management tools to reduce crop losses (Kroschel et al., Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013). Predicting changes in pest distribution and abundance in many agricultural systems have been done using models (Pearson & Dawson, Reference Pearson and Dawson2003; Estay et al., Reference Estay, Lima and Labra2009). Models are analytical tools that have been used to determine the risks associated with the behavior of agricultural pest under climate changes (Baker, Reference Baker1991; Kroschel et al., Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013). There are mainly two distinct modeling approaches used for the evaluation, understanding, and prediction, of the dynamics of insect populations in agro ecosystems and assessments of phytosanitary risks (Trnka et al., Reference Trnka, Muška, Semerádová, Dubrovský, Kocmánková and Žalud2007; Kroschel et al., Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013). The first approach is the inductive method, which uses programs such as CLIMEX and BIOCLIM. This approach matches the climate where an organism is normally found within a region to where it is not normally found using long-term meteorological data (Beaumont et al., Reference Beaumont, Hughes and Poulsen2005; Sutherst & Maywald, Reference Sutherst and Maywald2005; Peacock & Worner, Reference Peacock and Worner2006; Legaspi & Legaspi, Reference Legaspi and Legaspi2007; Trnka et al., Reference Trnka, Muška, Semerádová, Dubrovský, Kocmánková and Žalud2007). The second is the deductive method, which relies on phenology models (process-based climate response models) (Trnka et al., Reference Trnka, Muška, Semerádová, Dubrovský, Kocmánková and Žalud2007) to describe the basic physiological principles of the insect species growth. These principles include development time, survival and reproduction (Curry et al., Reference Curry, Feldman and Smith1978; Sporleder et al., Reference Sporleder, Kroschel, Quispe and Lagnaoui2004; Nietschke et al., Reference Nietschke, Borchert, Magarey, Calvin and Jones2007). This approach is based on detailed laboratory experiments that produces life-table parameters and allows the simulation of populations according to real or interpolated data for a given region and time (Sporleder et al., Reference Sporleder, Simon, Juarez, Kroschel, Kroschel and Lacey2008). Linear degree-day models have long been accepted as a basis for building phenology and population dynamic models (Roltsch et al., Reference Roltsch, Mayse and Clausen1990), but the nonlinearities at high and low temperatures have made them poor predictors of insect development. The linear models work well for intermediate temperature but produce error when the temperature fluctuates to the extremes (Stinner et al., Reference Stinner, Butler, Bacheler and Tuttle1975; Hilbert & Logan, Reference Hilbert and Logan1983). Due to nonlinearity at the high and low temperatures, non-linear models have been developed (Logan et al., Reference Logan, Wollkind, Hoyt and Tanigoshi1976; Sharpe & DeMichele, Reference Sharpe and DeMichele1977; Briere et al., Reference Briere, Pracros, Le Roux and Pierre1999) which includes stochastic function for variability in development times within a population among individuals (Sharpe et al., Reference Sharpe, Schoolfield and Butler1981; Wagner et al., Reference Wagner, Wu, Sharpe, Schoolfield and Coulson1984). Development of such phenology models requires knowledge on lower and upper developmental thresholds as well as data on development for each life stage (Wagner et al., Reference Wagner, Olson and Willers1991; Nietschke et al., Reference Nietschke, Borchert, Magarey, Calvin and Jones2007).
In this study, we focused on Chilo partellus (Swinhoe), a species that was accidentally introduced in Africa from Asia (Tams, Reference Tams1932), and is one of the most important lepidopteran stem borer pests of maize and sorghum in Eastern and Southern Africa. Adults of C. partellus live for about 2–5 days and do not disperse far from emergence sites. The larval stage passes through six instars and the total larval period ranges from 20 to 28 days. The total development from egg to adult takes 25–50 days with five or more successive generation during a single maize growing season (Kfir, Reference Kfir1997; Hutchison et al., Reference Hutchison, Venette, Bergvinson and Van den Berg2008). Until the year 1990, its distribution was limited to low- and mid-altitude areas (<1500 masl), the distribution attributed to favorable abiotic factors, mainly temperature and humidity (Seshu Reddy, Reference Seshu Reddy1983; Zhou et al., Reference Zhou, Overholt and Mochiah2001). However, it has been reported recently that its distribution is expanding to higher altitudes (Kfir, 1997, Reference Kfir, Overholt, Khan and Polaszek2002; Zhou et al., Reference Zhou, Overholt and Mochiah2001; Ong'amo et al., Reference Ong'amo, Le Ru, Dupas, Moyal, Calatayud and Silvain2006). The projected change in climate and in particular the temperature increase is likely to directly affect C. partellus distribution range and pest status. However, little is known on the thermal requirements for the development of C. partellus, which would be useful in the development of risk maps. This study was therefore designed to develop temperature-dependent population growth model for C. partellus and to predict the future population growth potential in various maize agro-ecological zones in Sub-Saharan Africa under climate change.
Material and methods
General rearing condition
The effect of temperature on the development, survivorship/mortality and reproduction were studied on cohorts of single life stages of C. partellus in controlled incubators (Elbanton, type: LTKB 650 and Sanyo, MLR-350H 553) at six constant temperatures (15, 18, 20, 25, 30, 32 and 35 °C), relative humidity of 75±5% and a photoperiod of L12:L12 h. Portable digital thermo-hygrometers were placed inside the incubators to monitor the temperature and relative humidity. The C. partellus population used in this study was directly provided by Animal Rearing and Containment Unit, (ARCU), icipe, from a colony that was initially started with about 200 individuals of both sexes, collected from maize stems in Coast Province of Kenya. To limit the risk of inbreeding, about 200 new individuals (both males and females) collected from the same province are added to the colony each year. About five to six generations of C. partellus are produced in the insectary every year.
Effect of temperature on development and survival of eggs, larvae and pupae
The pairs of C. partellus were put in transparent plastic rearing jars (9 cm diameter×16 cm high) where they were provided with water and an oviposition substrate. Oviposition substrate consisted of a wax paper cut rectangularly (15 cm×6 cm) and rolled helicoidally from top to bottom to form a cylindrical surrogate stem (Khan & Saxena, Reference Khan and Saxena1997). Freshly laid eggs (24 h old) were collected from the jars, counted and distributed in four replicates of 250 each. The eggs were later placed in glass vials (7.5 cm high×2.5 cm diameter) sealed with cotton wool and maintained at the six different temperature regimes in the incubators. The eggs were observed daily during which the development duration of each egg and the total number of eggs that hatched at each temperature regime was recorded. For the larval development, 200 newly hatched larvae (24 h old) were transferred individually with a camelhair brush into glass vials (7.5 cm high×2.5 cm diameter) containing artificial diet. The glass vials were sealed with cotton wool to prevent the larvae from escaping.
The glass vials were then placed in a rearing carton box measuring (27 cm length×27 cm width×8 cm high) and maintained in incubators where the larvae were observed daily while recording the mortality and developmental time until pupation. The larvae were later placed individually in the glass vials to avoid cannibalism. The newly developed pupae were removed from the artificial diet, sexed and transferred to an empty glass vial (measuring 7.5 cm high×2.5 cm diameter), sealed with cotton wool, where they were observed for emergence. Those that did not emerge were observed for a longer time (more than a month) and recorded as dead. Number of adult moths that emerged and their sex were recorded daily.
Effect of temperature on fecundity, oviposition and longevity of adults
On the day of emergence, one female and one male were paired and placed individually in transparent plastic jars (9 cm diameter×16 cm high) where they were provided with water and an oviposition substrate (Khan & Saxena, Reference Khan and Saxena1997). The wax papers were replaced each morning for egg counts. Cotton wool soaked in water was placed in the rearing jars to maintain the relative humidity. The survival time of individual adults per temperature was recorded by sex.
Data for evaluation of the model
In natural conditions insects develop under fluctuating temperatures. To evaluate if our simulations with data collected under constant temperatures predicted well the development times and mortality in immature life stages, we carried out life table experiments to collect data under naturally fluctuating temperature at the icipe compound. The protocol used was the same as under constant temperatures in the laboratory. Freshly laid eggs (24 h old) were collected from the jars, counted and distributed in four replicates of 250 each. The eggs were placed in the open air where they were observed daily during which the development duration of each egg and the total number of eggs that hatched was recorded. For the larval development, 200 newly hatched larvae (24 h old) were transferred individually with a camelhair brush into glass vials (7.5 cm high×2.5 cm diameter) containing artificial diet and placed in the open air where they were observed daily, recording development time. The newly developed pupae were removed from the artificial diet, sexed and transferred to an empty glass vial (measuring 7.5 cm high×2.5 cm diameter), sealed with cotton wool, where they were observed for emergence. HOBO U10 Temp/RH Data Logger (Part # U10-003) was used for recording the daily minimum and maximum temperature. The experiment was carried between 1 November 2011 and 10 October 2012 with an average minimum and maximum temperatures of 15–35 and 27–37 °C, respectively.
The C. partellus phenology model
The C. partellus phenology model was implemented using Insect Life Cycle Modeling software (ILCYM version 3.0) (Kroschel et al., Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013). The software has the tools for building process-based population models for the insect species. The model builder uses the same shape distribution approach combined with a rate summation and cohort updating for simulation of the population model. ILCYM has several non-linear functions that describe the temperature-dependency of different processes in the insect species’ life history. These include the development time and its variation between individuals in a population, mortality in each immature life-stage, senescence and reproduction frequencies of the adult according to temperatures. The model builder facilitates choosing the best fitting functions for describing these temperature-driven processes that yield the overall phenology model.
Temperature-dependent processes models and statistical analysis
The relationship between temperature-dependence of different processes in the C. partellus life history and different temperature regimes were analyzed by various non-linear models, using the ILCYM software (Tonnang et al., Reference Tonnang, Juarez, Carhuapoma, Gonzales, Mendoza, Sporleder, Simon and Kroschel2013). The statistical analysis implemented in this software selected the best-fitting model to quantify the effect of temperature on the development time, mortality, senescence and reproduction according to inbuilt model selections criteria. These included the Akaike's information criterion (AIC) and the coefficient of determination R 2, which explains how the models capture the variability within the data. A female ratio of 0.5 was established for all the temperatures that were studied.
Development time and its variation
The cumulative probability distribution of C. partellus development times under different temperatures were estimated and normalized. Frequency distribution of insects development times are usually skewed toward the longer times and it is assumed that development times of insects at different temperatures are of the same shape. The normalized development times were arranged in frequency distribution and fitted to each life stage development time in a parallel line approach using log-development as the explanatory variable. The fitted generalized linear models to the normalized development time were the logit model for the eggs, larvae and pupa with the mathematical expression given as
 $$F(x) = \displaystyle{1 \over {1 + \exp ( - (a_i + b{\rm Inx}))}}$$
$$F(x) = \displaystyle{1 \over {1 + \exp ( - (a_i + b{\rm Inx}))}}$$
where F(x) is the probability to complete development at time x, Inx is the natural logarithm of the development days observed, a is the intercept corresponding to the temperature (i) and b is the common slope of the regression model.
Development rate
Temperature-dependent development of insects from one stage to another does not follow a linear relationship and hence linear models are not good for phenology models. In describing the relationship between temperature and the development rate, ILCYM provides several non-linear functions including Sharpe and DeMichele (Sharpe & DeMichele, Reference Sharpe and DeMichele1977), Logan's model (Logan et al., Reference Logan, Wollkind, Hoyt and Tanigoshi1976) and other models that have been used successfully for many insect species. The development time d was used to calculate development rates. Development rate was fitted to various non-linear models and Logan et al. (Reference Logan, Wollkind, Hoyt and Tanigoshi1976) provided an excellent description of the temperature-dependent development rate of C. partellus across all temperatures tested for all developmental stages (table 1). The model is expressed as
 $$\; r(T) = Y\left\{ {\exp ({\rm \rho} T) - \exp \left( {{\rm \rho} T_{max} - \displaystyle{{T_{max} - \; T} \over V}} \right)} \right\}$$
$$\; r(T) = Y\left\{ {\exp ({\rm \rho} T) - \exp \left( {{\rm \rho} T_{max} - \displaystyle{{T_{max} - \; T} \over V}} \right)} \right\}$$
where r(T) is the rate of development at temperature T; T max is the upper development threshold Y, p and v are constants. The choice of the best-fitting function in ILCYM was done using the R 2 statistics or the AIC (Akaike, Reference Akaike, Petrov and Csaki1973). A simple linear regression was used to estimate the lower development threshold
 $$r(T) = a + b.T$$
$$r(T) = a + b.T$$
where a is the intercept and b is the slope.
Table 1. Estimated parameters (mean±SE) and coefficients of determination of the Logan 1 model fitted to median development rate and temperature for C. partellus immature life stages.

Mortality
The mortality rate in the immature life stage was calculated from the relative frequency of cohort survivors. The polynomial model was fitted by regression to describe the relationship between the mortality rate and the temperature for each life stage. The polynomial regression is expressed as
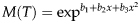 $$M(T) = \exp ^{b_1 + b_2 x + b_3 x^2} $$
$$M(T) = \exp ^{b_1 + b_2 x + b_3 x^2} $$
where M(T) is the rate of mortality at temperature T; and b i s are parameters to be estimated.
Adult life span and reproduction
The mean survival time of the adult was determined for both sexes and the inverse of the survival time was plotted against temperature. A modified four parameter Stinner model (Stinner et al., Reference Stinner, Gutierrez and Butler1974) was fitted to determine the relationship between the senescence rate of both the male and female adults and temperature. The mathematical expression of the model is given as
 $$S(T) = \displaystyle{{c_1} \over {1 + e^{(k_1 + k_2 T)}}} \; + \displaystyle{{c_2} \over {1 + e^{(k_1 + k_2 (T_0 - T))}}} $$
$$S(T) = \displaystyle{{c_1} \over {1 + e^{(k_1 + k_2 T)}}} \; + \displaystyle{{c_2} \over {1 + e^{(k_1 + k_2 (T_0 - T))}}} $$
where S(T) is the senescence rate at temperature T (°C), c 1 and c 2 are maximum and minimum temperatures, respectively, T o is the optimum temperature and k 1 and k 2 are constants. A polynomial regression was applied to determine the effect of temperature on the total number of eggs laid per female during her whole life span. The expression of the model is given as
 $$F(T) = b_{\rm 1} + b_{\rm 2}. x + b_{\rm 3}. x^{\rm 2} $$
$$F(T) = b_{\rm 1} + b_{\rm 2}. x + b_{\rm 3}. x^{\rm 2} $$
Life table parameters
Using the stochastic simulation tool in ILCYM the life parameters that is, gross reproductive rate (GRR), intrinsic rate of natural increase (r m), finite rate of increase (λ), net reproductive rate (R o ), mean generation time (T), and doubling time (Dt) were estimated based on the developed phenology model. The simulation is based on the rate summation and random determination for each individual's survival, and development to the next stage. The simulation started with 100 individuals from the egg stage for a given constant temperature regime and were performed over a temperature ranging from 18 to 35 °C with four repetitions at each temperature.
Evaluation of the models
The validation tool in ILCYM was used to evaluate whether the developed phenology models parameterized with the life table data collected under constant temperatures were comparable with data collected under fluctuating temperature conditions. Daily data on minimum and maximum temperature were obtained from experiments carried out under fluctuating temperatures at the icipe compound. The daily maximum and maximum were used in the stochastic simulation and the results compared with the life table from the constant temperatures. The minimum and maximum temperatures recorded during the experiment period ranged between 9.6–17.9 °C and 18.4–37.9 °C, respectively.
Potential population distribution and risk mapping
The Insect Life Cycle Modeling software (ILCYM version 3.0) (Kroschel et al., Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013) was used to generate C. partellus risk maps using the validated phenology model. Linked with GIS, the model allows the simulation of pest risk indices. The indices used for mapping included establishment index (ERI), generation index (GI) and activity index (AI) as calculated by Kroschel et al. (Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013), which are defined as follows
-
The ERI identifies the area in which the pest may survive and become established.
-
The GI estimates the mean number of generations that may be produced within a given year.
-
The AI is explicitly related to the finite rate of population increase, which takes the whole life history of the pest into consideration.
The simulation was carried out using the climate data obtained from WorldClim (http://www.worldclim.org/) and CCAFS (http://www.ccafs-climate.org) databases as described in Kroschel et al. (Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013). For Kenya, a spatial resolution of 2.5 arc-min was used whereas 10.0 arc-min was applied for Africa. In predicting the C. partellus response to climate change for the year 2050, we used downscaled data of the SRES-A1B (IPCC, Reference Pachauri and Reisinger2007; Kroschel et al., Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013) to project temperature changes. ‘The A1 scenario family develops into three groups that describe alternative directions of technological change in the energy system. The three A1 groups are distinguished by their technological emphasis: fossil intensive (A1FI), non-fossil energy sources (A1T), or a balance across all sources (A1B) (where balance is defined as not relying too heavily on one particular energy source, on the assumption that similar improvement rates apply to all energy supply and end-use technologies)’. www.ipcc.ch/ipccreports/tar/wg1/029.htm
Results
Development time
Chilo partellus individuals were able to develop across a wide range of temperatures and successful development from the egg to an adult was observed from 18 to 35 °C, respectively. The mean development times for each development stage and for each temperature are shown in table 2. Temperature significantly influenced the duration of each developmental stage. At temperatures where the development was complete (from egg to adulthood), the mean development times decreased with increasing temperature. At 15 and 38 °C, the eggs failed to hatch.
Table 2. Mean development time (in days) of C. partellus life stages at eight different constant temperatures.

Within a column means followed by the same letters are not significantly different (P=0.05), Student–Newman–Keuls test.
1 Eggs did not hatch at these temperatures.
Mortality of immature stages
The effect of temperature on the mortality of C. partellus immature life stages was described by polynomial functions (table 3 and fig. 1). Chilo partellus successfully developed from egg to adult between the temperature ranges of 18 to 35 °C but failed at 15 and 38 °C, as no egg hatched at these temperatures. The percentage mortality of eggs decreased at temperatures between 18 and 25 °C and increased thereafter. The minimum mortality of 24.6% was observed at 30 °C while the maximum mortality of 100% was reported at 15 and 38 °C (fig. 1). Mortality within the larval and pupa stages showed a similar temperature response. The highest mortality for both the larval and pupal stages was observed at the extreme temperatures. The minimum mortality for the larvae was 10.7% at 30 °C and for the pupa 10% at 32 °C.

Fig. 1. Temperature-dependent mortality rates of C. partellus immature life (eggs, larva and pupa).
Table 3. Estimated parameters (mean±SE) of the polynomial model fitted to mortality rate for eggs, larva and pupa stages of C. partellus.

Development rate
The development rates increased with temperature; however, the threshold temperature differed among the development stages. The estimated optimal temperatures for development were 33, 32 and 33 °C for eggs, larva, and pupa, respectively; at temperatures above the optima, the rate of development decreased sharply (fig. 2). The line regression estimated the lower threshold temperature as 10.5, 9.56 and 10.09 for eggs, larva and pupa, respectively.

Fig. 2. Temperature-dependent model of development rate (1/development time) for C. partellus eggs, larvae and pupa. The Logan model r(T) fit to the data.
Adult longevity and fecundity of C. partellus
The longevity of C. partellus adults was significantly affected by temperature (P<0.05). The mean longevity significantly decreased with increasing temperature for both males and females (table 2). The longevity for both males and females was 11 days at 18 °C and 6 days at 35 °C. The four-parameter model of Sharpe & DeMichele (Reference Sharpe and DeMichele1977) gave good fit to the observed mean senescence rate for both sexes (fig. 3). The effect of temperature on fecundity was described by a polynomial function (F=7.37; df=3,2; P=0.12). The average number of eggs laid per female significantly increased with increasing temperature between 15 and 25 and then decreased. At the lower and upper temperatures tested i.e., 18 and 35 °C, the fecundity per female was predicted to be 205.7 and 46, respectively. The maximum fecundity of 375.3 was observed at 25 °C (fig. 4A). The cumulative proportion of eggs produced per female and normalized female age was described by the gamma function. The oviposition was completed when the female reached their median life time (fig. 4B).

Fig. 3. Temperature-dependent senescence rate for adult female (A) and males (B) for C. partellus fitted to Sharpe and DeMichele model.

Fig. 4. (A) Temperature-dependent total egg production curve, (fitted curve: polynomial). (B) Cumulative proportion of egg production in relation to female age expressed as normalized time (fitted curve: gamma cumulative distribution function).
Life table parameters of C. partellus
The models established were compiled in an overall phenology model for stochastic simulation of C. partellus resulting in life table parameters (table 4). The intrinsic rate of natural increase (r m), the net reproductive rate (R o ), the finite rate of increase (λ), the mean generation time (Ί) and doubling time (Dt) were estimated for each temperature. The life-table parameters predicted show that C. partellus populations develop within the range of 18–35 °C (table 4). The optimum temperature for population development is at 30 °C with the highest intrinsic rate of increase. The net reproductive rate R o differed significantly among temperatures with the highest at 25 °C (56.72). The population reared at the lower and higher temperatures of 18 and 35 °C had smaller r m of 0.02 and 0.01 and R o of 7.14 and 0.511, respectively.
Table 4. Life table parameters of C. partellus at different constant temperatures. Intrinsic rate of increase (r m), net reproduction rate (R o ), finite rate of population increase (λ), gross reproduction rate (Ḡ), mean generation time (Ί), doubling time (Dt).

Within a column means followed by the same letters are not significantly different (P=0.05), Student–Newman–Keuls test.
Evaluation of the models
Simulations with data collected under constant temperatures predicted good development times and mortality in immature life stages when compared with the data collected under fluctuating temperatures (table 5).
Table 5. Observed and simulated development time and mortalities in C. partellus eggs larvae and pupae.

Average±SE: standard errors are calculated from the observed and simulated life table data.
Spatial mapping: changes in C. partellus distribution and abundance
In Kenya, where a lot of research has been done on the distribution of C. partellus, the result shows that under the current climate scenario (year 2000), C. partellus is established along the coastal region, and around Lake Victoria (fig. 5A) with an ERI between 0.7 and 1. Under the climate scenarios of the year 2050 (fig. 5B), there is indication of the boundary expansion of C. partellus to higher elevation areas, which is estimated to be prone to permanent establishment. The change in C. partellus ERI between 2000 and 2050 indicates that C. partellus may potentially disappear from the Coast and North Eastern part of Kenya (fig. 5C).

Fig. 5. Chilo partellus ERI in 2000 (A) and 2050 (B) and change of C. partellus ERI between 2000 and 2050 (C) according to ILCYM model predictions in Kenya. Indices>0.6 is associated with potential permanent establishment. The ERI identifies the area in which the insect may survive and become established permanently.
Under the present climate conditions of year 2000, the establishment risk index reflects well the current distribution of C. partellus in Eastern and Southern sub-Saharan countries, which includes Kenya, Tanzania, Uganda, Zambia, Botswana, Eritrea, Ethiopia, Mozambique and Somalia (fig. 6A, B). There is also an indication that C. partellus has a great value of ERI in several countries in West Africa (fig. 6A) although the species has never been recorded in there. Under the future scenario (2050), most areas in West Africa will likely have very little suitability for C. partellus establishment (0.5<ERI≤0.6), while most areas in Austral Africa, may potentially be much more suitable; in Eastern Africa the species will likely expand to higher altitude areas (0.7<ERI≤1) (fig. 6B, C) and disappear from dry lowland areas. Changes in GI will likely affect the abundance of C. partellus and the infestation in maize or other cereal crops available. According to the prediction, several countries in Eastern and Southern Africa will experience higher number of generations per year (8<GI≤10) in the year 2050 compared with the year 2000 (6<GI≤8) (fig. 7A–C). The risk maps developed for Africa with the climate data for the year 2000 and year 2050 show that areas with high-GI also have high AI (figs 7A–C and 8A–C). There will be a change in the AI from (9<AI≤12.0) under the current temperature (2000) to (13<AI≤15) in the year 2050 (fig. 8A–C). The change in GI indicates that in most of the maize production regions an increase of 1–2 generations can be expected.

Fig. 6. Chilo partellus ERI in 2000 (A) and 2050 (B) and change of C. partellus ERI between 2000 and 2050 (C) according to ILCYM model predictions in sub-Saharan Africa. Indices>0.6 is associated with potential permanent establishment. The ERI (survival) identifies the area in which the insect may survive and become established permanently.

Fig. 7. Chilo partellus GI in 2000 (A) and 2050 (B) and change of C. partellus GI between 2000 and 2050 (C) according to ILCYM model predictions in sub-Saharan Africa. The GI estimates the mean number of generations that may be produced within a given year.

Fig. 8. Chilo partellus AI in 2000 (A) and 2050 (B) and change of C. partellus AI between 2000 and 2050 (C) according to ILCYM model predictions in sub-Saharan Africa. The AI takes the whole life history into consideration; an index value of 3 would illustrate a potential population increases by a factor of 1000 within one year (all other population limiting factors including food availability are neglected).
Discussion and conclusion
Climate and especially temperature is known to have great effect on the insect's population growth and geographical distribution (Logan et al., Reference Logan, Regniere and Powell2003; Menendez, Reference Menendez2007; Deutsch et al., Reference Deutsch, Tewksbury, Huey, Sheldon, Ghalambor, Haak and Martin2008). There have been few studies on the effect of global warming on insect pest in tropical areas with the recent exception of hematophagous insects such as Anopheles spp. (Patz & Olson, Reference Patz and Olson2006) and Glossina pallidipes (Terblanche et al., Reference Terblanche, Clusella-Trullas, Deer and Chown2008), the coffee berry borer (Jaramillo et al., Reference Jaramillo, Chabi-Olaye, Kamonjo, Jaramillo, Vega, Poehling and Borgemeister2009) and the potato tuber moth (Kroschel et al., Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013). Tropical ectotherms are at most risk from climate change as they inhabit hot environments (Zeh et al., Reference Zeh, Bonilla, Su, Padua, Anderson, Kaur, Yangn and Zeh2012). Small increase in temperatures in the tropical will have great effect on the ectotherms since many species are already near their optimal temperature. Predicting the impact of climate change on the species populations based only on the magnitude and patterns of climate warming may not be sufficient (Tewksbury et al., Reference Tewksbury, Huey and Deutsch2008). The thermal sensitivity of the organism under study has to be considered (Kearney et al., Reference Kearney, Porter, Williams, Ritchie and Hoffmann2009; Kingsolver et al., Reference Kingsolver, Woods, Buckley, Potter, MacLean and Higgins2011). Models that include organism physiology suggest that impacts of climate warming may be more severe in the tropics (Tewksbury et al., Reference Tewksbury, Huey and Deutsch2008). In this study we report the first detailed prediction of the effect of temperature change on the distribution of C. partellus, one of the main stem borer pest in sub-Saharan Africa based on a process-based phenology model.
Physiological effects of temperature
Previous studies on the effect of temperature on the development and survival of C. partellus did not consider the temperature-dependent mortality rate at all immature stages (Singh, Reference Singh1991; Mbapila et al., Reference Mbapila, Overholt and Kayumbo2002; Tamiru et al., Reference Tamiru, Getu, Jembere and Bruce2012). In this study we predicted the life table parameters (i.e. development rate, mortality and fecundity) under a given temperature regime by taking the total life history into account and estimated the temperature-dependent population growth potential. Our results showed that the temperature affects the development and survival of the different stages of C. partellus. The development times of the immature life stages of C. partellus presented here do not differ substantially from the results presented by Mbapila et al., Reference Mbapila, Overholt and Kayumbo2002. Development times of the different stages decreased with increasing temperatures ranging from 18 to 35 °C. At the extreme temperatures, 15 and 38 °C, no egg could hatch and no larvae completed development. Previous studies reported the mean development time from egg to adult reared on artificial diet of C. partellus was 48.3 days at 25 °C (Mbapila et al., Reference Mbapila, Overholt and Kayumbo2002), which is almost similar to 47 days at 25 °C recorded in our study. The temperature-dependent mortality rates observed were high at the minimum (18 °C) and maximum (35 °C) temperatures. The study agrees with prediction of Stevens climatic variability hypothesis (Stevens, Reference Stevens1989; Jaramillo et al., Reference Jaramillo, Chabi-Olaye, Kamonjo, Jaramillo, Vega, Poehling and Borgemeister2009), which states that the thermal tolerance of an insect is directly proportional to the climate variability the organism is exposed to. The temperature limits for C. partellus survival are 14.9 °C and 38.2 °C. In this regard, C. partellus is likely to establish and become a serious pest between 20 and 32 °C with the most suitable temperature being at 30 °C where an intrinsic rate of increase is 0.123. These results are similar to findings of Singh (Reference Singh1991), who reported the optimum intrinsic rate of increase and net reproduction rate to occur at 30 °C (0.108 and 51.091).
Prediction of C. partellus distribution
Originating from the Indian Subcontinent, C. partellus is essentially a hot lowland area pest; since its first appearance in Malawi (Tams, Reference Tams1932), it expanded its distribution in the warm, low-altitude regions of Eastern and Southern Africa (Kfir et al., Reference Kfir, Overholt, Khan and Polaszek2002). The geographical distribution of C. partellus was thought to be dependent on the elevation (Harris & Nwanze, Reference Harris and Nwanze1992), but Sithole (Reference Sithole1989) argued that temperature, rainfall and humidity were the main factors that affect its distribution, with temperature being the most important. Temperature and elevation are highly correlated and therefore the two hypotheses are true. Most recent studies show that C. partellus has remained an important pest in low altitude areas but at the same time it is expanding its distribution into higher elevation areas where it used not to occur (Kfir, Reference Kfir1997; Guofa et al., Reference Guofa, Overholt and Mochiah2002; Ong'amo et al., Reference Ong'amo, Le Ru, Dupas, Moyal, Calatayud and Silvain2006).
Our prediction of the current distribution and abundance of C. partellus were found to agree with the recent survey results in Kenya (Guofa et al., Reference Guofa, Overholt and Mochiah2002; Ong'amo et al., Reference Ong'amo, Le Ru, Dupas, Moyal, Calatayud and Silvain2006) where it was found to be the dominant species at the dry mid altitude, dry transitional and lowland tropical zones. Chilo partellus has been reported in most countries in Eastern and Southern Africa (Kfir et al., Reference Kfir, Overholt, Khan and Polaszek2002). Using CLIMEX to map the potential geographical distribution of C. partellus in Africa, Hutchison et al. (Reference Hutchison, Venette, Bergvinson and Van den Berg2008) predicted very similar results. These results indicate the potential establishment of C. partellus in several West African countries where it is not currently known to occur (Kfir et al., Reference Kfir, Overholt, Khan and Polaszek2002). Similar results were reported by Overholt et al. (Reference Overholt, Songa, Ofomata, Jeske, Lyons and Miller2000), and Hutchison et al. (Reference Hutchison, Venette, Bergvinson and Van den Berg2008), who used the GIS model to predict distribution of C. partellus, based on the climate of the known C. partellus locations.
Our predictions on the future C. partellus expansion to higher elevation in Eastern Africa (Burundi, Ethiopia, Kenya, Uganda, Tanzania, Rwanda), and to mid elevation in Southern Africa (Mozambique, Zambia, Zimbabwe, South Africa) confirm previous observations by Kfir (Reference Kfir1997), Guofa et al. (Reference Guofa, Overholt and Mochiah2002) and Ong'amo et al. (Reference Ong'amo, Le Ru, Dupas, Moyal, Calatayud and Silvain2006). However, our predictions clearly indicate that C. partellus may disappear from moist lowland dry areas in Kenya and Somalia. Our predictions also clearly indicate an increase in the number of C. partellus generations and AI in most countries in East and Southern Africa suggesting that economic losses are likely to increase in areas where C. partellus is already established and extend in areas where it has not yet been recorded. According to our predictions, the three pest risk indices will decrease in future in West Africa suggesting the potential risk of invasion of C. partellus will decrease in this part of the continent. Contrary to West Africa, the three pest risk indices will increase significantly in Central Africa suggesting that C. partellus could potentially become an important maize pest in this part of the African continent, in case of any accidental introduction.
The eco-physiology of both insects and plants will predict the future distribution of insect pests when both insects pest and host plant are in close synchrony (Hodkinson, Reference Hodkinson1999; Crozier & Dwyer, Reference Crozier and Dwyer2006). In the case of C. partellus, which is a highly specialized herbivore, the effect of climate change on the insect and host plant cannot be separated. Previous research suggests that maize growing regions of sub-Saharan Africa will experience increased growing seasons temperature and frequency of droughts (IPCC, Reference Pachauri and Reisinger2007). The results of temperature simulation for 2050 across maize mega-environment within sub-Saharan Africa show a general trend of warming with the maximum temperatures predicted to increase by 2.6 °C and the minimum temperatures slightly lower with an average of 2.1 °C (Cairns et al., Reference Cairns, Sonder, Zaidi, Verhulst, Mahuku, Babu, Nair, Das, Govaerts, Vinayan, Rashid, Noor, Devi, San Vicente, Prasanna and Sparks2012). Our study reveals that expected change in temperatures may lead to change of ranges of C. partellus present distribution. Studies on the potential impacts of climate change show negative impact on the maize production leading to serious problem in food security (Muchena & Iglesias, Reference Muchena, Iglesias, Rosenzweig, Allen, Harper and Jones1995; Jones & Thornton, Reference Jones and Thornton2003; Abraham & Savage, Reference Abraham and Savage2006; Lobell et al., Reference Lobell, Bänziger, Magorokosho and Vivek2011). Increased temperatures can speed up the cycle of insects leading to a faster increase in pest population (Cairns et al., Reference Cairns, Sonder, Zaidi, Verhulst, Mahuku, Babu, Nair, Das, Govaerts, Vinayan, Rashid, Noor, Devi, San Vicente, Prasanna and Sparks2012). It has been estimated that a 2 °C increase in temperature has the potential to increase the number of generation per year (Porter et al., Reference Porter, Parry and Carter1991; Bale et al., Reference Bale, Masters, Hodkinson, Awmack, Bezemer, Brown, Buttefield, Buse, Coulson, Farrar, Good, Harrington, Hartley, Jones, Lindroth, Press, Symrnioudis, Watt and Whittaker2002). In regions where there is an abundance of host plants and climate is warmer enough for development, C. partellus normally develops continuously all year round. In regions with long dry periods in winter or in summer, the borer enters into a resting period (Kfir, Reference Kfir1997). Our prediction on the distribution of C. partellus with global climate change shows that the insect pest is expanding its distribution to the higher altitude. Not all maize varieties will respond similarly to the climate change, and altitudinal change of varieties as a coping strategy in a global warming is a key potential means of adaptation.
In this paper we considered one single climate variable (temperature); however, other environmental factors such as rainfall and relative humidity, may affect the establishment of C. partellus. Abraham et al. (Reference Abraham, Thomas, Karunakaran and Gopalakrishnan1972), through correlation studies, found that there was a joint influence of rainfall, relative humidity and mean minimum temperature on the stem borer infestation. There is need to consider the effect of rainfall on the establishment and abundance of C. partellus when predicting the future pest risk. Climate change could upset the balance between insect crops pests and the natural enemies that control their number and therefore make pest outbreaks more frequent and severe (Hance et al., Reference Hance, Van Baaren, Vernon and Boivin2007; Thomson et al., Reference Thomson, Macfadyen and Hoffmann2010). The interaction between insect pests and their natural enemies needs to be better understood in order to devise methods for management. The prediction of the present ERI for all sub-Saharan countries was validated using known distribution records from Kenya where several surveys have been carried out. As a consequence there are some limitations in the predictions generated. This limitation can be overcome by spatial point-by-point of given location to estimate the potential generation within a year based on the minimum and maximum temperatures of that particular location (Kroschel et al., Reference Kroschel, Sporleder, Tonnang, Juarez, Carhuapoma and Simon2013).
In conclusion, our study shows that expected temperature increase as predicted by Intergovernmental Panel on Climate Change (IPCC, Reference Houghton, Ding, Griggs, Noguer, van der Linden, Dai, Maskell and Johnson2001) may have serious potential consequences on C. partellus distribution in most sub-Saharan African countries. To alleviate the potentially negative effect caused by C. partellus on maize production, farmers, extension workers and other stakeholders will have to adapt relevant cultural practices (Challinor et al., Reference Challinor, Wheeler, Garforth, Craufurd and Kassam2007). Adaptation strategies should look at the use of resistant varieties and length of growing period; intercropping maize with non-host plants should also be considered as they significantly reduces the oviposition, infestation and damage due to borer compared with maize monocrop (Chabi-Olaye et al., Reference Chabi-Olaye, Nolte, Schulthess and Borgemeister2005). At least, removal of maize residues, important in sustaining carry-over populations of Cp (Kfir, Reference Kfir1990; Ebenebe et al., Reference Ebenebe, Van den Berg and Van der Linde2001) should be evaluated.
Acknowledgements
The first author of this study is a PhD student working under ‘Predicting climate-change-induced vulnerability of African agricultural systems to major insect pests through advanced insect phenology modeling, and decision aid development for adaptation planning’ project. The authors gratefully acknowledge the financial support provided by the Federal Ministry of Co-operation and Development (BMZ), Germany, for the project. The authors thank the two anonymous referees for their valuable comments and suggestions.













