Introduction
Flat grain beetles of the genus Cryptolestes Ganglbauer 1899 (Coleoptera: Laemophloeidae) are considered serious cosmopolitan secondary stored-product pests which feed on many kinds of agricultural products (Halstead, Reference Halstead1993). They are usually associated with cereal products (e.g., wheat, barley, oat and maize), but can also be found on beans, oilseeds, cassava, nuts, dried fruit, dried vegetable, or even liquorice apricots (Howe & Lefkowitch, Reference Howe and Lefkowitch1957; Thomas & Zimmerman, Reference Thomas and Zimmerman1989; Halstead, Reference Halstead1993). Economic losses caused by Cryptolestes species are exacerbated by the fact that both adults and larvae feed on agricultural products. Additionally, they play a role in food safety by transferring pathogenic microorganisms and contaminating stored products with filth fragments (Trematerra et al., Reference Trematerra, Stejskal and Hubert2011). Nine Cryptolestes species are considered important stored-product pests, of which Cryptolestes ferrugineus (Stephens) and Cryptolestes pusillus (Schönherr) are globally distributed, whereas the other Cryptolestes species are more restricted in their ranges and thus represent a potential invasive threat (Howe & Lefkowitch, Reference Howe and Lefkowitch1957; Halstead, Reference Halstead1993). For example, it was only recently confirmed that two exotic Cryptolestes, namely, Cryptolestes cornutus (Thomas & Zimmerman) and Cryptolestes klapperichi (Lefkovitch), were newly established in Florida (Thomas, Reference Thomas2005). However, Cryptolestes turcicus (Grouvelle), which originated from the temperate climatic zone of the northern hemisphere, did not establish in Australia until recently. This is despite the fact that climatic conditions in Australia support the establishment of this insect species. Cryptolestes species differ in biological characteristics, such as their reproduction cycles (Lefkovitch, Reference Lefkovitch1957; Rees, Reference Rees2004), as well as in their susceptibility to management tools used to control their populations (Lefkovitch, Reference Lefkovitch1965; Throne et al., Reference Throne, Doehlert and McMullen2002; Nayak et al., Reference Nayak, Holloway, Emery, Pavic, Bartlet and Collins2013). As a result, correct identification of different species and strains is important for minimizing their introduction and establishment in new areas, and for effectively managing them.
Individual species within Cryptolestes are very similar in their external traits and are difficult to identify. Identification keys for Cryptolestes species can be helpful, but the diagnostic morphological characteristics are mostly based on microscope examination of genitalia (Banks, Reference Banks1979; Halstead, Reference Halstead1993), which are not easy to recognize reliably and preparation of insect material for identification is time consuming and not straightforward. Under certain circumstances, only Cryptolestes fragments (e.g., legs or wings) or immature stages (eggs, larvae and pupae) may be available and these do not have sufficient characteristics for reliable morphological species identification (Kučerová & Stejskal, Reference Kučerová and Stejskal2002; Trematerra et al., Reference Trematerra, Stejskal and Hubert2011). Given the aforementioned challenges to identification of Cryptolestes species, it is logical to develop practical, rapid and accurate identification methods. The developed methods should allow for identification of species using both whole insects and insect parts of any life stages.
DNA-based approaches represent an alternative way for identification of organisms, among which it was demonstrated that DNA barcoding resolves most species (Waugh, Reference Waugh2007). A short fragment of mitochondrial cytochrome c oxidase subunit I (COI) gene is widely used as the barcode sequence for animals and is frequently employed in applied entomology too (Armstrong & Ball, Reference Armstrong and Ball2005; Nelson et al., Reference Nelson, Wallman and Dowton2007; Wells & Williams, Reference Wells and Williams2007; deWaard et al., Reference deWaard, Mitchell, Keena, Gopurenko, Boykin, Armstrong, Pogue, Lima, Floyd, Hanner and Humble2010; Jinbo et al., Reference Jinbo, Kato and Ito2011; Morag et al., Reference Morag, Saroya, Braverman, Klement and Gottlieb2012; Wang et al., Reference Wang, Li, Guo, Xing, Dong, Wang, Zhang, Liu, Zheng, Zhang, Zhu, Wu and Zhao2012; Jiang et al., Reference Jiang, Li, Deng, Wu, Liu and Buahom2013). Although species identification based on COI barcoding and species-specific primers has already been successfully applied for identification of different storage pests (Obrepalska-Steplowska et al., Reference Obrepalska-Steplowska, Nowaczyk, Holysz, Gawlak and Nawrot2008; Nowaczyk et al., Reference Nowaczyk, Obrepalska-Steplowska, Gawlak, Throne, Olejarski and Nawrot2009; Yang et al., Reference Yang, Kučerová, Li, Kalinovic, Stejskal, Opit and Cao2012, Reference Yang, Zhao, Kučerová, Stejskal, Opit, Qin, Cao, Li and Li2013; Wang et al., Reference Wang, Li, Zhang, Varadínová, Jiang, Kučerová, Stejskal, Opit, Cao and Li2014), more research needs to be conducted to enable this approach to be easily adopted for molecular identification of Cryptolestes species. Wang et al. (Reference Wang, Li, Zhang, Varadínová, Jiang, Kučerová, Stejskal, Opit, Cao and Li2014) evaluated the accuracy of DNA barcoding based on COI gene for discrimination of the five most common storage Cryptolestes species. In that study, they clearly demonstrated that COI barcoding is a suitable alternative to traditional morphological identification of Cryptolestes species.
In the present study, our goal was to refine the molecular approach of Wang et al. (Reference Wang, Li, Zhang, Varadínová, Jiang, Kučerová, Stejskal, Opit, Cao and Li2014) to make it more practical for identification purposes. This was achieved by investigating the feasibility of species-specific primers in conventional polymerase chain reaction (PCR) system for reliable and rapid identification of five common stored-product pest species of the genus Cryptolestes, both larvae and adults.
Materials and methods
Cryptolestes specimens
The larval specimens of five Cryptolestes species (table 1), namely, Cryptolestes capensis (Waltl), C. ferrugineus (Stephens), C. pusilloides (Steel & Howe), C. pusillus (Schönherr) and C. turcicus (Grouvelle), came from laboratory strains or collections from grain stores that were preserved in alcohol. Altogether eight larval strains were used in the study (table 1). There are one strain each of C. capensis (Czech Republic), C. pusilloides (China) and C. turcicus (Czech Republic); two strains of C. ferrugineus (China, Czech Republic) and three strains of C. pusillus (two from China and one from Czech Republic). Prior to DNA extraction, larval and adult specimens were regarded as the same species if they came from the same colony or collection site. The adult specimens were identified based on taxonomic keys (Lefkovitch, Reference Lefkovitch1962; Halstead, Reference Halstead1993).
Table 1. List of samples used in evaluation of Cryptolestes species specific primers.

DNA extraction and COI sequencing
Genomic DNA of larvae was extracted from whole larval specimens using commercial TIANamp Genomic DNA kit (TIANGEN, China) according to the manufacturer's protocol with only slight modification. During the last step of the extraction, DNA was eluted in 2 × 50 μl TE buffer in two subsequent centrifugation steps. Universal forward LCO1490 and reverse HCO2198 primers were used for COI amplification (Folmer et al., Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994). PCR was carried out based on methods by Wang et al. (Reference Wang, Li, Zhang, Varadínová, Jiang, Kučerová, Stejskal, Opit, Cao and Li2014) in Veriti TM 96-well Thermal Cycler (ABI, USA). PCR products were confirmed by 1% agarose gel electrophoresis, stained with ethidium bromide (EB) and visualized under UV light (Gel Logic 212 PRO, Carestream Health, Inc.). Products were sequenced in both directions by Sangon Biotech (Shanghai) Co., Ltd.
Sequence analysis
Altogether we obtained sequences from 49 individuals, at least four individuals from each collection site. Original chromatograms of sequences were manually checked to avoid reading errors and the low quality ones were excluded from further analysis. After assembling, products were cut to a final 658 base pair fragment of COI gene. Sequences from each locality were aligned and all the identified haplotypes were submitted to GeneBank. These analyses were performed in DNAMAN (version 6.0.3.99) and DnaSP (version 5.10). Correct species identification of larval strains was verified by aligning and constructing joined phylogenetic tree of larval sequences obtained in this study with adult sequences from GeneBank (Wang et al., Reference Wang, Li, Zhang, Varadínová, Jiang, Kučerová, Stejskal, Opit, Cao and Li2014). The phylogenetic tree was constructed in MEGA (version 5.2.1) software applying the neighbor-joining (NJ) and maximum parsimony (MP) methods.
Specific primer design, selection and sensitivity test
Species specific primers for each of the five studied Cryptolestes species were designed and evaluated as a new reliable method for rapid identification of storage Cryptolestes. First, the alignment of all haplotypes was visually inspected in Bioedit (version 7.1.3) and specific primers’ suitable areas were proposed according to three factors: (1) intraspecific uniformity of the proposed area, (2) diversity from other species in the proposed area and importantly, (3) according to the presence of unique single nucleotide polymorphism (SNPs). Final selection of at least three primer sets for each species from the proposed primer suitable areas was done in Oligo 7 (version 7.57). Internal stability, false priming sites, duplex formation, melting and optimal annealing temperatures were analyzed to ensure the quality of primers. All primers were synthesized by Sangon Biotech (Shanghai) Co., Ltd.
The specificity test of each primer pair in uniplex PCR assay was performed using the selection of 15 samples (table 1) which included ten larval haplotypes identified in this study and another five different haplotypes obtained from adult insects studied by Wang et al. (Reference Wang, Li, Zhang, Varadínová, Jiang, Kučerová, Stejskal, Opit, Cao and Li2014). PCR amplification in final reaction volume of 25 μl consisted of 12.5 μl MasterMix with dye, 10 μl ddH2O, 0.5 μl of specific forward primer, 0.5 μl of specific reverse primer and 1.5 μl of template DNA. In the first step of primer selection, PCR cycler conditions were initial denaturation at 94°C for 3 min, followed by 35 cycles of 94°C for 30 s, optimal annealing temperature (Ta) of primer pair for 30 s and 72°C for 30 s with the final extension at 72°C for 10 min. After separation in 1.5% agarose gel electrophoresis and staining in EB, products were confirmed under the UV light. If the primer set produced only species-specific amplicons under the above mentioned conditions, another PCR run under the same conditions, except for the annealing temperature set at 50°C in all cases, was performed. Repeating the primer specificity test for each haplotype three times – twice under the optimal conditions and once under sub-optimal (Ta = 50°C) but for all primer sets uniform conditions – enabled verification of reliability and reproducibility of the specific primers.
Sensitivity of the selected five Cryptolestes species-specific primers was determined for one haplotype from each species in the PCR run with a series of samples with decreasing DNA concentrations, while keeping the same concentration of primers. Adopted DNA concentrations, namely, 10, 5, 1, 0.5, 0.1 and 0.05 ng μl−1, were prepared by diluting 1 μl of original genomic extraction material with known DNA concentration in equivalent amount of TE buffer. In the case of the C. ferrugineus primer sensitivity test, we added another two concentrations, namely, 0.01 and 0.005 ng μl−1. Reaction volume for the PCR was again 25 μl, but the volume of individual components was the following: 12.5 μl MasterMix with dye, 10.5 μl ddH2O, 0.5 μl of specific forward primer, 0.5 μl of specific reverse primer and 1 μl of template DNA. The PCR conditions were as in the case of COI amplification and second run of primer specificity test. The validation of products was done using the 2% agarose gel electrophoresis.
Results
Cryptolestes larvae sequence analyses
The DNA concentration extracted from a single larva was 7.6 ng μl−1 on average and was enough to generate high quality PCR amplicons using the universal COI primers. We obtained 49 Cryptolestes larval COI sequences from eight populations in China and Czech Republic and 40 final sequences with 658-base pair length were used for further analysis. The phylogenetic analysis of our data and adult sequences (Wang et al., Reference Wang, Li, Zhang, Varadínová, Jiang, Kučerová, Stejskal, Opit, Cao and Li2014) revealed 100% congruence of larval sequences with expected species identification. Altogether, ten different larval haplotypes of five Cryptolestes species were revealed – one haplotype each of C. capensis (GenBank submission number KC977912), C. turcicus (KC977913) and C. pusilloides (KC977916); two haplotypes of C. ferrugineus (KC977914 and KC977915) and five haplotypes of C. pusillus (KC977917, KC977918, KC977919, KC977921 and KC977922).
Specific primer design, selection and sensitivity test
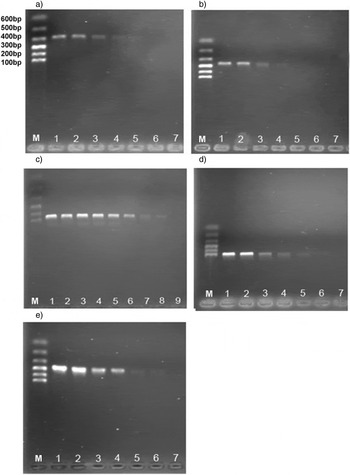
A set of 15 different Cryptolestes COI sequences, ten larval haplotypes obtained in this study and five different adult haplotypes obtained from GeneBank, were adopted for the purpose of developing an accurate and fast method for identifying five Cryptolestes species (fig. 1). This included one haplotype of C. capensis, two haplotypes each of C. turcicus and C. pusilloides, four haplotypes of C. ferrugineus and six haplotypes of C. pusillus (table 1). In the case of C. turcicus, two haplotypes represented populations from two continents (Asia, Europe). As for C. ferrugineus and C. pusillus, they represented populations from three continents (Asia, Europe and North America). The specificity test of the designed primers was assayed in uniplex PCR. Five primer pairs for reliable identification of five Cryptolestes species were selected and are listed in table 2. These primer pairs were designed to identify different Cryptolestes species regardless of their life stage or geographic origin. Although optimal annealing temperatures (Ta) for the selected primers varied from 50.2°C up to 52.7°C, we clearly demonstrated that each of them produced a species specific band without any other unwanted band or bands under the standardized conditions of Ta = 50.0°C (fig. 2). This indicates high specificity and reproducibility of primer pairs and a good potential for practical use.

Fig. 1. Aligned COI nucleotide sequences from five Cryptolestes species (C. capensis, C. turcicus, C. ferrugineus, C. pusilloides and C. pusillus) obtained in this study (accession numbers KC977912–KC977919, KC977921 and KC977922) and from the GenBank (accession numbers KC977933, KC977924, KC977925, KC436316 and KC977930). The dots indicate similarity between nucleotides. Framed regions indicate the positions of species-specific primers for each studied Cryptolestes species.

Fig. 2. Electrophoresis gels for five PCR reactions validating species specificity of five Cryptolestes primer pairs, Ta = 50°C. (a) Specific primers for C. capensis; (b) specific primers for C. turcicus; (c) specific primers for C. ferrugineus; (d) specific primers for C. pusilloides; (e) specific primers for C. pusillus. The order of samples and localities is the same as it is listed in table 1; 1: C. capensis; 2–3: C. turcicus; 4–7: C. ferrugineus; 8–9: C. pusilloides; 10–15: C. pusillus; 16: Control; M: DNA Marker I.
Table 2. Species specific primers designed for detection of five Cryptolestes species.

Because the concentration of genetic material extracted from one larva varied significantly among the samples and in few cases was no more than 0.15–0.30 ng μl−1, sensitivity of a selected specific primer set for each of the five studied species was determined using one sample from each species. In all species, DNA concentration of 10 and 5 ng μl−1 resulted in strong intensity bands. Lowering the concentration decreased the intensity of visualized bands markedly. Despite this, bands were still recognizable at concentration of 1 ng μl−1. Template concentrations of 0.1 and 0.05 ng μl−1 were not sufficient to produce visible amplicon except for C. ferrugineus specific primers, where 0.1 and 0.05 ng μl−1 concentrations still resulted in sufficient yield. Finally, the lower limit for detection was set to be 0.05 ng μl−1 for C. ferrugineus specific primers, 0.5 ng μl−1 for C. pusilloides and C. pusillus specific primers and 1 ng μl−1 for C. capensis and C. turcicus specific primers (fig. 3).

Fig. 3. Sensitivity tests for five specific primer sets: (a) C. capensis; (b) C. turcicus; (c) C. ferrugineus; (d) C. pusilloides; (e) C. pusillus. The concentration of template DNA from the lane 1 toward the lane 7 were 10, 5, 1, 0.5, 0.1, 0.05 ng μl−1 and control in the case of all species, except for C. ferrugineus where DNA concentrations from the lane 1 toward the lane 9 were following: 10, 5, 1, 0.5, 0.1, 0.05, 0.01, 0.005 ng μl−1 and control. Lane M: DNA Marker I.
Discussion
In the present study, we present a simple, rapid and accurate method based on DNA barcoding for identification of five common storage Cryptolestes species, namely, C. capensis, C. ferrugineus, C. pusilloides, C. pusillus and C. turcicus. Use of five primer pairs for the conventional PCR system is described and their specificity and sensitivity in the identification process is evaluated. In comparison with the study of Wang et al. (Reference Wang, Li, Zhang, Varadínová, Jiang, Kučerová, Stejskal, Opit, Cao and Li2014), the present identification system does not require sequencing, but only routine laboratory techniques such as DNA extraction, PCR and electrophoresis. Utilization of species-specific primers for identification is thus cost effective, time saving and does not compromise the reliability of identification. The PCR assay with the species-specific primers can be carried out within 3 h using unknown DNA of Cryptolestes species.
Molecular identification for Cryptolestes species using the species-specific primers proposed here plays an important role in routine species identification for pest management or quarantine operations, especially for the immature stages of insects which are generally tough to identify. Our data demonstrate that regardless of whether it is an adult or larva of Cryptolestes, it is possible to extract sufficient DNA for uniplex PCR assay with specific primers in order to perform the identification of the sample. We then speculate that the five species-specific primers of Cryptolestes can be feasible for identification of all ontogenetic stages.
When identifying species according to barcodes, misidentification is mostly caused by insufficient database information on the intraspecific or congeneric genetic diversity (Wiemers & Fiedler, Reference Wiemers and Fiedler2007). Reliability of species identification is dependent on the number of representative haplotypes included in barcode libraries (Meyer & Paulay, Reference Meyer and Paulay2005; Virgilio et al., Reference Virgilio, Backeljau, Nevado and de Meyer2010; Jinbo et al., Reference Jinbo, Kato and Ito2011). In the present study, two of five studied species, namely, C. capensis and C. pusilloides, were represented only by one geographical population. On the other hand, C. turcicus specific primers were evaluated on two geographically isolated populations (Asia and Europe). Furthermore, the analyzed genetic materials of C. ferrugineus and C. pusillus in both cases originated from four different populations from three geographically distant areas – Asia, Europe and North America. In all intraspecific populations from distinct continents, we identified different haplotypes of COI gene and designed primer pairs which are species-specific regardless of the origin of the population and the genetic variation within the species. Therefore, in our opinion, especially in the case of C. ferrugineus, C. pusillus and C. turcicus, the primer pairs tested in this study, can become universally applicable for the identification of Cryptolestes beetles of diverse geographic origins. Nevertheless, prospective research considering more geographical variability and larger sample sizes is necessary to support our assumption, especially for C. capensis and C. pusilloides. We also recognize that, only five species of flat grain beetles are included in our study, but generally, nine species are considered as stored-product pests (Rees, Reference Rees2004). To be of broad applicability, the sequences used for alignment and primer design should include more comprehensive taxon sampling over most of species’ distribution for such assay.
Sensitivity of primers was assessed using one haplotype per species. We found that the sensitivity of the five primer pairs was high and even a small quantity of genetic material of the studied Cryptolestes species can be easily detected. The detection limit was set to be different and varied from 1 ng μl−1 (C. capensis and C. turcicus) down to 0.5 ng μl−1 (C. pusilloides and C. pusillus) and 0.05 ng μl−1 (C. ferrugineus). This is particularly significant when checking imported stored products for the presence of Cryptolestes sample because it may be present in incomplete insect material which may not be sufficient to detect by universal primers.
Molecular identification of five Cryptolestes species using the species-specific COI markers proposed here represents a considerable improvement over currently available identification protocols. However, in the present study, we could not test the possible cross-reaction of the five species-specific primer pairs. Future work should also focus on developing the multiplex PCR, in which more than one pair of species-specific primer are used in a single PCR reaction.
Acknowledgements
This research was supported by the International Science and Technology Cooperation Program of China (2013DFG32350), the National Quality Supervision, Inspection and Quarantine Public Welfare Industry Research Project of China (201310075), the project KONTAKT – LH12160, the SVV 260 087/2014 project of the Charles University in Prague, and Oklahoma Agricultural Experiment Station (Project#: OKL02695). We thank Fan Jiang and Qianqian Yang from China Agricultural University for their excellent technical support with software analysis.