



Introduction
One of the consequences of rampant anthropization of landscapes is the increasing dependence of wildlife on human subsidies, a fact that may deeply shape foraging strategies and, ultimately, individual fitness (Oro et al. Reference Oro, Genovart, Tavecchia, Fowler and Martínez-Abraín2013 and references therein). A paradigmatic example of these modifications is the effects on avian scavengers. Subsidization increases the availability and predictability of resources both in space and time, which may directly affect populations and guild functionality and, ultimately, the ecological processes in which avian scavengers play a key role (Cortés-Avizanda et al. Reference Cortés-Avizanda, Jovani, Carrete and Donázar2012, Reference Cortés-Avizanda, Blanco, Devault, Markandya, Virani, Brandt and Donázar2016).
The potential effects of human subsidization may be particularly concerning for Old World vultures of the Gyps genus as these species have evolved in ecological systems characterized by randomly-distributed resources (Houston Reference Houston1974). Although some Gyps populations have recovered regionally, as in south-western Europe, most of the Old World Gyps populations have declined and many populations are below 10% of the original numbers, and thus the object of active conservation measures in many regions (Virani et al. Reference Virani, Kendall, Njoroge and Thomsett2011, Margalida and Ogada Reference Margalida and Ogada2018). The transcendence in the conservation of these species is disproportionately high; because of their large body-size, their sociality and relative higher abundance, they are among the main providers of ecosystem services (Rodríguez et al. Reference Rodríguez, Beard, Bennett, Cumming, Cork, Agard, Dobson and Peterson2006, Markandya et al. Reference Markandya, Taylor, Longo, Murty, Murty and Dhavala2008, Moleón et al. Reference Moleón, Sánchez-Zapata, Margalida, Carrete, Owen-Smith and Donázar2014, Pereira and Navarro Reference Pereira and Navarro2015, Sebastián-González et al. Reference Sebastián-González, Moleón, Gibert, Botella, Mateo-Tomás, Olea, Guimarães and Sánchez-Zapata2016, Gutiérrez-Cánovas et al. Reference Gutiérrez-Cánovas, Moleón, Mateo-Tomás, Olea, Sebastián-González and Sánchez-Zapata2020).
In Europe, human subsidization has been strongly dependent on changes to policies on sanitary regulations (Donázar et al. Reference Donázar, Margalida and Campión2009a). The Bovine Spongiform Encephalopathy (BSE) crisis led to the prohibition of the abandonment of livestock carcasses in the wild (EC 1774/2002; Donázar et al. Reference Donázar, Margalida, Carrete and Sanchez-Zapata2009b), which had widely-described detrimental consequences for scavenger species (Cortés-Avizanda et al. Reference Cortés-Avizanda, Carrete, Serrano and Donázar2009, Reference Cortés-Avizanda, Almaraz, Carrete, Sánchez-Zapata, Delgado, Hiraldo and Donázar2011, Reference Cortés-Avizanda, Jovani, Carrete and Donázar2012, Donázar et al. Reference Donázar, Cortés-Avizanda and Carrete2010, Moreno-Opo et al. Reference Moreno-Opo, Margalida, Arredondo, Guil, Martín, Higuero, Soria and Guzmán2010). Although still restrictive, subsequent regulations allowed the abandonment of livestock carcasses by farmers (CE 322/2003, CE 830/2005, CE 142/2011; Margalida et al. Reference Margalida, Carrete, Sánchez-Zapata and Donázar2012), which resulted in the recovery of demographic parameters of some avian scavenger populations (Margalida et al. Reference Margalida, Colomer and Oro2014, Donázar et al. Reference Donázar, Cortés-Avizanda, Ceballos, Arrondo, Grande and Serrano2020). In parallel, after having being closed during the sanitary crisis (Donázar et al. Reference Donázar, Margalida and Campión2009a), supplementary feeding stations, the so-called ‘vulture restaurants’, were re-opened and served with the aim of buffering the lack of food for avian scavengers (Cortés-Avizanda et al. Reference Cortés-Avizanda, Carrete and Donázar2010, Reference Cortés-Avizanda, Blanco, Devault, Markandya, Virani, Brandt and Donázar2016). Finally, during the last few decades, the growth of other sources of food such as landfills has been noticeable. These places were not created to feed wildlife but have unintentionally attracted individuals of numerous avian scavenger species, many of them threatened, such as vultures, with highly controversial consequences (Tauler et al. Reference Tauler, Real, Hernández-Matías, Aymerich, Baucells, Martorell and Santandreu2015, Plaza and Lambertucci Reference Plaza and Lambertucci2018).
Within this described scenario, to disentangle how Eurasian Griffon Vultures Gyps fulvus use trophic sources and subsidies of variable predictability is key to the future of science-based conservation decisions attaching European vulture populations. Moreover, it is necessary to know to what extent the actual network of legal feeding places is used by avian scavenger populations. It cannot be forgotten that conservation strategies of regional administrations on feeding management of avian scavengers in Spain pivot on the two legal alternatives existing currently, supplementary feeding places managed by authorities or NGOs, and extensive farms or hunting properties where carcass dumping has been authorised (Margalida et al. Reference Margalida, Oliva-Vidal, Llamas and Colomer2018). With these general purposes in mind, we take advantage of the information yielded by 35 adult Eurasian Griffon Vultures Gyps fulvus tagged with GPS/ACC devices in one of the most important areas for the target species in western Europe. In addition to a description of the general scenario, including legislative aspects, we hypothesise that both the intrinsic characteristics of the food sources and the individual traits would shape the use of potential feeding places by the tagged vultures. Specifically, we predict that the Griffon Vultures will feed preferentially at sites with larger amounts of food available and with a higher degree of predictability (Cortés-Avizanda et al. Reference Cortés-Avizanda, Carrete and Donázar2010). Even though the species show little reversed sexual dimorphism (Xirouchakis and Poulakakis Reference Xirouchakis and Poulakakis2008), there is evidence supporting differences in health and mortality linked to sex (Arrondo et al. Reference Arrondo, Sanz-Aguilar, Pérez-García, Cortés-Avizanda, Sánchez-Zapata and Donázar2020b, Gangoso et al. Reference Gangoso, Cortés-Avizanda, Sergiel, Pudifoot, Miranda, Muñoz, Delgado-González, Moleón, Sánchez-Zapata, Arrondo and Donázar2021) that may be associated to differences in micro-scale foraging behaviour. Consequently, we predict differences in resource use between males and females.
Methods
Focal species, study area and GPS data management
The Eurasian Griffon Vulture is a large obligate avian scavenger (up to 10–12 kg) adapted to travel long distances (Bildstein et al. Reference Bildstein, Bechard, Farmer and Newcomb2009, Harel et al. Reference Harel, Duriez, Spiegel, Fluhr, Horvitz, Getz, Bouten, Sarrazin, Hatzofe and Nathan2016a) in search of medium-sized and large vertebrate carcasses, mainly of domestic and wild ungulates (Ruxton and Houston Reference Ruxton and Houston2004). The species distribution covered Southern Europe, North Africa, the Middle East and Central Asia (Cramp and Simmons Reference Cramp and Simmons1980) but has drastically declined in most regions of the Western Palearctic. Currently, Spain holds the most abundant and healthiest European population with around 31,000 breeding pairs (Del Moral and Molina Reference Del Moral and Molina2018) with the bulk (>90%) of the European populations (Margalida et al. Reference Margalida, Donázar, Carrete and Sánchez-Zapata2010).
Our study was conducted in the upper Ebro Valley (northern Spain, covering parts of Aragón, Navarra and La Rioja regions; Figure 1). This is a region characterized by areas with little relief in the Ebro River Basin, many of them highly transformed for agricultural use, especially for irrigated crops (Lecina et al. Reference Lecina, Playán, Isidoro, Dechmi, Causapé and Faci2005). At the limits of the valley, there are mountains up to 1,500 m asl, generally covered with forests and grasslands (see Cortés-Avizanda et al. Reference Cortés-Avizanda, Colomer, Margalida, Ceballos and Donázar2015 for further details). Traditional sheep farming is still abundant although, in recent times, it has declined and been replaced by intensive farming, mainly of pigs (Martín-Queller et al. Reference Martín-Queller, Moreno-Mateos, Pedrocchi, Cervantes and Martínez2010). This region holds more than 8,000 breeding pairs of Eurasian Griffon Vultures (Del Moral and Molina Reference Del Moral and Molina2018) as well as healthy populations of other avian scavengers, thus constituting one of the most important regions for the conservation of scavengers in the Western Palearctic (see Cortés-Avizanda et al. Reference Cortés-Avizanda, Jovani, Carrete and Donázar2012).
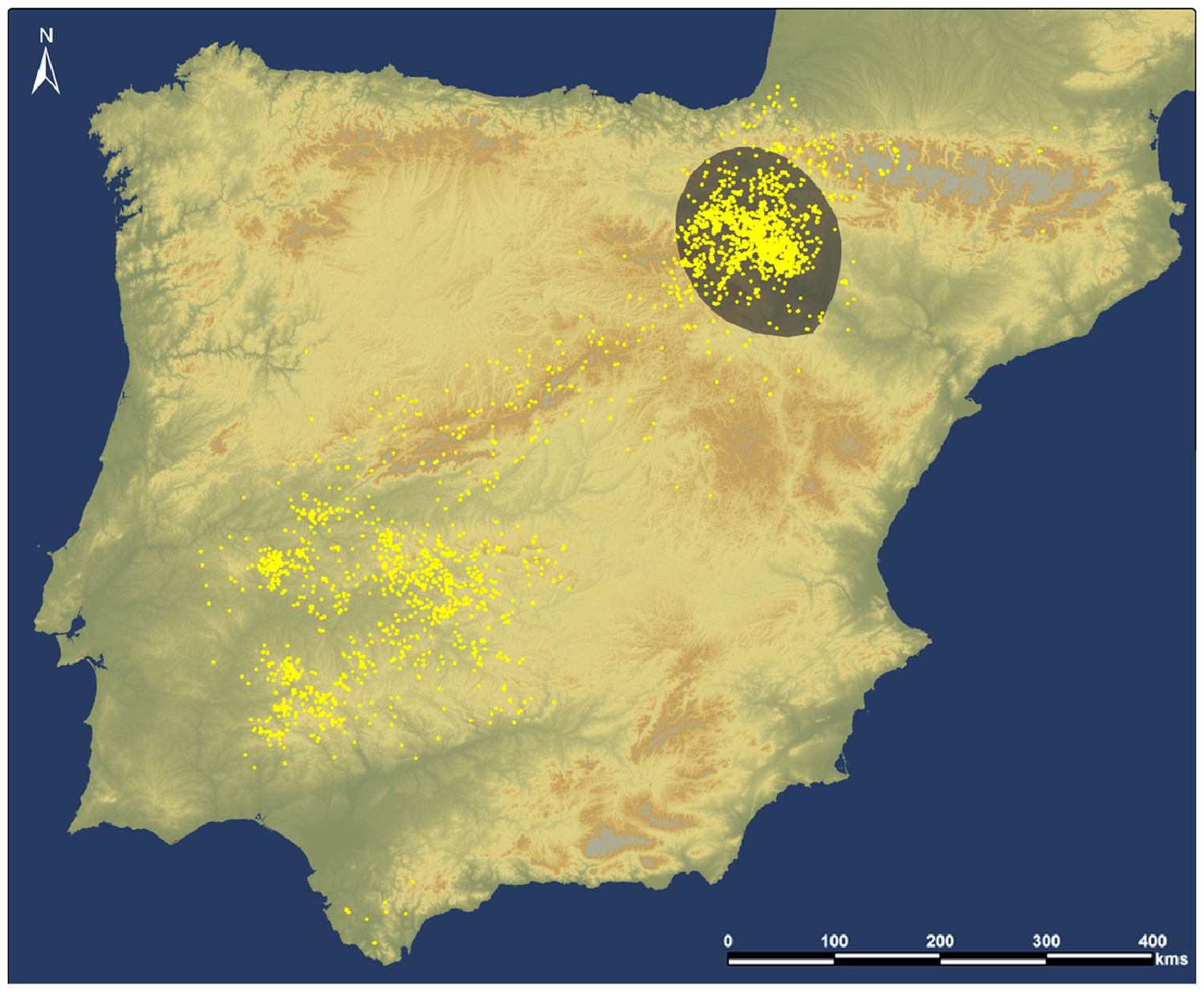
Figure 1. Study area. Yellow points represent the locations of all possible feeding events detected based on accelerometer algorithms (n = 6,300). In grey, we show our study area in the upper Ebro Basin (covering parts of Aragón, Navarra and La Rioja regions) where 72% of all possible feeding events concentrate (n = 4,553).
We captured 35 adult Griffon Vultures with cannon nets between 2015 and 2016 in the Bardenas Reales of Navarra Natural Park and Biosphere Reserve (north-eastern Spain; Figure 1). We equipped all individuals with 90g GPS/GPRS-GSM devices (E-obs digital telemetry; http://www.e-obs.de) attached by means of Teflon harnesses. The total system weighed less than 1% of the weight of the birds so that harmful effects were not expected (Sergio et al. Reference Sergio, Tavecchia, Tanferna, López Jiménez, Blas, De Stephanis, Marchant, Kumar and Hiraldo2015). All birds were sexed using molecular techniques (Wink et al. Reference Wink, Sauer-Gürth, Martinez, Doval, Blanco and Hatzofe1998). All the tagged birds were recorded breeding within a 50 km radius from the trapping point in years after marking. In those years when they did not nest, the individuals maintained their main centres of activity in colonies also within this same area (Arrondo et al. Reference Arrondo, Sanz-Aguilar, Pérez-García, Cortés-Avizanda, Sánchez-Zapata and Donázar2020b, Gangoso et al. Reference Gangoso, Cortés-Avizanda, Sergiel, Pudifoot, Miranda, Muñoz, Delgado-González, Moleón, Sánchez-Zapata, Arrondo and Donázar2021). Consequently, for each bird, and by means of field observations and accelerometer-GPS information, we determined the position of the active nests or, when the bird did not breed, the position of the main roosting site calculated as the main point within all nocturnal locations, managed with the ‘dbscan’ function from the fpc R package (Hennig Reference Hennig2019).
We programmed the GPS/GPRS-GSM devices to be activated one hour before sunrise and turned off one hour after sunset. The setting of the GPS was variable depending on weather conditions (Appendix S1 in the online supplementary material). Moreover, the accelerometers recorded data with a 10-second burst every 2.5 minutes with a frequency of 8.33 Hzs, except when we detected extremely low battery levels.
On the basis of the information of the accelerometers and by means of a supervised learning algorithm (http://accapp.move-ecol-minerva.huji.ac.il/; Nathan et al. Reference Nathan, Spiegel, Fortmann-Roe, Harel, Wikelski and Getz2012, Spiegel et al. Reference Spiegel, Harel, Getz and Nathan2013, Resheff et al. Reference Resheff, Rotics, Harel, Spiegel and Nathan2014, Harel et al. Reference Harel, Horvitz and Nathan2016b) we determined when the tagged birds were displaying apparent feeding behaviour on carcasses (Arkumarev et al. Reference Arkumarev, Dobrev, Stamenov, Terziev, Delchev and Stoychev2021a,Reference Arkumarev, Dobrev, Stamenov, Terziev, Delchev and Stoychevb). In total, we detected 6,300 potential feeding events in the Iberian Peninsula (Figure 1). We focused our study in the upper Ebro Basin, a region of 21,744 km2 where the breeding colonies and the bulk of the foraging activities of the birds concentrate (Arrondo et al. Reference Arrondo, Navarro, Perez-García, Mateo, Camarero, Martin-Doimeadios, Jiménez-Moreno, Cortés-Avizanda, Navas, García-Fernández, Sánchez-Zapata and Donázar2020a). The study area was defined trying to maximise the number of feeding sites and feeding events from the main breeding colonies and the capture area in Bardenas Reales Natural Park. Thus, our study area holds 4,553 (72%) of the total potential feeding events in Iberia. We visited 3,594 of them (78% of the study area) and found evidence of vultures feeding at 2,896 points (80% of the visited). In the rest of visited events, we could not determine if the vultures fed or not. From this departing point and considering that the birds repeatedly visited many of the locations where feeding events were identified (likely due to regular carcass disposal or abandonment), we defined 652 independent feeding sites, each one involving one or more feeding events.
During the field inspection of the confirmed feeding events, we identified the nature of the food source (hereafter “Resource”). We classified it into five categories: random carcass, carcass dumping site, intensive farm, extensive farm, and landfill (see Figure 2 for description of each category). Based on bone and skin remains, we also identified the species’ identity and the minimum number of carcasses present. Finally, thanks to the information provided by the regional administrations in the study area (Navarra, Aragón and Rioja), we determined the legal status of each place where carcasses were available. We established three categories: a) Lawful: extensive livestock exploitations where farmers have been authorized to abandon carcasses on the basis of regional dispositions (CE 1069/2009, CE 142/2011); b) Lawful-Government managed: supplementary feeding stations (vulture restaurants) created and maintained mainly by administrations; c) Lawless: sites without legal coverage; this includes intensive farms as well as extensive farms not under regulations allowing the abandonment of livestock carcasses. This also includes carcass dumping sites that have not been legalized. The information relative to the legal status of the feeding sites was determined departing from databases yielded by the regional administrations of Navarra, La Rioja and Aragón.

Figure 2. Types of feeding resources. Apart from the basic description, we detail their spatial and temporal predictability and if the disposal of animal remains for avian scavengers may be under legal coverage. Photos: from top to bottom; the top 3: José A. Donázar; 4: H. Garrido, 5: J.L. Rodriguez
Analytical procedures and model fitting
As a prior step for analytical purposes, and following the procedures of García-Alfonso et al. (Reference García-Alfonso, van Overveld, Gangoso, Serrano and Donázar2020), we split the data into two “Periods”: (1) from October to February (n = 5 months), corresponding to autumn-winter and (2) from March to September (n = 7 months), corresponding to spring–summer. These two periods correspond roughly to non-breeding and breeding seasons, respectively. We decided to split the year unevenly in order to maximize the breeding season since in September nestlings may still be dependent on adults if they result from late laying in March. Consequently, for the entire study we had data on five different periods (from October 2015 to September 2017), with each being a sampling unit. Fitting these periods as a random term (see below) allowed us to account for temporally contingent variation of the information.
We performed two sets of analyses based on two response variables calculated as proportion data:
(1) FEEDING SITE as the number of different tagged vultures present at each feeding site during each “Period” (numerator), controlling for the total number of tagged individuals with available information for that “Period” (denominator).
(2) VULTURE, indicating the individual use of resources, as the number of days each vulture visited each feeding site per “Period” (numerator), controlling for the total number of days with available information for the same vulture during that “Period” (denominator).
We defined 13 explanatory variables (Table 1) to evaluate seasonal variability, features of feeding sites (physiography and anthropization) and individual traits of focal vultures. “Resource” and “Biomass” covaried (Kruskal-Wallis χ2= 444.09, P <0.01) but as these two variables evaluated different features of the trophic availability to vultures, we decided to fit both alternatively in separate modelling procedures. Hence, we performed four different procedures according to combinations of “Resource” or “Biomass” for both FEEDING SITES and VULTURES. Also, anthropization, measured as the Human Footprint Index (HFP, Venter et al. Reference Venter, Sanderson, Magrach, Allan, Beher, Jones, Possingham, Laurance, Wood, Fekete, Levy and Watson2016), covaried with “Resource”, so “HFP” only was fitted in the “Biomass” models. Finally, we also detected that Kernel Density Estimator 95% (“KDE95”) covaried with Kernel Density Estimator 50% (“KDE50”), so both variables were independently fitted into the models. As we said above, the “Biomass” available at landfills was impossible to estimate so all the data related to this “Resource” (n = 16) were removed from the dataset used in the “Biomass” models (see above for details). In all the models, “Biomass” was fitted both as a linear and as a quadratic term and we only included the interactions of explanatory variables that we considered could have biological significance: “Season:Resource” and “Season:Biomass” in the FEEDING SITE analysis and “Sex:Biomass”, “Sex:Resource” and “Distance:Resource” in the VULTURE analysis.
Table 1. Explanatory variables used to determine the selection of feeding sites by tagged griffon vultures.

(*) Used in VULTURE analysis
(**) Used in FEEDING SITE and VULTURE analyses
Both response variables were modelled as proportion data with a denominator by means of Generalized Linear Mixed Models (GLMMs) with binomial error distribution and logit link function. “Feeding site ID” and “Period ID” were fitted as random terms within the FEEDING SITE analysis. Additionally, “Individual ID” was fitted as a random term within the VULTURE analysis (Table 1). We standardised all continuous explanatory variables to a mean of 0 with a variance of 1 by subtracting the means and dividing by the standard deviations (Gelman and Hill Reference Gelman and Hill2007, Bolker et al. Reference Bolker, Brooks, Clark, Geange, Poulsen, Stevens and White2009, Bolker Reference Bolker2020). Models were fitted with all possible combinations of explanatory variables (from only the intercept to all considered fixed effects), except for pairs of variables with a Spearman’s correlation coefficient higher than |0.5| that were never included in the same model to avoid collinearity problems (Graham Reference Graham2003). We based the model selection on the Akaike´s Information Criterion corrected for small sample sizes (AICc) (Sugiura Reference Sugiura1978). The resulting models with ΔAICc <2 were averaged by means of a model averaging procedure. All statistical analyses were performed using R statistical software version 3.6.0 with the stats package (R Core Team 2013) for confidence intervals, lme4 package (Bates et al. Reference Bates, Maechler, Bolker and Walker2015) for the GLMM analysis, AICcmodavg for model ranking (Mazerolle Reference Mazerolle2019) and the dredge function from the MuMIn package (Barton Reference Barton2019) for model selection. Moran´s I index from the nfc package (Bjornstad and Cai Reference Bjornstad and Cai2019) were used to examine the spatial autocorrelation (Appendix S2).
Results
Of the 652 places visited by the targeted birds, 42% correspond to carcass dumping sites and 29%, 17.5%, 9% and 2.5% to random carcasses, intensive farms, extensive farms and landfills, respectively (Figure 3). Attending to the total number of visits made to each type of resource, it was observed that 36% were concentrated in carcass dumping sites, followed by intensive farms (32%), extensive farms (11.6%), landfills (11%) and random carcasses (9.4%). Moreover, most of the biomass was found in carcass dumping sites and intensive farms (Figure 3). Carcasses found at intensive farms were mainly pigs whereas at random feeding sites, half of the available biomass was sheep, as also occurs in extensive farms (Table 2). In carcass dumping sites many different species were found, including wild ungulates disposed of by hunters (Table 2). Remarkably, only 9% of the total feeding sites visited had legal coverage with 5% corresponding to carcass dumping sites and 4% to extensive farms (Figure 3). In relation to the total number of extensive farms and carcass dumping sites, 34.5% and 60.7% of them, respectively, had legal coverage.
Table 2. Percentages of different prey species identified in each type of “Resource” (landfills excluded). Based on the minimum number of individuals identified by bone and skin remains.


Figure 3. Use of different types of food resources by adult GPS-tagged griffon vultures. Above: Percentages of feeding points belonging to each category; in light grey/red: proportion of these points where the disposal of carcasses for avian scavengers was under legal coverage. Centre: Number of visits performed by individual vultures to each food resource. Below: amount of “Biomass” (kg) per category (logarithmic scale).
Modelling of FEEDING SITE with the variable “Resource” yielded two alternative models with ΔAICc <2 (Appendix S3). The model averaging showed that livestock dumping sites seemed to gather more feeding birds, particularly in summer, and that in this season landfills also received a higher number of tagged individuals (Table 3). When we fitted “Biomass” (instead of “Resource”) we obtained three alternative models. Averaging showed a quadratic response of “Biomass”, indicating that greater numbers of vultures preferentially visited those feeding sites with intermediate amounts of food, especially in summer (Appendix S3, Table 3).
Table 3. Results of the full coefficients of model average for the response variables FEEDING SITES and VULTURE. Informative parameters, i.e. variables explaining sufficient deviance to provide a net reduction in AICc (Burnham and Anderson Reference Burnham and Anderson2002, Arnold Reference Arnold2010) according to the confidence interval 85%, are shown in bold. Rs = Resource; Ss = Season; Br = Breeding status.

The VULTURE approach including “Resource” yielded eight alternative models (Appendix S3). Model averaging showed that landfills, carcass dumping sites and intensive farms were more visited by breeding birds and during winter. The fitting of “Biomass” (instead of “Resource”) resulted in seven alternative models (Appendix S3). Model averaging (Table 3) revealed again a quadratic response of “Biomass”. Also, more visits took place close to the nest or the main roosting site, during winter, and by breeding birds. Notably, males visited sites with more “Biomass” more frequently (Figure 4). Finally, there was a slight positive relationship to the degree of anthropization (human footprint). Moreover, it can be highlighted that the standard deviation associated to the random factors “Individual” and “Semester” was notably higher than that relative to “Site”, revealing important individual and temporal variations in the observed patterns of resource use (Appendix S4).

Figure 4. Predictive values of the models relative to the response variable VULTURE. We show the change in probability of use in relation to the amount of carrion (“Biomass”) for individuals of both sexes and during both seasons. Left: Breeding birds; Right: Non-breeding birds.
Discussion
Our results show that the GPS-tagged vultures preferably visited those feeding sites with high predictability, both in space and time, of resources such as carcass dumping sites and intensive farms. Indeed, we found that most of the visited sites did not have legal coverage (91%). Moreover, the use of feeding sites was not homogeneous between individual birds, with effects of environmental variables (“Season”) and individual traits (“Breeding status” and “Sex”) (see also Moreno-Opo et al. Reference Moreno-Opo, Trujillano and Margalida2015). Furthermore, we found that the response of our tagged birds to the amount of carrion (“Biomass”) was quadratic, suggesting that factors operating in highly clumped places, such as intraspecific competition (see below), may be playing a key role in the selection of feeding places by these obligate top avian scavengers. All of these results have decisive implications from a conservation point of view and open questions on how to address the management of the multiplicity of potential food resources, many of which are not under legal coverage.
We detected that intensive farms and carcass dumping sites may be equally attractive to Griffon Vultures, coinciding with higher availability of biomass. However, it was remarkable that intensive farms showed relatively higher numbers of visits in relation to their frequency within the total number of feeding sites. This finding is closely related to another striking result: the quadratic response to the variable “Biomass”, which is consistent regardless of the analytical approach (FEEDING SITE or VULTURE). In our opinion, these results may be conditioned by the existence of high intraspecific competition at sites with very abundant and clumped resources, places that are known to attract larger numbers of feeders (mainly juveniles, García-Heras et al. Reference García-Heras, Cortés-Avizanda and Donázar2013, van Overveld et al. Reference van Overveld, García-Alfonso, Dingemanse, Bouten, Gangoso, la Riva, Serrano and Donázar2018). Moreover, points with a lesser amount of food (e.g. intensive farms) may have daily discharges of carcasses, compared to points with higher amounts (e.g. carcass dumping sites) where disposals are more spaced through time. In addition, and very interestingly, this quadratic response of vultures to the biomass showed sex asymmetries, with males visiting sites with larger amounts of food more frequently. The stress of facing anthropogenic disturbances may be sex-dependent (e.g. Wasser et al. Reference Wasser, Houston, Koehler, Cadd and Fain1997, Hayward et al. Reference Hayward, Booth and Wasser2010, Ahlering et al. Reference Ahlering, Maldonado, Eggert, Fleischer, Western and Brown2013, Strasser and Heath Reference Strasser and Heath2013). Previous research done with the same marked Griffon Vultures has hypothesized that male Griffon Vultures could be bolder than females when facing risky situations (Gangoso et al. Reference Gangoso, Cortés-Avizanda, Sergiel, Pudifoot, Miranda, Muñoz, Delgado-González, Moleón, Sánchez-Zapata, Arrondo and Donázar2021). It is also possible that, despite the low reversed sexual dimorphism, males could be displaced by dominant females from the more favourable (less crowded) feeding places as has been observed in other species of vultures (van Overveld et al. Reference van Overveld, García-Alfonso, Dingemanse, Bouten, Gangoso, la Riva, Serrano and Donázar2018).
The avoidance of crowded feeding sites may also explain why we found an appreciable use of randomly distributed resources. It cannot be disregarded that this category is of greater importance than indicated by the accelerometer data since small items can be quickly consumed and disappear without a trace, especially since our field survey effort has been lower in high mountain areas where in summer there are important cattle herds (MARM 2011). The reported random carcasses corresponded mostly to domestic ungulates (mainly sheep) maintained in extensive and semi-extensive regimes, a particular characteristic of the traditional agro-grazing systems of the steppe cereal landscapes of the Ebro Valley and surrounding Mediterranean mountains ( Donázar et al. Reference Donázar, Naveso, Tella and Campión1996, Cortés-Avizanda et al. Reference Cortés-Avizanda, Jovani, Carrete and Donázar2012). These carcasses correspond to animals that die during herds transit to pasture areas. Consequently, the overall importance of extensive livestock may be greater than the specific category reflected in our analyses. In this sense, we should add that, although our work covers a very important proportion of the feeding events of the GPS-tagged vultures, it is true that the birds perform long-distance displacement to other regions of Iberia (Figure 1).
On the other hand, a portion of the random carcasses corresponded to wild mammals, mainly wild rabbits Oryctolagus cuniculus. The importance of this species (despite its low weight; 1 kg) has increased during the last decades probably because of the increasing competition for food resources among avian scavengers in the study area (see Donázar et al. Reference Donázar, Cortés-Avizanda and Carrete2010). Moreover, experimental approaches made in the study area with rabbit carcasses show how up to the 30% of these baits were consumed by Griffon Vultures (Donázar et al. Reference Donázar, Cortés-Avizanda and Carrete2010). In addition, as is logical, the time of consumption of a rabbit carcass is very short (a few minutes) all which highlights how these randomly found resources can be underestimated. In general, the remains of other wild mammals, like wild boars Sus scrofa, were of little importance. In our study area, rewilding processes, understood as abandonment of land uses (Jørgensen Reference Jørgensen2015, Perino et al. Reference Perino, Pereira, Navarro, Fernández, Bullock, Ceauşu, Cortés-Avizanda, Van Klink, Kuemmerle, Lomba, Pe’er, Plieninger, Benayas, Sandom, Svenning and Wheeler2019) is widespread, especially in mountain areas, but contrary to what happens in other regions of Iberia (Martin-Díaz et al. Reference Martin-Díaz, Cortés-Avizanda, Serrano, Arrondo, Sánchez-Zapata and Donázar2020), hunting activity is comparatively lower and the remains of hunted animals are generally moved to carcass dumping sites.
Although there are only 16 landfills in our study area, we observed a remarkable use of these places by our tagged vultures. The use of landfills by avian scavengers was described decades ago (Donázar Reference Donázar1992) but the frequency of the phenomenon seems to have increased progressively (Tauler-Ametller et al. Reference Tauler-Ametller, Hernández-Matías, Pretus and Real2017, McGrady et al. Reference McGrady, Karelus, Rayaleh, Sarrouf Willson, Meyburg, Oli and Bildstein2018, Plaza and Lambertucci Reference Plaza and Lambertucci2018), probably due to the increase in landfill discharges, the growth of vulture populations and the intraguild competition for resources (Donázar et al. Reference Donázar, Cortés-Avizanda and Carrete2010). It was impossible to determine the amount of resources available in landfills but it is reasonable to think that it is certainly lower that in carcass dumping sites and intensive farms, so the preference of birds when selecting these sites could be explained by social context, which creates the highest predictability (discharges of new garbage occur on a daily basis). The conservation consequences of this attraction to landfills may be important. Individuals that frequent landfills are exposed to a higher risk of pathogen infections, suffer bioaccumulation of certain contaminants or death by ingestion of baits with rodenticides, pharmacy/veterinarian drugs and other chemical compounds (Plaza and Lambertucci Reference Plaza and Lambertucci2017, Tongue et al. Reference Tongue, Reynolds, Fernie and Harrad2019).
Environmental effects also contribute to the observed patterns of resource use. In relation to seasonality, an apparent contradiction appears, since the probability that a feeding site was visited increased in summer (FEEDING SITE), but the intensity of use (VULTURE) was higher in winter for breeding birds. This matches with the patterns found by (Arkumarev et al. Reference Arkumarev, Dobrev, Stamenov, Terziev, Delchev and Stoychev2021a) in a Bulgarian population of Griffon Vultures. During summer, the vultures would leverage better weather conditions to perform long-distance flights showing an increase in their home ranges (Harel et al. Reference Harel, Duriez, Spiegel, Fluhr, Horvitz, Getz, Bouten, Sarrazin, Hatzofe and Nathan2016a), which would facilitate the use of more feeding sites. During winter, on the contrary, worse flight conditions and attachment to early breeding, tasks demanding high nest attendance (pair courtship, incubation), would determine the intensive exploitation of resources located at the vicinity of the colonies (see also Martin-Díaz et al. Reference Martin-Díaz, Cortés-Avizanda, Serrano, Arrondo, Sánchez-Zapata and Donázar2020). We also found a slight positive effect of human footprint reinforcing previous findings showing the dependence of this avian scavenger population on resources located in very anthropized landscapes (Arrondo et al. Reference Arrondo, Sanz-Aguilar, Pérez-García, Cortés-Avizanda, Sánchez-Zapata and Donázar2020b, Cortés-Avizanda et al. Reference Cortés-Avizanda, Colomer, Margalida, Ceballos and Donázar2015).
Finally, there is a non-negligible variability linked to the individual and to time (estimated through the contribution of random factors) that merits discussion. This suggests that there may be individual specialization in the use of certain resources, something that has been reported among several taxa (Bolnick et al. Reference Bolnick, Svanbäck, Fordyce, Yang, Davis, Hulsey and Forister2003) but which seems surprising in a scavenger bird that is also social at feeding grounds. On the other hand, the temporal variation (accounted in “Period”) points to possible changes in the availability of resources throughout the study period. This could, for example, be linked to changes in livestock mortality determined by the environment (Vitali et al. Reference Vitali, Segnalini, Bertocchi, Bernabucci, Nardone and Lacetera2009). Obviously, future research is necessary to deep into these relevant questions.
Perspectives
Our results suggest that the feeding points supported by current sanitary regulations contribute very little to covering the food demand of the studied population of Eurasian griffon vultures, since less than 10% of the feeding events took place at these sites, included in this percentage those supplementary feeding stations created for conservation purposes. In part, this is because these new sanitary regulations have been applied irregularly by the regional administrations of the study area (Donázar et al. Reference Donázar, Cortés-Avizanda, Ceballos, Arrondo, Grande and Serrano2020). It highlights the current difficulties represented by the irregular transposition of the European regulations to the regions thus compromising desirable transboundary conservation strategies through increasing the authorized points of safety carrion and favoring more natural sources ( Donázar et al. Reference Donázar, Margalida and Campión2009a, Margalida and Colomer Reference Margalida and Colomer2012, Margalida et al. Reference Margalida, Oliva-Vidal, Llamas and Colomer2018, Arrondo et al. Reference Arrondo, Moleón, Cortés-Avizanda, Jiménez, Beja, Sánchez-Zapata and Donázar2018).
The existence and maintenance of food sources without legal coverage is a double-edged sword from a conservation point of view. Quantitatively, these may have favored demographic parameters, in particular preadult survival, as has been found in other vulture species (Oro et al. Reference Oro, Margalida, Carrete, Heredia and Donázar2008). In fact, the spectacular growth (more than 10 times higher) of the Griffon Vulture populations in northern regions of Spain in the last four decades could be partially explained by extraordinary availability of food yielded by intensive farms and clumped dumping sites (Donázar and Fernández Reference Donázar and Fernández1990, Parra and Tellería Reference Parra and Tellería2004, Del Moral and Molina Reference Del Moral and Molina2018). Currently, however, Griffon Vulture populations in our study area and probably in other regions of Europe, although still apparently healthy (but see Gangoso et al. Reference Gangoso, Cortés-Avizanda, Sergiel, Pudifoot, Miranda, Muñoz, Delgado-González, Moleón, Sánchez-Zapata, Arrondo and Donázar2021), are suffering from heavy unnatural mortality linked to foraging activities in very anthropized areas where intensive farming predominates (Arrondo et al. Reference Arrondo, Sanz-Aguilar, Pérez-García, Cortés-Avizanda, Sánchez-Zapata and Donázar2020b). In addition, to feed on intensively-reared livestock increases the risk of intoxication and diseases because of the existence of pharmaceuticals and pathogens in the remains of the abandoned carcasses (Casas-Díaz et al. Reference Casas-Díaz, Cristòfol, Cuenca, Agustí, Carneiro, Marco, Lavín and Margalida2016, Blanco et al. Reference Blanco, Junza and Barrón2017, Reference Blanco, Cortés-Avizanda, Frías, Arrondo and Donázar2019) to the point that they can cause large-scale extinctions (Watson et al. Reference Watson, Gilbert, Oaks and Virani2004, Gilbert et al. Reference Gilbert, Watson, Virani, Oaks, Ahmed, Chaudhry, Arshad, Mahmood, Ali and Khan2006).
We highlight the need for research focusing on the feasibility of long-term stability of the population of Griffon Vultures in our study area and in many regions of Spain where the birds increasingly depend on intensive farming located in anthropized areas. It is clear that the transposition of permissible sanitary regulations is currently insufficient. Novel initiatives are required to extend permissive sanitary regulations to the largest possible number of farmers maintaining extensive exploitations but it must be taken into account that this likely cannot be the sole solution in the long-term as these farming practices are in widespread regression (Martín-Queller et al. Reference Martín-Queller, Moreno-Mateos, Pedrocchi, Cervantes and Martínez2010). In addition, and because of the health of avian scavenger populations, it would be desirable to implement the regulations prohibiting the abandonment of carcasses of intensively farmed animals. This cannot be done, however, without carefully evaluating the possible effects of these measures on the demography of the vulture populations and on the perception of farmers towards the services provided by the birds (Cortés-Avizanda et al. Reference Cortés-Avizanda, McKeen, Ceballos, Pereira and Martín-López2022). It could be argued that the loss of this resource could be partially compensated for by the carcasses of wild ungulates currently under expansion in many mountain areas (Blázquez and Sánchez-Zapata Reference Blázquez, Sánchez-Zapata, Donázar, Margalida and Campión2009, Sánchez-Zapata et al. Reference Sánchez-Zapata, Eguía, Blázquez, Moleón and Botella2010, Margalida et al. Reference Margalida, Colomer and Sanuy2011, Cortés-Avizanda et al. Reference Cortés-Avizanda, Colomer, Margalida, Ceballos and Donázar2015). This, however, deserves more research because as noted above, this can be very dependent on hunting practices. In addition, carcasses resulting from predation events that are important in other European regions (Arkumarev et al. Reference Arkumarev, Dobrev, Stamenov, Terziev, Delchev and Stoychev2021b) are not available in our study area and large regions of Spain because large predators have long been extirpated.
Our results draw an uncertain picture for the medium-term future of scavenger birds in increasingly transformed European regions. We believe that, in the long-term, the conservation of vulture populations must be framed by "One Health" strategies that ensure a supply of livestock carrion free of veterinary drugs (Margalida et al. Reference Margalida, Bogliani, Bowden, Donázar, Genero, Gilbert, Karesh, Kock, Lubroth, Manteca, Naidoo, Neimanis, Sánchez-Zapata, Taggart, Vaarten, Yon, Kuiken and Green2014a). On the other hand, and also in the long term, it is necessary to prevent possible risks arising from profound socio-economic changes affecting the ecosystem services provided by vultures to farmers. It will be necessary to carefully assess the ecological role that both intensive and extensive livestock farming practices have in the viability of vulture populations as well as the risks associated with the potential disruptions affecting this complex and fragile system.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0959270921000575
Acknowledgements
We thank L. Lopo, J.M Vadillo from the Navara government and M. Alcántara from the Aragon regional government, Consultora CMC Sistemas de Mejora, S.L. for providing information about the authorised feeding points in their corresponding region. Funding was provided by the Comunidad de Bardenas Reales de Navarra and the project RTI2018‐099609‐B‐C21 (Spanish Ministry of Economy and Competitiveness and EU/FEDER). ACA was supported by a PostDoc contract of Excellence project of Junta de Andalucía and a PostDoc contract EMERGIA of Junta de Andalucía.









