Introduction
Bird extinctions are rare occurrences on continental landmasses compared to islands (Manne et al. Reference Manne, Brooks and Pimm1999, Sekercioglu et al. Reference Sekercioglu, Daily and Ehrlich2004). Over 90% of bird extinctions during the past 400 years have been of island-restricted species (King Reference King1978–1979, Stattersfield Reference Stattersfield and Mountfort1987, Johnson and Stattersfield Reference Johnson and Stattersfield1990). Compared to continental species, island birds have generally lost some defences to potentially detrimental factors including habitat destruction, lower numbers, introduced species, naiveté to predators and humans, as well as demographic constraints and ranges restricted to a small land area (Diamond and Veitch Reference Diamond and Veitch1981, Johnson and Stattersfield Reference Johnson and Stattersfield1990, Manne et al. Reference Manne, Brooks and Pimm1999, Blackburn et al. Reference Blackburn, Cassey, Duncan, Evans and Gaston2004, Duncan and Blackburn Reference Duncan and Blackburn2007). Avian population declines can be damaging to ecosystems in which birds historically were the primary, and sometimes only, terrestrial vertebrates to facilitate essential ecological process as consumers, predators, pollinators, and seed dispersers. For example, declining bird populations in the Mariana Islands were shown to adversely affect native seed dispersal while releasing depredation pressure on spider populations (Rogers et al. Reference Rogers, Hille Ris Lambers, Miller and Tewksbury2012, Caves et al. Reference Caves, Jennings, Hille Ris Lambers, Tewksbury and Rogers2013, Wandrag et al. Reference Wandrag, Dunham, Duncan and Rogers2017).
In the Mariana Islands, 12 native bird species are currently federally listed as Endangered and nine species are listed as ‘Critically Endangered’, ‘Endangered’ or ‘Extinct in the Wild’ on the IUCN Red List of Threatened Species (IUCN 2019). Understanding the status of a species is particularly important for island endemic and rare species and is required by land managers and policy makers to inform decisions related to conservation. Surveys based on robust methodologies permit reliable estimation of species status, which can be used to set baselines, assess population trends, establish population threshold levels, and set alert limits for management actions and priorities (Nichols and Williams Reference Nichols and Williams2006). Population status can also be used to focus conservation effort and funding where they are most needed and where they will be most effective, as well as update and inform local, regional, and global extinction risks. The first quantitative bird surveys in the Mariana Islands were conducted in 1981 and 1982 as part of the Micronesian forest bird surveys organized by the U.S. Fish and Wildlife Service (USFWS; Engbring and Ramsey Reference Engbring and Ramsey1984, Engbring et al. Reference Engbring, Ramsey and Wildman1986). Surveys were conducted on Guam, Rota, Aguiguan, Tinian, and Saipan and have served as a baseline for population status and trend assessment. The USFWS and Commonwealth of the Northern Mariana Islands (CNMI) Department of Lands and Natural Resources Division of Fish and Wildlife (DFW) have subsequently repeated the surveys several times, although with varying frequency and coverage owing to the difficult logistics and expense involved in sampling these remote islands.
Saitoh et al. (Reference Saitoh, Cibois, Kobayashi, Pasquet and Thibault2012) described four reed-warbler species for the Mariana Islands; the Guam Reed-warbler Acrocephalus luscinius, Aguijuan Reed-warbler A. nijoi, hereby referred to as the Aguiguan Reed-warbler, Nightingale Reed-warbler A. hiwae, and Pagan Reed-warbler A. yamashinae. This contrasts with the previous taxonomic classification by Mayr et al. (Reference Mayr, Traylor and Watson1986) who designated only one species, the Nightingale Reed-warbler Acrocephalus luscinia, split into three subspecies. The Nightingale Reed-warbler under the previous classification was federally listed as endangered in 1970 due to its disappearance on Guam and concern about its status in the CNMI (USFWS 1970). Historically the Nightingale Reed-warbler occurred on five islands in the Mariana archipelago: Guam, Aguiguan, Saipan, Pagan, and Alamagan. Prehistorically, a species of reed-warbler occurred on Tinian (Steadman Reference Steadman1995, USFWS 1998). Currently, following the taxonomy of Saitoh et al. (Reference Saitoh, Cibois, Kobayashi, Pasquet and Thibault2012), only the Nightingale Reed-warbler A. hiwae is extant and the other three species are believed to be extinct on Guam, Aguiguan and Pagan. We provide a description of island-specific reed-warbler trends in Appendix S1 in the online supplementary material.
Here we present results from resource surveys of the islands of Aguiguan, Alamagan, and Pagan in the Mariana Islands conducted between 2008 and 2010 to evaluate potential resource effects from proposed military training in the region. This was primarily an opportunity to conduct a range-wide survey of the Nightingale Reed-warbler on Alamagan to update the 10-year old population estimate and information pertaining to habitat use. We also took the opportunity to conduct surveys for Aguiguan and Pagan Reed-warblers. We used these survey data to determine the current archipelago-wide status and distribution of reed-warblers in the Mariana Islands, compare the results to past survey estimates, and compute the survey effort needed to confirm extinctions on unoccupied islands. This fundamental population information is relevant to the conservation of reed-warblers throughout the Mariana Islands. Our results can be used to determine habitat restoration needs, identify which islands may be most suitable for future conservation introductions, and determine the most important threats to address.
Methods
Surveys
Surveys were conducted in 2008 and 2010 for reed-warblers on Aguiguan, Alamagan, and Pagan where they occurred in the past 40 years (Figure 1; see Appendix S2 for location and description of the Mariana Islands). Island-wide point-transect distance sampling was conducted at 80 stations on five transects on Aguiguan from 25–27 June 2008 (Amidon et al. Reference Amidon, Camp, Marshall, Pratt, Williams, Radley and Cruz2014), at 72 stations along 10 transects on Alamagan from 17–24 May 2010, and at 144 stations along 13 transects on Pagan from 1925 June 2010. Call playback surveys were additionally conducted at all stations on Aguiguan on 20–22 August 2008, and at 10 stations on Lower Lake and five stations on Upper Lake on Pagan from 20–21 June 2010. In 2007, island-wide point-transect distance sampling counts were conducted on Saipan (Camp et al. Reference Camp, Pratt, Marshall, Amidon and Williams2009), and in 2008 on Tinian (Camp et al. Reference Camp, Amidon, Marshall and Pratt2012). No surveys were conducted on Guam. The point-transect, directed counts and occupancy data for Alamagan are available from the U.S. Geological Survey <https://doi.org/10.5066/P94Y0LIJ> (Camp Reference Camp2020).
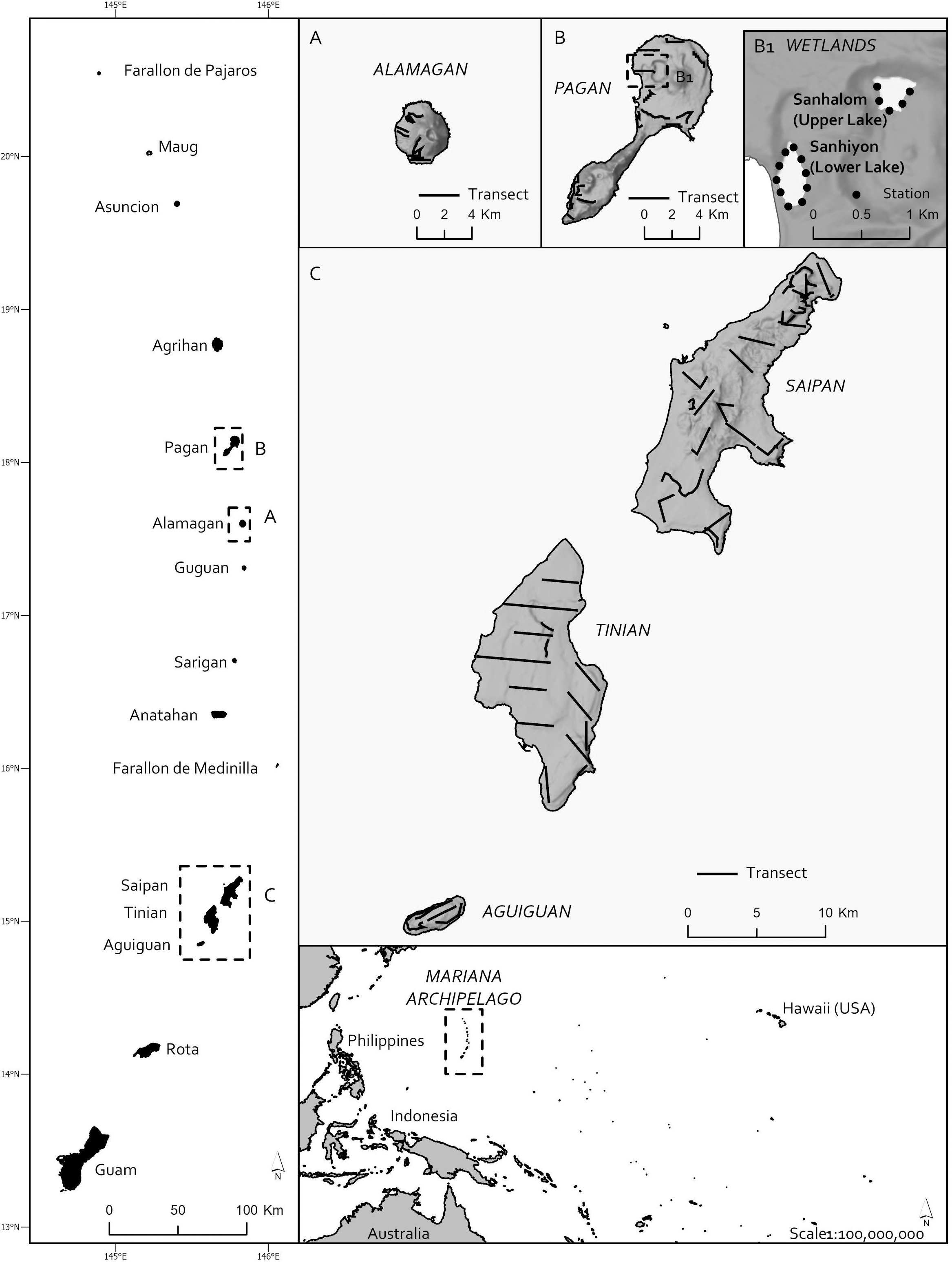
Figure 1. Maps of the Mariana archipelago and reed-warbler survey areas on Pagan, Alamagan, Saipan, Tinian, and Aguiguan, Commonwealth of the Northern Mariana Islands.
Point-Transect Surveys:
Prior to conducting surveys observers participated in at least three days of calibration involving distance estimation and bird vocalisation training in different habitat cover types. Point-transect distance sampling surveys were conducted for forest birds following standard point-transect methods, consisting of 8-minute counts at each station (150 m apart) and estimating and recording the horizontal distances to each bird heard or seen (see Engbring et al. Reference Engbring, Ramsey and Wildman1986 for additional details). Sampling conditions and habitat type were recorded at each station and were available for use as model covariates. Rangefinders were available for estimating distances to birds for surveys on Alamagan and Pagan. Surveys on Aguiguan occurred along transects established by Engbring et al. (Reference Engbring, Ramsey and Wildman1986) with additional transects added to improve survey coverage (Amidon et al. Reference Amidon, Camp, Marshall, Pratt, Williams, Radley and Cruz2014). Surveys on Alamagan and Pagan were conducted along transects established in forested areas on each island. The starting point for each transect was determined by selecting random points on a 150-m grid using a Geographic Information System (GIS). The remaining stations on each transect were placed at 150-m intervals to maximize coverage of forested areas. Counts commenced at sunrise, continued for 4 hours, and were conducted only under favourable weather conditions (halting when conditions exceeded light rain and Beaufort scale 3 winds). The habitat at each station was classified into one of the following cover types: native forest, coconut forest, secondary forest (following the Forest Service classification as mixed introduced forest, Casuarina equisetifolia thicket, agroforest, and agroforest-coconut; Liu and Fischer Reference Liu and Fischer2006), residential, open, or tangantangan Leucaena leucocephala thicket.
Population density by habitat cover type was calculated as birds per km2, and abundance (i.e. total number of birds) was obtained by multiplying densities by habitat cover type area. Density was calculated using the program Distance, version 6.0, release 2 (Thomas et al. Reference Thomas, Buckland, Rexstad, Laake, Strindberg, Hedley, Bishop, Marques and Burnham2010), from a global detection function where data were post-stratified by survey and cover type. Stations were usually sampled only once during an annual survey; but when sampled more than once, survey effort was adjusted by the number of times the station was counted. Data were right-truncated where detection probability was about 0.1 to facilitate model fitting (Buckland et al. Reference Buckland, Anderson, Burnham, Laake, Borchers and Thomas2001). Candidate models included half-normal and hazard-rate detection functions with expansion series of order two (Buckland et al. Reference Buckland, Anderson, Burnham, Laake, Borchers and Thomas2001 pp 361, 365). Sampling covariates were modelled in the multiple covariate distance engine of the Distance program (Thomas et al. Reference Thomas, Buckland, Rexstad, Laake, Strindberg, Hedley, Bishop, Marques and Burnham2010). The model with the lowest sample size corrected Akaike Information Criterion (AICc) was used to select the detection function that best approximated the data (Burnham and Anderson Reference Burnham and Anderson2002). Covariates were used to generate the global detection function when the best approximating model was improved by four or more AICc units. Model diagnostics were assessed and indicated adequate fit (Appendix S3). Variances and confidence intervals were derived by bootstrapping methods with 999 iterations (Thomas et al. Reference Thomas, Buckland, Rexstad, Laake, Strindberg, Hedley, Bishop, Marques and Burnham2010).
Directed Playback Surveys:
Directed playback surveys consisted of broadcasting digitally recorded Nightingale Reed-warbler calls from Saipan. The call was played on an electronic game caller (Foxpro NX3TM). On Aguiguan, the same transects used for the point-transect surveys were used for the directed playback surveys. The vocalisation of a singing male reed-warbler was broadcast for one minute, followed by three minutes of silent observation. All reed-warblers responding were recorded, including their distance, direction and time of detection (playback survey detections were analysed separately from the point-transect detections). Observers used rangefinders to assist with distance estimation and a compass for determining direction of the bird (based on compass bearing). Weather and habitat sampling conditions were likewise recorded at each station. Counts commenced at sunrise and continued until 11h00 and were conducted only under favourable weather conditions. Survey effort was recorded in hours as the difference between the start and end times for each survey.
Observers established stations and conducted directed playback surveys for reed-warblers at Laguna Sanhiyon on Pagan (Figure 1). These directed surveys were conducted in concurrence with surveys for the Mariana Common Moorhen Gallinula chloropus guami. Once counts for the moorhen at stations 1, 2 and 3 were done, another observer began the reed-warbler surveys at station 1. The observer listened for reed-warblers during the 3-minute survey period (30 seconds of playback followed by 2 minutes and 30 seconds of observation) and recorded the horizontal distances of any reed-warblers heard/and or seen, direction of the bird from the observer, and time of detection. The observer then walked 150 m along the lake to the next reed-warbler station, conducted the next playback and continued around the rest of the lake to the east side to finish at station 10. Reed-warbler counts at Laguna Sanhiyon started at 07h05 and ended at 08h19, after which observers walked to Laguna Sanhalom (Figure 1) and followed the same protocol establishing stations starting on the west side of the lake and ending on the east side. Reed-warbler counts at Laguna Sanhalom started at 11h00 (station 1) and ended at 11h48 (station 5). The next day, observers started surveys at Laguna Sanhalom following the same protocol as the first day, starting again on the west side of the lake at 08h06 and ending on the east at 09h15. Observers then walked to Laguna Sanhiyon where reed-warbler surveys were conducted from 11h46 to 12h56, starting at station 6 to 10 and ending with stations 1 to 5.
Following Reynolds and Snetsinger (Reference Reynolds and Snetsinger2001), we calculated detection probabilities to estimate the likelihood of extinction (extirpation) of reed-warblers on Aguiguan and Pagan. Scott et al. (Reference Scott, Mountainspring, Ramsey and Kepler1986) calculated the probability (p) of detecting one bird from a randomly distributed population of n individuals as:
 $$ p=1-{\left(1-\frac{a}{A}\right)}^n $$
$$ p=1-{\left(1-\frac{a}{A}\right)}^n $$
The effective search area (a) for reed-warbler on Aguiguan was approximated as 125.9 hectares (ha) by calculating the area for the effective detection radius (EDR) (71 m; Camp et al. Reference Camp, Pratt, Marshall, Amidon and Williams2009) using ArcMap 9.2 (ESRI, Redlands, CA) and XTools Pro 5.2.0 (Data East, LLC, Novosibirsk, Russia). The last known range of the Aguiguan Reed-warbler (A) was estimated as 435 ha based on the total available forest habitat (native, secondary and tangatangan) on the island (Amidon et al. Reference Amidon, Metevier and Miller2017). We set the hypothetical population size (n) at 15 as that was the last estimated number of birds on Aguiguan (Engbring et al. Reference Engbring, Ramsey and Wildman1986).
For Pagan, the effective search area for lake and forest, and lake-only was 34.4 ha and 1.8 ha, respectively, and approximated by calculating the area for the reed-warbler on Alamagan EDR (36 m; see Results). Detection probabilities were calculated for lake and forest, and lake-only searches. For lake and forest, the range of the Pagan Reed-warbler was estimated as 1,981.5 ha (lake habitat = 10.5 ha, forested habitat = 1,971.0 ha). For lake only, the range of the Pagan Reed-warbler was estimated as 10.5 ha. We set the hypothetical population size (n) at 20 as that was the last estimated number of birds on Pagan (Reichel et al. Reference Reichel, Wiles and Glass1992).
Using Reed’s (Reference Reed1996) modification of Guyann et al.’s (Reference Guyann, Downing and Askew1985) statistical methods to infer extinction, we also calculated the minimum number of visits, Nmin needed for 95% (α = 0.05) and 99% (α = 0.01) probability of detection as:
 $$ {N}_{min}=\frac{ln\alpha}{\mathit{\ln}\left(1-p\right)} $$
$$ {N}_{min}=\frac{ln\alpha}{\mathit{\ln}\left(1-p\right)} $$
where n is the number of independent visits. We defined one visit as 10 hours of search effort.
Occupancy surveys:
Surveys to determine reed-warbler occupancy were conducted on Alamagan in concurrence with the point-transect survey described above. All stations were sampled four times with sampling distributed across two time periods (06h00–11h00 and 14h00–18h00). All surveys were conducted by one observer and on Pagan followed playback methods described above (i.e. 3-minute survey period). Occupancy models included the following site covariates for occupancy probability (ψ) and occurrence probability (p): vegetation type (ψ), understorey closure/openness (ψ), canopy height (ψ), canopy cover (ψ), and time of day (morning or afternoon; p) (see Appendix S4 for a list of models with covariates). Vegetation type categories are coconut forest (CF), secondary forest (SF), and native forest (NF). Understorey openness is ranked from 1 (closed/dense) to 5 (open/sparse). Canopy height is ranked from 1 (short; < 3 m), 2 (mid; 3 – 10 m) and 3 (tall; > 10 m). Canopy cover categories ranged from very scattered (< 5% cover), scattered (5–25% cover), open (25–60% cover), and closed (> 60% cover). Occupancy models were first ranked according to AICc. Model goodness-of-fit statistics (ĉ) were assessed with a parametric bootstrap procedure (Mackenzie and Bailey 2004) in which a Pearson chi-square test statistic with a P-value ≥ 0.05 indicated an adequate model fit.
Results
Point-transect and directed surveys
We detected 60 Nightingale Reed-warblers from 72 stations during the 2010 point-transect survey on Alamagan. Because insufficient numbers of detections were made to reliably model a detection function (< 100 detections; Buckland et al. Reference Buckland, Anderson, Burnham, Laake, Borchers and Thomas2001), we added an additional 40 detections from the Division of Fish and Wildlife surveys of Alamagan (DFW 2000), and 46 detections from the 2007 Saipan surveys described in Camp et al. (Reference Camp, Pratt, Marshall, Amidon and Williams2009). These additional data were collected using the same survey methods, in similar sampling conditions and habitat cover types, and roughly during the same time of year. The count length of the DFW surveys was only 5 minutes; therefore, we included detections from the first 5 minutes of the 2010 survey (53), yielding a total of 139 detections to model the detection function. Truncation was set at 55.3 m, leaving 124 detections for population estimation. The hazard-rate detection function was the best-fit model, and the covariate “survey” further improved the model (Appendix S3). The effective detection radius was estimated to be 36 m. Although not statistically significant because the confidence intervals overlap, the Alamagan reed-warbler point estimates decreased from 1,125 to 946 individuals between 2000 and 2010 (Table 1).
Table 1. Population size estimates of Nightingale Reed-warbler on Alamagan. Density (birds ha-1) and abundance (density x area) with variance estimates were calculated for the 2000 Commonwealth of the Northern Mariana Islands Division of Fish and Wildlife and 2010 U. S. Fish and Wildlife Service surveys. Area for both surveys equalled 485 ha.

There were no reed-warbler detections on Aguiguan or Pagan during either point-transect or directed playback counts. During the directed surveys on Aguiguan, one vocalisation was recorded by an observer who thought they might have detected an Aguiguan Reed-warbler. This vocalisation was later definitively identified as that of a Golden White-eye Cleptornis marchei, whose warbling call can sometimes be mistaken for song of the male Nightingale Reed-warbler (APM pers. obs). Search effort totalled 47 hours for the June and August 2008 surveys on Aguiguan. We calculated the likelihood of detecting one reed-warbler during the 2008 surveys on Aguiguan to be 99% if the actual population was 15, while the likelihood of detecting one reed-warbler was 46% if the population on Aguiguan was two (Appendix S5).
The total time we spent searching during the lake and forest surveys on Pagan was 42.3 hours and covered 34.4 ha, whereas 1.5 hours were spent during the lake-only surveys and covered 1.8 ha. The likelihood of detecting one Pagan Reed-warbler ranged from slightly better than 3% to almost 98%, depending on the survey and the starting population (Appendix S5). If a population of two individuals persisted around the lakes, an additional eight searches would be needed to achieve a 95% likelihood of detecting an individual. If two individuals occurred in suitable habitat anywhere on Pagan (around the lakes and in the forest) 86 additional searches would be needed to achieve a 95% detection probability. Therefore, we cannot definitively infer extirpation of the Pagan Reed-warbler based on the 2010 surveys.
Occupancy survey
The top-ranked models included the site covariates canopy height and vegetation type (Table 2, Appendix S4). Although the confidence intervals overlapped considerably because of small sample sizes among habitat strata, occupancy of the reed-warbler on Alamagan appeared to be tentatively correlated with vegetation type (Table 2). Mean estimates ranged from a low of 0.19 (95% CI = 0.09–0.35) in native forest (n = 58), up to 0.49 (95% CI = 0.17–0.83) in coconut forest (n = 10), and 0.69 (95% CI = 0.02–1.00) in secondary forest (n = 2). Reed-warbler occurrence also appeared associated with canopy height, with higher mean occupancy in taller vegetation.
Table 2. Summary of reed-warbler occupancy (ψ) estimates and 95% confidence limits (lower and upper) for existing combinations of site covariate values. Results are shown only for the top-ranked subset of models as indicated by shading in Appendix S4. Results are sorted by ascending occupancy estimate within each model. Habitat variable “Canopy Height” is ranked from 1 (short; < 3 m), 2 (mid; 3 – 10 m) and 3 (tall; > 10 m), and “Vegetation” categories are coconut forest (CF), secondary forest (SF), and native forest (NF).

Occupancy models identified the time of day during which surveys were conducted as having a moderate effect on reed-warbler detectability. Although confidence intervals overlapped, the model ψ (canopy height-vegetation) p (time) showed a slightly higher mean in the morning (P = 0.43; 95% CI = 0.21–0.68) than in the afternoon (P = 0.31; 95% CI = 0.14–0.56) (Table 3).
Table 3. Summary of reed-warbler detection probability (p) estimates and 95% confidence limits (lower and upper) for the sampling covariate describing the time of day during which a survey was conducted. Results are shown only for the top-ranked subset of models as indicated by shading in Appendix S4.

Discussion
The Nightingale Reed-warbler population on Alamagan has declined slightly to between 428 and 1,762 birds (95% CI) in 2010 since it was last surveyed in 2000 and estimated at between 504 and 1,539 birds (95% CI) (Table 1). This slight decline was not significant but is concerning, especially given a similar decline on Saipan (estimated at 1,686 to 3,956 birds in 2007 (Camp et al. Reference Camp, Pratt, Marshall, Amidon and Williams2009). The global population was estimated at between 1,019 and 6,356 birds (mean estimate 3,688) by combining the 2010 Alamagan and 2007 Saipan abundances. The Alamagan population is therefore ~25 % of the global population (946 of 3,688 birds). The current population has declined by more than 1,000 birds since the first quantitative surveys were conducted in 1982 (Engbring et al. Reference Engbring, Ramsey and Wildman1986), which equates to a 24% decline in 28 years. This decline is conservative as Engbring et al. did not survey Alamagan. Restricted to two islands, the species remains listed as ‘Critically Endangered’ by IUCN (Birdlife International 2016). Declines in reed-warblers in the Marianas are not unique as similar declines have occurred for most forest birds and across most of the islands in the archipelago (Esselstyn et al. Reference Esselstyn, Cruz, Williams and Hawley2003, Camp et al. Reference Camp, Pratt, Marshall, Amidon and Williams2009, Reference Camp, Amidon, Marshall and Pratt2012, Reference Camp, Brinck, Gorresen, Amidon, Radley, Berkowitz and Banko2015; Amidon et al. Reference Amidon, Camp, Marshall, Pratt, Williams, Radley and Cruz2014), and is a pattern common to many insular bird populations (Steadman Reference Steadman2006, Wood et al. Reference Wood, Alcover, Blackburn, Bover, Duncan, Hume, Louys, Meijer, Rando and Wilmshurst2017).
Our results indicate that the Aguiguan and Pagan Reed-warblers are likely extinct, which agrees with previous assessments of reed-warbler population on those islands (Reichel et al. Reference Reichel, Wiles and Glass1992). We found little evidence that a population of even a few individuals persists on Aguiguan. Because a relatively small number of surveys would be required to detect at least one bird at a 99% probability of detection, we believe that the Aguiguan Reed-warbler is extinct. In contrast, a relatively large number of surveys, between 80 and 860 hrs, is required to conclusively determine that the Pagan Reed-warbler is extinct. Regardless, we believe the species is likely extinct because it was historically associated with the wetlands on the island, which were heavily surveyed in 2010. Although we did not survey on Guam, Guam Reed-warblers have not been detected either during annual Christmas Bird Counts or on bird surveys conducted by the Guam Division of Aquatic and Wildlife Resource since the species was last sighted in 1969 (Reichel et al. Reference Reichel, Wiles and Glass1992), which further confirms it is likely extinct. Archaeological and paleontological investigations reveal that an Acrocephalus sp. reed-warbler occurred on Tinian prehistorically but is now extinct (Steadman Reference Steadman2006). Reed-warblers were not detected on Tinian during the 2008 survey (Camp et al. Reference Camp, Amidon, Marshall and Pratt2012) nor during a subsequent 2013 survey conducted for the Commonwealth of the Mariana Islands Joint Military Training (CJMT) Environmental Impact Statement (EIS) (Naval Facilities Engineering Command, Pacific 2014).
The Nightingale Reed-warbler population on Alamagan appears to have varied little in the last decade, but may likely show declines in the near future due to habitat loss and fragmentation as a result of continued grazing by feral ungulates (Kessler Reference Kessler, Veitch and Clout2002). Alamagan is not currently inhabited by humans and does not support agriculture or urban habitats. Forested habitat covers approximately 35% of the island (Amidon et al. Reference Amidon, Metevier and Miller2017) with the remaining area dominated by grassland or barren land-cover not suitable for reed-warblers. We found the greatest Nightingale Reed-warbler occupancy in secondary forest with lesser occupancy in native and coconut forests. Ungulates directly affect vegetation through browsing that suppress forest regeneration, as well as indirect effect attributable to changes in the forest floor microclimate leading to desiccation and subsequent loss of soil. This situation may result in the conversion of forest into non-forested land cover (Worthington et al. Reference Worthington, Marshall, Wiles and Kessler2001, Kessler Reference Kessler, Veitch and Clout2002). Based on our observations of habitat cover types during our surveys, the effects of ungulates are particularly obvious in the lower elevations of Alamagan with loss of understorey, the occurrence of large open areas, and forest fragmentation. Kessler (Reference Kessler, Veitch and Clout2002) showed that ungulate removal on the Mariana island of Sarigan resulted in rapid regeneration of forest cover and a reversal of forest loss. Without a concerted effort to control and eradicate ungulates on Alamagan, a quarter of the global Nightingale Reed-warbler population could be lost, which would have serious implications for the species’ long-term conservation. The Nightingale Reed-warbler, moreover, is declining on Saipan where the largest population of the species occurs (Camp et al. Reference Camp, Pratt, Marshall, Amidon and Williams2009). This is likely a result of invasive species and human development resulting in forest habitat conversion and degradation. Camp et al. (Reference Camp, Pratt, Marshall, Amidon and Williams2009) found that Nightingale Reed-warblers were least likely to occur in residential areas and golf courses, where their populations were substantially declining on Saipan. These areas have subsequently expanded with the increasing human population on the island.
Management and conservation efforts have not been successful in arresting declining abundances and curbing island extirpations of reed-warblers in the Mariana Islands. Threats to reed-warblers include the loss and degradation of habitat due to agriculture, urbanization, grazing by feral ungulates, depredation by introduced mammals, lack of a local conservation ethic (e.g. Sussman et al. Reference Sussman, Ha and Henry2015), lack of law enforcement (e.g. Sussman et al. Reference Sussman, Ha and Henry2015), and demographic constraints associated with small island land area (USFWS 1998, Graham et al. Reference Graham, Gruner, Lim and Gillespie2017). To meet the recovery goals specified by USFWS (1998) it is pertinent that the distribution and population numbers of reed-warblers increase. Ensuring that the Alamagan population remains stable, and preferably increases, may be achieved by establishment of fenced areas that protect the remaining forested areas for Nightingale Reed-warbler use. Identification of suitable areas may be achieved through the development and implementation of a forest restoration plan with preference to protection of tall-stature native and secondary forest, and ravines forested with pago Hibiscus tiliaceus. Arresting the decline of the Nightingale Reed-warbler on Saipan will be more difficult but could benefit from habitat protection and restoration, and predator removal.
Development of island-wide conservation plans identifying critical areas for reed-warblers to persist on both Alamagan and Saipan would benefit the long-term survival of the species. Prudent conservation planning could include developing and implementing a translocation plan to re-establish reed-warbler populations on the previously occupied islands of Aguiguan, Tinian, and Pagan. Reviewing and evaluating the Mariana Avifauna Conservation (MAC) Working Group (2013), specifically the conservation introduction plan, could benefit a variety of native bird species and help managers better understand and target re-establishment goals. Habitat restoration along with predator control and feral ungulate removal following methods developed by Kessler (Reference Kessler, Veitch and Clout2002) would benefit translocation success. Potential translocations could benefit from ecological research into habitat use differences on Saipan versus Alamagan and conducting further genetic comparisons. Taxonomic classification using mitochondrial (mtDNA) sequencing was not definitive from a single specimen for which positive results were obtained and the relationship of Aguiguan Reed-warbler within the Micronesian clade of the Pacific Acrocephalus radiation has not been resolved (Saitoh et al. Reference Saitoh, Cibois, Kobayashi, Pasquet and Thibault2012). In addition to the management efforts necessary to meet reed-warbler recovery and delisting goals, continued monitoring with increased frequency of island-wide surveys (which include surveys for Nightingale Reed-warblers) on Alamagan and Saipan would facilitate assessing the effectiveness of management and conservation actions (Camp et al. Reference Camp, Pratt, Marshall, Amidon and Williams2009).
As with all avian species in the Mariana archipelago, another major concern to reed-warbler recovery is the potential for the introduction of the brown tree snake Boiga irregularis from Guam and its subsequent colonization of other islands in the CNMI. The brown tree snake has caused the extinction or extirpation of most of the land birds of Guam, probably including the Guam Reed-warbler (Savidge Reference Savidge1987, Reichel et al. Reference Reichel, Wiles and Glass1992, Rodda et al. Reference Rodda, Fritts and Chiszar1997, Wiles et al. Reference Wiles, Bart, Beck and Aguon2003, Rodda and Savidge Reference Rodda and Savidge2007). Interdiction and control of the brown tree snake at Guam ports helps to ensure that this highly invasive species does not spread. Brown tree snakes, however, have been found on both Tinian and Saipan (Rodda and Savidge Reference Rodda and Savidge2007). Saipan has a particularly long history of brown tree snake sightings, and while many have recently been discounted, there are many confirmed sightings (Brown Treesnake Technical Working Group [BTTWG] 2015). Recovery of the Nightingale Reed-warbler would be severely compromised if the brown tree snake were to become established on Saipan. Recovery of the species would be further hampered if the snake became established on Aguiguan and Pagan, the two proposed as target islands by the Mariana Avifauna Conservation Working Group (2013) for re-establishing reed-warbler populations via translocation.
Cibois et al. (Reference Cibois, Beadell, Graves, Pasquet, Slikas, Sonsthagen, Thibault and Fleischer2011) conclude that island-dwelling Acrocephalus are a good candidate species for conservation translocation programs. As a group, reed-warblers can colonize remote islands that have limited or diverse habitats and food resources, do particularly well on islands that are species-poor and presumably with low potential for inter-species competition, and can colonize islands multiple times (Cibois et al. Reference Cibois, Beadell, Graves, Pasquet, Slikas, Sonsthagen, Thibault and Fleischer2011). If the reed-warbler population on either Alamagan or Saipan were to crash, individuals from the other island could serve as a founder population without homogenizing the species’ genetics. The genetics of the reed-warbler on Tinian are unknown, and may have been either A. hiwae, A. nijoi, or an island-specific endemic species (Saitoh et al. Reference Saitoh, Cibois, Kobayashi, Pasquet and Thibault2012). Therefore, translocating birds to Tinian from either Alamagan or Saipan is genetically feasible once the habitat requirements are identified and restored.
Our results indicate that the Aguiguan and Pagan Reed-warblers are likely extinct and molecular techniques cannot be solely used to select candidate species for intra-island translocations. Saitoh et al. (Reference Saitoh, Cibois, Kobayashi, Pasquet and Thibault2012) also compared molecular and morphological characteristics and found that both the Guam and Aguiguan Reed-warblers were genetically similar to Nightingale Reed-warbler, and that the Guam and Nightingale Reed-warblers are morphologically indistinguishable. Consequently, individuals from Saipan could serve as a founder population for re-establishment of reed-warblers on Guam. The range of size and bill shape of the Aguiguan Reed-warbler are similar to those of the Nightingale Reed-warbler (Saitoh et al. Reference Saitoh, Cibois, Kobayashi, Pasquet and Thibault2012; based on measurement of three individuals). Reed-warbler on Saipan could therefore also serve as a founder population for Aguiguan.
Saitoh et al. (Reference Saitoh, Cibois, Kobayashi, Pasquet and Thibault2012) noted that the Pagan Reed-warbler was genetically distinct, and morphologically much smaller than individuals on the other Mariana islands. This morphological difference could be that A. yamashinae is more closely related to A. rehsei and A. mendanae from Nauru and the southern Marquesas, respectively (Cibois et al. Reference Cibois, Beadell, Graves, Pasquet, Slikas, Sonsthagen, Thibault and Fleischer2011) and potentially be a result of the Pagan Reed-warblers strong association with wetlands, whereas birds on other Mariana islands tend to be associated with more diverse terrestrial habitats. Larger reed-warblers with comparably larger bills tend to occur on islands with more diverse habitat cover and presumably consume a wider range of prey (Leisler and Schulze-Hagen Reference Leisler and Schulze-Hagen2011). Thus, Nightingale Reed-warblers translocated to Pagan may have a greater chance of establishing if they are smaller-sized birds like the extinct Pagan species and from Saipan where individuals are known to utilize wetland habitats. Alternatively, larger-sized birds from Alamagan could be translocated that have adapted to non-native and secondary native forest habitats, which may allow them to occupy the more diverse terrestrial habitats on Pagan. Regardless of the origin of the founder population, successful colonization of translocated Nightingale Reed-warblers to Pagan, Guam or Aguiguan may benefit if suitable protected habitat includes restoring native habitats, controlling ungulates, and eliminating predators. The remaining Nightingale Reed-warblers on Saipan and Alamagan could also benefit from an increase in their range and distribution across the archipelago.
Supplementary Material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0959270921000034.
Acknowledgements
This study was produced by the USFWS and U.S. Geological Survey (USGS)-Pacific Island Ecosystems Research Center (PIERC). The U.S. Navy and Pacific Islands Fish and Wildlife Office (PIFWO) provided funding for our work. Data were analysed by PIERC, PIFWO and the University of Hawaii at Hilo-Hawaii Cooperative Studies Unit. We thank the many field biologists who organized or collected the data, especially Major B. Adams, T. Castro, L. Corin, C. Eggleston, C. Eggleston, S. Hudson, S. Kremer, M. Laut, J. Lepson, G. Martin, T. Robert, and R. Rounds. We are also indebted to those in the Marianas who cut transects over the years; the Secretary of the CNMI DFW for assistance with permits; PIFWO managers and administration; C. Kessler and C. Clark provided logistical support; Americopters and Hansen Helicopters; and the Cabras Marine on Guam, captain, and crew of the MV Micronesian; the U.S. Navy for providing access to their leased land; the USGS for volcano monitoring; the U.S. Coast Guard for support; the Mayor and people of Tinian; and the Mayor and people of the Northern Islands for their support and hospitality. The findings and conclusions in this article are those of the author(s) and do not necessarily represent the views of the U.S. Fish and Wildlife Service. Any use of trade, product, or firm names in this publication is for descriptive purposes only and does not imply endorsement by the U.S. Government.