Introduction
Historical background
The Northern Bald Ibis (NBI), Geronticus eremita, is an ‘Endangered’ species (BirdLife International 2018). Until the beginning of the 17th century, it also bred in Central Europe (Schenker Reference Schenker1977, Böhm and Pegoraro Reference Böhm and Pegoraro2011). Many human activities have reduced NBI populations (Collar and Stuart Reference Collar and Stuart1985). During the early decades of the 20th century, birds were shot for the skin collections of European museums (Meade-Waldo Reference Meade-Waldo1903, Pallary Reference Pallary1922, Chabot Reference Chabot1923, Hartert and Jourdain Reference Hartert and Jourdain1923, Bédé Reference Bédé1926, Berlioz Reference Berlioz1926, Hartert Reference Hartert1928, Meinertzhagen Reference Meinertzhagen1940). Illegal hunting and poaching, including egg collecting and consumption, were widespread. Furthermore, eggs and juvenile and adult birds were collected for zoological gardens. For instance, the NBI colony in the Zoological Garden in Basel, Switzerland was built up with 38 birds imported between 1949 and 1954 from the Zoo Aïn-Sebaâ in Casablanca (Zoo Basel pers. comm. 2017). The Basel colony has been reproducing regularly since 1957.
Other important factors contributing to the decline of the NBI were the massive application of pesticides by aircraft to control desert locusts and use of DDT in anti-malaria campaigns (Hirsch Reference Hirsch1980) as well as changes in land use (for Morocco see below) (Collar and Stuart Reference Collar and Stuart1985).
Road construction, dam construction and drowning of breeding sites as well as alterations to feeding habitats and disturbance by tourist activities have further reduced breeding colonies since the 1960s (Brosset Reference Brosset1957, Reference Brosset1961, Ruthke Reference Ruthke1966, Hirsch Reference Hirsch1979, Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003). Only a few breeding sites were lost due to natural erosion processes (e.g. a collapse of a steep riverbank) (Brosset and Petter Reference Brosset and Petter1966). As a result, only two separate breeding populations remained in the wild by the end of the 20th century. A western population survived in Morocco and Algeria (Algerian population now probably extinct; Fellous Reference Fellous2004, Reference Fellous2011) and an eastern population in south-east Turkey and Syria (Collar and Stuart Reference Collar and Stuart1985, Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003, Thévenot et al. Reference Thévenot, Vernon and Bergier2003, Serra et al. Reference Serra, Peske, Abdallah, al Qaim and Kanani2009, Hatipoglu Reference Hatipoglu, Boehm and Bowden2016). The eastern population migrates along the Red Sea to the main wintering area in Ethiopia (Lindsell et al. Reference Lindsell, Serra, Peske, Abdullah, Al Qaim, Kanani and Wondafrash2009, Serra et al. Reference Serra, Lindsell, Peske, Fritz, Bowden, Bruschini, Welch, Tavares and Wondafrash2014). The western surviving population in Morocco is not migratory, but shows post-breeding dispersal mainly along the Atlantic coast and occasionally inland (Collar and Stuart Reference Collar and Stuart1985, Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003, Thévenot et al. Reference Thévenot, Vernon and Bergier2003, Reference Thévenot, Bergier and Qninba2015).
The massive decline of the eastern population, especially of the Birecik colony in south-east Turkey, is well documented (Kumerloeve Reference Kumerloeve1962, Hirsch Reference Hirsch1980, Akçakaya Reference Akçakaya1990, Pegoraro Reference Pegoraro1996). Today, only a semiwild colony remains near Birecik (Hatipoglu Reference Hatipoglu, Boehm and Bowden2016). A breeding colony discovered in 2002 in the Syrian Desert near Palmyra is most likely extinct by now (Serra et al. Reference Serra, Abdallah, Assaed, Abdallah, Al Qaim, Fayad and Williamson2004, Reference Serra, Peske, Abdallah, al Qaim and Kanani2009, Reference Serra, Lindsell, Peske, Fritz, Bowden, Bruschini, Welch, Tavares and Wondafrash2014, Serra Reference Serra2017).
Intensification of land-use
To illustrate land-use changes and their effects on the landscape of Morocco in greater detail, key elements of this development are briefly discussed.
Infrastructure:
The three most important road types in Morocco (i.e. national roads and highways, regional and provincial roads; Table 6) increased significantly during the last few decades, providing a much denser road network than in the past. Thus, the total length of asphalt roads in Morocco amounted to 11,500 km in 1954, 21,367 km in 1960, 28,000 km in 1992 and 42,000 km or 74% of the total road network of Morocco in 2015 (Abécassis Reference Abécassis2009, Ministère de l‘Equipement 2017).
In addition, a high number of dams were constructed for water use, further documenting the dynamic development of Morocco. From 1929 to 1969, 17 large dams were built, and from 1970 to 2009, 113 new dams were realised (Secrétariat d’Etat auprès du Ministère de l’Equipement 2017). These numerous dams have multiple functions, but irrigation of agricultural areas (63%) and reservoirs for drinking water (36%) are predominant.
Demographic growth:
Demographic growth and urbanisation increased simultaneously. The human population of Morocco amounted to 12.3 million in 1960, and 70.6% of the population was rural. In 1990, the population had increased to 24.8 million (51.6% rural) and in 2016 to 35.2 million (39.3% rural) (Banque Mondiale Reference Banque2017).
Agriculture:
According to the last general nationwide census carried out in Morocco in 1996 and 1997 (Akesbi Reference Akesbi2001), small farms (41%; less than 3 ha rain-fed and less than 1 ha irrigated land) and small to medium-sized farms (57%; 3–50 ha rain-fed and 1–20 ha irrigated land) prevailed. Only 2% of all farms were large (> 50 ha rain-fed and > 20 ha irrigated land, Useful Agriculture Surface (UAS), Akesbi Reference Akesbi2001). However, the proportion of cultivated land in Morocco used by small farms was only 8%, whereas small to medium-sized and large farms used 70% and 22% of the cultivated land, respectively. In general, the small to medium-sized farms are family farms (Akesbi Reference Akesbi2001). In 1996, large regional and local irrigation projects covered 585,000 ha and 325,000 ha, respectively (Ragala and Refass Reference Ragala and Refass2002). Since 2000, the irrigated area has been growing constantly. Following the national plan Maroc Vert (Green Morocco), drip irrigation has been especially promoted since 2008. The irrigated area increased from 100,000 ha in 2003 to 250,000 ha in 2010 and to 483,000 ha in 2016 (Ministère de l’Agriculture 2011, 2017).
In 2016, a total of 8.7 million ha was used as agricultural land in Morocco (UAS; Ministère de l’Agriculture 2017). Fifty-two percent of this area was used for growing cereals, 15% for plantations (citrus, olive trees and fruit trees), 20% was fallow land, 5% was used for growing fodder and 8% for legumes, market gardening and other small crops.
Actual situation
In Morocco, approximately 70 breeding colonies were originally known. In 1940, the NBI population in Morocco consisted of about 1,500 birds (Hirsch Reference Hirsch1976). However, the population declined to about 600 individuals in 1975 (Hirsch Reference Hirsch1976), including 198 breeding pairs dispersed over 13 breeding sites (Hirsch Reference Hirsch1979). Thus, only four colonies have survived on the Atlantic coast in south-west Morocco, three in the Souss-Massa National Park (SMNP) and one in Tamri (Table 2) (Bowden and Smith Reference Bowden and Smith1997, Thévenot et al. Reference Thévenot, Vernon and Bergier2003, Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003, Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008). Finally, the creation of the Souss-Massa National Park in 1991 and conservation measures implemented since 1993 resulted in an increase of the last wild NBIs from 59 breeding pairs in 1997 to 147 breeding pairs and a total of 708 individuals in 2018, including non-breeding and juvenile birds (El Bekkay et al. Reference El Bekkay, Oubrou, Ribi, Smith and Bowden2007, Oubrou and El Bekkay Reference Oubrou and El Bekkay2018). In 2017, two new small breeding colonies on the Atlantic coast north of Tamri were discovered (Aourir et al. Reference Aourir, Bousadik, El Bekkay, Oubrou, Znari and Qninba2017) and in 2018 two additional new sites were found in the SMNP on the Atlantic coast (Oubrou and El Bekkay Reference Oubrou and El Bekkay2018).
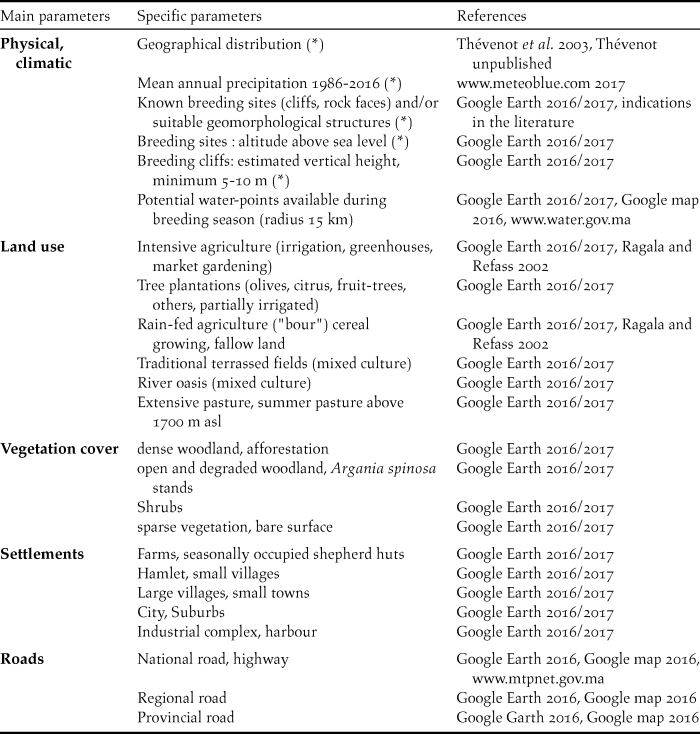
This paper investigates the abiotic conditions of the 72 Moroccan breeding colonies of the NBI that have been known since 1900 using qualitative and semi-quantitative parameters. In particular, geographical distribution, physical and climatic conditions such as the type of nesting cliffs, elevation (metres asl) of breeding sites and mean annual precipitation are considered. For the 28 known breeding colonies still in existence after 1977 (Appendix S1 in the online supplementary material), a more detailed habitat analysis within a perimeter of up to 5 km and > 5–15 km around the breeding sites was carried out, including land-use types as well as other landscape elements such as vegetation cover or urbanisation, to improve our knowledge of the ecological requirements of the NBI in Morocco. Hence the aim of this paper covers the following topics: 1) a better understanding of ecological requirements of the NBI in Morocco; 2) identification of possible key factors for the survival of the 28 breeding colonies still in existence after 1977; and 3) recommendations for a possible future reintroduction or translocation of NBI populations in the inland and/or at sea cliffs.
Methods
Details of the 72 known sites of breeding colonies between 1900 and 2016 are indicated in Thévenot et al. (Reference Thévenot, Vernon and Bergier2003), complemented by precise coordinates (Thévenot unpubl. data; Appendix S1). Due to different accuracy levels of the coordinates (Thévenot unpubl. data), locations of the breeding sites were characterised by the following three perimeter categories: perimeter 1: radius = 0–1 km, perimeter 2: radius > 1–5 km and perimeter 3: radius > 5–10 (-20) km.
Breeding sites within perimeter 1 could be identified with a high level of confidence. Sites within perimeter 2 could also be defined quite accurately. In some cases, precise indications from historical references were available. However, breeding sites of colonies within perimeter 3 are not known and indications of geomorphological structures as potential breeding sites are uncertain. For this reason, sites within perimeter 3 are dealt with separately.
Using satellite images from Google Earth (2016/2017), geomorphology, especially steep rock structures and their suitability as potential breeding sites, was assessed for each of the 72 known breeding sites. Terrestrial photos posted on Google Earth were used for characterising local conditions of breeding sites in cases where photos could be clearly attributed to the respective landscape.
Additional information on actual land use, vegetation and geomorphological conditions was gained by A. S. during a journey to Morocco along the south-western Atlantic coast, in the southern foothills of the High Atlas, the western Middle Atlas and the northern foothills of the High Atlas in April 2017. A second journey in April 2018 covered the Haouz and Oued N’fiss south-west of Marrakech, the Atlantic coast between Aglou plage and Tifnite and the Tamri area. Specifically, the following breeding sites and land-use patterns were checked on site in their wider surroundings (see Appendix S1 for numbers): in 2017 numbers 11–16, 34, 36–39, 44, 45 and 67–72 and in 2018 numbers 13–23 and 58. This information supplemented the analysis of breeding sites of the NBI.
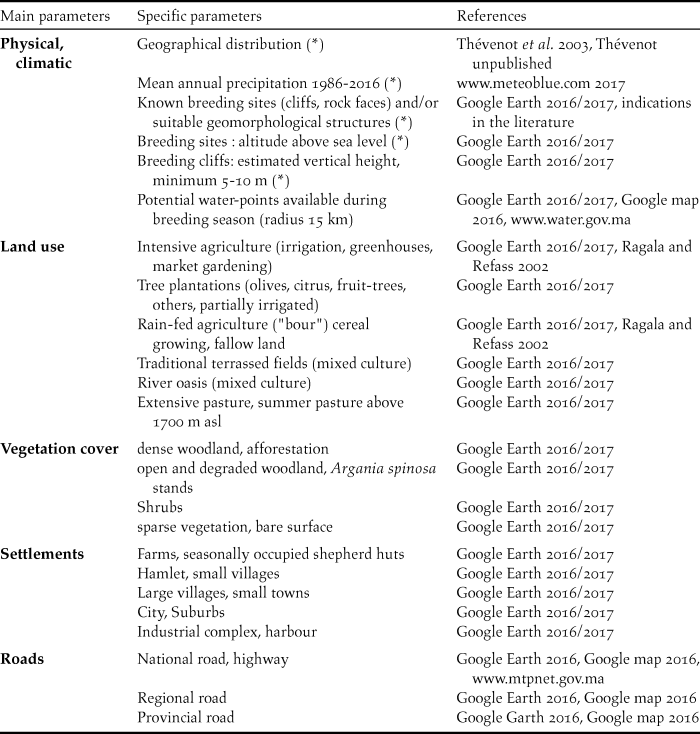
Breeding sites of the NBI were analysed separately on three levels: (A) 72 breeding sites known since 1900 (perimeter 1–2 and 3); (B) 28 breeding sites still occupied after 1977 (subset of A, perimeter 1–2) and (C) 4 breeding sites still occupied at present (subset of B, perimeter 1). For group (A), the geographical distribution, physical and climatic characteristics were examined, whereas for groups (B) and (C), a more detailed habitat analysis using Google Earth satellite images was carried out (Table 1).
Table 1. Parameters analysed for known breeding sites from 1900 to 2016 (*) and from 1977 to 2016, with corresponding references.
Data on mean annual precipitation for the years 1994–2016 for all 72 breeding sites were provided by meteoblue (meteoblue.com 2017). Meteoblue climate diagrams are based on 30 years of hourly weather model simulations. Simulated weather data have a spatial resolution of approximately 30 km and may not reproduce all local weather effects, such as thunderstorms, local winds, etc.
For the four remaining breeding colonies on the Atlantic coast, three in the SMNP near Oulad Noumer and one near Tamri, a possible correlation between quarterly precipitation (meteoblue.com 2017) and breeding success (number of fledglings) was tested covering the years 1994–2016. Furthermore, we tested if the number of reproducing pairs was correlated with the number of fledglings at the two locations.
Availability of fresh water in the vicinity of the breeding colonies has a positive effect on the breeding success of the NBI (Smith et al. Reference Smith, Aghnaj, El Bekkay, Oubrou, Ribi, Armesto and Bowden2008). For the 28 breeding colonies still in existence after 1977, potential available fresh water sources within a radius of 15 km around breeding sites were identified, using Google Earth satellite images. A clear distinction between permanent and temporary water bodies is difficult and varies according to the annual amount of precipitation, so the level of accuracy in the classification of water bodies is limited (Table 4).
Data analysis
We used R 3.3.1 and the R-package mgcv to perform a general additive model to analyse the number of fledglings. The model used the descriptive variables location, assessment year and precipitation in the fourth quarter of the previous year and the first and the second quarter of the same year during which fledglings hatched. Two specific smoothing curves were fitted for Tamri and Oulad Noumer for precipitation in the fourth and second quarter. The model was selected based on the Akaike information criterion (AIC) out of various models including different combinations of the precipitation in the fourth, first and second quarters. Visual inspection of the residual plots was applied to test for any obvious deviations from homoscedasticity or normality and temporal autocorrelation. Pearson Product-Moment Correlations were used to test for a correlation between the number of breeding pairs and fledglings.
Results
Sites of breeding colonies with their respective perimeters were evaluated using physical, climatic, habitat and landscape characteristics as well as landuse patterns. The corresponding findings are summarised in Tables 2–6 and Figure 1.
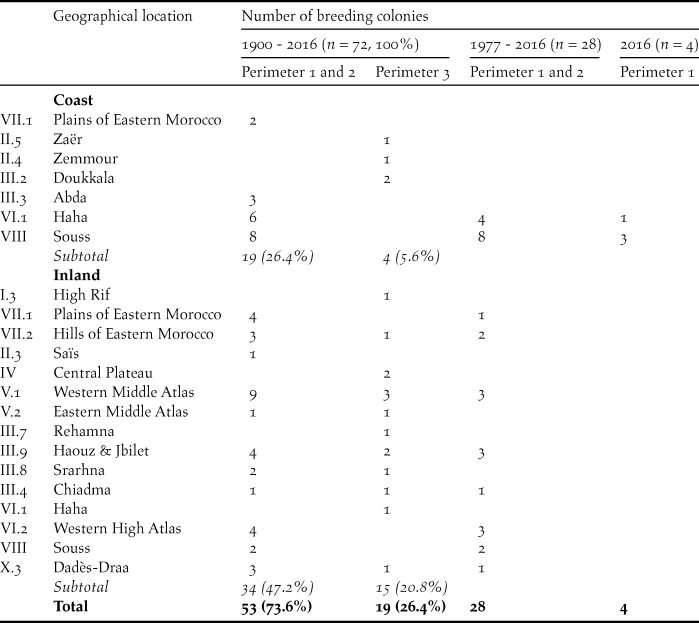
Table 2. Geographical location and accuracy of location (perimeter 1: 0–1 km, perimeter 2: > 1–5 km, perimeter 3: > 5–10 (20) km, see methods) of known breeding sites of the NBI in Morocco since 1900. Roman numerals of geographical locations according to Thévenot et al. (Reference Thévenot, Vernon and Bergier2003) and Thévenot (unpublished).
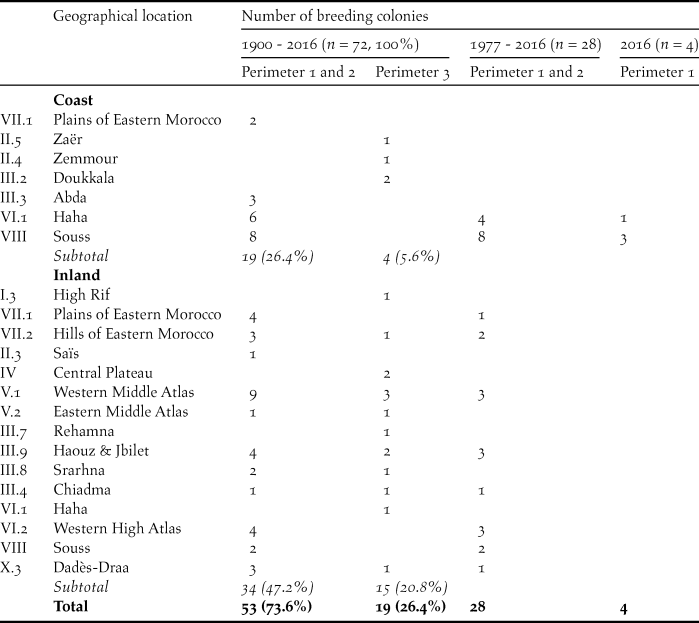
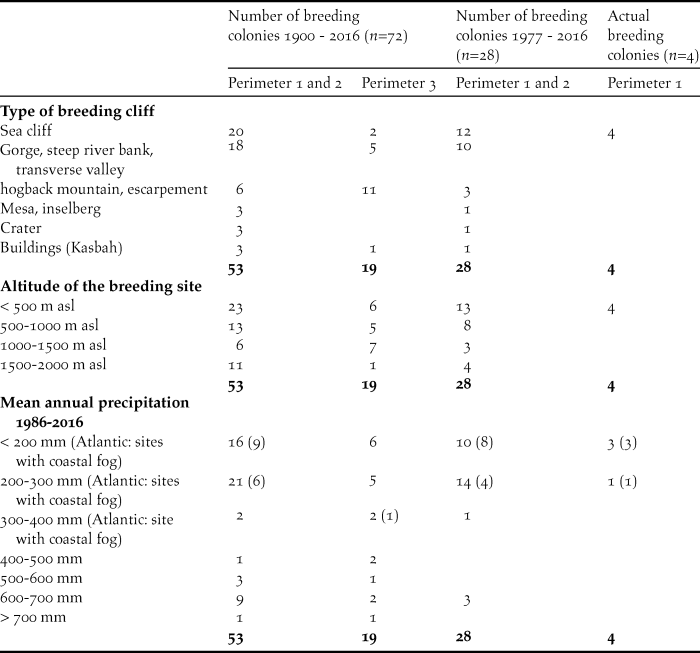
Table 3. Selected physical and climatic parameters of known breeding sites of the NBI in Morocco since 1900.
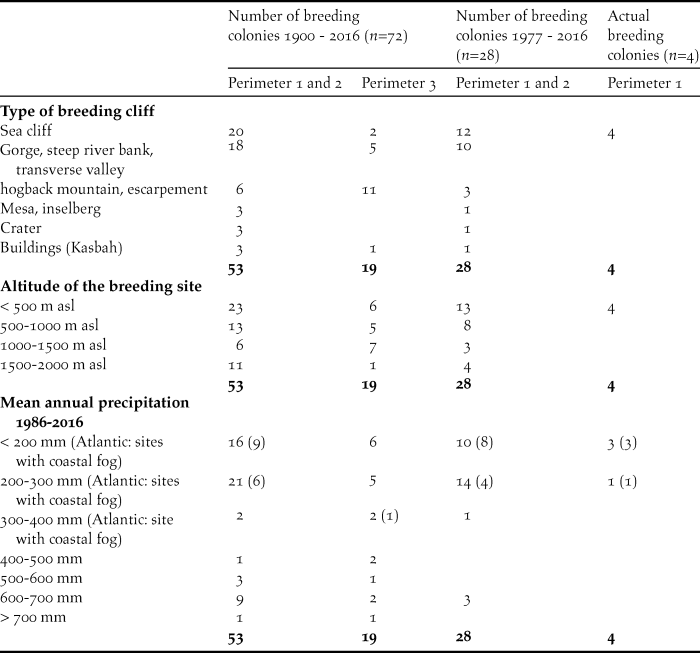
Table 4. Site characteristics of the 28 breeding colonies of the NBI remaining since 1977. Available water bodies during the breeding season. Multiple mentions are possible.

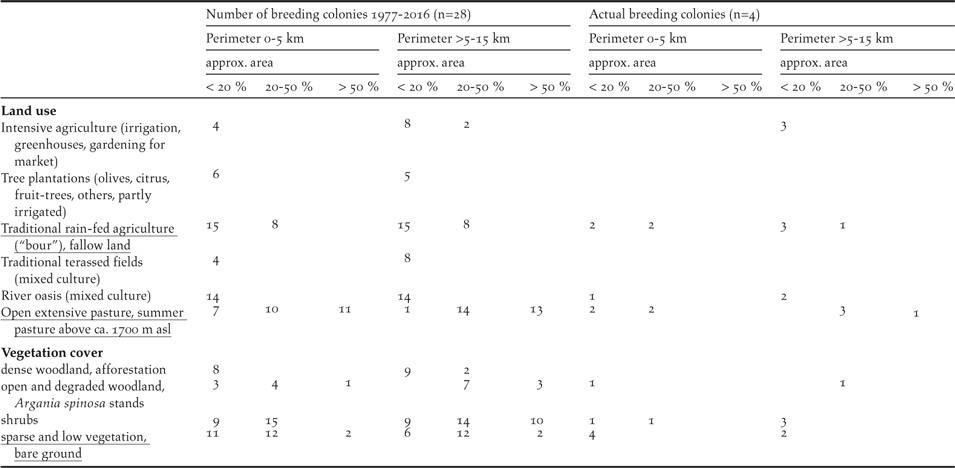
Table 5. Site characteristics of the 28 breeding colonies of the NBI remaining since 1977. Landuse and vegetation cover. Suitable feeding habitats are underlined. Multiple mentions are possible.
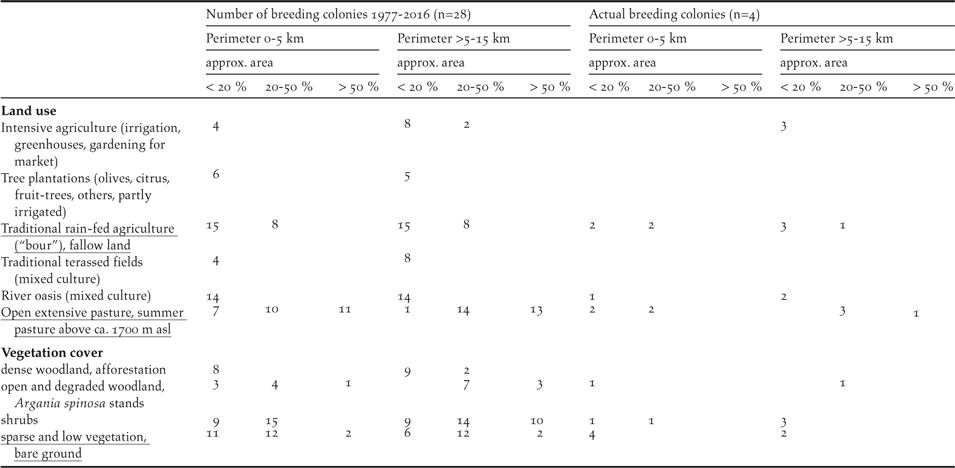
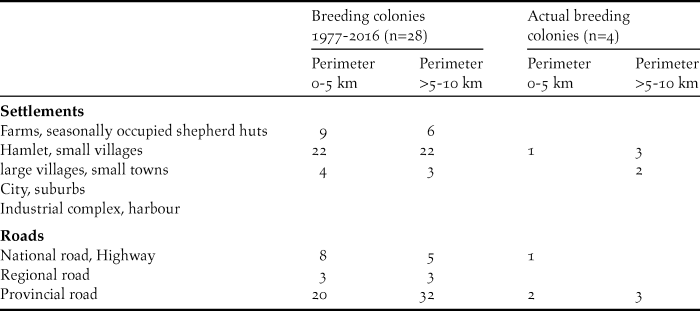
Table 6. Site characteristics of the 28 breeding colonies of the NBI remaining since 1977. Type of settlements and categories of roads. Multiple mentions are possible. Indications for perimeter 0–5 km are not included in perimeter >5–10 km.
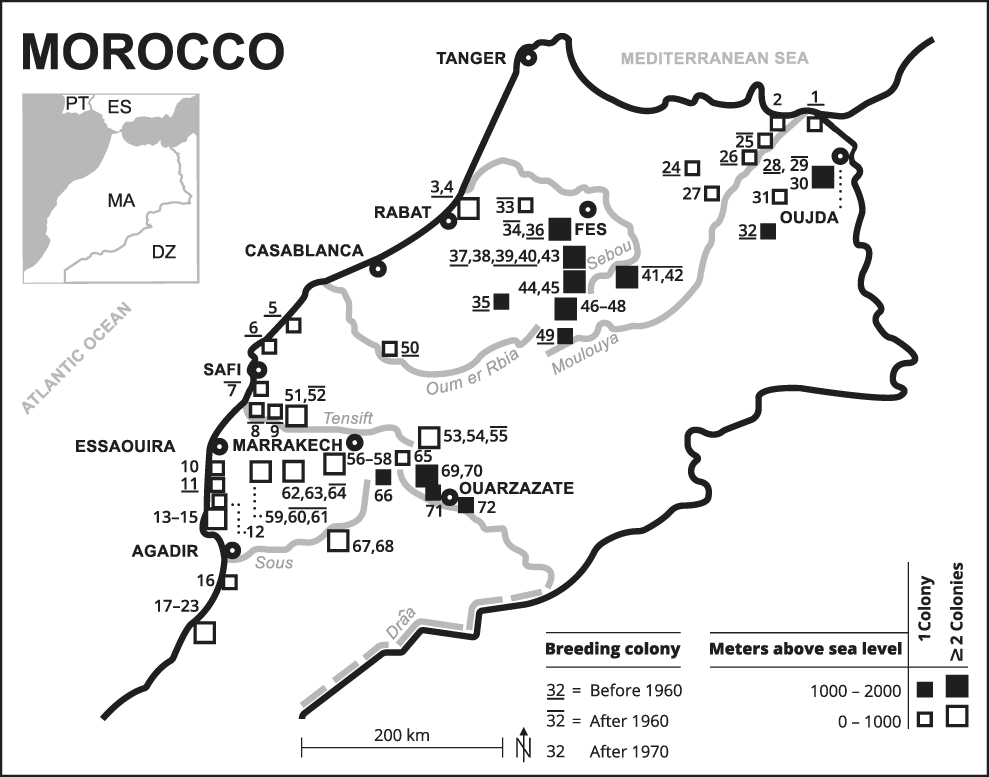
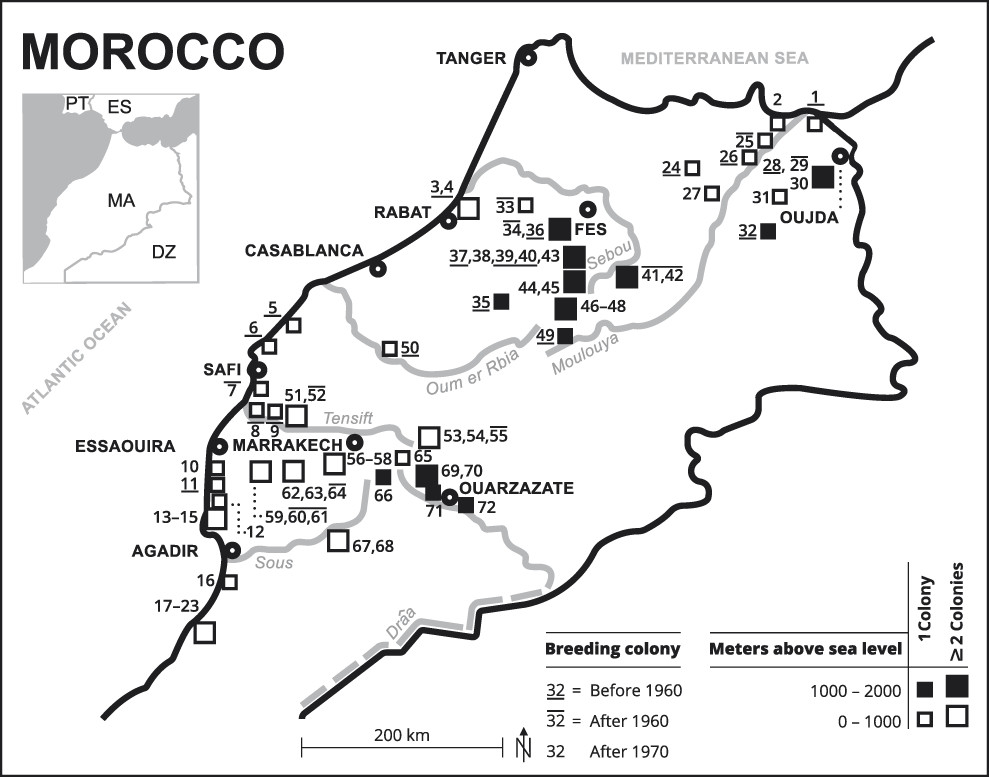
Figure 1. Breeding colonies of the NBI Geronticus eremita known since 1900 in Morocco (see Appendix S1 for numbers). The southern part of the Kingdom of Morocco without breeding sites is not shown.
Analysis of the 72 breeding sites (1900–2016)
Breeding colonies are dispersed over all of Morocco excluding the central Rif Mountains and the eastern steppe plateau (Table 2). However, a higher concentration of breeding colonies is found along the Atlantic coast, in the Souss-Massa region and in the Haha country (n = 14, 19%), in the Middle Atlas (n = 14, 19%), in the western High Atlas and the foothills of Haouz and Jbilet hills (n = 10, 14%) and in the plains and hills of Eastern Morocco (n = 8, 11%). The few known breeding colonies in the surroundings of cities (Rabat-Salé, El-Jadida, Safi, Meknes and El Hajeb) had disappeared already by the 1960s, whereas breeding colonies in more remote sites survived for longer (Figure 1).
The most frequent type of breeding sites (Table 3) are cliffs on the Atlantic and Mediterranean coasts, followed by steep slopes of perennial rivers and dry valleys, gorge-like structures and transverse valleys (perimeters 1 and 2: n = 38, 72%; perimeter 3: n = 7, 37%). However, escarpements and hogback mountains are additional important breeding sites (perimeters 1 and 2: n = 6, 8%; perimeter 3: n = 11, 15%). The most important breeding sites since 1977 are cliffs on the Alantic coast, steep riverbanks and gorge-like structures (perimeters 1 and 2: n = 22, 78%).
The elevation above sea level (asl) of breeding sites varies from sea level up to 2,000 m in the Middle Atlas. However, the majority of the breeding colonies occurred below 1,000 m (perimeters 1 and 2: n = 36, 68%; perimeter 3: n = 11, 58%) and most of the breeding colonies remaining since 1977 were also located below 1,000 m (perimeters 1 and 2: n = 21, 75%).
In the majority of the breeding sites known since 1900, mean annual precipitation from 1986–2016 did not exceed 300 mm (perimeters 1 and 2: n = 37, 70%; perimeter 3: n = 11, 58%). For most of the breeding sites still existing after 1977, annual precipitation was less than 300 mm (perimeters 1 and 2: n = 24, 85%). Twelve breeding sites on the south-west coast of the cold Atlantic Ocean benefit from frequent coastal fog and the formation of dew, which improves the ecophysiological conditions in this semiarid to arid coastal area.
Analysis of the 28 remaining breeding sites after 1977
The majority of the available water bodies (Table 4) in the surroundings of the breeding sites still in existence after 1977 are perennial rivers and temporary rivers with water that remains locally in the otherwise dry riverbed (n = 24, 69%). Small and large dam lakes and a large irrigation channel are also located within a perimeter of 15 km (n = 6, 17%). For eight breeding sites, only one temporary water body was available within a perimeter of 15 km, whereas for two SMNP breeding sites, not even a single water body existed within a perimeter of 15 km (data not shown).
Land use and vegetation cover within the two considered perimeters of the breeding sites often form mosaic-like patterns (Table 5). Of the six land-use categories and the four vegetation cover categories assessed using Google Earth, two and one, respectively, are potential feeding habitats for the NBI (see Table 5 underlines).
Within the perimeter of 0–5 km and a coverage of < 20% around the 28 breeding colonies still in existence after 1977, 33 (41%) of the 81 areas identified in the 10 categories belong to these three suitable feeding habitats. In the case of 20%–50% coverage, 30 (61%) of 49 areas are suitable, and in the case of > 50% coverage, 13 (93%) of 14 areas are suitable. For the perimeter > 5–15 km the corresponding figures are 22 (29%) of 75 areas, 34 (58%) of 59 areas and 15 (54%) of 28 areas.
Farms, seasonally occupied shepherd huts, hamlets and small villages are the most abundant type of settlements in the vicinity of the breeding sites (Table 6). Only a few breeding sites are located within the vicinity of large villages and small towns. The density of roads within the indicated perimeters of the breeding sites is high. However, smaller, provincial roads are the most frequent road type. Note that at four breeding colonies (three colonies in the SMNP, one at El Azib, see Appendix S1) none of the three road types is present within a perimeter 0–5 km (data not shown).
Rainfall and breeding success 1994–2016
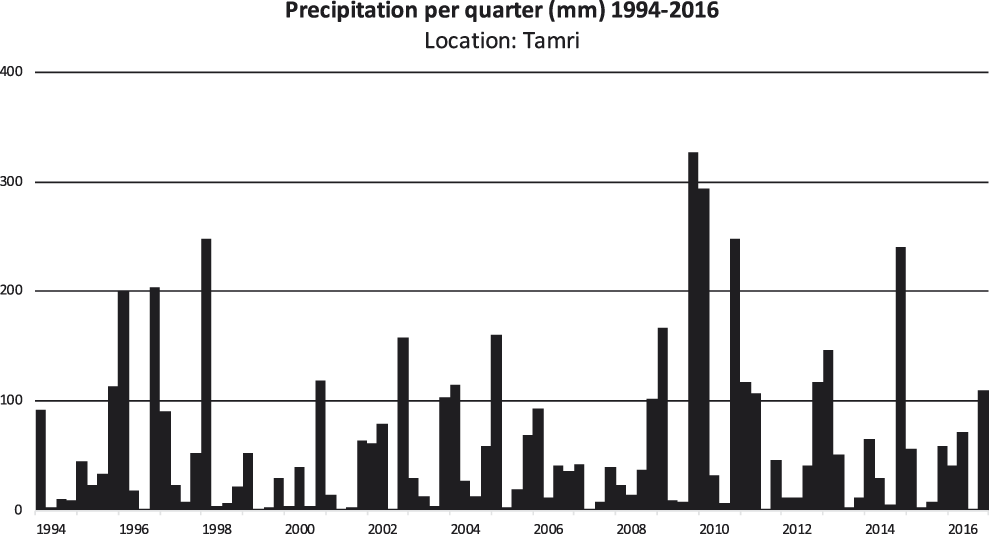
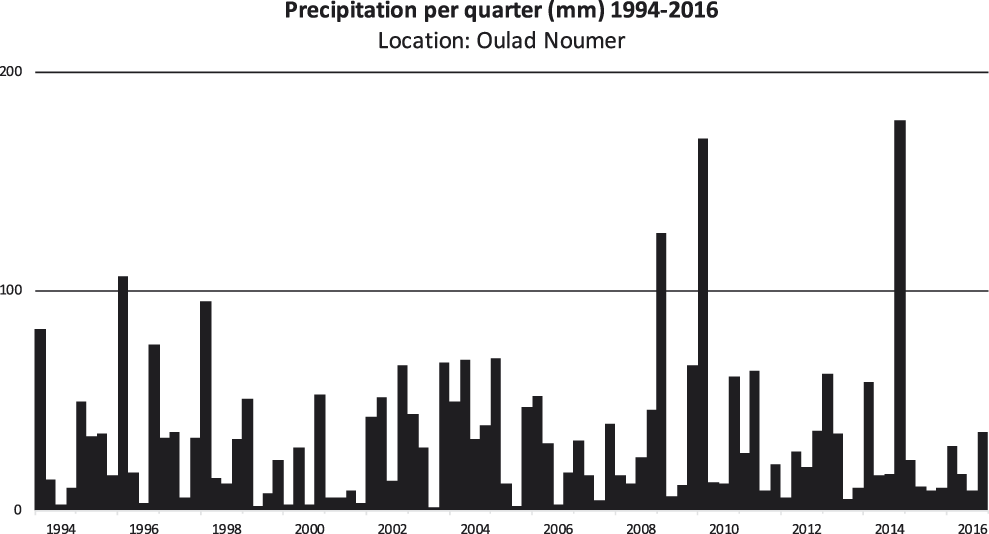
Quarterly precipitation for the period 1994–2016 is presented for two parts of the Atlantic coast that still have breeding colonies, Tamri (Figure 2) and Oulad Noumer, which are south of the estuary of the Massa River (Figure 3). At Tamri, the two driest years were 1999 (86 mm) and 2001 (82 mm). The two wettest years were 2009 (510 mm) and 2010 (580) mm. The mean annual precipitation from 1986–2016 was 243 mm. The two driest years at Oulad Noumer were 2001 (23 mm) and 2015 (52 mm) and the two wettest years were 2010 (255 mm) and 2014 (269 mm). The mean annual precipitation at Oulad Noumer from 1986–2016 was 140 mm.
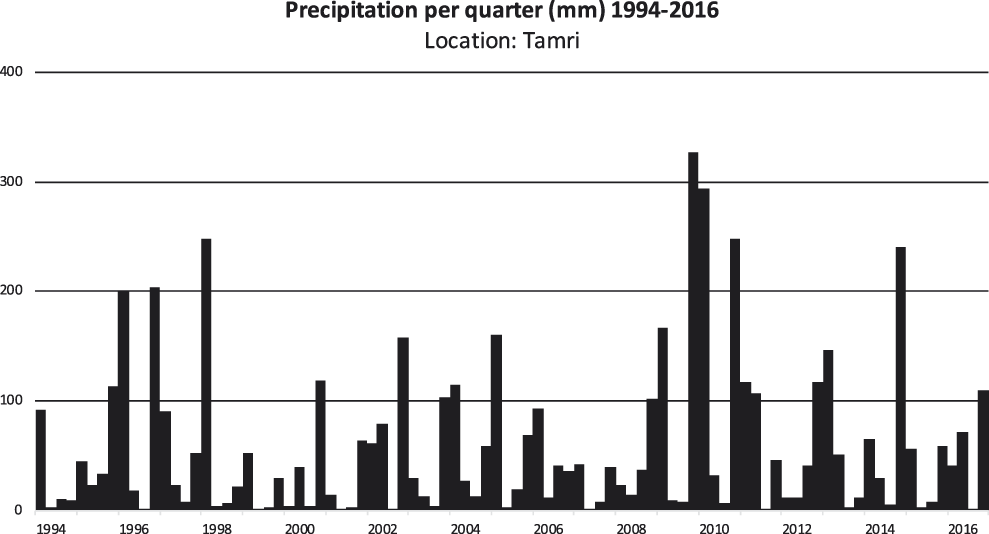
Figure 2. Quarterly precipitation for the period 1994–2016 at Tamri (30°41’ N, 9°49’ W). Precipitation data (meteoblue.com 2017). The spatial resolution of the weather model simulation is 30 km for the years 1994–2007 and 12 km for the years 2008–2016 (meteoblue.com 2017).

Figure 3. Quarterly precipitation for the period 1994–2016 at Oulad Noumer (29°53’ N, 9°42’ W). Precipitation data (meteoblue.com 2017). The spatial resolution of the weather model simulation is 30 km for the years 1994–2007 and 12 km for the years 2008–2016 (meteoblue.com 2017).
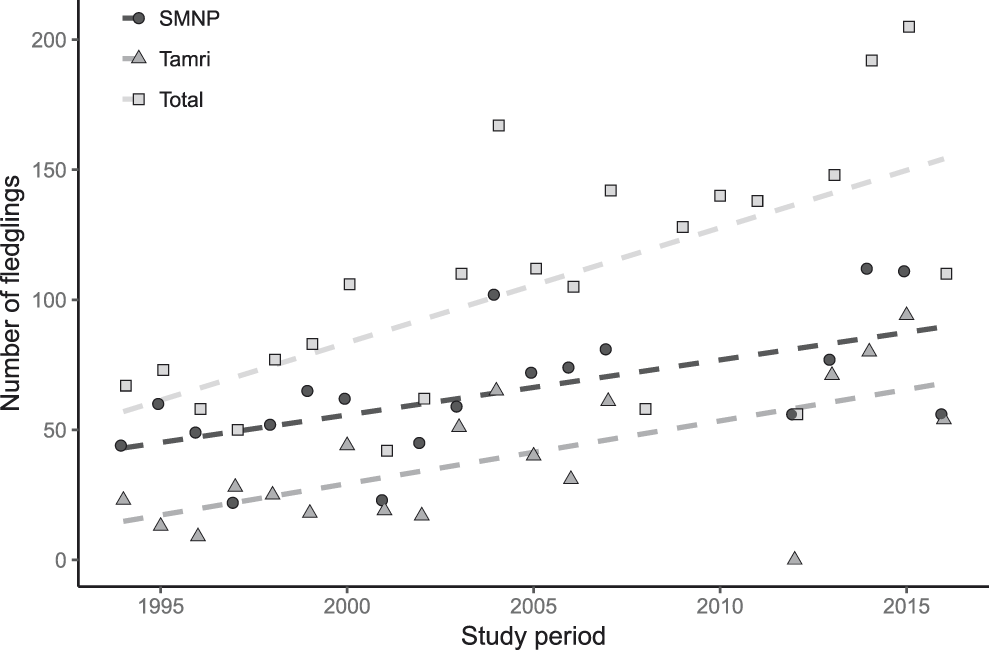
During the time period from 1994 to 2016 (Figure 4), the total number of fledglings varied from 50 (1997) to 205 (2015). Note the reduction by more than 50 fledglings from one year to the next in 2001 and 2005, and by about 100 fledglings in 2008, 2012 and 2016. On the other hand, a substantial increase by 75 fledglings occured within a single year in 2009 and another increase by over 100 fledglings within two years from 2002 to 2004 and 2012 to 2014.

Figure 4. Breeding success of NBI colonies from 1994–2016 in Souss-Massa National Park (SMNP) and Tamri (Bowden et al. Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008, Oubrou and El Bekkay Reference Oubrou and El Bekkay2013, Reference Oubrou and El Bekkay2014, Reference Oubrou and El Bekkay2015, Reference Oubrou and El Bekkay2016). For the years 2008–2011 the total was estimated according Oubrou and El Bekkay (Reference Oubrou and El Bekkay2014). Showing the trend from linear regression.
The number of breeding pairs (Oubrou and El Bekkay Reference Oubrou and El Bekkay2016) increased steadily from 65 pairs in 1994 to 116 in 2015, which is the maximum number of breeding pairs since 1994. However, two massive declines interrupted this increase, one in 1997 following a severe mortality incident in May 1996 with an unknown cause that reduced the number of breeding pairs by 18 (Touti et al. Reference Touti, Oumellouk, Bowden, Kirkwood and Smith1999), and one in 2012 with a reduction by more than 40 breeding pairs. The reason for this later decline is also not clear. In 2012, 43 breeding pairs were observed at Tamri, of which 11 pairs laid eggs. However, no hatchlings were observed in 2012 (Oubrou and El Bekkay Reference Oubrou and El Bekkay2013). The reason for this reproductive failure is also unclear.
Correlation between number of breeding pairs and fledglings
In Tamri, the number of fledglings was positively correlated with the number of reproducing breeding pairs (pairs whose females laid eggs; t = 8.66, R = 0.91, df = 16, P < 0.0001). However, this correlation was not found for SMNP (t = 0.71, R = 0.17, df = 16, P = 0.49)
Number of fledglings and precipitation
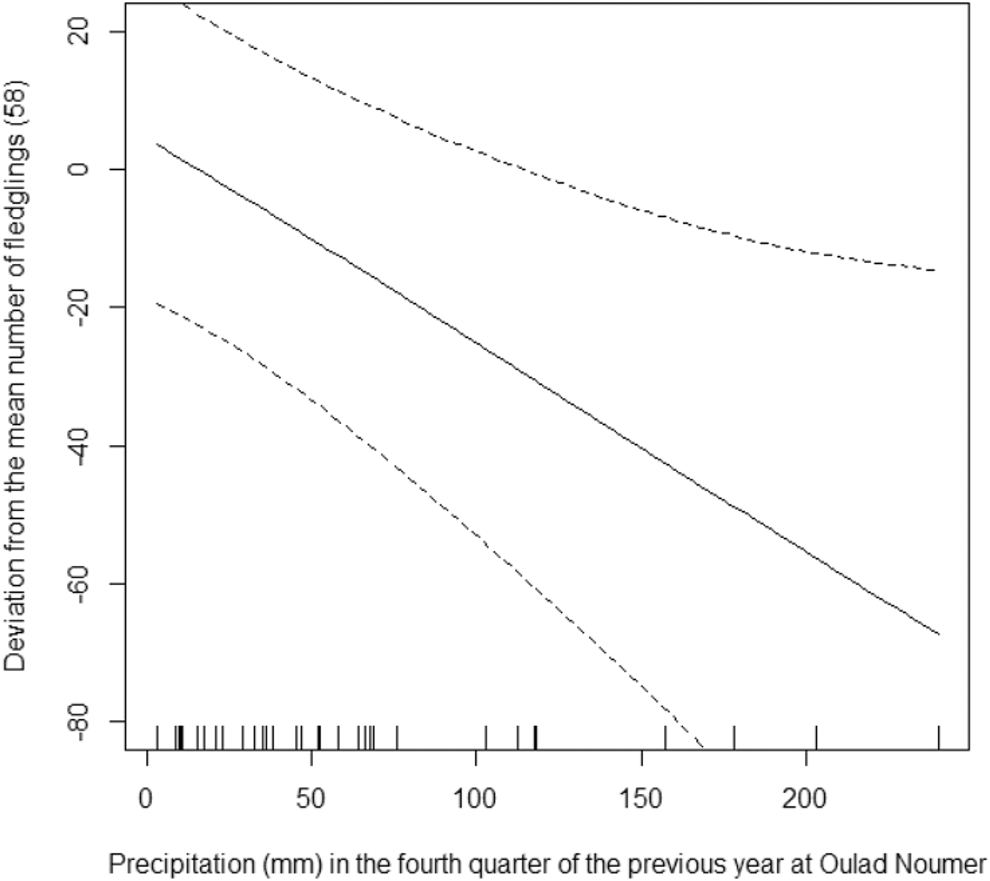
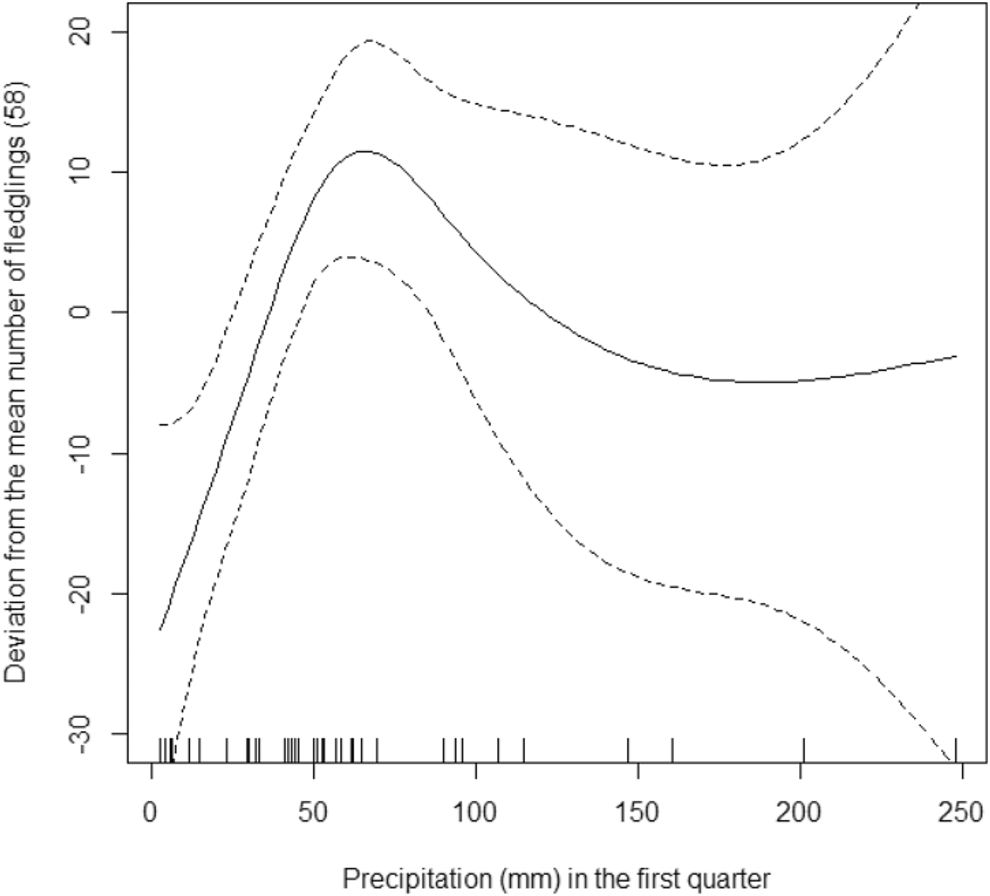
The model that we used explains 80.8% of the deviance in the data (R2 = 0.68). On average, there were 58 fledglings per year in Oulad Noumer and 19 fewer in Tamri, but this difference was not statistically significant (F1,36 = 0.00, P = 0.951). The number of fledglings increased significantly from 1995 to 2016 (F1,36 = 13.38, P = 0.001, Figure 4) by 11 fledglings per year. The number of fledglings in Tamri was not significantly related to precipitation in the fourth quarter of the previous year (F0.8,36 = 0.53, P = 0.073). In contrast, the number of fledglings in Oulad Noumer significantly decreased with increasing precipitation in the fourth quarter of the previous year (F1,36 = 1.89, P = 0.002, Figure 5). The number of fledglings was significantly related to precipitation in the first quarter of the year the fledglings hatched (F3.2,36 = 2.79, P = 0.009, Figure 6). The number of fledglings was not significantly related to precipitation in the second quarter in Oulad Noumer (F3.8,36 = 1.64, P = 0.057), but there was a relationship in Tamri (F3.2,36 = 2.18, P = 0.028), although there was not a clear trend and extremely large confidence intervals (figure not shown).
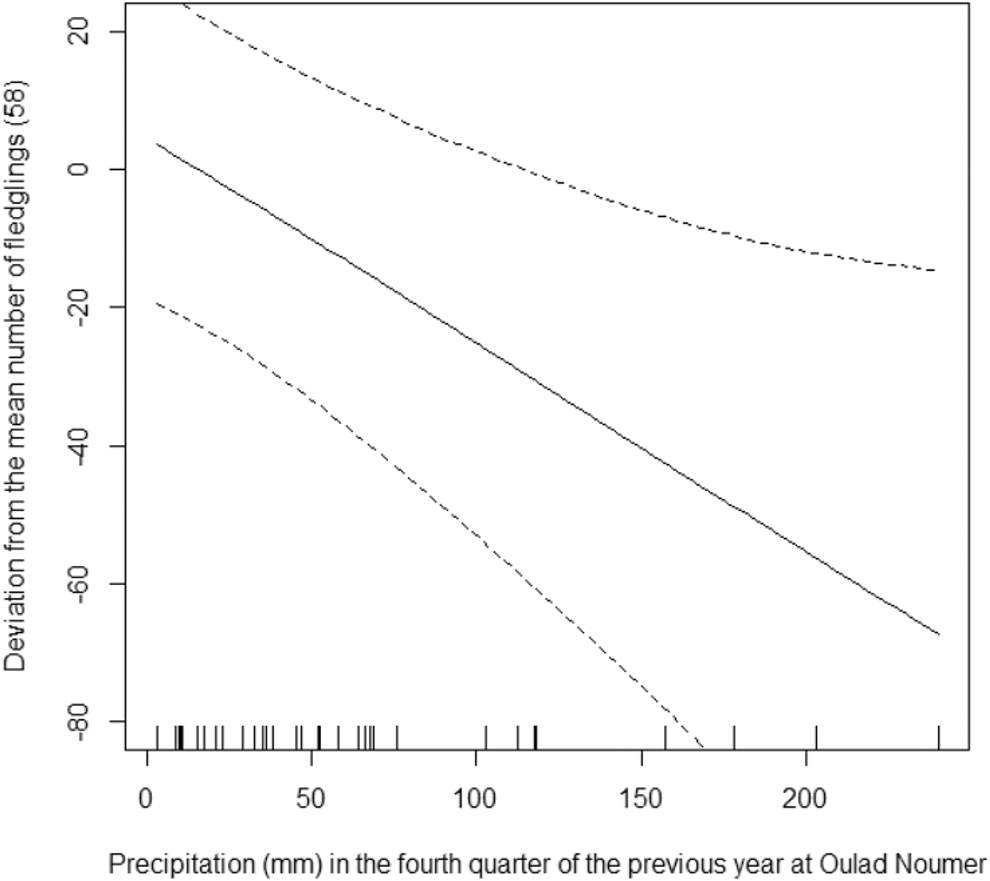
Figure 5. Effect of precipitation in the fourth quarter of the previous year on the number of fledglings (deviation from mean) at Oulad Noumer, with 95% confidence intervals.
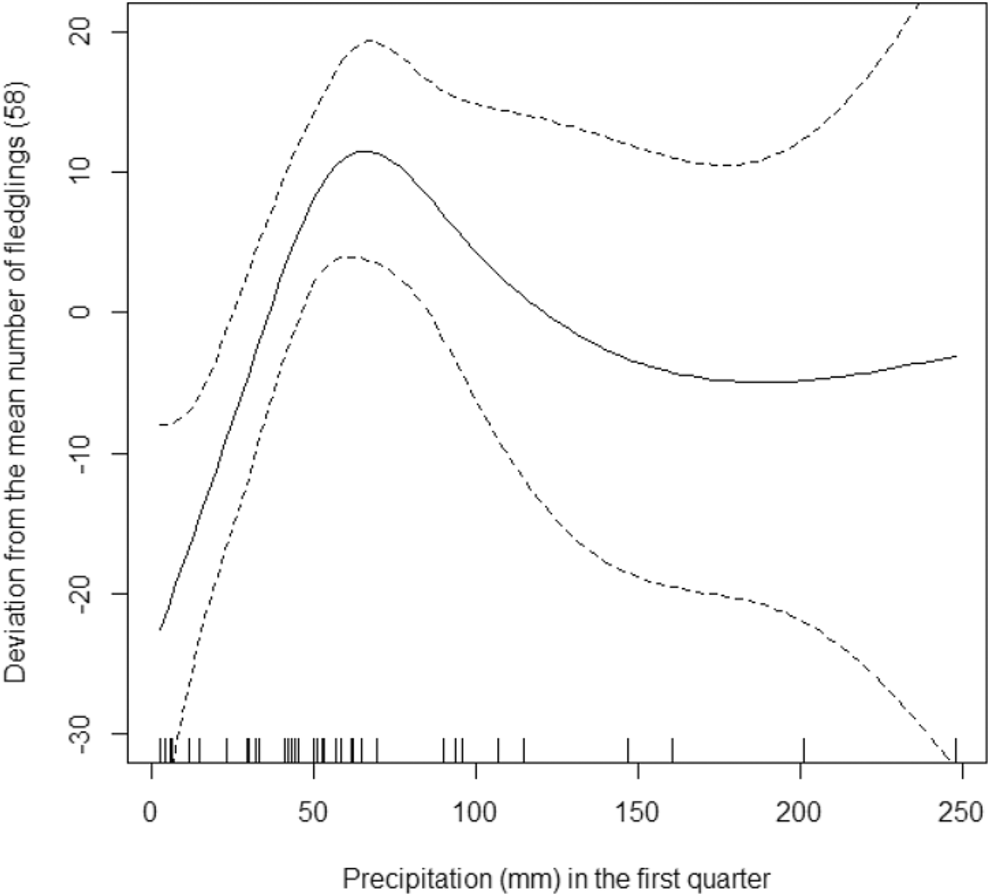
Figure 6. Effect of precipitation in the first quarter of the current year on the number of fledglings (deviation from mean) at Oulad Noumer and Tamri, with 95% confidence intervals.
Discussion
Known breeding sites of the NBI in Morocco and Algeria (Fellous Reference Fellous2004) that were occupied between 1900 and the 1960s were often located near important roads and in the vicinity of larger settlements where the probability of detection was generally higher. Following a marked decline during the 1970s, an intensive search in different regions of Morocco led to the discovery of a few breeding colonies that were previously unknown (Hirsch Reference Hirsch1976, Reference Hirsch1978). However, since the 1970s the population of Morocco, and hence urban settlements, has been increasing significantly and agriculture has intensified. This development and its associated growing disturbance correlates well and can be implicated in the further reduction of NBI breeding colonies, even in remote, less accessible, less disturbed and drier regions.
General conclusions
Geographical locations of Moroccan breeding colonies of the NBI share a common pattern: breeding colonies are often located at the transition zone of two distinct landscape types such as plains and foothills of the High Atlas, plains and isolated hills and plateaus and hilly area. Plains bordering hills and mountains are mostly used as agricultural land, whereas extensively grazed pastures and areas with sparse vegetation dominate in the foothills of mountains, isolated hills and plateaus. In the vicinity of breeding sites along the steep cliffs of the Atlantic coast, a small strip of coastal steppe is present.
Breeding colonies are located at the northern and southern fringe of the High Atlas, but not in valleys in the inner part of the Atlas Mountains. The indication of a breeding colony of the NBI in the Middle Atlas near Aït Kermousse south of Boulemane at 2,300 m asl (Ruthke Reference Ruthke1966) is probably not correct. Most likely, this breeding colony was located on an isolated hogback mountain (Google Earth 33°12’46” N, 4°39’35” W), 1,800 m asl and 5 km south of Aït Karmosse.
Steep rock faces with suitable ledges and small cavities are essential habitat structures for breeding colonies of the NBI. Such breeding sites are frequently present at cliffs along the coasts of the Atlantic Ocean and the Mediterranean Sea, and furthermore in areas with calcareous rock (Ragala and Refass Reference Ragala and Refass2002). In inland Morocco, steep rock faces and valleys with gorges and steep riverbanks are the preferred nesting sites. Water bodies in the vicinity of the breeding colonies obviously favour breeding success (Smith et al. Reference Smith, Aghnaj, El Bekkay, Oubrou, Ribi, Armesto and Bowden2008).
The elevations of the known Moroccan breeding colonies vary from sea level at the coasts of the Atlantic Ocean and the Mediterranean Sea to 2000 m asl in the Middle Atlas. Consequently, mean annual precipitation also varies from about 150 mm to more than 700 mm in the Middle Atlas. Following elevation and climatic conditions, egg deposition begins at the end of February and the beginning of March at the Atlantic coast (Bowden and Smith Reference Bowden and Smith1997, Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003), but is delayed until April in the formerly occupied Middle Atlas (Rencurel Reference Rencurel1974). High interannual variability of rainfall is the norm in semiarid and arid regions, which also affects food availability, particularly the availability of arthropods (Hunter et al. Reference Hunter, Walker and Elder2001, Meserve et al. Reference Meserve, Vasquez, Kelt, Gutierrez and Milstead2016, Palmer Reference Palmer2010). However, additional humidity brought in by frequent coastal fog and dew mitigates dry conditions on the Atlantic coast in southwest Morocco. In addition, mean annual precipitation can be affected by climatic change. Thus, the drainage basin of the river Souss shows a slight reduction in rainfall compared to earlier decades of the 20th century (Ait Brahim et al. Reference Ait Brahim, Seif-Ennasr, Malki, N’da, Choukr-Allah, El Morjani, Sifeddine, Abahous, Bouchaou and Choukr-Allah2017).
Furthermore, appropriate feeding grounds in the vicinity of the breeding sites are essential for the long-term survival of NBI colonies. Such feeding grounds are treeless, extensively grazed summer pastures in the Middle Atlas and traditional rain-fed agriculture and fallow land. The preferred feeding ground along the Atlantic coast is a narrow strip of sandy littoral steppe and fallow land. The three potentially suitable forage habitats of the NBI are present in a substantial proportion of the 28 breeding colonies still in existence after 1977 (see Results, Table 5).
Note that none of the three most frequented road types are present within a perimeter of 0–5 km at four of the 28 breeding colonies remaining after 1977, including two Massa colonies still in existence today. An increasing density of well-maintained roads is clearly associated with the increasing disturbance by human activities.
Tourism (i.e. infrastructure for tourists such as resort construction as well as tourist activities) is further threatening breeding colonies of the NBI. Problematic activities include kitesurfing at beaches, disturbance from birdwatchers and photo-naturalists, local fishermen near breeding colonies (Bowden et al. Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008) and sudden, unexpected activities like paragliding and paramotor flights along the coast and a variety of off-road activities (e.g. bikes, quads and SUVs).
Breeding colonies still persisting since 1977 are often located in remote, less disturbed areas in semiarid regions or in regions in the Middle Atlas where traditional agriculture and extensive grazing are still in use. However, breeding sites in arid regions must be considered refuge sites with reduced habitat quality due to the high variability of annual rainfall. This appears to be the case not only in Morocco but also in Algeria (Collar and Stuart Reference Collar and Stuart1985, Fellous Reference Fellous2004) and Syria (Serra et al. Reference Serra, Peske, Abdallah, al Qaim and Kanani2009).
The four remaining breeding colonies today (three at SMNP and one near Tamri) have no particular habitat characteristics that differ from the original sites. All actual colonies are located on sea cliffs, below 500 m, close to water bodies with rainfed agriculture and fallow land and extensive pastures in the nearby surroundings. However, they differ clearly in the amount of annual rainfall (Tamri: average about 240 mm, SMNP: average about 140 mm). The main factors in their survival was their remoteness from civilization and that they were protected since 1995 (Bowden and Smith Reference Bowden and Smith1997) by an efficient observation system with wardens. The low amout of rainfall at SMNP means that the three colonies located in this reserve are actually living under rather marginal ecological conditions (see below). In contrast, the colony at Tamri has better ecological conditions with a higher amount of annual rainfall. The two new colonies founded in 2017 are also located in the same coastal region as the Tamri colony.
However, the reason for the complete breeding failure in the colony near Tamri in 2012 is not clear. Possible causes are a high level of disturbance at the beginning of the breeding period, predators (e.g. Lanner Falcon Falco biarmicus, Raven Corvus corax; and Pharaoh Eagle Owl Bubo ascalaphus; C. Bowden pers. comm.) or competition with other nesting bird species (e.g. Great Cormorant Phalacrocorax carbo maroccanus). Increasing human disturbance (e.g. tourists and local fishermen), unusually low precipitation (only 23 mm from January to June 2012; meteoblue.com 2017) and cold and persistent strong winds from the Atlantic Ocean along with coastal fog when the chicks were small or a combination of these factors are additional possible causes.
In spite of this sobering breeding failure of the colony at Tamri, the fact that the remaining NBI colonies have survived under distinct ecological conditions that are not different from the many extinct colonies in Morocco is also a reason for hope. It shows that there is still potential for recolonisation of formerly inhabited, suitable sites, or for new colonization of sites with appropriate habitat conditions, as the encouraging example of the two new colonies north of Tamri proves.
Correlation between number of breeding pairs and fledglings
As expected, the number of fledglings was closely and positively correlated with the number of reproducing pairs in Tamri. However, this correlation was not found at SMNP, indicating that ecological factors other than the simple number of reproducing pairs are more important for fledgling survival. Such factors may include population structure, climatic conditions including cold and persistent strong winds from the Atlantic, predator pressure or disturbance by paragliders along the coast during the breeding period of the NBI. This observation is also consistent with the different effects of rainfall on the number of fledglings at the two locations (see below).
Number of fledglings and precipitation
The clear increase in the number of birds from 1994 to 2016 in the remaining populations of the NBI is also reflected in the number of fledglings. This is a hopeful perspective, as it confirms that the conservation measures that were applied in the recent past had a positive effect on NBI populations and should also be followed in the future. Although the SMNP seems to be a site with poorer habitat quality, the higher number of breeding pairs than in Tamri is also reflected in the higher number of fledglings, even if it is not statistically significant in the applied model.
As expected in an arid region, rainfall clearly affects the number of fledglings, as it also affects food availability, particularly the availability of arthropods. However, this relationship is not a simple positive correlation (i.e. higher precipitation = more fledglings) as one might expect. This positive correlation is only true for the first quarter of the year in which fledglings hatched (Figure 4). Surprisingly, the number of fledglings decreases in Oulad Noumer with increasing amounts of rainfall during the last quarter of the previous year (Figure 3). A possible explanation of this rather unexpected pattern may include the following factors: (i) Increasing rainfall later in the year at SMNP motivates farmers to cultivate fallow land more intensely than in years with less precipitation, thereby not only increasing disturbance, but more importantly, strongly reducing the area of fallow land and favourable feeding grounds for the NBI. This is especially true for the most suitable feeding grounds north of the Oued Massa estuary (Rice et al. Reference Rice, Aghnaj, Bowden, Smith, Fox, Moore, Chamberlain and Wilson2002, Bowden et al. Reference Bowden, Smith, El Bekkay, Oubrou, Aghnaj and Jimenez-Armesto2008), which is interestingly also characterised by a particular soil type with a moderately deep A-horizon above marine, calcareous sand (Staimesse and Billaux Reference Staimesse and Billaux1978). (ii) Consistent with these factors is the observation of dead, most likely starved fledglings in nests at SMNP, whereas at Tamri fledglings mostly disappeared overnight, possibly due to predation or loss of fledglings by falling out of nests (Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003). (iii) Interspecific predation and inter- and intraspecific competition for breeding places may affect SMNP and Tamri populations in similar ways.
The contrasting situation regarding rainfall in Oulad Noumer and Tamri may also at least partly explain the difference in the correlation between reproducing breeding pairs and fledglings in these locations, again indicating that the Tamri colony, although smaller, is actually living under more favourable ecological conditions than the colony in SMNP.
Conclusions for conservation
Effective protection measures in south-west Morocco during the last two decades have not only prevented a further decline of the last wild population of the NBI, but have even caused a steady growth of these populations. Maintaining the wardening system is important and essential for the recovery of the population. In 2017, two new sites with small breeding colonies were detected on the Atlantic coast (Aourir et al. Reference Aourir, Bousadik, El Bekkay, Oubrou, Znari and Qninba2017). Further foundations of new populations on the Atlantic coast or in inland Morocco seem possible, even if such sites seem hard to predict at present.
A clear long-term goal of the International Single Species Action Plan (Bowden Reference Bowden2015) is to safeguard and if ever possible to increase wild populations of the NBI in Morocco and to manage and build up the semi-wild population in Turkey. The most desirable outcome would be natural, spontaneous recolonisation of still appropriate breeding sites in Morocco. However, natural recolonisation, particularly in inland Morocco, may be slow or not occur at all. Thus, additional measures including translocation projects may be necessary (Boehm Reference Boehm, Boehm and Bowden2016, Bowden et al. Reference Bowden, Boehm, Lopez, Hatipoglu, Boehm and Bowden2016, Dereliev Reference Dereliev, Boehm and Bowden2016). The assessment of former breeding sites in Morocco revealed two positive key factors: (I) No human disturbances in the immediate vicinity of the nesting sites and (II) suitable feeding habitats within a relatively short distance of 5–15 km.
Based on the available data, only a general assessment of the suitability of former breeding sites for future translocation projects is possible. However, in the case of possible future translocation activities, a preceding detailed on-site assessment is mandatory. For future recolonisation inland and along the coast, suitable feeding habitats within a radius of 5–15 km provide favourable conditions for successful chick rearing, while far away feeding grounds with a radius of 25 km and more lead to corresponding additional costs (time and flight energy) which may have a negative effect on the breeding success. The restriction to a distance of 5–15 km for suitable foraging habitats corresponds to published field observations (Rencurel Reference Rencurel1974, Collar & Stuart Reference Collar and Stuart1985, Akçakaya Reference Akçakaya1990, Boehm Reference Boehm, Boehm and Bowden2016, Yeniyurt et al. Reference Yeniyurt, Oppel, Isfendiyaroglu, Ozkinaci, Erkol and Bowden2017) and a questionnaire (Fellous Reference Fellous2011). Distant feeding habitats 25 to 35 km away probably reduce the breeding success due to higher exposure to predation (Serra et al. Reference Serra, Peske, Abdallah, al Qaim and Kanani2009) and deaths of chicks in the nest suggesting starvation (Bowden et al. Reference Bowden, Aghnaj, Smith and Ribi2003).
Based on the earlier distribution of breeding colonies (see Figure 1) in the years after 1960 and 1970, three regions for successful long-term establishment of a NBI population can be identified, the Atlantic coast with adjacent areas in the south-west of Morocco, the western Middle Atlas and the north-eastern Moulouya valley with adjacent plains and scattered island-like hill ranges.
With regard to the two key factors mentioned above, two types of breeding site can be identified. (A) Isolated and scattered breeding sites such as No. 59 and 66. The critical component of this type is its remoteness and difficult accessibility. (B) Easily accessible breeding sites such as No. 45 and 67. For these breeding sites a potentially higher level of human disturbance in the immediate vicinity of the nesting site is the main problem. On the other hand, this type offers advantages for required activities such as monitoring of the breeding success by locally recruited persons, wardening and close supervision as well as local awareness or guided tours for birdwatchers. As a recent example of the NBI reintroduction programme in Andalusia/Spain shows (Lopez and Quevedo Reference Lopez, Quevedo, Boehm and Bowden2016), a breeding colony can also settle in the immediate vicinity of a busy road (M. Quevedo, pers. comm. 2019) if other factors such as suitable forage habitats are present.
The translocation of breeding colonies in the previously occupied Middle Atlas at elevations between 1,000 and 2,000 m asl is desirable. However, it is uncertain if inexperienced adult NBI and their offspring can successfully avoid the harsh climatic conditions during the winter months by migrating to climatically more favourable regions as the NBI populations in the Middle Atlas did in the past.
Based on our detailed knowledge of the ecological requirements of the NBI, we state the following recommendations for possible translocation projects in Morocco some time in the future, and with adaptations to the local conditions, possibly also in Algeria, Turkey and perhaps Syria. For the latter two, migration remains a complex problem (Lindsell et al. Reference Lindsell, Serra, Peske, Abdullah, Al Qaim, Kanani and Wondafrash2009, Serra et al. Reference Serra, Lindsell, Peske, Fritz, Bowden, Bruschini, Welch, Tavares and Wondafrash2014). A possible approach could be the encouraging experience with human-led migration (Fritz et al. Reference Fritz, Hofmann, Unsöld, Boehm and Bowden2016).
General recommendations:
• Any proposed reintroduction site should be subject to a detailed feasibility study in line with AEWA guidelines (Dereliev Reference Dereliev, Boehm and Bowden2016, Seddon Reference Seddon, Bowden and Bowden2016) before other work begins. Such a feasibility study incorporates the following points.
• Benefit from the lessons learned in reintroduction projects in Central Europe (Fritz et al. Reference Fritz, Hofmann, Unsöld, Boehm and Bowden2016) and southern Spain (Lopez and Quevedo Reference Lopez, Quevedo, Boehm and Bowden2016), attempts to restore free-flying NBI colonies in Birecik/south-east Turkey (Akçakaya Reference Akçakaya1990, Hatipoglu Reference Hatipoglu, Boehm and Bowden2016) and from experiences supplementing juvenile birds to the remaining NBI colonies in Syria (Fritz and Riedler Reference Fritz and Riedler2010).
• A future cooperation with the National Zoo at Rabat and the new zoo Aïn Sebaâ at Casablanca should be examined.
Specific recommendations:
• Use experience and lessons learned from the Moroccan National Park team (system of wardens from local villages, awareness building and collecting data).
• Draw necessary conclusions from the failed project in Beshar el Kheir at Ain Tija-Mezguitem in north-east Morocco (poor breeding success and high mortality, unsuitable site) (Pfistermueller Reference Pfistermueller, Boehm and Bowden2009).
• Criteria for identifying potential breeding sites:
◦ Sites should be suitable cliffs and steep rock faces with low erosion risk.
◦ There should be no or only minor disturbance in the immediate vicinity of the breeding colonies, and if necessary, use a system of local rangers.
◦ There should be an appropriate, suitable feeding habitat within a site perimeter of up to 25 km (i.e. open, extensively cultivated areas with traditional agriculture and fallow land, treeless extensive summer pastures, open steppe and low and scattered vegetation).
◦ Water bodies should be available with fresh water within a perimeter of 5–10 km during the breeding season.
◦ Sites should be found without nearby powerlines and existing powerlines within a perimeter of 10–15 km should be mapped. If necessary, change dangerous line pole construction (OFEV et al. 2009, Prinsen et al. Reference Prinsen, Smallie, Boere, Pires and Compilers)2012).
◦ Choose sites near or in protected areas such as national parks, Sites d’intérêts biologiques et écologiques SIBE (Benabid Reference Benabid2000), Ramsar wetlands, Important Bird Areas IBA (Magin Reference Magin, Fishpool and Evans2001), hunting reserves, areas with prohibited access due to military use or land owned by ‘Eaux et Forêts’ and controlled by their personnel.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0959270919000170
Acknowledgements
We thank Madeleine Leutenegger from the Basel Zoo for species records in the archive of the Basel Zoo. Thanks to Tobias Roth, University of Basel, for his support with Figure 4 and to Martina L. Schenker for creating the distribution map. We appreciate the constructive comments of two anonymous reviewers and Chris Bowden, RSPB on an earlier version of this manuscript.