


Reading is a complex cognitive process that includes visual analysis of print, recognition of word form, and conversion from orthography to phonology and meaning. This process of reading is universal across different languages with accommodation to special features of each language at each level of the process (Perfetti & Liu, Reference Perfetti and Liu2005). Neuroimaging studies have identified a neural circuit involved in reading which includes left temporo-occipital regions for visual-orthographic recognition, left inferior parietal lobule for conversion from orthography to phonology and semantics, left superior temporal gyrus for phonological representation, left middle temporal gyrus for semantic representation, and left inferior frontal gyrus for semantic, syntactic and phonological processing (Pugh, Shaywitz, Shaywitz, Constable, Skudlarski, Fulbright, Bronen, Shankweiler, Katz, Fletcher & Gore, Reference Pugh, Shaywitz, Shaywitz, Constable, Skudlarski, Fulbright, Bronen, Shankweiler, Katz, Fletcher and Gore1996; Fiez & Petersen, Reference Fiez and Petersen1998; Booth, Burman, Meyer, Gitelman, Parrish & Mesulam, Reference Booth, Burman, Meyer, Gitelman, Parrish and Mesulam2002) (Figure 1). Learning to read in a second language follows the same cognitive processes and therefore provides a great model to study the process of reading acquisition in the brain (Xue, Mei, Chen, Lu, Poldrack & Dong, Reference Xue, Mei, Chen, Lu, Poldrack and Dong2010). The neural network of reading shows an inverted U-shaped function with the acquisition of expertise in the second language reading (Price, Reference Price2013). The rising part of the inverted U is illustrated by studies showing that learning to read is related to increased activation in the left temporo-occipital regions including the visual-word-form-area, irrespective of whether learning is focused on lexical or sublexical strategies (Mei, Xue, Lu, He, Zhang, Xue, Chen & Dong, Reference Mei, Xue, Lu, He, Zhang, Xue, Chen and Dong2013; Pugh, Landi, Preston, Mencl, Austin, Sibley, Fulbright, Seidenberg, Grigorenko, Constable, Molfese & Frost, Reference Pugh, Landi, Preston, Mencl, Austin, Sibley, Fulbright, Seidenberg, Grigorenko, Constable, Molfese and Frost2013), suggesting this region is more sensitive to the new script after initial learning during which meaning and phonology are attached to it. The falling part of the inverted U is illustrated by studies showing that activation in the left temporo-occipital regions decreases with the familiarity of the script (Twomey, Kawabata Duncan, Hogan, Morita, Umeda, Sakai & Devlin, Reference Twomey, Kawabata Duncan, Hogan, Morita, Umeda, Sakai and Devlin2013) and studies that show reduced activation in other parts of the reading network. A study that looked at the acquisition of literacy in a group of adults who were illiterate in their first acquired language but literate in their second language – which was German – (Abutalebi, Keim, Brambati, Tettamanti, Cappa, De Bleser & Perani, Reference Abutalebi, Keim, Brambati, Tettamanti, Cappa, De Bleser and Perani2007) has found that with learning of the orthography of their first language, there is reduced activation in left middle, inferior frontal gyrus, left superior temporal gyrus, left precuneus, left inferior parietal lobule and left inferior fusiform gyrus. Thus, the buildup of a new orthographic lexicon may be achieved through the interplay of these brain areas that sub-serve different aspects of reading and reading acquisition. Another training study found that learning to read Chinese in a group of English speakers is related with reduced brain activation in left superior parietal lobule, left fusiform gyrus and left inferior frontal gyrus (Deng, Booth, Chou, Ding & Peng, Reference Deng, Booth, Chou, Ding and Peng2008). Taken together, training studies suggest that there is an inverted U-shaped function in the reading network with increased expertise in second language reading; however a longitudinal study that can capture the complete learning curve in the brain is still much needed.
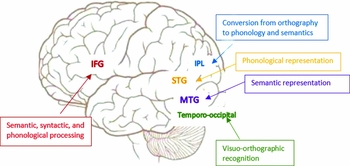
Figure 1. Demonstration of important regions in the reading network.
Orthographic transparency influences brain activation in L2 reading
Over and above the inverted U-shaped learning function, brain activation also varies with the reading strategy adopted. According to dual route models of reading, single words can be read in two distinct ways: a direct ‘lexical’ route and an indirect ‘sublexical’ route (Coltheart, Curtis, Atkins & Haller, Reference Coltheart, Curtis, Atkins and Haller1993). Orthographic differences across languages impose differential weighting on distinct component processes, and consequently on different routes/pathways during word-reading tasks (Coltheart & Crain, Reference Coltheart and Crain2012). The dorsal pathway including the left temporo-parietal region is engaged in the sublexical route while the ventral pathway including the left fusiform gyrus and left inferior frontal gyrus is engaged in the lexical route (Paulesu, McCrory, Fazio, Menoncello, Brunswick, Cappa, Cotelli, Cossu, Corte, Lorusso, Pesenti, Gallagher, Perani, Price, Frith & Frith, Reference Paulesu, McCrory, Fazio, Menoncello, Brunswick, Cappa, Cotelli, Cossu, Corte, Lorusso, Pesenti, Gallagher, Perani, Price, Frith and Frith2000; Bolger, Hornickel, Cone, Burman & Booth, Reference Bolger, Hornickel, Cone, Burman and Booth2008; Zhang, Chen, Xue, Lu, Mei, Xue, Wei, He, Li & Dong, Reference Zhang, Chen, Xue, Lu, Mei, Xue, Wei, He, Li and Dong2014). Readers of transparent orthographies such as Italian and Hindi are thought to rely on spelling-to-sound assembly and show increased activation in phonologically tuned areas along the dorsal pathway including the left temporo-parietal region (Paulesu et al., Reference Paulesu, McCrory, Fazio, Menoncello, Brunswick, Cappa, Cotelli, Cossu, Corte, Lorusso, Pesenti, Gallagher, Perani, Price, Frith and Frith2000, Sakurai, Momose, Iwata, Sudo, Ohtomo & Kanazawa, Reference Sakurai, Momose, Iwata, Sudo, Ohtomo and Kanazawa2000), whereas reading an opaque orthography such as English is thought to rely more on lexically mediated processing associated with increased activation of semantically tuned regions along the ventral pathway including the left fusiform gyrus and inferior frontal gyrus (Tokunaga, Nishikawa, Ikejiri, Nakagawa, Yasuno, Hashikawa, Nishimura, Sugita & Takeda, Reference Tokunaga, Nishikawa, Ikejiri, Nakagawa, Yasuno, Hashikawa, Nishimura, Sugita and Takeda1999; Paulesu et al., Reference Paulesu, McCrory, Fazio, Menoncello, Brunswick, Cappa, Cotelli, Cossu, Corte, Lorusso, Pesenti, Gallagher, Perani, Price, Frith and Frith2000; Chen, Fu, Iversen, Smith & Matthews, Reference Chen, Fu, Iversen, Smith and Matthews2002; Chen, Vaid, Bortfeld & Boas, Reference Chen, Vaid, Bortfeld and Boas2008).
Bilingual studies have demonstrated the distinct pathways corresponding to different orthographic transparency in a within-subject manner (Cherodath & Singh, Reference Cherodath and Singh2015). For example, quite a few studies have found that left middle/inferior frontal gyrus and left fusiform gyrus are more involved in the more opaque language in a bilingual context, greater for Chinese than for English in English–Chinese bilinguals (Nelson, Liu, Fiez & Perfetti, Reference Nelson, Liu, Fiez and Perfetti2009), greater for Urdu than for Hindi in Urdu–Hindi equal bilinguals (Kumar, Reference Kumar2014), greater for English than for Spanish in Spanish–English equal bilinguals (Jamal, Piche, Napoliello, Perfetti & Eden, Reference Jamal, Piche, Napoliello, Perfetti and Eden2012), greater for English than French in French–English bilinguals (Berken, Gracco, Chen, Watkins, Baum, Callahan & Klein, Reference Berken, Gracco, Chen, Watkins, Baum, Callahan and Klein2015), and greater for English than for Hindi in English–Hindi bilinguals (Das, Padakannaya, Pugh & Singh, Reference Das, Padakannaya, Pugh and Singh2011). In contrast, the dorsal pathway including the left temporo-parietal regions is more involved in the more transparent language: greater for Spanish than for English in Spanish–English equal bilinguals (Jamal et al., Reference Jamal, Piche, Napoliello, Perfetti and Eden2012), greater for German than for French in French–German bilinguals (Buetler, de Leon Rodriguez, Laganaro, Muri, Spierer & Annoni, Reference Buetler, de Leon Rodriguez, Laganaro, Muri, Spierer and Annoni2014), and greater for Hindi than for English in English–Hindi bilinguals (Das et al., Reference Das, Padakannaya, Pugh and Singh2011). Taken together, orthographic transparency plays an important role in determining brain activation during word reading in both L1 and L2.
In the bilingual situation, however, whether the transparent language is acquired first or the opaque language is acquired first also makes a difference in brain activation of L2. When L1 is more opaque, for example, in Chinese–English bilinguals, English reading evokes a network which is identical to Chinese reading (Chee, Caplan, Soon, Sriram, Tan, Thiel & Weekes, Reference Chee, Caplan, Soon, Sriram, Tan, Thiel and Weekes1999; Tan, Spinks, Feng, Siok, Perfetti, Xiong, Fox, Gao & Kalogirou, Reference Tan, Spinks, Feng, Siok, Perfetti, Xiong, Fox, Gao and Kalogirou2003; Cao, Tao, Liu, Perfetti & Booth, Reference Cao, Tao, Liu, Perfetti and Booth2013), suggesting that the existing mechanisms are sufficient for the new L2. When L1 is more transparent than L2, for example, in English–Chinese bilinguals, significant accommodations were observed including greater involvement of the left middle frontal gyrus and the right fusiform gyrus for Chinese than English reading (Liu, Dunlap, Fiez & Perfetti, Reference Liu, Dunlap, Fiez and Perfetti2007; Nelson et al., Reference Nelson, Liu, Fiez and Perfetti2009), suggesting that additional neural resources are required for the new L2. One possible explanation is that when L2 is more transparent than L1 (i.e., in Chinese–English bilinguals), the existing L1 system is sophisticated enough for the more regular mapping in L2; while when L2 is more opaque than L1 (i.e., in English–Chinese bilinguals), additional new mechanisms are required in order to deal with the arbitrary mapping between orthographic and phonology in L2. Future research needs to test this hypothesis by comparing two bilingual groups with the same L2 but different L1. In one group, L2 is more opaque than L1 and in the other group, L2 is more transparent than L1, such as Chinese–English and Spanish–English bilinguals.
Proficiency effect in the bilingual brain
Proficiency is another intriguing factor that influences brain activation in the bilingual brain. General proficiency effects have been reported that L2 reading is related with an extended network than L1 reading including but not limited to the left prefrontal cortex and anterior cingulate cortex, because of the lower proficiency in L2 than in L1 (Stowe & Sabourin, Reference Stowe and Sabourin2005, Indefrey, Reference Indefrey2006, Abutalebi, Reference Abutalebi2008, Sebastian, 2011). However, even when proficiency is matched, there seems to be greater activation in the attention network for L2 than L1. For example, Kovelman and colleagues showed that bilateral dorsal lateral prefrontal cortex and inferior frontal cortex are more involved in bilinguals than in monolinguals in English semantic processing (Kovelman, Shalinsky, Berens & Petitto, Reference Kovelman, Shalinsky, Berens and Petitto2008) and English syntactic processing (Kovelman, Baker & Petitto, Reference Kovelman, Baker and Petitto2008) when comparing early Spanish–English bilinguals to English monolinguals with matched proficiency on English. Taken together, one fundamental difference between L1 processing and L2 processing is the greater involvement of the attentional control network in L2 which is not completely explainable by proficiency.
Studies have also examined whether Higher L2 proficiency is related with greater similarity to L1 or greater accommodation to L2. There has been evidence that higher proficiency in the second language is related with greater similarity to brain activation in the first language during semantic judgment in a group of English–German bilinguals (Stein, Federspiel, Koenig, Wirth, Lehmann, Wiest, Strik, Brandeis & Dierks, Reference Stein, Federspiel, Koenig, Wirth, Lehmann, Wiest, Strik, Brandeis and Dierks2009), and during visual word rhyming judgment in a group of late Chinese–English bilinguals (Cao et al., Reference Cao, Tao, Liu, Perfetti and Booth2013). In contrast, there are also studies that have found higher proficiency in the second language is related with greater accommodation to L2. Studies found that in late English–Chinese bilinguals, high proficiency in L2, Chinese was associated with increased brain activation in two ‘Chinese’ regions, right superior parietal lobule and right lingual gyrus in a lexical decision task (Cao, Vu, Chan, Lawrence, Harris, Guan, Xu & Perfetti, Reference Cao, Vu, Chan, Lawrence, Harris, Guan, Xu and Perfetti2013). In another ‘Chinese’ region (i.e., left superior parietal lobule), there was increased activation with greater proficiency improvement from early learning to late learning in a meaning match judgment task (Deng et al., Reference Deng, Booth, Chou, Ding and Peng2008). The three ‘Chinese’ regions from the two studies mentioned above are found to be more involved in Chinese L1 readers than in English L1 readers (Bolger, Perfetti & Schneider, Reference Bolger, Perfetti and Schneider2005; Tan, Laird, Karl & Fox, Reference Tan, Laird, Karl and Fox2005), suggesting that higher proficiency in L2 is related with greater accommodation to the new language. In consistent, a recent study on late Chinese–English bilinguals has found that higher proficiency in L2 (i.e., English) is characterized by greater connectivity with the left supramarginal gyrus and left superior temporal gyrus (Cao, Kim, Liu & Liu, Reference Cao, Kim, Liu and Liu2014). Both of these regions have been found to be more involved in English L1 readers than in Chinese L1 readers (Bolger et al., Reference Bolger, Perfetti and Schneider2005; Tan et al., Reference Tan, Laird, Karl and Fox2005; Cao, Brennan & Booth, Reference Cao, Brennan and Booth2015), presumably because this temporo-parietal region is involved in fine-grained mapping between orthography and phonology such as grapheme-phoneme-correspondence in reading (Bookheimer, Zeffiro, Blaxton, Gaillard & Theodore, Reference Bookheimer, Zeffiro, Blaxton, Gaillard and Theodore1995; Booth et al., Reference Booth, Burman, Meyer, Gitelman, Parrish and Mesulam2002). In summary, previous studies have produced controversial findings about whether high proficiency in L2 is related with greater similarity to L1 or greater accommodation to L2.
AOA of L2 affects how L2 is processed in the brain
Another factor that determines brain activation in L2 processing is age of acquisition. Even when proficiency is matched in early and late bilinguals, brain activation for L2 varies according to the age of acquisition. One study found that early simultaneous bilinguals showed distinct activation for English and Hindi with greater activation in left inferior temporal gyrus for English and greater activation in left inferior parietal lobule for Hindi in a word reading task. However, late bilinguals with a comparable proficiency level to the early bilinguals activated similar network for both languages, suggesting greater accommodation in early bilinguals than in late bilinguals (Das et al., Reference Das, Padakannaya, Pugh and Singh2011). On the other hand, there is also well documented evidence that accommodation is more prominent in late bilinguals and assimilation is more prominent in early bilinguals. A study by Kim et al. (Kim, Relkin, Lee & Hirsch, Reference Kim, Relkin, Lee and Hirsch1997) found that late bilinguals showed distinct activation within the Broca's area, while early bilinguals showed an overlap between two languages in Broca's area in a silent internally expressive linguistic task (Kim et al., Reference Kim, Relkin, Lee and Hirsch1997). Another piece of evidence for greater assimilation in early bilingual and greater accommodation in late bilinguals comes from a sentence reading study, in which researchers found that for both grammatical and semantic processing, early bilinguals showed little or no distinction between L1 (i.e., Italian) and L2 (i.e., German), while late bilinguals showed greater involvement of left inferior frontal gyrus in L2 than in L1, even when their proficiency was comparable to the early bilinguals (Wartenburger, Heekeren, Abutalebi, Cappa, Villringer & Perani, Reference Wartenburger, Heekeren, Abutalebi, Cappa, Villringer and Perani2003). Another study found increased activation in left IFG, left fusiform gyrus, left premotor cortex, orthography phonology mapping region, speech-motor regions in L2 reading aloud than L1 in sequential late French–English bilinguals, while early simultaneous bilinguals showed an overlapped network (Berken et al., Reference Berken, Gracco, Chen, Watkins, Baum, Callahan and Klein2015). While the studies mentioned above suggest controversial conclusions about how AOA influences assimilation and accommodation, there is also research that suggests AOA does not play a role in the pattern of assimilation and accommodation. By directly comparing early and late bilinguals, one study found that similar networks were involved in both groups for both L1 and L2, suggesting that for both early and late bilinguals, assimilation is dominant (Chee, Tan & Thiel, Reference Chee, Tan and Thiel1999; Yang, Tan & Li, Reference Yang, Tan and Li2011). Taken together, findings of how AOA influences assimilation and accommodation are not consistent.
L1 influences how L2 is being learned in the brain
The established L1 skills actually constrain what can be learned in L2 and how L2 can be learned (Li & Farkas, Reference Li and Farkas2002). One fMRI study found supporting evidence for cross-linguistic influences of L1 on L2. In this study, it was found that native Chinese speakers showed greater activation in left middle frontal gyrus than native Korean speakers in reading Japanese kana, with matched AOA and proficiency in the two bilingual groups (Yokoyama, Kim, Uchida, Miyamoto, Yoshimoto & Kawashima, Reference Yokoyama, Kim, Uchida, Miyamoto, Yoshimoto and Kawashima2013). Left middle frontal gyrus has been found to be more involved in Chinese reading than other languages, probably due to the unique whole-character to whole-syllable mapping procedure in reading (Berken et al., Reference Berken, Gracco, Chen, Watkins, Baum, Callahan and Klein2015). The finding from Yokoyama et al.’s study suggests that the brain regions and cognitive strategies involved in reading a L2 is constrained by L1. In learning to read a second language, the properties of the L2 writing system and the distance between L1 and L2 also affects the learning process (Gleitman, Reference Gleitman1985, Bassetti, Reference Bassetti2008). Jeong et al. (Jeong, Sugiura, Sassa, Haji, Usui, Taira, Horie, Sato & Kawashima, Reference Jeong, Sugiura, Sassa, Haji, Usui, Taira, Horie, Sato and Kawashima2007) found that native Korean speakers showed greater similarity to their native reading network when reading Japanese sentences than when reading English sentences. This was interpreted as a reflection of greater grammar similarity between Korean and Japanese than that between Korean and English. When the study was conducted among native Chinese speakers, however, it was found that there was greater similarity to native reading network when reading English sentences than reading Japanese sentences (Jeong, Sugiura, Sassa, Yokoyama, Horie, Sato, Taira & Kawashima, Reference Jeong, Sugiura, Sassa, Yokoyama, Horie, Sato, Taira and Kawashima2007). Again it is because Chinese and English grammar is more similar than Chinese and Japanese grammar. Therefore, a limited number of studies tend to suggest that short distance between L1 and L2 is associated with greater similarity and long distance is associated with greater accommodation.
Learning a second language changes how the first language is processed
Traditionally, researchers have not entertained the idea that one's native language is something subject to influence from the second language, but more recent evidence does suggests that L1 is more permeable than we thought, and is such that L2 to L1 influences not only exist for early simultaneous bilinguals, but also for late, successive bilinguals (Pavlenko & Malt, Reference Pavlenko and Malt2011). Very few neuroimaging studies have examined how L1 network changes after a second language is acquired. One study found that left ventral prefrontal activation for first language reading increases with second language vocabulary knowledge (Nosarti, Mechelli, Green & Price, Reference Nosarti, Mechelli, Green and Price2009). One recent study also found greater activation in left inferior frontal gyrus, superior temporal gyrus, inferior parietal lobule and right homologues in bilinguals than monolinguals in reading regular, irregular, nonsense words in L1 (Jasinska & Petitto, Reference Jasinska and Petitto2014). It suggests that the experience of exposure to two phonological systems results in greater activation in the phonological processing regions in the brain. Another study found that long-term experience of Chinese is associated with increased involvement of the right fusiform gyrus in native English speakers, suggesting that L2 can modulate how L1 is processed at the visual-orthographic level as well (Mei, Xue, Lu, Chen, Wei, He & Dong, Reference Mei, Xue, Lu, Chen, Wei, He and Dong2015). In summary, it suggests that knowing a second language fundamentally changes how the brain processes languages and that the greater activation in the language network and brain regions involved in monitoring and attention may be related with the enhanced linguistic competition and cognitive processing.

