



Introduction
The late pre-Hispanic people of the Sierras of Córdoba (Argentina, 1500–360 years BP) practised a mixed foraging and cultivation economy with a flexible mobility pattern that included semi-permanent villages and small sites scattered across the landscape (Medina et al. Reference Medina, Pastor and Recalde2016; Figure 1). The bone-tool technology indicates a diverse subsistence pattern adapted for the processing of resources and by-products acquired through a broad-spectrum farming and foraging base (Medina et al. Reference Medina, Buc and Pastor2014). There are few technological studies focused on the form and function of these tools, however, even though they are a common artefact-class in late pre-Hispanic assemblages (Serrano Reference Serrano1945; Marcellino et al. Reference Marcellino, Berberián and Perez1967; Berberián Reference Berberián1984; Laguens & Bonnin Reference Laguens and Bonnin2009).

Figure 1 Satellite imagery showing the geographic location of Boyo Paso 2 in the Sierras of Córdoba, Argentina (© Google Earth).
Large animal bones, used as raw materials or food, are a conspicuous part of the late pre-Hispanic archaeological record (Medina & Pastor Reference Medina and Pastor2012), yet, as a result of poor preservation, plant food remains are not. Consequently, the roles of wild and domesticated plant resources have been difficult to ascertain, and the first archaeological evidence for plant consumption has only recently been obtained. Maize (Zea mays), squash (Cucurbita sp.), beans (Phaseolus spp.), quinoa (Chenopodium quinoa var. quinoa), ajara (Chenopodium quinoa var. melanospermum), carob (Prosopis spp.), chañar (Geoffroea decorticans) and mistol (Sarcomphalus mistol) have been identified in late pre-Hispanic sediments and on various tools (Medina et al. Reference Medina, Lopez and Berberián2009; Pastor & López Reference Pastor and López2010; López & Recalde Reference –2016; Medina et al. Reference Medina, Pastor and Recalde2016). The role of tubers and other underground plant organs as components of late pre-Hispanic diets has often been downplayed, although they are mentioned in sixteenth- and seventeenth-century AD ethnohistoric records (see Berberián Reference Berberián1984). Recently, however, starch grains from potatoes (Solanum cf. S. tuberosum), and probably also sweet potatoes (cf. Ipomea batata) or manioc (cf. Manihot sp.), have been identified in food residues from the pottery assemblage at the northen Sierras of Córdoba Quebrada Norte 7 site (López & Recalde Reference López and Recalde2015).
Here we present the results of a full morphological, use-wear and residue analysis of a notched bone tool recovered from the late pre-Hispanic site of Boyo Paso 2 (Sierras of Córdoba), revealing an unexpected use for the tool. The results challenge conventional assessments of the role of plant foods in pre-Hispanic subsistence practices (Figures 1–2). The artefact presented here is signficant for three reasons. First, it is of an uncommon type, as assemblages are usually dominated by awls and projectile points (Medina et al. Reference Medina, Buc and Pastor2014). Second, to avoid the limitations of functional assessment of tool use based only on morphology, we apply a multi-proxy approach of the sort typically reserved for lithic technologies (Kononenko et al. Reference Kononenko, Torrence, Barton and Hennell2010; Barton & Torrence Reference Barton and Torrence2015). Finally, the application of use-wear and residue analyses offers the potential to reveal a range of activities conducted at Boyo Paso 2 that could otherwise leave little or no trace in the archaeological record.

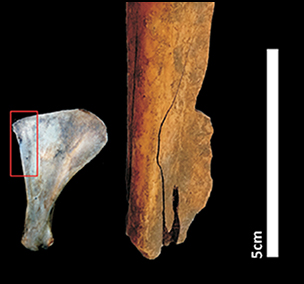
Figure 2 The notched bone tool made from a camelid scapula and recovered from Boyo Paso 2. The arrows show the active part of the tool that was subject to sonication. The image on the left shows which part of the scapula was used for the tool (photograph by Natacha Buc).
The site and its chronology
Boyo Paso 2 is an open-air site located at 1160m asl in the eastern Salsacate Valley of the Sierras of Córdoba (Figure 1). It is surrounded by sloping topography with organic-rich soils, well suited for farming, and by a xerophytic forest dominated by small trees with edible fruits such as carob, chañar and piquillín (Condalia microphylla). Recent excavations have revealed a lightly compacted living floor, or surface, with postholes, a semi-subterranean structure and abundant refuse spread across the exposed area, including the macroremains of edible plants (maize, beans and mistol), bones from a variety of animals and evidence of pottery making. The existence of habitational structures, such as pit-houses, is inferred from the presence of 23 postholes (Figure 3), although these overlap in such a way that it is impossible to determine the size and shape of the structures with confidence. The absence of well-defined hearth features makes identification even more difficult and suggests that the habitational structures were not intended for long-term residence.

Figure 3 Plain view of the archaeological floor from Boyo Paso 2 with the excavation units, postholes and other key features. The arrow shows the location of the notched bone tool referred to in the text.
Artefacts abandoned as refuse were found on the living floor, including the bone tool analysed here (Figure 3), grinding tools and fragmented pottery vessels (Figure 3) (Medina et al. Reference Medina, Pastor and Recalde2016). Among other bone tools that were recovered, awls, spatulas, projectile points and manufacturing debris were also recognised. The dated samples from the floor surface were modelled at 95.4% in OxCal v.4.3, using ShCal13 calibration curve (Bronk Ramsey Reference Bronk Ramsey2009; Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013). A wood charcoal sample was dated to 750±70 BP (LP-2932: 1205–1400 AD); a common bean remain was dated to 866±39 BP (AA110929: 1156–1277 AD); a maize remain was dated to 878±18 BP (AA110928: 1162–1265 AD); wood charcoals samples were also dated to 1060±50 BP (LP-3122: 897–1155 AD) and 1500±80 BP (LP-3107: 411–765 AD), suggesting that the deposit was formed over an extended period of time, even though many of the artefacts appear to have been found in situ (Medina et al. Reference Medina, Pastor and Recalde2016). Consequently, Boyo Paso 2 has been interpreted as a seasonally reoccupied encampment where small groups of people, practising a mixed foraging and cultivation economy, gathered and took part in a range of activities.
Methodology
On discovery, the bone tool was immediately sealed in a bag to avoid external and cross-specimen contamination (Allen & Ussher Reference Allen and Ussher2013). Once in the laboratory, the tool was handled with starch-free gloves and processed for microfossil extraction. Phytoliths and starch-grain food residues were recovered according to the protocols used by Piperno et al. (Reference Piperno, Ranere, Holst and Hansell2000), wherein stepwise extractions were performed to assess whether microfossil remains were associated with tool use. First, the tool was washed, and residues were examined to identify microfossils incorporated incidentally rather than those derived from use of the tool. The working or active edge was then subjected to sonic extraction to remove all remaining microfossils (Figure 2). It was assumed that the microfossils obtained using the ultrasonic bath were in close association with the serrated edge of the bone tool, demonstrating a clear relationship to tool function (Perry Reference –2007; Lanjegans Reference Lanjegans2011; Allen & Ussher Reference Allen and Ussher2013).
The residue samples obtained by both methods were filtered and dried for 24 hours. They were then mounted on glass slides using immersion oil, and scanned with an optical microscope at 500–1000× magnification under transmitted and polarised light. Phytoliths and starch grains were observed, photographed, described and compared with a reference collection, published atlases and keys for taxonomic identification (i.e. Loy Reference Loy1994; Cortella & Pochettino Reference Cortella and Pochettino1995; Piperno & Holst Reference Piperno and Holst1998; Perry Reference Perry2004; Piperno Reference Piperno2006; Korstanje & Babot Reference Korstanje and Babot2007; Babot & Del Reference Babot and Del2009). Tubers in the reference collection vary in provenance, including some purchased at traditional markets (e.g. O. tuberosa), and others collected during archaeological fieldwork (e.g. wild Oxalis spp., Pterocactus sp. and Maihueniopsis darwinii) (Figure 4). Modifications produced by post-harvest processing activities, such as grinding and boiling, were also assessed (Henry et al. Reference Henry, Hudson and Piperno2009; Crowther Reference Crowther2012; Li et al. Reference Li, Dhital and Hasjim2014; Barton & Torrence Reference Barton and Torrence2015).

Figure 4 Reference collection of modern edible underground plant organs under transmitted (a, c, e and g) and polarised light (b, d, f and g). Figure 4a–b) cultivated Oxalis tuberosa; c–d) unidentified wild Oxalis sp.; e–f) Pterocactus tuberosum; g–h) Maihuenopsis darwinii. Scale bar=20 µm.
The starch grains were documented with detailed descriptions of morphology, size, contours and surface traits, along with the shape and size of the hilum, the visibility of the lamellae and fissures, birefringence properties, extinction cross features, grain visibility under normal and polarised light, and the packing of compounded grains (Loy Reference Loy1994; Piperno & Holst Reference Piperno and Holst1998; Perry Reference Perry2004; Korstanje & Babot Reference Korstanje and Babot2007). The phytolith description follows the International Code for Phytolith Nomenclature (Madella et al. Reference Madella, Alexandre and Ball2005), including three-dimensional shape, symmetry and surface ornamentation.
The anatomical and taxonomic identification of the bone used to make the tool was based on an ungulate reference collection. Morphological and metric analyses were undertaken according to standard parameters (Buc Reference Buc2012). The process of manufacture was studied according to Legrand (Reference Legrand2007), Averbouh (Reference Averbouh2000), Kononenko et al. (Reference Kononenko, Torrence, Barton and Hennell2010), Buc (Reference Buc2012) and Anderson (Reference Anderson2013). Marks that are unrelated to tool use or manufacture (e.g. instances of weathering gauged using the Behrensmeyer (Reference Behrensmeyer1978) stage index or anthropomorphic modifications to bone surface) are described to assess any potential effects on use-wear analysis, and to explore questions of raw material acquisition (Behrensmeyer Reference Behrensmeyer1978; Lyman Reference Lyman1994).
Finally, the use-wear pattern of the active edge was described and photographed using binocular and metallurgical microscopes at 20–200× magnification (Legrand Reference Legrand2007; Buc Reference Buc2012). This was then compared with experimental analogues to identify potential tool functions (see Griffitts Reference Griffitts2006; Kononenko et al. Reference Kononenko, Torrence, Barton and Hennell2010; Stone Reference Stone2011; Buc Reference Buc2012; Anderson Reference Anderson2013).
Results
The tool was fashioned from the left scapula of a guanaco (Lama cf. L. guanicoe), a large mammal well represented in the faunal assemblage of Boyo Paso 2. Cut marks on the blade suggest that the scapula came from an animal killed for food, probably one that remained on site after primary butchering.
The tool was manufactured on the posterior border of the scapula, conserving the axillary border, the subscapular fossa and the posterior angle (Figure 2). Fifteen closely spaced notches were incised on the infraspinatous fossa using a stone tool to create a serrated edge (Figure 2). The highest bone-weathering stage displayed by the tool is the Behrensmeyer (Reference Behrensmeyer1978) stage 1, with cracking parallel to the fibre structure that has not excessively affected the bone surface and therefore does not affect the use-wear analysis. Likewise, the low level of weathering suggests that the artefact was exposed to sunlight, precipitation and other climate-related agents for only a short span of time, a condition that reinforces the short-term site-use interpretation.
The notches and areas between them appear shiny to the naked eye and exhibit polish under magnification (Figures 2 & 5a). When observed under a metallurgical microscope, narrow, plain, parallel striations that emanate from the notches and run perpendicular to the centre of the tool are visible (Figure 5b). Striations coming from different notches sometimes overlap (Figure 5b–e). The micro-relief is shiny, rounded and uninvasive (Figure 5d). Striations and topography are morphologically similar to those recorded on experimental bone tools used for plant-scraping activities, where the artefact is held transverse to the direction of motion (Figure 5f–h; see also Griffitts Reference Griffitts2006).

Figure 5 Use-wear pattern analysis of the notched bone tool from Boyo Paso 2: a) shiny surface (circled) visible on a notch viewed using binocular microscope (20×); b, c, e) parallel striations emanating from the notches viewed by metallurgical microscope (100×); d) non-invasive shiny surface on notches viewed by metallurgical microscope (100×); f–g) parallel striations on experimental bone tool used on silica-rich plants (photographs by N. Buc); h) non-invasive shiny surface on experimental bone tool used on silica-rich plants (from Stone Reference Stone2011: tab. 7.3). Arrows indicate striations.
A total of 39 starch grains were recovered from the active edge of the bone tool, showing that wear traces were an effective trap for the processed plant microfossils. Most of these (n=33) are similar to those from Oxalis tuberosa, an endemic tuber crop from South America and one of the most important members of the Andean root crop complex (Cortella & Pochettino Reference Cortella and Pochettino1995). The absence of features required for species-level identification and the lack of comparative material from local wild specimens such as O. conorrhiza and O. lasiopetala (Giorgis et al. Reference Giorgis, Cingolani, Chiarini, Chiapella, Barboza, Ariza, Morero, Gurvich, Tecco, Subilis and Cabido2011), however, means that they can only be tentatively identified as aff. Oxalis sp.
These microfossils are asymmetrical single starch grains with irregular shapes (conical, elongated and pear-shaped) (Figure 6). Grain sizes range from 10–50µm. Eccentric extinction crosses have irregular, dark broken arms intersecting at more than one point. The intensity of birefringence was high, and lamellae were not visible. Eccentric lineal hilum was viewed only in some grains. The absence of fissures, fractures, relief alterations, gelatinisation and damaged extinction crosses shows that these Oxalis starch grains have not been affected by processing or cooking activities such as dehydratation (for storage), boiling or toasting. This suggests that Oxalis sp. tubers were processed raw, and that the bone tool was used to peel the epidermis to reach the inner part of the tuber. Significantly, starch grains were absent from the washed residues resulting from post-depositional contamination, reinforcing the suggestion that the notched bone tool was used for wild or domestic Oxalis tuber processing.

Figure 6 Starch grains identified (aff. Oxalis sp.) from the notched bone tool from Boyo Paso 2 under transmitted (a, c and e) and polarised light (b, d and f); scale bar=20 µm.
A few phytoliths identified as wavy-top rondels, which only occure in maize, were recovered from the notched edge (Figure 7). Maize cob microfossils were also identified in the washed residues used as control samples, which makes the relationship between maize and the bone tool more difficult to clarify. Thus, these maize phytoliths are interpreted as potential contaminants derived from plant material that came into contact with the tool through deposition on the floor. The absence of maize starch grains close to the active edge, where they would be expected if the tool had been used for cob-related activities (Barton & Torrence Reference Barton and Torrence2015), supports this interpretation.

Figure 7 Identified phytoliths: maize-only, wavy-top rondel, recovered from the notched edge of the tool. Scale bar=20 µm.
Discussion
Bone tools with serrated working edges are uncommon in late pre-Hispanic assemblages from the Sierras of Córdoba. The only other well-known artefact made using a camelid scapula similar to that described here was recovered from the pit-house floor of Potrero de Garay (310±75 years BP; Berberián Reference Berberián1984), a site located approximately 70km from Boyo Paso 2. Unlike the Boyo Paso 2 find, the notches and the use-wear traces from the Potrero de Garay artefact are concentrated on the cranial border of the blade, with the epiphysis conserved for handling (Figure 8).

Figure 8 The camelid scapula notched bone tool from Potrero de Garay, Sierras of Córdoba (photograph by Sebastián Pastor).
The abundance of starch grains and the use-wear pattern around the serrated edge support the interpretation that the serrated bone tool from Boyo Paso 2 was used for peeling Oxalis sp. tubers, an activity with low archaeological visibility due to the poor preservation of soft-tissue, underground plant organs. Thus, despite the morphological similarities to tools used to strip off hulled grains of wheat (Triticum spp.), or to process fibres, skins and tendons, during the Neolithic or Formative transition (Stordeur & Anderson-Gerfaud Reference Stordeur and Anderson-Gerfaud1985; Moore Reference Moore1999; Northe Reference Northe1999; Luik et al. Reference Luik, Ots and Maldre2011; Stone Reference Stone2011; Anderson Reference Anderson2013; Capriles Reference Capriles2014; López Campeny Reference López Campeny2016), the analyses of the Boyo Paso 2 tool suggest a different use. Currently, the most similar functional analogue is the macropod scapula used as a yam (Dioscorea sp.) grater by the recent northern Australia foragers, although this tool is not fully described (Beck & Torrence Reference Beck and Torrence2006: 60).
Recent studies have reported the earlier than expected importance to the pre-Hispanic populations of South America of wild and domestic plant foods, particularly those with underground storage organs (Korstanje Reference Korstanje2015; Ochoa & Ladio Reference Ochoa and Ladio2015; Pochettino Reference Pochettino2015). For example, wild potatoes were used at the Late Pleistocene settlement of Monterverde c. 13 000 years BP, in southern Chile (Ugent et al. Reference Ugent, Dillehay and Ramirez1987). Tubers, roots and rhizomes have been well documented at several archaeological sites ranging from lowlands to higher-altitude areas in the Andes and from the Early Holocene (10 000–7000 BP) to the Inca period, suggesting a temporal trend towards the domestication of various genera such as S. tuberosum, O. tuberosa, Ullucus tuberosus, I. batata, M. esculenta, Canna edulis, Maranta arundinacea, Dioscorea spp. and Tropaeolum tuberosum (Cortella & Pochettino Reference Cortella and Pochettino1995; Iriarte Reference Iriarte2006; Babot & Del Reference Babot and Del2009, Reference –2011; Llano Reference Llano2014; Ochoa & Ladio Reference Ochoa and Ladio2015; Pochettino Reference Pochettino2015). The earliest evidence of Oxalis tuber consumption comes from the Guitarrero Cave and has been dated to 10 500–7500 years BP (Cortella & Pochettino Reference Cortella and Pochettino1995). More recently, wild or possibly cultivated Oxalis sp. has been recorded from grinding tools recovered from north-western Argentina (Babot & Del Reference Babot and Del2009, Reference –2011), but until now it has never been identified at sites from the Sierras of Córdoba or on bone-tool technology in South America.
These results also highlight the disproportionate attention afforded to maize in Argentinian archaeology, a trend that is now being questioned through archaeobotanical and palaeodietary isotope studies (e.g. Korstanje Reference Korstanje2015; Killian Galván et al. Reference Killian Galvan, Samec and Panarello2016). In this context, the identification of Oxalis in the Sierras of Córdoba, whether wild or domestic, is a significant discovery because the contribution of domesticated plants in the late pre-Hispanic diet—and especially maize—has been the subject of much recent debate (Pastor & López Reference Pastor and López2010; Medina et al. Reference Medina, Pastor and Recalde2016, Reference Medina, Grill, Fernandez and López2017). It is clear that late pre-Hispanic people had experience with cultigens (Pastor & López Reference Pastor and López2010). Palaeoecological and settlement pattern data indicate that extensive cultivation was being practised, and that the groups residing in the region were organised as seasonal or occasional food producers (Medina et al. Reference Medina, Pastor and Recalde2016). The archaeological evidence of pre-Hispanic crops in the Sierras of Córdoba does not support maize farming as the main economic or subsistence strategy of late pre-Hispanic people. Instead, the importance of cultigens fluctuated as part of a diverse foraging and cultivation economy, in which wild plant and animal resources continued to be extensively used.
Thus, several different lines of evidence combine to question the ‘maize-centric’ approach that has dominated discussion of the archaeology of the Sierras of Córdoba:
1) Maize macroremains are relatively few in number. They are only preserved when carbonised, which limits our ability to quantify the contribution of maize to diet, even when maize phytoliths and starch grains are a common food residue on pottery and grinding tool assemblages.
2) Archaeobotanical evidence, however, now provides robust evidence that maize made a contribution to diet alongside a variety of domesticated tubers, beans, squashes, pseudo-cereals and a diversity of wild plants (Medina et al. Reference Medina, Lopez and Berberián2009, Reference Medina, Grill, Fernandez and López2017; Pastor & López Reference Pastor and López2010; López et al. Reference López, Medina and Rivero2015; López & Recalde Reference López and Recalde2015, Reference –2016).
3) Identification of potato and sweet potato or manioc starch grains provides evidence for the calorific significance of root and tuber crops in pre-Hispanic diets (López & Recalde Reference López and Recalde2015).
4) High isotope values of 3C plants in human bones indicate the relative importance of foraging and other crops compared with non-indigenous 4C isotope values associated with maize farming (Laguens & Bonnin Reference Laguens and Bonnin2009; Laguens et al. Reference Laguens, Fabra, Santos and Demarchi2009). Further, as the researchers have assumed that the 4C signature is a de facto reflection of maize, this may have resulted in the systematic neglect of the potential role of Chenopodiaceae-Amaranthaceae, Cyperaceae or Paniceae plants with 4C photosynthetic pathways and Crassulacean Acid Metabolism plants, such as cacti, which may have entered the human food chain (Cadwallader et al. Reference Cadwallader, Beresford-Jones, Whaley and O’Connell2012).
5) It has proven difficult to locate evidence for maize cultivation near late pre-Hispanic sites. Arroyo Tala Cañada 1 is a notable exception, where a ploughsoil feature containing microfossils of maize leaves was recorded and dated to 1028±40 years BP (Pastor & Berberián Reference Pastor and Berberián2007). No maize pollen grains have been identified in late pre-Hispanic sediments, even when pollen spectra were dominated by crop weeds or other Andean cultivars (Medina et al. Reference Medina, Grill, Fernandez and López2017).
6) The absence of durable and obvious agricultural features, such as terraces or retaining walls, implies low-investment cultivation that did not require significant expenditure of time for subsistence activities compared to foraging, which provided the bulk of the overall diet (Medina et al. Reference Medina, Pastor and Recalde2016).
7) Finally, the absence of diagnostic features on microfossils of potentially cultivable plant species (e.g. quinoa, squash, green bean and now Oxalis sp.) should be interpreted as evidence of plant husbandry involving cultigens and quasi-cultigens, rather than as a simple dichotomy of domestic crops vs wild stands. This reduces the relative contribution of domesticated crops to overall diet, and shows that ancient people were actively transforming and maintaining the landscape for the purpose of growing their food (López et al. Reference López, Medina and Rivero2015).
Conclusion
Use-wear patterns and starch grains identified on the active edge of a pre-Hispanic notched bone tool from Boyo Paso 2 in the Sierras of Córdoba indicate that the tool was used for peeling wild or domesticated Oxalis sp. tubers. The results shows how reliance on morphological criteria alone can be misleading when trying to establish tool function (Perry Reference –2007; Kononenko et al. Reference Kononenko, Torrence, Barton and Hennell2010; Allen & Ussher Reference Allen and Ussher2013). The study also challenges the traditional interpretation of pre-Hispanic peoples as being reliant upon maize through the identification of evidence for plant foods and subsistence activities that typically have poor archaeological visibility. In this respect, food-residue analysis provides robust archaeological evidence of Oxalis sp. tuber processing in the Sierras de Córdoba, and, along with the identification of potatoes and sweet potatoes or manioc (López & Recalde Reference López and Recalde2015), demands a reconsideration of the role that underground plant organs played in late pre-Hispanic subsistence, although the taxonomic status of Oxalis sp. as domestic or wild requires further investigation.
The technological analysis presented here reinforces the suggestion that late pre-Hispanic bone-tool assemblages were dominated by expedient artefacts that could perform different tasks with minimal labour investment (sensu Nelson Reference Nelson1991), including the stripping of Oxalis sp. tubers for food. Moreover, these tools appear to have been fashioned from domestic refuse, such as a camelid scapula, linking them to the needs of the small co-residential groups who occasionally settled in semi-permanent villages. Notched bone tools and tuber remains have not previously been recorded to the late pre-Hispanic Period. Thus, the adoption of a broad-scale foraging and cultivation base from c. 1500 years BP was accompanied by the development of new types of bone tool for processing the plant foods that were now integrated into the pre-Hispanic diet.
Food preparation is a well-documented domestic activity in which late pre-Hispanic people made full use of tools, including ceramic cooking vessels, stone artefacts and grinding tools (Medina et al. Reference Medina, Lopez and Berberián2009, Reference Medina, Pastor and Recalde2016). Tools made of bone, and other organic materials, used in cooking-related activities are limited or have low visibility in the record. At Boyo Paso 2, use-wear and starch-grain analysis suggest that this camelid scapula bone tool must be added to the inventory of kitchen tools. Extending this innovative multi-proxy approach to other archaeological bone tools may shed light on underrepresented dietary components and multi-resource economies, and help move away from outdated models that place unwarranted emphasis on maize as the dominant dietary staple.
Acknowledgements
We are grateful for the financial support from the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET PIP 112-200801-02678) and the Agencia Nacional de Promoción Científica y Tecnológica (PICT-2012-0995). Our thanks also extend to Selena Vitezović, Pablo Demaio, Cecilia Trillo, Aylen Capparelli, Nora Franco, Verónica Lema, Sebastián Pastor, Robin Torrence and Peter White, who provided professional advice, equipment and helped greatly to improve the original article.








