



Introduction
Pathological analyses can be used to reveal a broader picture of the health and existence of a population. The unusual occurrence of diseases in certain areas is particularly interesting, as they may reflect processes of migration and a diffusion of genes. Anaemia is rarely reported in the archaeological material from the Middle Euphrates Valley (more precisely, the Syrian lower Euphrates) (Tomczyk Reference Tomczyk2013). Defined as a pathological deficiency in haemoglobin, anaemia can be either genetic (hereditary) or acquired. The most common hereditary forms of anaemia are sickle cell anaemia (sicklemia) and thalassaemia (of which there are several variations) (Walker et al. Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009).
Acquired forms of anaemia are caused by blood loss and nutritional deficiency, including chronic gastrointestinal infections (Pasvol & Abdalla Reference Pasvol, Abdalla, Guerrant, Walker and Weller1999). Numerous authors stress that the best indicators for haematopoietic diseases are cribra orbitalia (CO)—pitting on the superior wall of the orbit; it is thought to be the result of an expansion in haematopoietic bone marrow—and porotic hyperostosis (PH)—pitting on the outer table of the frontal and/or parietal bones; the bones are thickened by the expanded diploic layer and the outer table overlying the lesion is reabsorbed completely (Keenleyside & Panayotova Reference Keenleyside and Panayotova2006; Walker et al. Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009). Although the association between these two types of lesion is not very strong (in many skeletal collections, CO is the more common type, while in others, PH predominates), their presence in skeletal material is often used to identify cases of anaemia (e.g. Salvadei et al. Reference Salvadei, Ricci and Manzi2001; Walker et al. Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009).
There is some controversy regarding the differential diagnosis of anaemia in archaeological materials without considering other variables such as biochemistry (Ortner & Putschar Reference Ortner and Putschar1985). There are, however, important publications that identify the specific features of different forms of anaemia: Hershkovitz et al. (Reference Hershkovitz, Bruce, Rothschild, Latimer, Dutour, Léonetti, Greenwald, Rothschild and Jellema1997) proposed the osteological list of changes associated with diseases of the hematopoietic system. This means that various types of anaemia may be recognised following the meticulous examination of bones.
Recent analysis of cranial abnormalities in an individual from the Middle Euphrates Valley allows us to trace the presence of thalassaemia in this region during the late Roman period. The geographic position of the Middle Euphrates Valley made this region a ‘bridge’ that connected southern Mesopotamia with western Syria. This means that the basis of the livelihood of the local population was not restricted to agriculture and grazing, but included the control of trade and military routes (Postgate Reference Postgate1992). It is therefore possible that the examined individual was not a native of this area, a hypothesis made testable through molecular analysis.
Archaeological context
Tell Masaikh is an archaeological site on the eastern bank of the Euphrates, approximately 6km upstream from the modern Tell Ashara (Figure 1). It was first mentioned as Kar-Ashurnasirpal (Assurnasirpal's Quay; see Masetti-Rouault Reference Masetti-Rouault2008, Reference Masetti-Rouault2010) in Assurnasirpal II's inscription (yearbook) (883–859 BC), a powerful Assyrian king who conquered Syria. In this inscription, we can read the following: “I founded two cities on the Euphrates, one on this bank of the Euphrates [which] I called Kar-Ashurnasirpal [and] one on the other bank of the Euphrates [which] I called Nebarti-Assur” (Grayson Reference Grayson1991: 216). Insufficient information currently exists regarding the dating of the final Assyrian presence at Tell Masaikh (Masetti-Rouault Reference Masetti-Rouault2010). It is known, however, that by the time of the decline of antiquity, when the border between the Roman Empire and the Parthian State (later the state of the Sassanids) ran through northern Mesopotamia, Tell Masaikh was re-occupied (Weiss Reference Weiss1991). In the thirteenth century, a Mongol invasion led by Genghis Khan caused the near total annihilation of the local population. Tell Masaikh has shown no significant indications of urban settlement since the twelfth or thirteenth century, although there is evidence of industrial use, and a Muslim cemetery at the site that is still in use today (Masetti-Rouault Reference Masetti-Rouault2008).

Figure 1. Map of the Middle Euphrates Valley with inset showing the archaeological site.
Materials and methods
During the 2006 excavation season, fragments of skeleton that exhibited pathological changes in the cranial and postcranial bones were found at Tell Masaikh (MK 11G107). The analysed remains were dated to the late Roman period (AD 200–400), indicated both by the grave deposits (e.g. jars) and the east–west orientation of the grave. Moreover, the grave was located in a terrace known to be from the Roman period (Masetti-Rouault Reference Masetti-Rouault2008).
The preserved material included the neurocranium, with a fragment of right temporal, zygomatic and maxillary bone, as well as the body of a mandible. From the postcranial skeleton, the following elements were preserved: proximal epiphysis of the left humerus, both epiphyses of the left ulna and radius, fragments of metacarpals and phalanges from both of the hands, diaphysis and distal epiphysis of the right femur, both epiphyses of the right fibula, right patella, elements of ribs, the sternum, the right scapula and both clavicles (Figure 2).

Figure 2. Diagram of skeleton with preserved areas of bones indicated by shading (drawing by J. Tomczyk).
The sex of the individual was determined on the basis of a morphological assessment of the skull (White & Folkens Reference White and Folkens2000). The medial epiphyses of the clavicles were fully fused; there are, however, still faint signs that remain visible after this union (Walker & Lovejoy Reference Walker and Lovejoy1985). Using these criteria, it was determined that the individual was an adult male (about 30 years old). Radiological examinations were performed at the Radiological Diagnostic Center in Maiadin (Syria). The neurocranium, the right maxilla, the first right rib and the sternum were used in these tests. All of the steps of the molecular analysis were carried out in a laboratory dedicated to ancient DNA (aDNA) work in the Department of Molecular Biology at the Medical University of Lodz (Poland). The teeth (15, 18, 44 and 45—according to the Federation Dentair International numeration) of the individual studied were collected with care and transferred to separate and sterile containers at the archaeological site, delivered to the laboratory and kept frozen until the beginning of the isolation procedure. First, each tooth was mechanically cleaned to remove surface contamination, and then washed in NaClO before finally being washed in alcohol. This was followed by exposure to UV light for 30 minutes per surface, before the samples were powdered in a freezer mill SPEX SamplePrep 6770 and subjected to the DNA isolation procedure. Samples were decalcified with a 0.5M (pH 8) EDTA solution for 48 hours, followed by incubation with proteinase K and PTB for the next 2 hours at 560°C. The obtained solution was then subjected to DNA isolation via the MagNA Pure Compact Nucleic Acid Purification System (Roche) according to the procedures listed in the manufacturer's manual.
Multiple negative controls for all of the steps of the procedure were included. Each of the four analysed teeth were powdered separately by two molecular biologists exhibiting different mtDNA haplogroups (U5 and C). Two independent extractions and the sequence analysis from the different teeth were carried out according to Winters et al.’s (Reference Winters, Barta, Monroe and Kemp2011) protocol. Obtained mtDNA data were compared with the haplotypes of the people involved (archaeologist and two molecular biologists) to test for sample contamination. All of the equipment, including a laminar flow hood (Heraeus) and other work surfaces were decontaminated under UV and with DNA-ExitusPlus.
The hypervariable regions (HVR I and HVR II) of the mtDNA control region were amplified as four overlapping fragments using the following primer pairs:
-
• L16112 (5’-CGTACATTACTGCCAGCC-3’)
-
• H16262 (5’-TGGTATCCTAGTGGGTGAG-3’)
-
• L16251 (5’-CACACATCAAC TGCAACTCC-3’)
-
• H16380 (5’-TCAAGGGACCCCTATCTGAG-3’)
-
• L65 (5’-GCATTTGGTATTTTCGTCTGG-3’)
-
• H156 (5’-TTGAACGTAGGTGCGATAAAT-3’)
-
• L141 (5’-CGCAGTATCTGTCTTTGATTCC-3’)
-
• H256 (5’-CTGTGTGGAAAGTGGCTGTG-3’)
These yielded products of 186, 171, 130 and 156bp, respectively. PCRs (polymerase chain reactions) were performed in 25µl, including 2–4µl of the sample extract, and standard reagents with Taq Gold (Applied Biosystems) for 38 cycles at an annealing temperature of 540°C for HVR I, and 570°C for HVR II. Negative controls accompanied each step of the PCR reaction in order to monitor the presence of contaminating DNA. Approximately 4µl of each PCR product was visualised on a silver-stained 10% polyacrylamide gel to confirm amplification efficiency. Amplicons were cleaned on a spin column (Clean-up, A&A Biotechnology), and then extended using the BigDye 3.1 termination-ready reaction mix. Each sequence reaction (20μl) contained 4μl of BigDye mix, 30ng of primer and 50–70ng of product.
The cycling conditions used were as follows: initial denaturation at 950°C for 5 minutes, followed by 38 cycles at 950°C for 30 seconds, 560°C for 8 seconds and 600°C for 4 minutes. The extended products were cleaned up using spin columns (ExTerminator, A&A Biotechnology), dried in a Speed-Vac system, re-suspended in 20μl of deionised formamide and analysed on an ABI Prism 310Genetic Analyser. The sequences were edited using BioEdit and MEGA 4: Molecular Evolutionary Genetics Analysis, and the haplogroups were determined by using two online databases: mtDNAmanager and HaploGrep.
Results
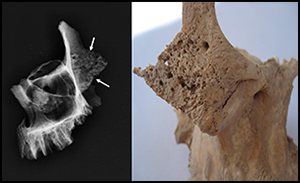
During the macroscopic assessment, CO was detected on the preserved right orbital roof, and was classified as ‘cribrotic type’ Nr 2 (small, as well as large and isolated foramina) using the revised version of Nathan and Haas's (Reference Nathan and Haas1966) and Stuart-Macadam's criteria (Reference Stuart-Macadam1985). PH was present on the neurocranium, affecting the area of the frontal bones near the coronal suture and the entire area of the parietal bones (Figure 3). The surface of the right temporal bone in the vicinity of the tympanic plane was irregular and canalised (Figure 4). The mandibular fossa showed subtle periosteal reactions and proliferation of new bone. Significant changes were found on the right zygomatic bone: the macrostructure was strongly perforated, with evidence of thinning of the cortical bone and resorption of the trabecular bone (Figure 5). Such large macroscopic changes were not visible on the maxilla, although subtle proliferation of the new bone was visible around the infraorbital foramen. Further changes were observed on the body of the sternum, which was thickened in the inferior part. The ribs, especially the right first rib, were enlarged, with particular reference to the posterior and lateral parts.

Figure 3. Superior view of skull with inset showing porosity; this is due to exposure of the marrow rather than taphonomic processes (photograph by J. Tomczyk).

Figure 4. Lateral view of the right temporal bone; arrows indicate granular osteoporosis (photograph by J. Tomczyk).

Figure 5. Anterior view of the zygomatic bones and oblique films of the maxilla and zygomatic bones; the features of granular osteoporosis are present in both bones, and are most pronounced in the hypertrophied body of the zygomatic bone (arrows); there is a remodelling of the maxillary sinus walls, with the reduction of its airspace (images by J. Tomczyk and P. Palczewski).
The radiographic analysis showed expansion of the diploic space of the calvarial bones, with thinning of the outer table and features of granular osteoporosis. An increase in skull-vault thickness, as the result of an increase of the diploic layer of bone, is a feature seen in many haematological lesions. The ratio of compact to diploic tissue in the frontal bone, calculated according to the methodology described by Stuart-Macadam (Reference Stuart-Macadam1987), was 1:4.2. The changes were most prominent in the parietal bones, where an area of what has been called ‘hair-on-end’ (Stuart-Macadam Reference Stuart-Macadam1987) phenomenon was present (Figure 6). The occipital bone was not affected. Granular osteoporosis was also present on the facial bones, especially within the body of the zygomatic bone, with its expansion and decreased number of trabeculae. Some of the trabeculae were hypertrophied. The walls of the maxillary sinus were expanded, leading to the reduction of airspace (Figure 5). The thickening of the sternum and the right first rib proved to be due to the expansion of the marrow space, with a subtle endosteal scalloping and periosteal new bone formation (calcified, uninterrupted periosteal reactions); the first rib showed the ‘rib-within-a-rib’ appearance, as with Lawson et al. (Reference Lawson, Ablow and Pearson1981) (Figure 7).

Figure 6. Lateral film of the skull, showing: a) the neurocranium, except for the occipital bone, illustrates the widening of the diploic space with increased bone porosity and thinning of the outer table, which is most pronounced in the parietal bones; b) the magnification of the parietal bone depicts an area of the ‘hair-on-end’ appearance with vertical striations crossing the thinned outer table; the white arrow indicates the outer table; the black arrow, the inner table; and the arrowheads point to the area of ‘hair-on-end appearance’ (image by P. Palczewski).

Figure 7. Lateral film of the sternum (A) and anteroposterior film of the first right rib (B); there are calcified, uninterrupted periosteal reactions on the anterior and posterior surfaces of the body of the sternum and along the widened first rib, giving the ‘rib-within-a-rib’ appearance (arrows) (image by P. Palczewski).
It is well established that in the average cell, mitochondrial DNA (mtDNA) is represented by hundreds of molecules in contrast to only one nuclear molecule. This fact, in addition to a number of agents that degrade the chemical structure of DNA, strongly limits the possibility of isolating nuclear DNA from skeletal material. Nevertheless, we attempted to identify thalassaemia genetically by identifying genes that code for a and b chains of the haemoglobin molecule. We were unsuccessful because it was impossible to obtain a sufficient amount of amplifiable nuclear DNA in order to confirm the presence of one of the numerous mutations (more than 120 for α-thalassaemia and more than 250 for β-thalassaemia) responsible for the disease phenotype (Filon et al. Reference Filon, Faerman, Smith and Oppenheim1995). We did, however, successfully amplify mtDNA and establish the HVR sequence-related haplotype, which shows the following differences from CRS: 16223T, 16266T, 16289G and 73G. As the site itself is located in a warm and arid region where conditions favour the chemical degradation of DNA molecules, we have taken special measures to authenticate analysed sequences. Applied procedure included the analysis of two separate DNA extracts from each of the four teeth. As a result, each extract produced the same sequence data, which were totally different from those of the three people involved in the extraction process. Attempts were made to establish the haplogroup of the individual using online databases. While mtDNAmanager (Lee et al. Reference Lee, Song, Ha, Cho, Yang and Shin2008) was not able to evaluate the probable haplogroup, the HaploGrep (Kloss-Brandstatter et al. Reference Kloss-Brandstatter, Pacher, Schonherr, Weissensteiner, Binna, Specht and Kronenberg2010) database showed M4b1 (previously M65a) as the most probable. It is noteworthy that the mutation 16311, rooting to the M4b1 haplogroup, was not identified in the HVR of the studied individual, yet it is most probably the effect of a reverse mutation at one of the sites in the mtDNA sequence that is sensitive to mutation (‘hotspots’), of which an example appears in an individual living today in Tibet (Kong et al. Reference Kong, Sun, Wang, Zhao, Wang, Zhong, Hao, Pan, Wang, Cheng, Zhu, Wu, Liu, Jin, Yao and Zhang2011).
Discussion
The analysis of the individual from Tell Masaikh reveals a complex picture that seems best explained by the malfunction of the haematopoietic system. Many authors stress that the best indicators of haematopoietic diseases are found in the skull, which is partly due to the regression of the red marrow from the extra-axial skeleton with age (e.g. Stuart-Macadam Reference Stuart-Macadam1985). In our case, we observed CO and PH, which affected both the calvarial and facial bones.
Both CO and PH are produced by the expansion of the diploë of the skull in response to marrow hypertrophy (Ponec & Resnick Reference Ponec and Resnick1984). The significance of both lesions is unclear (Walker et al. Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009). As epidemiological studies show that CO and PH may arise in response to iron-deficiency anaemia (Oxenham & Cavill Reference Oxenham and Cavill2010), they are commonly used by anthropologists as an indicator for the assessment of the health and nutritional status of historical populations (Stuart-Macadam Reference Stuart-Macadam1985). On the other hand, there are numerous studies relating both lesions to cases of genetic anaemia in the Mediterranean region (Salvadei et al. Reference Salvadei, Ricci and Manzi2001; Keenleyside & Panayotova Reference Keenleyside and Panayotova2006). The fact that osteological material is often fragmented or even destroyed makes the differential diagnosis between acquired and genetic anaemia in palaeopathology studies difficult (Lewis Reference Lewis2012). Another difficulty arises from the clinical evidence, which suggests that both CO and PH often have multifactorial aetiologies (Lagia Reference Lagia, Eliopoulos and Manolis2007; Walker et al. Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009). Some authors suggest, however, that the intensity and distribution of skeletal changes may help in differentiating between types of anaemia (Hershkovitz et al. Reference Hershkovitz, Bruce, Rothschild, Latimer, Dutour, Léonetti, Greenwald, Rothschild and Jellema1997).
Analysis of the Tell Masaikh individual suggests that thalassaemia is evidenced by the presence of the ‘hair-on-end’ phenomenon. This sign, referring to the vertical striations in the calvarial bones that appear to extend behind the outer table, arises following expansive marrow hyperplasia (which expands beyond the natural limitations of the bone), detaches and lifts the periosteum (Resnick Reference Resnick2002). It is almost never seen in iron-deficiency anaemia, and very rarely (2–8% of cases) seen in sickle cell anaemia (e.g. Bassimitici et al. Reference Bassimitici, Yucel-Eroglu and Akalar1996). On the other hand, the ‘hair-on-end’ phenomenon is very common in individuals with thalassaemia (Hershkovitz et al. Reference Hershkovitz, Bruce, Rothschild, Latimer, Dutour, Léonetti, Greenwald, Rothschild and Jellema1997; Walker et al. Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009).
Another finding suggestive of thalassaemia is apparent in the facial bones (Stuart-Macadam Reference Stuart-Macadam1987; Hershkovitz et al. Reference Hershkovitz, Bruce, Rothschild, Latimer, Dutour, Léonetti, Greenwald, Rothschild and Jellema1997). Many thalassaemic patients display changes in the maxillary region (Resnick Reference Resnick2002). In addition to the reduction of sinus airspace caused by expanded walls, maxillary alternations may also produce lateral displacement of both the orbits, leading to hypertelorism, and of the alveolar processes and dental structures, resulting in malocclusion of the jaw (Resnick Reference Resnick2002). As substantial fragments of facial bone were missing from the Tell Masaikh individual, we were not able to observe typical deformities such as hypertelorism. There were, however, features of granular osteoporosis, bone remodelling and periosteal new bone formation evident in the preserved fragments. Changes observed in the sternum and the ribs (thickening and periosteal reactions) lack specificity (Hershkovitz et al. Reference Hershkovitz, Bruce, Rothschild, Latimer, Dutour, Léonetti, Greenwald, Rothschild and Jellema1997), but they do reflect the same ‘hair-on-end’ phenomenon—the proliferation of bone marrow beyond the natural limitations of the bone, with the detachment of the periosteum and subsequent new bone apposition. The differential diagnosis of PH includes hypovitaminoses, caused by deficiencies in one or more vitamins.
Scurvy may lead to CO and PH of the cranial and flat bones, which is most probably related to a chronic bleeding from fragile vessels and a hypertrophy of reparative vessels (Brickley et al. Reference Brickley and Ives2006). Contrary to genetic anaemias, marrow hyperplasia is unusual for scurvy, and should be expected where anaemia is also a prevalent cause of death (Walker et al. Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009; Brown & Ortner Reference Brown and Ortner2011). Scurvy is less common in adults than in children, in whom it affects the growing parts of the skeleton. In the postcranial skeleton of children, a widening of the metaphyses and ribs at the costochondral junctions may be observed (Brickley & Ives Reference Brickley and Ives2006; Brown & Ortner Reference Brown and Ortner2011)—none of these characteristics were present in the studied individual.
Megaloblastic anaemia should be considered when the features of bone marrow hyperplasia are encountered in infants, but it is very uncommon for B12 deficiency to produce skeletal changes in adults (Resnick Reference Resnick2002; Walker et al. Reference Walker, Bathurst, Richman, Gjerdrum and Andrushko2009). Overall, the aforementioned findings make it very probable that the individual, whose remains were recovered from Tell Masaikh, suffered from thalassaemia.
Thalassaemia belongs to the group of disorders known as the haemoglobinopathies, in which the cause of the disease is the reduced or imbalanced synthesis of globin-chains (alpha (α) or beta (β)), resulting in a reduced haemoglobin content and ability to transport oxygen (Hilliard & Berkow Reference Hilliard and Berkow1996). The division of thalassaemia is related to the damage of either the α or β chains. Homozygous individuals with α-thalassaemia die at the foetal stage because their bodies do not synthesise normal haemoglobin, but heterozygous individuals show no signs of pathology. β-thalassaemia is an autosomal recessive disorder characterised by microcytosis and haemolytic anaemia. Although the literature refers to both of these types of thalassaemia, cases of α-thalassaemia are rare in practice (Hilliard & Berkow Reference Hilliard and Berkow1996). Thus, our case is more probably β-thalassaemia than α-thalassaemia.
The terms thalassaemia major, minor and intermedia all relate to defects in the synthesis of β-haemoglobin, and describe one sequence of the disease as well as its symptoms. In homozygous individuals, the severe disease thalassaemia major is present, which causes high mortality among infants and children, but is quite rare in adults. There is hence little chance of finding this type of thalassaemia in skeletal material. The most prominent changes of thalassaemia major are observed in the skull (e.g. Hershkovitz et al. Reference Hershkovitz, Bruce, Rothschild, Latimer, Dutour, Léonetti, Greenwald, Rothschild and Jellema1997; Resnick Reference Resnick2002). The expansion of the diploë is frequent, leading to a thickening of the vault area. Hyperplasia of the compact bone causes a reduction in the pneumatised parts of the frontal and temporal bones. The expansion of the facial bones (especially the maxilla) leads to classic facial deformities and reduces the volume of the maxillary sinuses.
In adults (heterozygotes), the milder form (thalassaemia minor) is usually present, which does not leave clear traces on bone tissue (Lagia et al. Reference Lagia, Eliopoulos and Manolis2007; Lewis Reference Lewis2012). Finally, thalassaemia intermedia is extremely heterogeneous, and encompasses a wide spectrum of phenotypes, ranging in severity from severe anaemia with bone changes (similar to thalassaemia major) to moderate lesions without skeletal abnormalities (Lagia et al. Reference Lagia, Eliopoulos and Manolis2007). People with this type of thalassaemia can survive to later life. Lagia et al. (Reference Lagia, Eliopoulos and Manolis2007) suggest that thalassaemia intermedia is the type most commonly encountered in antiquity. Likewise, we think that the individual from Tell Masaikh most probably suffered from this type of disease.
Many authors have observed a similar pattern in the geographic distribution of thalassaemia and malaria, as the former has a protective effect against Plasmodium infection (e.g. Hershkovitz & Edelson Reference Hershkovitz and Edelson1991; Keenleyside & Panayotova Reference Keenleyside and Panayotova2006). The Tell Masaikh area, located 500km away from the Mediterranean Basin, has never been conducive to the development of malaria. Such areas are restricted to northern (Al Hasakah) and western (the area around Aleppo) Syria, close to the borders with Turkey, Lebanon or Israel (Filon et al. Reference Filon, Faerman, Smith and Oppenheim1995). Climate conditions in the Middle Euphrates Valley have not changed significantly in the last 6000 years (Kirleis & Herles Reference Kirleis and Herles2007). Due to the hydrological and climatic conditions (low average annual rainfall: 100–150mm per year), the Middle Euphrates Valley was generally described as a steppic desert. It is very probable, however, that the valley was not an area where thalassaemia commonly occurred during the late Roman period, when this individual lived.
It is worth emphasising that the Tell Masaikh individual lived during the late Roman period, at the time when northern Mesopotamia was on the boundary between the Roman Empire and the Parthian (later Sassanid) State. During this period, the Middle Euphrates Valley began to play the role of the main military, and trade (in times of peace), route between the two rival empires. Events such as Trajan's expeditions (113–117 AD), or the capture of Dura Europos by the Romans (165 AD), a city less than 40km away from Terqa (see Figure 1), are only some examples of the hostilities between the two empires. It is therefore possible that the individual from Tell Masaikh was not a native of this area. A similar phenomenon is observed in Britain, where genetic anaemia was not endemic and its presence is argued to reflect migration from other regions (Lewis Reference Lewis2012).
Most of the common, contemporarily observed branches of mtDNA in the Near East belong to the super-haplogroups M and N (Thangaraj et al. Reference Thangaraj, Chaubey, Kivisild, Reddy, Singh, Rasalkar and Singh2005). The present-day Near Eastern gene pool most frequently exhibits the haplogroups HV (30%), H (25%), U (22%), J (10%) and T (9%), and, at a lower frequency, K, U1, U2, U3, U4, U5, U7, N1b, I, W and X (Richards et al. Reference Richards, Macaulay, Hickey, Vega, Sykes, Guida, Rengo, Sellitto, Cruciani, Kivisild, Villems, Thomas, Rychkov, Rychkov, Rychkov, Gölge, Dimitrov, Hill, Bradley, Romano, Calì, Vona, Demaine, Papiha, Triantaphyllidis, Stefanescu, Hatina, Belledi, Di Rienzo, Oppenheim, Nørby, Al-Zaheri, Santachiara-Benerecetti, Scozzari, Torroni and Bandelt2000). There is not much data, however, regarding ancient mtDNA sequences from the Near East, probably because of the difficulties in extracting amplifiable fragments from human remains. Nevertheless, a few publications present data on mtDNA isolated from skeletons unearthed in arid and semi-arid regions, such as Syria (Tomczyk et al. Reference Tomczyk, Jędrychowska-Dańska, Płoszaj and Witas2011; Witas et al. Reference Witas, Tomczyk, Jędrychowska-Dańska, Chaubey and Płoszaj2013) and Egypt (Graver et al. Reference Graver, Molto, Parr, Walters, Praymak and Maki2001). A specific haplotype found in the Tell Masaikh skeleton, considered to be the M4b1 haplogroup with a mutation at position 16289G, was identified among contemporary inhabitants of the Indian subcontinent. Haplogroup M4b was also determined in a few cases from the Arabian Peninsula and the Syrian region (Abu-Amero et al. Reference Abu-Amero, Larruga, Cabrera and Gonzalez2008). The observed haplotypes were, however, distinct from that of the Tell Masaikh individual identified herein, which is, in turn, different both to those already found in aDNA from Syria (Fernandez Reference Fernandez2005) and among the genes of the modern Syrian population (Abu-Amero et al. Reference Abu-Amero, Cabrera, Larruga, Osman, Gonzalez and Al-Obeidan2011). All of these suggest that the analysed individual was probably a newcomer to the area, or a local with ancestors whose maternal lineage originated in Asia, most probably in the Indian Peninsula, where this specific haplogroup can still be identified (Kong et al. Reference Kong, Sun, Wang, Zhao, Wang, Zhong, Hao, Pan, Wang, Cheng, Zhu, Wu, Liu, Jin, Yao and Zhang2011). Their origins may well confirm theories of trade and military contacts between the Indian Peninsula and Syria. One example of these highly active trade relations was the Silk Route, one of the oldest trade routes connecting the Eastern and the Western worlds (Whitfield & Sims-Williams Reference Whitfield and Sims-Williams2004). Palanichamy et al. (Reference Palanichamy, Mitra, Debnath, Agrawal, Chaudhuri and Zhang2014) indicate that during the first centuries AD, trade between Rome and southern India intensified, and many goods (pearls, ivory, textiles) were imported from this region. These trade routes would have also served as routes for the migration of peoples and diseases.
Conclusion
This paper has provided evidence for a probable case of thalassaemia in the Middle Euphrates Valley. Despite difficulties in confirming a diagnosis of genetic anaemia in palaeopathology studies, skeletal changes in the Tell Masaikh individual (especially the widening of the diploë exhibiting ‘hair-on-end’ phenomenon and the remodelling of the facial bones) suggest evidence of a malfunction of the haematopoietic system, most probably as a consequence of thalassaemia intermedia. Further molecular analyses were prompted following the observation of macroscopic evidence of these alterations. The results of those studies show signs of a disease that is uncommon in this particular area. Evidence of this disease strongly suggests the migration of this individual and perhaps others to the region from an area outside it. The identification of a haplogroup indicating an eastern, and not Mediterranean, origin for the Tell Masaikh individual was unexpected, but may explain this unusual occurrence of thalassaemia.
Acknowledgements
The authors would like to thank Maria Grazia Masetti-Rouault (École des Hautes Études, Paris) who has given much advice and additional information. Many thanks are also dedicated to the medical staff from Maiadin (Syria) who helped perform X-ray tests of the materials. Last but not least, we would like to thank Theya Molleson (Natural History Museum, London) for her valuable comments and advice. This research was financed by scientific projects N303 319837 and N109 286737 from the Polish Ministry of Science and Higher Education.







