


Introduction
A principal aim of ecology is to understand the links between species and habitats. This is hard to achieve, however, when organisms or the regions they inhabit are remote or otherwise difficult to observe. One group of such animals that are particularly abundant in the Southern Ocean are the surface feeding, pelagic seabirds. Because these birds routinely range hundreds to thousands of kilometres from their breeding colonies in search of food it has become the norm to study their foraging ecology using remote sensing technology. Typically the approach is as follows: miniature devices attached to the birds record their movements and behaviour, while satellites simultaneously measure the dynamic biological and physical properties of the wide areas of ocean in which they forage (e.g. Wakefield et al. Reference Wakefield, Phillips, Trathan, Arata, Gales, Huin, Robertson, Waugh, Weimerskirch and Matthiopoulos2011). As such, the habitats of pelagic seabirds (and many other wide-ranging higher marine predators) are frequently described in terms of superficial properties of the ocean measured using satellite remote sensing. These include sea surface temperature, sea surface height, chlorophyll a and so on, as well as their derivatives, including primary productivity, mesoscale turbulence etc. (reviewed by Wakefield et al. Reference Wakefield, Phillips and Matthiopoulos2009).
Despite the insights gained using this approach, the availability of food at or near the surface of the ocean is not solely dependent on processes readily measurable from space. In particular, upward benthic-pelagic coupling can have a major influence on the productivity of shelf sea ecosystems (Smith et al. Reference Smith, Mincks and DeMaster2006). It follows that depth and substrate type could also influence the distribution of surface-feeding or shallow diving seabirds even if they cannot access the seabed itself. Though it has long been recognized that bathymetry can play an important role in defining pelagic seabird habitat suitability, variability in substrate type and small-scale bathymetry have rarely been explicitly considered in analyses of habitat preference of surface-feeding, pelagic seabirds. This is despite the fact that mid-water and epipelagic fisheries for the same species preyed upon by seabirds often target waters overlaying particular substrate types or bathymetric features (Greenstreet et al. Reference Greenstreet, Spence, Shanks and McMillan1999). In Antarctic waters, examples of upward benthic-pelagic coupling caused by such features include currents interacting with complex topography, resulting in nutrient transport from the seabed into the water column (Sedwick et al. Reference Sedwick, DiTullio and Mackey2000, Atkinson et al. Reference Atkinson, Whitehouse, Priddle, Cripps, Ward and Brandon2001), and euphausiids feeding on accumulations of biogenic detritus on the seabed and then migrating to the surface, where they are consumed by higher predators (Gutt & Siegel Reference Gutt and Siegel1994, Smith et al. Reference Smith, Mincks and DeMaster2006). It is possible therefore, that seabed characteristics, such as substrate type, may in part determine habitat quality for surface-feeding pelagic seabirds. In turn, this implies that substrate may be a useful predictor of these species’ distribution.
The black-browed albatross Thalassarche melanophris (Temminck, 1828) is one of the world's largest and most wide-ranging pelagic seabirds. It breeds on sub-Antarctic islands, with population centres in the Falkland Islands (c. 414 000 pairs), southern Chile (c. 123 000 pairs) and South Georgia (c. 74 000 pairs) (Wakefield et al. Reference Wakefield, Phillips, Trathan, Arata, Gales, Huin, Robertson, Waugh, Weimerskirch and Matthiopoulos2011). During foraging trips, breeding black-browed albatrosses show a marked preference for shelf and shelf slope habitats (Cherel et al. Reference Cherel, Weimerskirch and Trouve2000, Wakefield et al. Reference Wakefield, Phillips, Trathan, Arata, Gales, Huin, Robertson, Waugh, Weimerskirch and Matthiopoulos2011), where they principally catch fish, cephalopods and crustaceans (Reid et al. Reference Reid, Croxall and Prince1996, Cherel & Klages Reference Cherel and Klages1998, Cherel et al. Reference Cherel, Weimerskirch and Trouve2000). During non-breeding periods birds forage in temperate neritic, and in the case of some populations, oceanic waters but little is known of their diet during these stages (Phillips et al. Reference Phillips, Silk, Croxall, Afanasyev and Bennett2005). Their limited diving capabilities mean that they capture prey either at or within 5 m of the surface (Prince et al. Reference Prince, Huin and Weimerskirch1994). Despite this constraint, they frequently consume benthic or semi-pelagic organisms, such as nototheniid and channichthyid icefish (Cherel & Klages Reference Cherel and Klages1998, Cherel et al. Reference Cherel, Weimerskirch and Trouve2000). In some instances, black-browed albatrosses obtain such prey by scavenging discards from fishing vessels, which in some areas can make up a significant proportion of their diet (Reid et al. Reference Reid, Croxall and Prince1996). However, these prey may also be consumed by birds in the absence of fishing activity (Cherel et al. Reference Cherel, Weimerskirch and Trouve2000). Therefore, although the mechanisms by which benthic prey become accessible at the surface are unclear, it is possible that black-browed albatross distribution may be influenced by that of particular benthic habitats.
The need to understand black-browed albatross habitat use, especially where this overlaps with commercial fishing grounds, has become more than academic over recent years. This is because incidental bycatch by both longline and trawl fisheries is leading the species into a potentially terminal decline, such that since 2002 it has been formally recognized as endangered (www.iucnredlist.org, accessed May 2011). During the breeding season, the foraging areas used by birds from the South Georgia population, on which our study focuses, are also used by mid-water trawlers targeting Antarctic krill Euphausia superba (Dana, 1852) and mackerel icefish Champsocephalus gunnari (Lönnberg, 1905) (Kock Reference Kock2001). The potential for overlap between these trawlers and black-browed albatrosses is heightened because krill are a major component of the diet both of this species and mackerel icefish (Kock et al. Reference Kock, Wilhelms, Everson and Groger1994, Cherel & Klages Reference Cherel and Klages1998).
Until recently, little was known about the benthic habitats surrounding South Georgia. However, analysis of high-resolution multibeam swath bathymetry data collected over the shelf has now revealed a series of banks and ridges, rising from a few metres to > 75 m above the surrounding seabed (Graham et al. Reference Graham, Fretwell, Larter, Hodgson, Wilson, Tate and Morris2008, Fretwell et al. Reference Fretwell, Tate, Deen and Belchier2009). These features, which are concentrated along the shelf edge, especially to the north and west of the islands, are thought to comprise terminal moraines formed during the last glacial maximum at the seaward margins of once extensive ice sheets (Graham et al. Reference Graham, Fretwell, Larter, Hodgson, Wilson, Tate and Morris2008). As such, they probably form a substrate of unconsolidated cobbles and boulders, that support a richer fauna than surrounding bedrock habitats (Barnes Reference Barnes2008).
Here, we use seabed substrate data as a habitat proxy, and highly accurate GPS tracking to test the hypothesis that black-browed albatrosses forage in a non-random manner with respect to benthic habitats on the South Georgia shelf. We then use auxiliary logger data to examine variability in prey capture and activity with respect to habitat type. In addition, we investigate potentially confounding interactions between tracked birds and fishing vessels operating within the study area. We discuss our findings in the context of possible mechanisms of upward benthic-pelagic coupling and their implications for understanding pelagic seabird-habitat relationships and seabird-fishery conflicts in the Southern Ocean and beyond.
Materials and methods
Context, definitions and instrumentation
Fieldwork was carried out at Bird Island, South Georgia (54°00′S, 38°03′W) between January and March 2008. During January, adult black-browed albatrosses at South Georgia are engaged in rearing small, recently hatched chicks. In this period (hereafter ‘brood-guard’) parents alternate between foraging at-sea and remaining at the nest to brood and feed their chicks (i.e. one or other parent always attends the chick). Brood-guard foraging trips are typically short (mean duration from Bird Island c. 50 hours, mean maximum distance from the colony c. 300 km) and mainly to neritic or shelf break waters surrounding South Georgia but they may also be directed to other local oceanic waters (Phillips et al. Reference Phillips, Silk, Phalan, Catry and Croxall2004). From the end of January onwards chicks are large enough to be left unattended and both parents forage at the same time to meet the energetic demands of both themselves and their offspring. During this period (hereafter ‘post-brood’) they continue to make trips to local waters but also travel to more distant patches of neritic, shelf break and oceanic habitat (mean trip duration c. 75 hours, mean maximum distance from the colony c. 470 km) (Phillips et al. Reference Phillips, Silk, Phalan, Catry and Croxall2004).
During fieldwork, we deployed GPS loggers programmed to record positions every 5, 15 or 30 minutes on a total of 49 black-browed albatrosses. These were either Mini-GPSlogs (29 g, ‘earth & OCEAN Technologies’, Kiel, Germany), GPS-DLs (38 g, New Behaviour AG, Zurich, Switzerland) or GPS-RF tags (36 g, e-obs GmbH, Munich, Germany). Ten birds were also equipped with a Stomach Temperature Archival Unit (STAU) (MiniTemp, 33–37 g, ‘earth & OCEAN Technologies’, Kiel, Germany) set to log the temperature in the bird's proventriculus at intervals of 1, 2 or 20 seconds. These data were later analysed to determine the timing of prey ingestion and the approximate mass of prey items (see Catry et al. (Reference Catry, Phillips, Phalan, Silk and Croxall2004) for details of deployment and recovery techniques). All birds also carried a Mk7 combined light and immersion logger (4 g, British Antarctic Survey, Cambridge, UK) and during post-brood, to facilitate recapture, a VHF transmitter (17 g, Sirtrack, Havelock North, New Zealand). These devices were attached to plastic bands placed around the tarsus of the birds’ legs. The Mk7 records the time of transition (3 sec resolution) between wet/dry states providing the new state lasts more than 6 sec. A state change of less than 6 sec is ignored. Of the birds not equipped with a STAU, seven were equipped with a second small GPS logger set to record locations every 1–5 seconds (8 or 23 g, Technosmart, Rome, Italy, data not presented here). The mass of birds was measured prior to instrument attachment. Median total mass of instruments and attachment material was 68 g or 2% of body mass (range 46–105 g, 1–3%). To establish whether tracking had any deleterious effects on breeding performance, nests of tracked birds were matched with a nearby control, and both nests visited regularly until the end of the breeding season to establish whether the chicks fledged.
Calibration of the STAUs, as well as feeding trails (data not presented here), was performed following Catry et al. (Reference Catry, Phillips, Phalan, Silk and Croxall2004). Briefly, three non-breeding adult black-browed albatrosses were captured in the colony and taken to a purpose built wooden holding pen (floor area 25 m2, height 2 m). While birds were in the holding pen they did not attempt to escape or fly but spent most of their time sitting on the ground. After being left for 3–4 h to become accustomed to their surroundings the birds were induced to swallow a STAU. Following a period of either 2.5 h (one bird) or 12 h (two birds) they were then fed two meals of mackerel icefish, two meals of the squid Illex argentinus (Castellanos, 1960), three meals of Antarctic krill Euphausia superba (Dana, 1850) and one meal of the mantle tissue of the Cnidarian Periphylla periphylla (Péron & Lesueur, 1810). Meal masses were between 1.3 and 105.8 g. In addition birds ingested 100 ml of seawater. This and the meals were all at a known temperature of between -0.3 and 3.4°C. An interval of at least an hour was left between meals in order to allow the temperature of the proventriculus, meal and STAU to equilibrate. The birds did not show any signs of undue stress during these procedures, did not attempt to regurgitate any meals and settled down immediately after being fed. At the end of the feeding trials STAUs were recovered and the birds were released back into the colony. The time elapsed between capture and release for the three birds was 28, 40 and 41 h respectively.
Fisheries data
The waters around South Georgia fall within the Fisheries Conservation and Management Zone (FCMZ) of the South Georgia and South Sandwich Islands and are subject to conservation measures set by the Commission for the Conservation of Antarctic Marine Living Resources (CCAMLR). The Government of South Georgia and the South Sandwich Islands issues licences to a limited number of fishing boats to operate in the FCMZ each year and sets catch limits for different target species. During the study period (January–March) longlining is prohibited in order to mitigate seabird bycatch but a limited amount of mid-water trawling for mackerel icefish and Antarctic krill is permitted. The locations of all sets and hauls made by boats engaged in these fisheries are reported to CCAMLR, along with catch totals. We obtained these data from CCAMLR for the period 2000–08. Illegal fishing is effectively prevented in South Georgia waters by regular fisheries patrols and surveillance by licensed fishing vessels so these data are thought to summarize all fishing activity in the area.
Habitat data
Hereafter we refer to shelf (or neritic) waters as those shallower then 500 m, which is the approximate depth of the South Georgia shelf break. We refer to upper shelf slope waters as those 500–1000 m deep and oceanic waters as those > 1000 m deep. We use the term moraine bank to refer to those features identified as such on the South Georgia shelf by Graham et al. (Reference Graham, Fretwell, Larter, Hodgson, Wilson, Tate and Morris2008). We downloaded high-resolution (150 m cell size) bathymetric data compiled by the British Antarctic Survey (BAS, Cambridge, UK) from single and multi-beam swath bathymetry (Fretwell et al. Reference Fretwell, Tate, Deen and Belchier2009) from http://www.antarctica.ac.uk/bas_research/data/online_resources/sgbd/ (accessed November 2010). We also obtained an ESRI shape file defining the extent of benthic moraine banks on the South Georgia shelf identified by Graham et al. (Reference Graham, Fretwell, Larter, Hodgson, Wilson, Tate and Morris2008) from BAS. This study did not include the shallow waters around Shag Rocks, 270 km to the WNW of Bird Island. Therefore, we had no information on the presence or absence of submarine moraine features in this area or indeed in shelf waters elsewhere that may be utilized by black-browed albatrosses breeding at Bird Island. For this reason we confined our analysis of albatross movements, habitat use and prey ingestion to waters within 120 km of Bird Island (Fig. 1, hereafter the study area). The shape and extent of this area was defined to exclude the Shag Rocks shelf, omit areas of the South Georgia shelf that have poor bathymetric data coverage (Graham et al. Reference Graham, Fretwell, Larter, Hodgson, Wilson, Tate and Morris2008, fig. 2) and reflect the fact that breeding albatrosses are central place foragers (Wakefield et al. Reference Wakefield, Phillips and Matthiopoulos2009). We also excluded waters within 3 km of Bird Island from the study area because albatrosses tended to spend time on the surface of the sea close to the colony. The function of this behaviour is unclear but such rafting by seabirds is not thought to be associated with foraging (Waugh et al. Reference Waugh, Weimerskirch, Cherel and Prince2000).
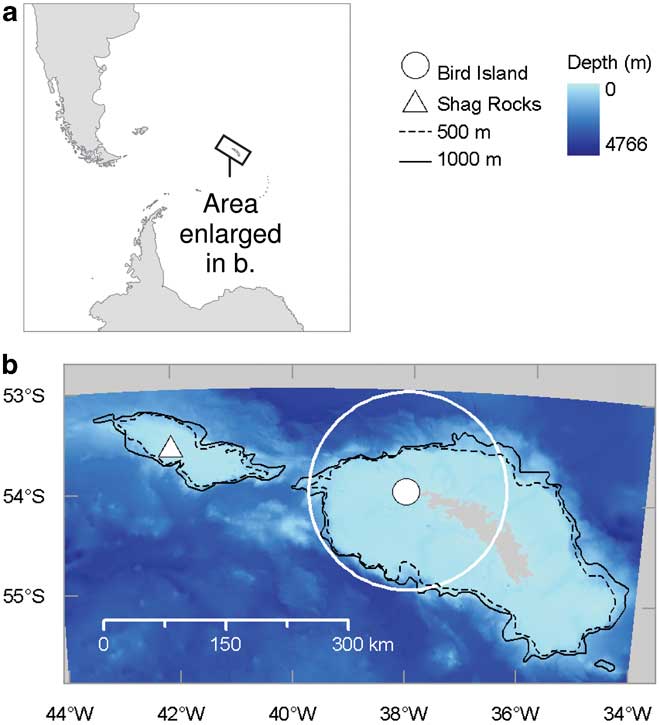
Fig. 1 Location of the study area and places mentioned in the text. High-resolution bathymetric data were not available for areas shaded grey in b.

Fig. 2 Benthic moraine banks on the South Georgia shelf identified by Graham et al. (Reference Graham, Fretwell, Larter, Hodgson, Wilson, Tate and Morris2008), overlaid with the distribution of icefish trawling activity (contours containing 50 and 90% kernel density of all haul locations reported between 2000 and 2008). Numbers indicate moraine banks or groups of banks discussed in the text.
Analysis
We reduced all GPS tracking data to the same temporal resolution (one location every 30 minutes), if necessary, by retaining only every second or every sixth location. In order to test whether black-browed albatrosses used waters overlaying moraine banks more frequently than they would by chance we first compared their use of moraine bank areas to their availability (Johnson Reference Johnson1980). Adopting a case-control design (Aarts et al. Reference Aarts, MacKenzie, McConnell, Fedak and Matthiopoulos2008, Wakefield et al. Reference Wakefield, Phillips, Trathan, Arata, Gales, Huin, Robertson, Waugh, Weimerskirch and Matthiopoulos2011), we regarded GPS points as indicative of a bird's presence at a location. We then generated pseudo-absence locations quasi-randomly as controls for each presence location, at a rate inversely proportional to distance from Bird Island. This resulted in a null model, in which the probability of an albatross using a location is inversely proportional to its distance from the colony (i.e. locations close to the colony are more easily accessible and therefore more probably used by chance than those further away). For each presence location a total of four pseudo-absence locations were generated, this number being chosen to give a reasonable representation of available habitat and to stay within computational limits (Aarts et al. Reference Aarts, MacKenzie, McConnell, Fedak and Matthiopoulos2008). We then modelled the presence/absence of birds as a function of substrate type and depth. The latter covariate was included because moraine banks on the South Georgia shelf tend to occur just landward of the shelf break (Graham et al. Reference Graham, Fretwell, Larter, Hodgson, Wilson, Tate and Morris2008). It is possible that a preference for this feature, like that expressed by black-browed albatrosses foraging from Kerguelen (Cherel et al. Reference Cherel, Weimerskirch and Trouve2000, Pinaud & Weimerskirch Reference Pinaud and Weimerskirch2007), could give the spurious impression that moraine banks were preferred simply because they are adjacent to the shelf break. Models were structured as mixed-effects generalized additive models with a binomial error structure, fitted using the package gamm4 run in the statistical software R. Individual bird was specified as a random effect, and substrate and depth as fixed-effects. Substrate was classified in two ways: firstly, substrate I was defined according to whether locations overlay a moraine bank or not. Secondly, to account for potential local advection of prey associated with the presence of the banks, substrate II was defined according to whether locations lay within a 2 km buffer surrounding the moraine banks.
Four models were considered: models Ia and Ib were fitted to locations of all birds within the study area. We then assumed that birds recorded on the same day as and within 33 km of the locations of icefish hauls, potentially interacted with fishing vessels (the distance of 33 km is based on a maximum trawl time during 2008 of 3 h and the assumption that working trawlers travelled at ≤ 5 knots). We therefore fitted models IIa and IIb to locations from birds during periods when they did not have the potential for interaction with a working trawler.
In order to examine activity patterns with respect to habitat and daylight, bird activity at each tracking location was determined using immersion logger data. The proportion of locations when the loggers were wet was calculated for each bird within each habitat type and for periods of darkness and light. This index was used as an estimate of the proportion of time spent on the water in each category within these habitats. Locations were classified as dark if the sun was < 6° below the horizon (i.e. between the end of evening, and the start of morning civil twilight) or light otherwise. Times of civil twilight were calculated using standard astronomical algorithms.
Analysis of STAU data was carried out using methods described by Wilson et al. (Reference Wilson, Putz, Gremillet, Culik, Kierspel, Regel, Bost, Lage and Cooper1995). Following prey ingestion, proventriculus temperature typically falls precipitously and then rises exponentially (a precipitous drop, exponential rise or PDER event). The timing of each PDER event, and the integral (INT) above the curve of temperature vs time for that event were estimated using the program MultiTrace Temp (Jensen Software Systems, Laboe, Germany). The mass M of food ingested was estimated according to:
where SHC f is the specific heat capacity of the prey item, which was assumed to be 4.0 J g-1 °C-1 (Putz et al. Reference Putz, Wilson, Charrassin, Raclot, Lage, Le Maho, Kierspel, Culik and Adelung1998); T a is the temperature to which the food must be heated; and T f is the temperature of the food. The constant m is defined as:
where E is the energy required to heat the food to the equilibrium temperature of the proventriculus, which can be estimated by:
We estimated m by evaluating the gradient of INT vs E for prey items of known mass ingested by the three black-browed albatrosses during the feeding trials (Wilson et al. Reference Wilson, Putz, Gremillet, Culik, Kierspel, Regel, Bost, Lage and Cooper1995, Catry et al. Reference Catry, Phillips, Phalan, Silk and Croxall2004). For free-ranging birds, T f was assumed to be the equal to the sea surface temperature (SST) at the location where prey items were captured. These locations were estimated by linear interpolation between the GPS locations preceding and following PDER events. T f was then determined from weekly mean Optimally Interpolated SST data, obtained on a 1° grid from the National Oceanic and Atmospheric Administration, Boulder, CO (http://www.esrl.noaa.gov/psd/data/gridded/data.noaa.oisst.v2.html, accessed November 2010). Where n multiple-PDER events overlapped one another, the total meal mass M was calculated and then the mass of individual prey items estimated as M/n.
Results
Fishing activity
Between 2000 and 2008 trawlers targeting icefish concentrated their activity over moraine banks landward of the South Georgia shelf break. Two distinct areas were targeted: the first coinciding with a concentration of banks to the north-west of South Georgia (banks 1 and 2, Fig. 2) and the second a large bank to the north-east of the island (bank 4). During the study period (January–March 2008) four trawlers operated on the South Georgia shelf. Fishing activity was not continuous during this time but was largely carried out by two boats in two non-overlapping bouts. The first boat made 40 hauls between 19 January and 2 February. These were predominantly in an area 35 km north of Bird Island, where no moraine banks have been identified. However, a small number of hauls were also made in the study area over bank 1, north-west of the island. Two further boats made three additional hauls during this period. After a pause of two weeks, fishing resumed on 16 February and continued until 1 March. During this time, one boat carried out 31 hauls, which were predominantly over a single moraine bank in the study area 40 km to the WNW of Bird Island (bank 2). A total of 750t of mackerel icefish was caught during the study period (CCAMLR 2009). No fishing for krill on the South Georgia shelf was reported in 2008.
Albatross habitat use and behaviour
A total of 49 foraging trips made by breeding adult black-browed albatrosses were tracked using GPS loggers. The percentage of chicks successfully fledged by the 49 pairs that included a tracked bird (65%) did not differ significantly from that of the matched controls (61%, χ2 = 0.001, n = 98 pairs, P = 0.971). Furthermore, the fledging success of chicks of tracked birds was not significantly related to instrument mass (generalized linear model with quasibinomial errors, F 1,47 = 1.502, P = 0.224). Foraging trips lasted between 0.6 and 18.7 days (median 3.1 days). The maximum distances from the colony reached by birds ranged from 96–2191 km (median 433 km). Although most trips were to the Shag Rocks or more distant areas, 13 were confined entirely to the South Georgia shelf. During 11 of the latter, ≥ 75% of the birds’ time was spent within the study area. Some birds on longer trips also spent an appreciable amount of time on the South Georgia shelf, rather than commuting rapidly through the area.
Moraine banks in the study area had a gently sloping relief and the maximum gradient of each of the banks had a median value of 6° (range 1–15°). Moraine banks were located in waters with a mean depth of 261 m (range 143–320 m) and on average rose to a maximum of 54 m above the surrounding seabed (range 13–161 m). Within the study area, black-browed albatross spatial usage was highest around the colony and towards the shelf break. A total of 29 birds visited waters overlaying moraine banks and 39 birds visited waters overlaying either the banks or the 2 km buffer zone. Banks 1, 2, 3 and 5 were used most frequently (Fig. 3) but bank 4 and other banks outside the study area were also visited by one or two birds each. Overall, 7% of albatross locations within the study area overlaid moraine banks (ranging from 0–40% for individual birds) and 17% overlaid or were within 2 km of the edges of moraine banks (range for individual birds 0–53%). In comparison, the banks themselves comprised only 2% of the seabed in the study area (or 7% when the 2 km buffer was included). Only 1% of pseudo-absence locations (see Materials and methods) were over the banks, and an additional 2% over the 2 km buffer. Models Ia and Ib, fitted to all tracking data from the study area (49 birds) indicated that moraine banks and moraine banks plus the 2 km buffer were used significantly more frequently than would be expected by chance (Table I). In addition, locations close to the shelf break were used more frequently than expected (Fig. 4).

Fig. 3 Half hourly locations (black dots) of black-browed albatrosses GPS-tracked from Bird Island, South Georgia within the study area (dashed line). a. Locations from all birds tracked (n = 49), and b. locations only from birds that remained at least 33 km from icefish hauls sites (blue triangles indicate locations where the other birds came within 33 km of an icefish haul, remaining symbols are as in Fig. 2).
Table I Fixed effects in mixed-effects Generalized Additive Models of black-browed albatross habitat use in waters between 3 and 120 km from Bird Island, South Georgia. Parameters are expressed in logits (i.e. on the scale of the Generalised Additive Model canonical link function for the binomial distribution). Logits may be transformed to the scale of the response variable according to the formula logit-1(α) = exp(α)/1 + exp(α), where α in the parameter value. SE = standard error, z = the Wald statistic.
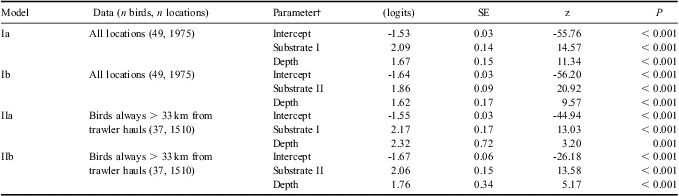
†Substrate I defined as 1 if locations overlay a moraine bank or 0 otherwise. Substrate II defined as 1 if locations overlay a moraine bank plus a 2 km buffer or 0 otherwise.

Fig. 4 Variation in habitat use with depth by black-browed albatrosses within 120 km of Bird Island, South Georgia, predicted by a mixed-effects Generalized Additive Model (model Ia). The y-axis shows the partial residual once the effects of substrate type have been removed and is proportional to habitat use vs availability. Dashed lines indicate estimated 95% confidence intervals. The approximate depth of the South Georgia shelf break is indicated by the vertical dot-dashed line.
Within the study area, during daylight, birds spent little time on the water (median 14%, 1st and 3rd quartiles 8% and 29%, range 0–69%). Conversely, during darkness they spent the majority of their time on the water (median 81%, 1st and 3rd quartiles 58% and 92%, range 0–100%). The activity of birds while in the vicinity of the moraine banks followed this overall pattern and did not differ significantly from their behaviour elsewhere (paired t-test, percentage of locations within the 2 km buffer wet vs percentage of locations outside the 2 km buffer wet, t 38 = -0.586, P = 0.561).
A total of 12 tracked birds came within 33 km of icefish trawlers engaged in hauling. However, across the study period and site as a whole, potential fishery-albatross interactions were rather limited. They occurred almost exclusively in the area of shelf to the north and west of Bird Island, some in the vicinity of banks 1 and 2 (Fig. 3b), during two distinct periods: firstly from 19–31 January and secondly from 21–28 February (Fig. 5). Considering only the 37 birds that did not at any point come within 33 km of an icefish haul, 8% of locations within the study area overlay moraine banks (range for individual birds 0–40%) and 19% overlay or were within the 2 km buffer (individual range 0–53%). Models IIa and IIb indicate that the rate of use of moraine bank habitats by birds which did not interact with fishing boats was higher than expected by chance (Table I).

Fig. 5 Number of GPS-tracked black-browed albatrosses within the study area during each day of the study and the number of those birds that could potentially have interacted with trawlers. Potential interaction was deemed to be possible if a tracked bird came within 33 km of the location of a haul on the day in question.
Prey capture
The value of the constant m (± 95% confidence interval (CI)), estimated following feeding trials with three non-breeding adult black-browed albatrosses, was 0.0083 ± 0.0009°C s J-1 (linear regression, r 2 = 0.95, F 1,19 = 377.8, P < 0.001). Single meal masses estimated for captive birds using this value had a median prediction error of ± 23% of their true value (lower and upper quartiles 11 and 45%, n = 20). STAU data were successfully obtained from seven free-ranging birds (the other two birds regurgitated their STAU at sea and one logger malfunctioned). Four of these birds commuted over the South Georgia shelf to forage in more distant waters (data not shown here), ingesting from three to seven prey items on their way through the study area. Of the remaining three birds, two (#22 and #27) made complex looping trips, foraging primarily over banks 1 and 2 to the west and north-west of Bird Island, returning to the vicinity of the colony on a number of occasions before finally returning to the colony itself (Fig. 6). Bird #22 ingested 20 items within the 2 km buffer around the moraine banks and 18 elsewhere. Bird #27 ingested 63 and 24 items, respectively, in these habitats. The third bird visited the shelf north-east of Bird Island beyond the study area, where it ingested five items, but did not forage over any moraine banks (not illustrated). Birds #22 and #27 were tracked between 14 and 22 March. They are unlikely to have interacted with any fishing vessels during this period, as none were reported to be present in the study area (Fig. 5).

Fig. 6 Detail of single foraging trips made by two black-browed albatrosses (birds #22 and #27) from Bird Island, South Georgia in relation to moraine banks showing the estimated location and mass of prey items taken in foraging events.
The average estimated mass of the144 individual prey items ingested in the study area was 19.7 g (95% CIs 0.3–68.5 g, masses log-normally distributed, Anderson-Darling test A = 2.395, P < 0.001). The mean mass of prey items ingested by birds #22 and #27 within the 2 km buffer around the moraine banks did not differ significantly from those ingested elsewhere (Fig. 7, one-way ANOVA for each bird, F 1,36 = 2.09, P = 0.157; F 1,85 = 1.15, P = 0.286). Considering data from all birds monitored with STAUs, the majority of prey (56% of individual items, 75% by mass) was ingested during daylight. Bird #22 only visited the banks and 2 km buffer during the day and so all prey taken in this habitat was caught in daylight. Bird #27, which visited the banks during both light and darkness, ingested 98% of prey items (78% by mass) in this habitat during daylight. Regurgitates collected from 20 instrumented birds after they returned from foraging indicated that they had fed primarily on krill, followed by fish and cephalopods (49, 40 and 11% of regurgitates by weight respectively). Of these birds, eight foraged primarily in the study area, only one of which regurgitated any cephalopod remains. Regurgitates from bird #22 predominantly comprised unidentified fish remains (95% by mass) and a small amount of krill. Bird #27 exclusively regurgitated krill.

Fig. 7 Estimated mass of prey items ingested by two black-browed albatrosses (birds #22 and #27) from Bird Island, South Georgia caught a. within 2 km of moraine banks, and b. beyond 2 km from moraine banks. Note use of logarithmic size categories.
Discussion
Our tracking data clearly indicate that while on foraging trips in the vicinity of South Georgia, breeding black-browed albatrosses spend more time in waters overlaying moraine banks than expected by chance. By combining data from highly accurate GPS loggers and hydrographic surveys, it was possible to relate the movements of individual wide-ranging marine predators to habitat at a finer scale than has hitherto been possible. Furthermore, although the sample size was low, the information on prey ingestion derived from stomach temperature loggers was invaluable for showing that black-browed albatrosses visiting waters overlaying moraine banks forage actively, in some cases in intensive bouts. Before exploring the reasons for the relationship between benthic habitat and albatross distribution, and its possible implications, we discuss the limitations of our study and some relevant inferences about prey in relation to habitat.
Limitations of the study
Firstly, given a lack of detailed benthic habitat data, we used a necessarily coarse habitat classification scheme (i.e. waters overlaying moraine banks, or not overlaying such banks). If the benthic habitats of the South Georgia shelf are mapped in more detail and our understanding of the mechanisms linking these to prey availability in superficial waters advances (Smith et al. Reference Smith, Mincks and DeMaster2006), less arbitrary, more biologically meaningful habitat distinctions should be possible (Aarts et al. Reference Aarts, MacKenzie, McConnell, Fedak and Matthiopoulos2008).
Secondly, we quantified habitat use within a relatively small area compared to the total area utilized by black-browed albatrosses foraging from South Georgia (cf. our Fig. 3 and fig. 1 in Phillips et al. Reference Phillips, Silk, Phalan, Catry and Croxall2004). Our models performed well for the shelf and slope region but poorly beyond this, where presence locations were sparse (Fig. 4). Previously we have shown that at the mega- to mesoscale, chick-rearing black-browed albatrosses prefer in ascending order oceanic, upper shelf slope, shelf break and neritic habitats (Wakefield et al. Reference Wakefield, Phillips, Trathan, Arata, Gales, Huin, Robertson, Waugh, Weimerskirch and Matthiopoulos2011). Birds from South Georgia travel as far away as the Antarctic Peninsula and the Patagonian shelf to access the latter types. In these areas the tendency to forage over moraine banks documented in the present study may be replaced by other sub-mesoscale habitat preferences. Therefore, data on seabed substrate throughout shelf waters accessible to this and other populations would be required to establish the generality of our findings. Similarly, the tendency for black-browed albatrosses from some populations to forage at the shelf break had the potential to confound our findings (Cherel et al. Reference Cherel, Weimerskirch and Trouve2000, Wakefield et al. Reference Wakefield, Phillips, Trathan, Arata, Gales, Huin, Robertson, Waugh, Weimerskirch and Matthiopoulos2011). This is because terminal moraines from the Last Glacial Maximum also tend to occur at shelf margins (Syvitski Reference Syvitski1991). However, the inclusion of depth, in addition to habitat type, in our models effectively captured variability in spatial usage due to proximity to the shelf break (Fig. 4). Furthermore, visual examination of the GPS, STAU and habitat data shows a remarkable congruence between the tracks of the birds and the distribution of the banks within the shelf break zone (Figs 3 & 6).
Finally, we analysed albatross habitat use in the vicinity of South Georgia. However, as noted above, many of the tracked birds flew rapidly through the study area, commuting to more distant foraging sites. Excluding these birds from our analysis would have allowed us to focus only on the behaviour of the birds that spent the majority of their time in the study area. However, it would have been inappropriate given that both black-browed albatrosses and their close relatives grey-headed albatrosses Thalassarche chrysostoma (Forster, 1785) sometimes forage (albeit to a limited extent) during the commuting stages of provisioning trips (Catry et al. Reference Catry, Phillips, Phalan, Silk and Croxall2004, Wakefield, unpublished data). Indeed, we noted that some of the longer-ranging birds in this study also visited the moraine banks. Hence, habitat selection may have been made even while the birds were primarily concerned with commuting.
Type of prey captured over the banks
The diet of the birds observed during the study was typical of that of chick-rearing black-browed albatrosses from South Georgia during years of high krill abundance, being dominated by krill and fish (Reid et al. Reference Reid, Croxall and Prince1996). Cephalopods, the other major prey taxon taken by this population, were extremely rare in the diet of the birds that foraged predominantly in local neritic waters. In a previous studies, fish caught by black-browed albatrosses had estimated masses ranging from 0.5–2012 g, with the vast majority weighing > 50 g (Reid et al. Reference Reid, Croxall and Prince1996, Cherel et al. Reference Cherel, Weimerskirch and Trouve2000), whereas the mass of individual krill fed to chicks at Bird Island typically ranges from 1–22 g (Croxall et al. Reference Croxall, Prince and Reid1997). Comparing these figures to the distribution of prey masses estimated using the STAU data (Fig. 7) it seems that the prey captured by birds #22 and #27 in waters overlaying moraine banks probably consisted of krill and fish in approximately equal proportions. A closer examination of the PDER events recorded both on and off the moraine banks shows that in some instances many small prey items were ingested in quick succession. We interpret this as krill or other small shoaling prey being caught rapidly, one by one.
Do albatrosses use the banks because of attraction to trawlers?
It is highly unlikely that black-browed albatrosses spend more time foraging over moraine banks than over surrounding habitats by chance, so some explanation for why this occurs is needed. One possibility is that birds are attracted to trawlers, which either target the banks or associated shoals of mackerel icefish. It is well known that black-browed albatrosses scavenge from trawlers (and as a consequence are frequently killed by trawl warps) (Sullivan et al. Reference Sullivan, Reid and Bugoni2006). However, such attraction to trawlers could at best only account for a small proportion of the birds’ association with moraine banks during our study because this occurred when fishing boats were both present and absent. For example, during our study bank 5 was visited by two tracked albatrosses but no fishing boats (Figs 2 & 3). Furthermore, recent research in the Falklands indicates a great deal of individual variability in associations between black-browed albatrosses and fishing vessels, with the majority of birds in a given population not targeting vessels even if they are easily within foraging range (Granadeiro et al. Reference Granadeiro, Phillips, Brickle and Catry2011).
However, we caution firstly, that our analysis relied on the accuracy of reported set and haul data, which in the past have contained errors (Kock Reference Kock2001) and secondly, that we had no information on the movements of trawlers while not engaged in fishing. As such, it is possible that birds were attracted to fishing boats not actively trawling but perhaps engaged in prospecting for fish on the banks. However, this is unlikely firstly, because black-browed albatrosses are primarily attracted to working (and especially actively discarding) trawlers (Sullivan et al. Reference Sullivan, Reid and Bugoni2006), and secondly because fishing boats were entirely absent from the study area for the majority of the study's duration (Fig. 5).
Do albatrosses use the banks because of upward benthic-pelagic coupling?
Most documented instances of benthic-pelagic coupling usually involve a downwards transfer of energy through the water column, such that production at higher trophic levels in benthic ecosystems is regulated by primary production in epipelagic waters (Graf Reference Graf1992). However, processes occurring at or near the seabed may also affect biomass in epipelagic waters, giving rise to upward benthic-pelagic coupling (Smith et al. Reference Smith, Mincks and DeMaster2006). As such, variability in prey abundance in surface waters may be mechanistically linked to the patchiness of benthic habitats, explaining the observed distribution of surface-feeding black-browed albatrosses. What might these links be? We discuss three, not necessarily mutually exclusive, hypotheses.
The first process by which the banks could give rise to higher prey availability in surface waters is through the upward export of enhanced benthic production. Barnes (Reference Barnes2008) described how substrates comprised of boulders, presumably themselves glacial moraines, on shelf and shelf slopes of the Shag Rocks, to the west of South Georgia, sustained a much richer fauna than surrounding habitats. On the boulders a cryptofauna of filter feeders that exploit the interstitial spaces characteristic of such large grained sediments predominates. In other shelf seas demersal fish are known to be more abundant in such topographically complex, mixed glacial sediment habitats (Todd & Kostylev Reference Todd and Kostylev2011). It is possible therefore, that black-browed albatrosses are attracted to the banks in order to predate demersal fish, which frequently appear in their diet (Reid et al. Reference Reid, Croxall and Prince1996, Cherel et al. Reference Cherel, Weimerskirch and Trouve2000). Although it is not clear how they would access such prey (Cherel et al. Reference Cherel, Weimerskirch and Trouve2000), some fish taken by albatrosses during foraging trips from South Georgia feed both benthically and near the surface (Collins et al. Reference Collins, Shreeve, Fielding and Thurston2008). However, this would offer a rather incomplete explanation because it is probable that a large proportion of the prey caught by black-browed albatrosses on the banks was krill rather than fish.
A second mechanism could potentially enhance the availability of krill (and also therefore planktivorous fish) in surface waters overlaying the banks. Although krill primarily feed pelagically, they have also been observed grazing photodetritus (the remains of phytoplankton) on the seabed at depths well in excess of that of the South Georgia shelf (Gutt & Siegel Reference Gutt and Siegel1994). Gutt (Reference Gutt2000) pointed out that krill swarms migrating to the surface, which they are able to do rapidly (Clarke & Tyler Reference Clarke and Tyler2008), can transfer energy from the seabed to surface-feeding predators. It is possible that as currents pass over the moraine banks, eddies cause photodetritus to accumulate in their lee or in the interstitial spaces between their component cobbles and boulders. This would provide a more profitable grazing habitat for krill swarms than the surrounding seafloor. Krill would themselves become available in surface waters due to habitual diel vertical migrations (Everson Reference Everson1983). As yet, little is known about the frequency of benthic feeding in krill so this is a very speculative hypothesis.
Finally, the banks may cause local enhancement of phytoplankton growth in the epipelagic zone. Currents on the South Georgia shelf have velocities in the order of 1–15 cm s-1 (Atkinson et al. Reference Atkinson, Whitehouse, Priddle, Cripps, Ward and Brandon2001). The turbulence these create as they pass over the banks, some of which rise to over 100 m above the seabed, could conceivably enhance vertical mixing, re-supplying limiting nutrients to the photic zone in a similar process to those occurring around oceanic seamounts (Lueck & Mudge Reference Lueck and Mudge1997). Although South Georgia shelf waters are not thought to be significantly nutrient limited (Atkinson et al. Reference Atkinson, Whitehouse, Priddle, Cripps, Ward and Brandon2001), this process could become important at the outer shelf, where the influence of nutrient poor oceanic waters is most apparent. Indeed, it would be facilitated by the weak degree of stratification of these waters (Sedwick et al. Reference Sedwick, DiTullio and Mackey2000). Similarly, the dynamics of the South Georgia shelf break front, a feature that also gives rise to localized nutrient upwelling, are tied to small-scale bathymetry (Brandon et al. Reference Brandon, Murphy, Whitehouse, Trathan, Murray, Bone and Priddle1999). Increased stocks of phytoplankton due to such nutrient re-supply would sustain higher levels of secondary production, principally of Antarctic krill, a major component in the diet of both black-browed albatrosses and mackerel icefish (Kock et al. Reference Kock, Wilhelms, Everson and Groger1994, Croxall et al. Reference Croxall, Prince and Reid1997). Furthermore, turbulence caused by the banks could result in the retention of phytoplankton and zooplankton in their vicinity (Barth et al. Reference Barth, Pierce and Castelao2005).
Wider implications
Links between the spatial usage of surface-feeding, pelagic seabirds and the distribution of benthic habitats are perhaps counter intuitive but deserve wider investigation. Yet, to our knowledge, this is the first time a study such as ours has been carried out. In part this is because tracking devices and remote sensing (now the predominant tools used to study pelagic seabird habitat use) were not hitherto accurate enough to examine sub-mesoscale habitat interactions (Wakefield et al. Reference Wakefield, Phillips and Matthiopoulos2009). However, GPS tracking now allows individual seabird movements to be recorded to an accuracy of c. ± 5 m, and equally accurate acoustic mapping of seabed habitats is now becoming available. In this context, it is notable that moraine banks are a common feature of continental shelves (Syvitski Reference Syvitski1991), with well-known examples including the Grand Banks of Canada and the Dogger Bank of the North Sea. Furthermore, the prey of many surface-feeding pelagic seabirds, especially those that routinely forage in neritic waters have well-known preferences for habitats characterized by particular seabed sediments (e.g. Frederiksen et al. Reference Frederiksen, Wright, Harris, Mavor, Heubeck and Wanless2005).
A useful way of contextualising the results of fine-scale studies of pelagic seabird habitat use such as ours is within the conceptual framework of hierarchical patches (Fauchald et al. Reference Fauchald, Erikstad and Skarsfjord2000). For example, as we have previously shown, at the meso- to macroscale chick-rearing black-browed albatrosses from South Georgia prefer to forage in neritic and shelf break waters (Wakefield et al. Reference Wakefield, Phillips, Trathan, Arata, Gales, Huin, Robertson, Waugh, Weimerskirch and Matthiopoulos2011). Our present findings imply that within these neritic habitat patches they also show preference at the sub-mesoscale, in this case for waters overlaying a particular benthic habitat. This supports the view that at large to medium spatial scales the distribution of predators reflects that of prey (Fauchald et al. Reference Fauchald, Erikstad and Skarsfjord2000). Indeed, black-browed albatrosses’ putative mesoscale preference for shelf break and upper slope habitats (Cherel et al. Reference Cherel, Weimerskirch and Trouve2002, Wakefield et al. Reference Wakefield, Phillips, Trathan, Arata, Gales, Huin, Robertson, Waugh, Weimerskirch and Matthiopoulos2011) may in fact arise due to a sub-mesoscale preference for moraine substrates, which predominantly occur at shelf edges (Syvitski Reference Syvitski1991). Hence, an understanding of such fine-scale habitat use may be informative not only about the ecology of seabirds but also about processes of benthic-pelagic coupling, which are clearly deserving of further study.
Acknowledgements
The authors gratefully acknowledge generous financial support provided for the fieldwork element of this study by the Antarctic Science Bursary and the Russell Trust, as well as logistical support provided in kind by the UK Antarctic Funding Initiative Collaborative Gearing Scheme. This study was partly carried out while the first author was in receipt of a UK Natural Environment Research Council CASE studentship (NER/S/A/2005/13648). We thank Peter Fretwell for providing a shapefile depicting the extent of moraine banks around South Georgia and Dr Jose Xavier for identifying prey samples. We thank the Government of South Georgia and South Sandwich Islands for permission to use fishery effort data. We are very grateful to Ewan Edwards, Derren Fox, Fabrice le Bouard, Donald Malone, Chris Martin, Felice Prospero-Porti, Robin Snape and John Withers for assisting with fieldwork at Bird Island and to Prof John Croxall for supporting long-term studies there. We thank two anonymous referees for helpful comments on an earlier draft of this manuscript.








