


Significant outcomes
∙ Localised effects of ischaemia in the prefrontal cortex (PFC) can be studied using endothelin-1 (ET-1) microinjections
∙ Bilateral ET-1 injections produce infarcts confined to the PFC
∙ ET-1 injected rats demonstrate several behaviours indicative of heightened anxiety independent of changes in locomotor activity
Limitations
∙ Enhanced anxiety following ischaemic lesions can only be inferred from rat behaviour
∙ Effects of the lesions on secondary brain regions such as the amygdala was not examined
∙ No evaluations of altered neurochemistry were performed
Introduction
Stroke is one of the main forms of death and disability, with an estimated 5.7 million deaths worldwide and 16 million new cases reported each year [Reference Strong, Mathers and Bonita1]. Among stroke survivors motor dysfunction is often the first recognisable sign of long-term disability, and preclinical stroke research has been very successful at modeling and replicating many of these signs [Reference Kleim, Boychuk and Adkins2]. Less well studied, however, are complex cognitive deficits and disturbances of mood including anxiety/depression post-stroke. Up to one third of stroke survivors suffer from such deficits, that often manifest later in the recovery period [Reference Tatemichi, Desmond, Stern, Paik, Sano and Bagiella3]. One reason for this lack of data specific to post-stroke deficits in mood and cognition is that the most common models of ischaemic stroke in rodents produce significant damage to multiple brain regions which may confound behavioural testing [Reference Liu and McCullough4].
ET-1, the most biologically prominent of the four discovered isoforms of endothelin, is a potent vasoconstrictor that can be used directly to occlude blood vessels in vivo [Reference Sharkey and Butcher5,Reference Luscher and Barton6]. Since its discovery and subsequent characterisation, this compound has been used in a host of vascular impairment models in order to study the effects of stroke on behaviour in vivo [Reference Thiyagarajan and Sharma7,Reference Livingston-Thomas, McGuire, Doucette and Tasker8].
In rats, the PFC is a brain region strongly implicated in higher order (executive) cognition, anxiety, and depression [Reference Birrell and Brown9–Reference Bissiere, McAllister, Olpe and Cryan11] although the immediate and delayed consequences of ischaemia in the PFC are largely unexplored.
Aim of the study
To determine if bilateral microinjections of ET-1 into the medial prefrontal cortex (mPFC) of rats would result in significant deficits in two behavioural tasks linked to cognition or anxiety.
Material and methods
Animals
All procedures were conducted in accordance with the guidelines of the Canadian Council on Animal Care and were approved in advance by the University of Prince Edward Island Animal Care Committee. Adult male Sprague-Dawley rats (n=21, 126–150 g on arrival) were purchased from Charles River Laboratories (Montreal, Canada) and singly housed on a 12 h light/dark cycle (lights on at 06:00 and off at 18:00) with food (Purina rat chow) and water available ad libitum until training and testing. Animals were housed singly post-surgery to avoid aggression between conspecifics. Upon arrival, the animals were handled by the experimenter for 5 min each day for three days during an acclimation period. All surgeries, behavioural training, and behavioural testing occurred during the light phase of the light/dark cycle.
Surgical methods
Rats were randomly divided into groups receiving ET-1 (n=11) or vehicle [artificial cerebrospinal fluid (aCSF)] (n=10). Briefly, each rat was anaesthetised using isoflurane induction (4%) and maintenance (2–3%) and mounted on a stereotaxic frame. Topical Xylocaine (AstraZeneca, Mississauga, Canada) was applied to the shaved head area and a 2 cm incision was made 5 min later. Two small burr holes were then drilled into the cranium and a 26 gauge needle and a 10 µl syringe was lowered into each injection site (coordinates A/P +3.0; M/L ±0.7; D/V −4.5, all coordinates relative to bregma as per Paxinos and Watson [Reference Paxinos and Watson12]) and was left for 1 min before ET-1 injection. After one minute, 1 µl of ET-1 (400 pmol) dissolved in aCSF was injected at a rate of 0.5 µl/min. Once injected, the needle was left undisturbed for 4 min to allow for drug dispersal at the injection site, and then the needle was slowly retracted from the brain. The same procedure was repeated for the second injection point. After the ET-1 injections, the incision site was sutured, Xylocaine was reapplied and each animal was given a 2.0 mg/kg subcutaneous injection of butorphanol tartrate and monitored for signs of pain and distress. During surgeries, one animal from each group died, and subsequent to the surgeries on post-surgery day (PSD) 4 one animal from the ET-1 group had to be euthanised for humane reasons, leaving n=9 for each experimental group.
Behavioural testing
Rats were tested in two paradigms; a test of temporal object recognition (TOR) (executive function) and in an elevated plus maze (EPM) (anxiety). The testing procedure for the temporal object recognition task was modified from Hannesson et al. [Reference Hannesson, Howland and Phillips13] and consisted of placing the rat in a round, empty, fibreglass container (148.6 cm in diameter; walls 63.5 cm in height) to which they had been acclimated on PSD5 and 6. Testing began on PSD7 and consisted of placing the animal in the maze with a pair of identical objects, crafted from Lego objects of different shapes, sizes and colours, positioned equidistant from the centre of the maze and the maze walls. The rat was allowed to explore for 4 min before being returned to their home cage. After a delay of 60 min, the animal was placed back in the maze with a new pair of identical objects for a further 4 min. After an additional delay of 180 min, the animal was returned to the maze with one object from each of the two previous trials for 3 min and the time spent exploring each of the objects was recorded. Exploration of an object was defined as the animal’s head being within at least 2 cm of the object’s surface, and scoring was done with AnyMaze software (Stoelting, Wood Dale, IL, USA). TOR testing took place on PSD7, PSD14, PSD21, and PSD28, with different sets of objects being used at randomised locations for each day of testing.
Testing in the EPM consisted of placing the animal in the central region of a platform (elevated 83 cm from the floor) connected to four arms (112 cm long×10 cm wide). Two of the arms were exposed (open arms), and the other two arms had 28 cm high metal walls (closed arms). Rats were given 5 min to explore the maze and behaviour was video recorded using a webcam (Logitech Deluxe S7500, Lausanne, Switzerland) suspended over the centre of the maze, connected to a laptop, using ANY-Maze Software version 4.76 (Stoelting Co., Wood Dale, IL, USA). The variables recorded were the overall percentage of time spent in the closed arms, the amount of time spent immobile, the amount of time exhibiting ‘freezing’ behaviour (defined as the amount of time that no discernable movement was detected from the animal after 250 ms of non-movement), the number of freezing episodes, the amount of distance moved throughout the trial, and the number of rearing motions made during testing. Animals were tested three times over the course of the experiment; pre-surgery and on post-surgery (PSD) days 8 and 29.
Histology
On PSD29 animals were anaesthetised with isoflurane and euthanised by decapitation. Brains were removed and immersion fixed in 10% formalin. Brain tissues were sectioned, mounted, stained with cresyl violet and infarcts quantified by taking each digital image and tracing the area of tissue which displayed a lack of, or abnormal, cresyl violet staining or altered cytological architecture. Tracing was performed using ImageJ software, version 1.45s (ImageJ; National Institutes of Health, Bethesda, MD USA). Once each area had been traced, the volume of infarct damage was quantified by summing each individually calculated area and multiplying the area by the distance between each section (100 µm).
Data analysis
Statistical analyses were performed using Graph Pad Prism, Version 5.00 (Graph Pad, La Jolla, CA, USA) and employed a two-way repeated measures analysis of variance (ANOVA) comparing the day of testing with the surgery group. Post-hoc comparisons were made using Bonferroni post-test, one-way ANOVAs, or t-tests where appropriate. In all analyses statistical significance was considered at p⩽0.05.
Results
Histology
Figure 1a and b demonstrate the extent of damage produced following bilateral ET-1 injections into the mPFC as examined 29 days post stroke surgery. Figure 1a provides an overview of the estimated volume of damage every 200 µm, demonstrating the largest amount of damage occurred between +3.3 and +2.7 mm anterior to bregma, coinciding with the bilateral injections placed at +3.0 mm anterior to bregma. Figure 1b gives an overview of the damage, with five coronal sections through the infarcted area.
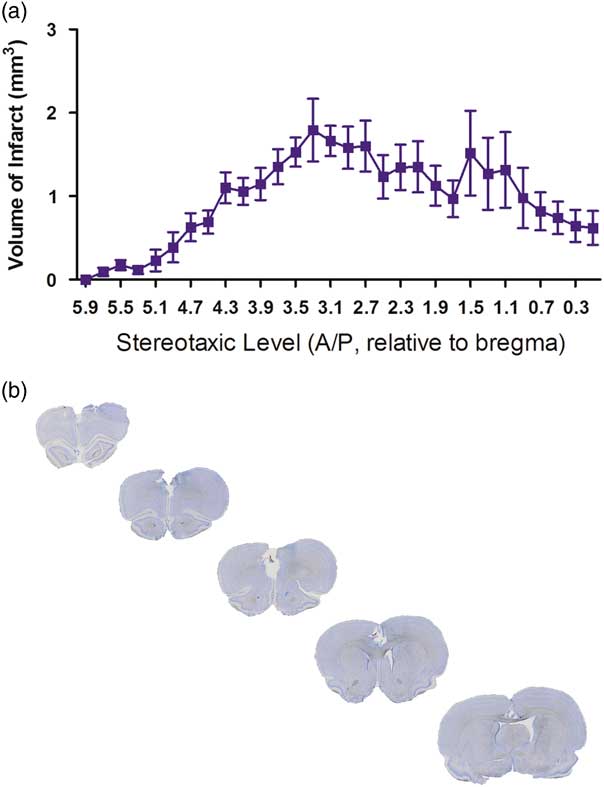
Fig. 1 Histology of the lesion. (a) Line graph indicating the relative volume of damage at different stereotaxic levels in the endothelin-1 (ET-1) group (n=9). (b) A representative example of the ischaemic insult within the medial prefrontal cortex displayed through five successive coronal sections stained with cresyl violet.
Behavioural testing
Testing in the TOR paradigm revealed no significant differences in object preference except on PSD14 when lesioned animals spent significantly more time with the first presented object than the second (38.8±12.4 s versus 7.6±3.7 s; t (16)=2.411, p<0.05) and lesioned animals spent significantly less time with the second object (sham 18.5±2.6 s; stroke 7.6±3.7 s; t (16)=2.351, p<0.05). At no other time point did either experimental group, sham or stroke, spend more time with the first object presented than the second object. Similarly there were no differences in total mobility between groups with the exception of PSD14 where lesioned animals moved less than sham animals in the maze (sham 14.2±1.6 m; stroke 5.3±1.4 m; t (16)=4.186, p<0.05).
In contrast, a number of significant differences were found between groups within the EPM. A two-way repeated measures ANOVA comparing the effects of the surgical procedure and the day of testing found a significant effect due to the testing day in the amount of time spent immobile in the maze (F(2,16)=7.856, p<0.05). Upon post-hoc analysis this effect was only seen in stroke animals comparing between pre-surgery and PSD8 (t (8)=3.561, p<0.05), as well as between PSD29 and PSD8 (t (8)=2.957, p<0.05) (Fig. 2a). No statistically significant effects were seen between days in the sham group. There was also a difference in freezing behaviour in the stroke animals; a two-way repeated measures ANOVA comparing the effects of the surgical procedure and the day of testing found a significant effect due to the testing day for the number of freezing events (F(2,16)=6.336, p<0.05). Upon post-hoc analysis this effect was only significant in the stroke animals comparing pre-surgery (17.67±2.96) and PSD8 (31.89±5.37) (t (8)=3.060, p<0.05), wherein animals displayed more freezing behaviours on PSD8 than pre-surgery. With regard to rearing behaviour while in the maze, two-way repeated measures ANOVA revealed statistically significant differences between sham and stroke group behaviour (F(1,16)=7.545, p<0.05) (Fig. 2b). Further Bonferroni post-hoc analysis revealed that these differences were only significant between groups on PSD8 (t (8)=3.023, p<0.05).
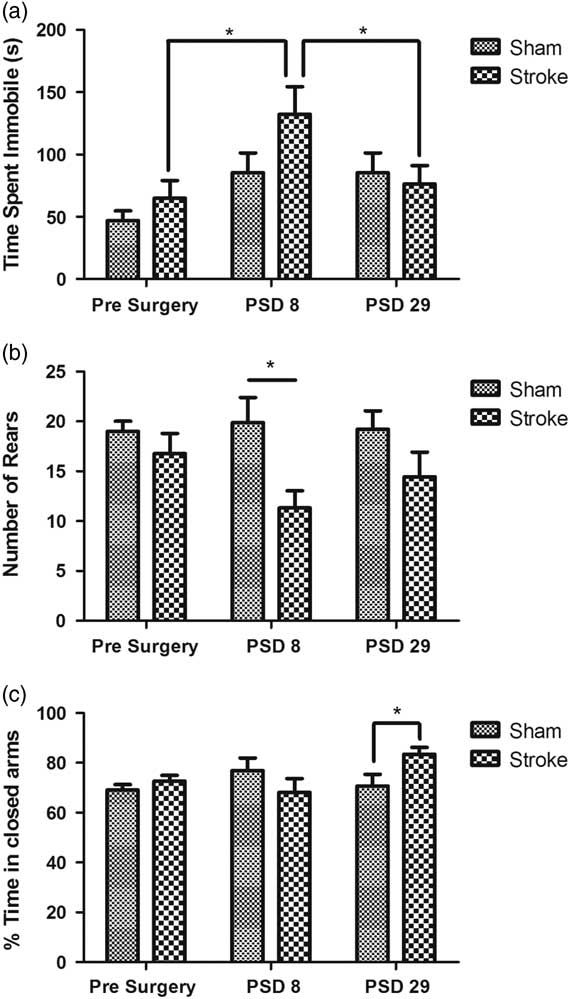
Fig. 2 Behaviour in the elevated plus maze (EPM). (a) Time spent immobile in the EPM pre- and post-surgery. (b) Number of rearing actions performed in the EPM. (c) Per cent of time spent in the closed arms of the EPM pre- and post-surgery. Bars indicate mean±SEM of time spent in closed arms, with n=9 for each group. *p<0.05 as indicated.
With respect to the relative per cent of time spent in the closed and open arms of the EPM, generally regarded as a measure of anxiety, a two-way ANOVA revealed an interactive effect between the day tested and the surgery group (F(2,16)=3.775, p<0.05), so subsequent analyses looked at each of these factors separately. No significant differences were detected between testing days but an unpaired t-test revealed a significant difference between surgical groups on PSD29, with animals in the stroke group spending more time in the closed arms of the maze than the sham group (t (16)=2.340, p<0.05). There was no significant difference between sham or stroke groups pre-surgery or on PSD8 (Fig. 2c).
Discussion
Much of the research into cognitive dysfunction post-stroke has examined deficits in working memory, spatial memory, and learning ability [Reference Jiwa, Garrard and Hainsworth14]. This may be because one of the most prominent areas of the brain that is damaged following global hypoperfusion and middle cerebral artery occlusion is the hippocampus, an area intricately involved with spatial and working memory [Reference Butler, Kassed, Sanberg, Willing and Pennypacker15,Reference Hartman, Lee, Zipfel and Wozniak16]. The PFC, however, is thought to regulate many higher-order cognitive functions, as well as to regulate behaviours associated with changes in emotions [Reference Uylings, Groenewegen and Kolb17,Reference Salzman and Fusi18]. For example, damage to the PFC has resulted in changes in aggressive behaviours in rats as well as humans [Reference Kolb and Nonneman19,Reference Giancola20]. In the current study animals with lesions to the medial PFC responded differently to anxiogenic situations, in that animals with mPFC lesions were more likely to spend time in the closed arms of the maze on PSD29 as well as exhibit anxiety behaviours on PSD8 as evidenced by a lower number of rearing actions, more time spent immobile and more freezing episodes throughout the EPM trials. Our contention that reduced mobility and rearing is due to ‘anxious’ behaviour rather than a non-specific reduction in locomotion is supported by two of our observations, namely (1) that we saw no significant changes in locomotor activity in the TOR arena on either PSD7 or 28 (data not shown), and (2) that these activity effects in the EPM were seen on PSD8 but not on PSD29 when the closed versus open arm data were collected. This is consistent with existing literature reporting that animals with chemically induced lesions to the PFC have either increased levels of spontaneous movement or no changes in movement overall [Reference Livingston-Thomas, Jeffers, Nguemeni, Shoichet, Morshead and Corbett21,Reference Shah and Treit22]. Thus the avoidance of the open arm on PSD29 appears to reflect a true anxiogenic response to the maze that develops over time and manifests subsequent to an early ‘anxious’ response.
Although not specifically examined in the current study, the behavioural changes seen in the EPM could also be due to secondary damage in brain regions other than the PFC. Lesions to the PFC have been shown to alter reciprocal connections with the amygdala [Reference Motzkin, Philippi, Wolf, Baskaya and Koenigs23], a brain region integrally involved in anxiogenic response. The involvement of the amygdala in the observed behavioural changes is a distinct possibility and future studies examining secondary damage to this structure are certainly warranted. Similarly our data suggest that increased anxiety develops slowly over time post-stroke. Examination of EPM behaviour at later time points is probably warranted. Alternately, these differences in EPM behaviour could be due to repeated tests in the maze, as this test is subject to ‘one-trial tolerance’ if done repeatedly [Reference File24]. Although there was a minimum of 10 days between testing pre-surgery and PSD8, the location of the testing was not changed, therefore one-trial tolerance could have affected the results.
The inability to detect consistent significant effects in the temporal object recognition test in the current study is surprising because prior literature using other methods of PFC lesioning has reported an effect [Reference Barker, Bird, Alexander and Warburton25]. Possible explanations for the difference include the size of the testing arena, the time delay between presentation of objects, and the objects themselves. More specifically, the objects used in the object recognition task, Lego® objects of varying colours and configurations, may not have been dissimilar enough from each other to be discerned by the test subjects. As well, the large size of the maze itself could have provided less incentive for the rodents to explore the maze, and not enough time with the objects themselves. Further experiments are required to determine which, if any, of these variables are responsible for the lack of effect in this task, and to further refine our methodology to demonstrate consistent recency discrimination performance.
The literature regarding prefrontal cortical manipulations on anxiety responses is mixed. One study demonstrated that animals with depleted levels of dopamine in the PFC displayed anxiogenic-like behaviours, spending more time in the closed arms of the EPM [Reference Espejo10]. However, other lesion experiments have produced evidence to the contrary [Reference Livingston-Thomas, Jeffers, Nguemeni, Shoichet, Morshead and Corbett21,Reference Shah and Treit22] when using different testing protocols and experimental designs. The effect(s) of these differences on anxiety behaviour in this model require(s) further investigation. The results of the current study support the conclusion that mPFC lesions are anxiogenic, and further, that this effect is relevant to the changes in emotionality that often follow ischaemic stroke.
Authors’ Contribution:
R.A.D. performed the experiment and analysed the data. R.A.D. and R.A.T. jointly designed the experiment and co-wrote the manuscript.
Financial Support
This work was supported by internal research funds provided by the University of PEI. RAD was the recipient of a Canada Graduate Scholarship provided by the Natural Sciences and Engineering Research Council of Canada.
Conflicts of Interest
The authors have no conflicts of interest to declare.
Ethical Standards
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional guides on the care and use of laboratory animals. The studies described were conducted in compliance with the ARRIVE guidelines.


